Recombinant Production, NMR Solution Structure, and Membrane Interaction of the Phα1β Toxin, a TRPA1 Modulator from the Brazilian Armed Spider Phoneutria nigriventer

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Expression and Purification
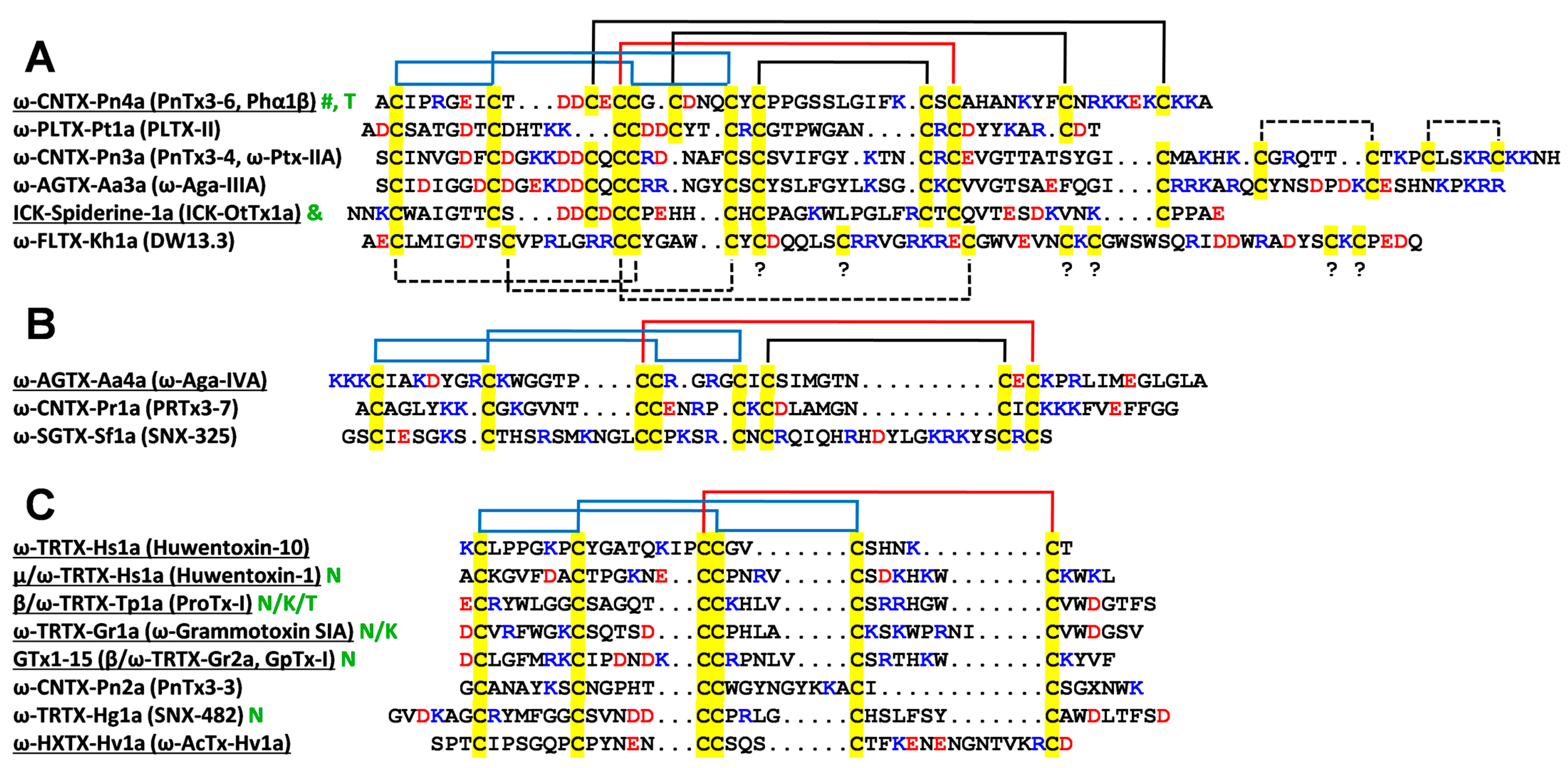
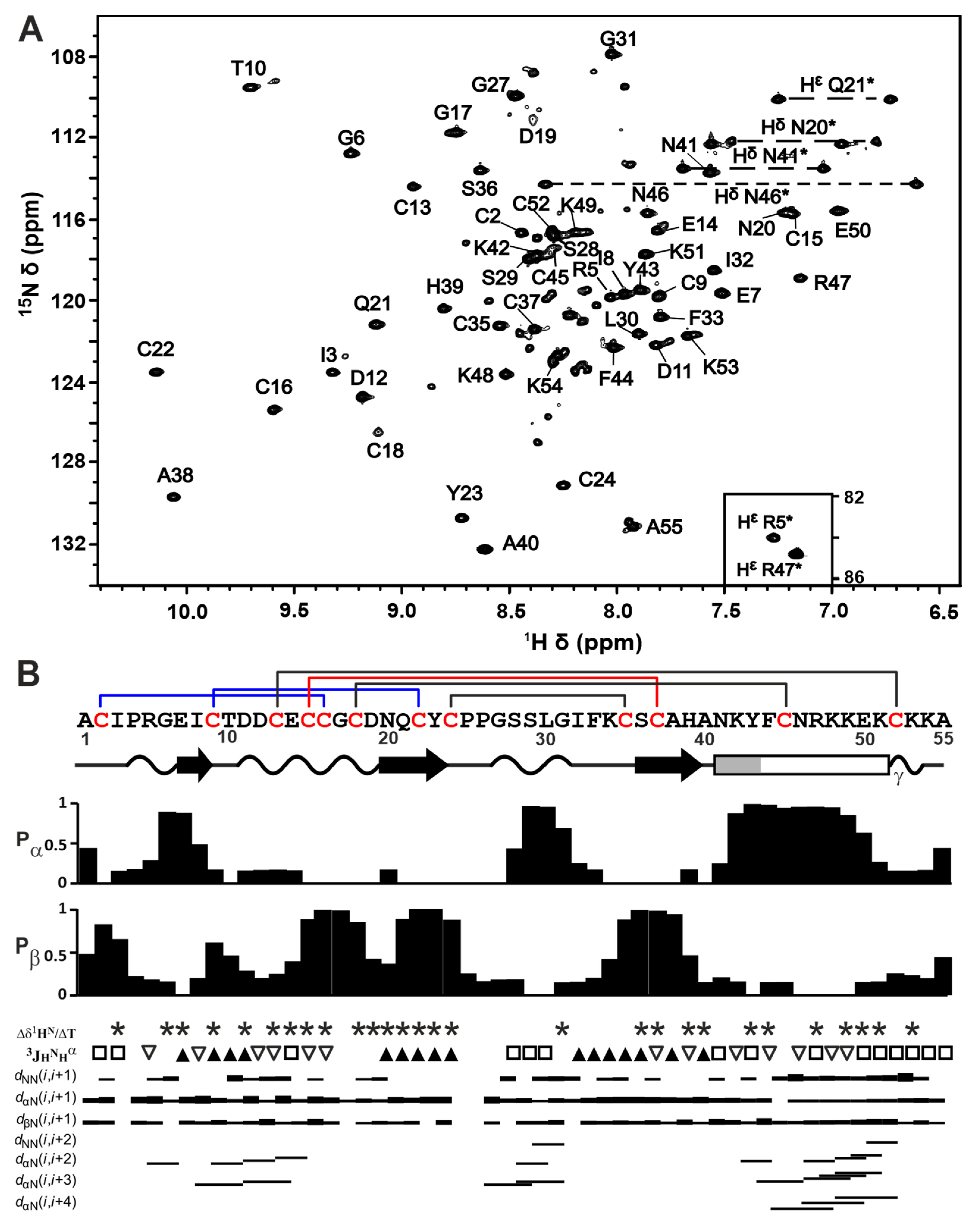
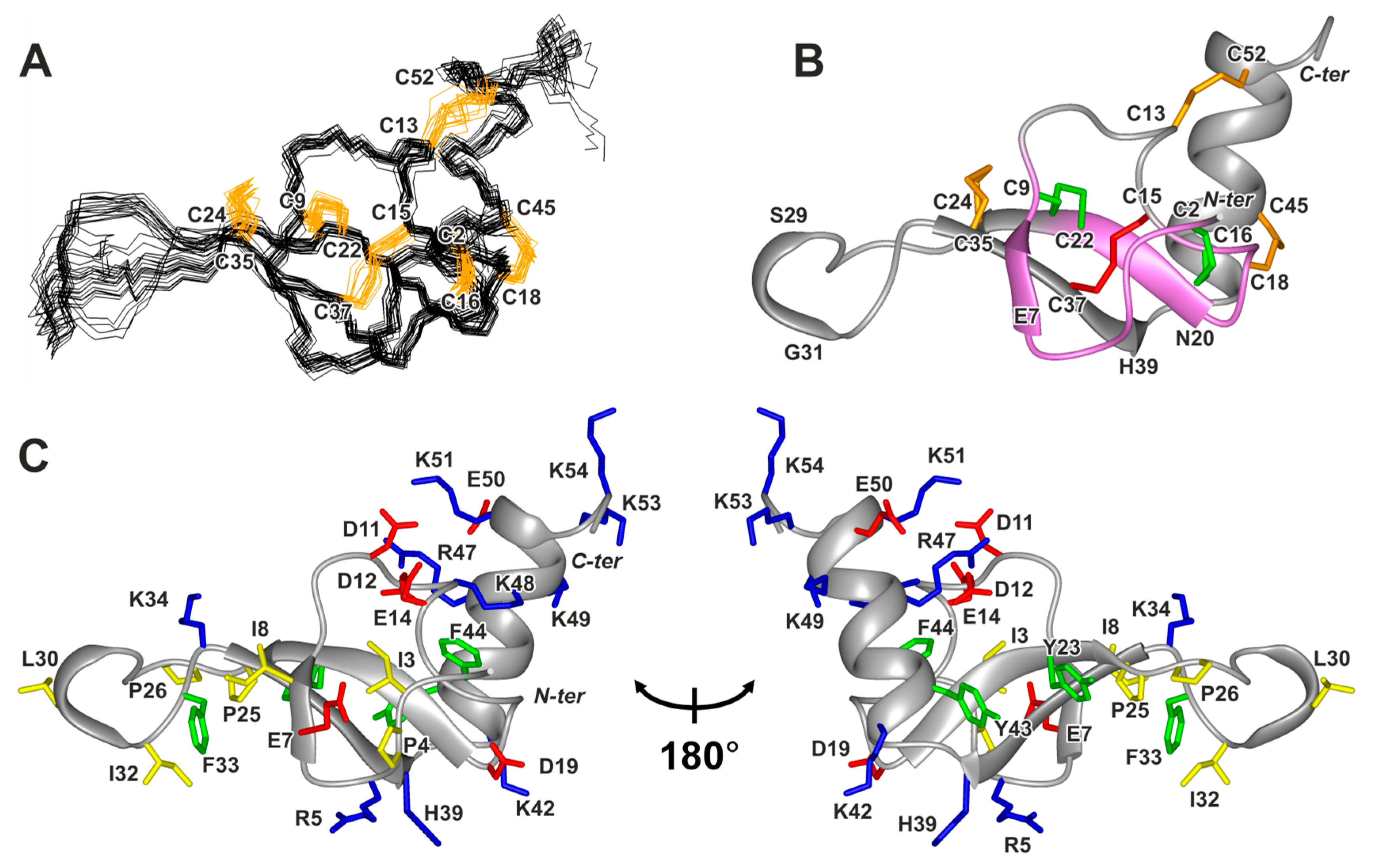
2.2. Spatial Structure of Phα1β

2.3. Backbone Dynamics of Phα1β
2.4. Properties of the Phα1β Surface
2.5. Interaction of Phα1β with Lipid Bilayers
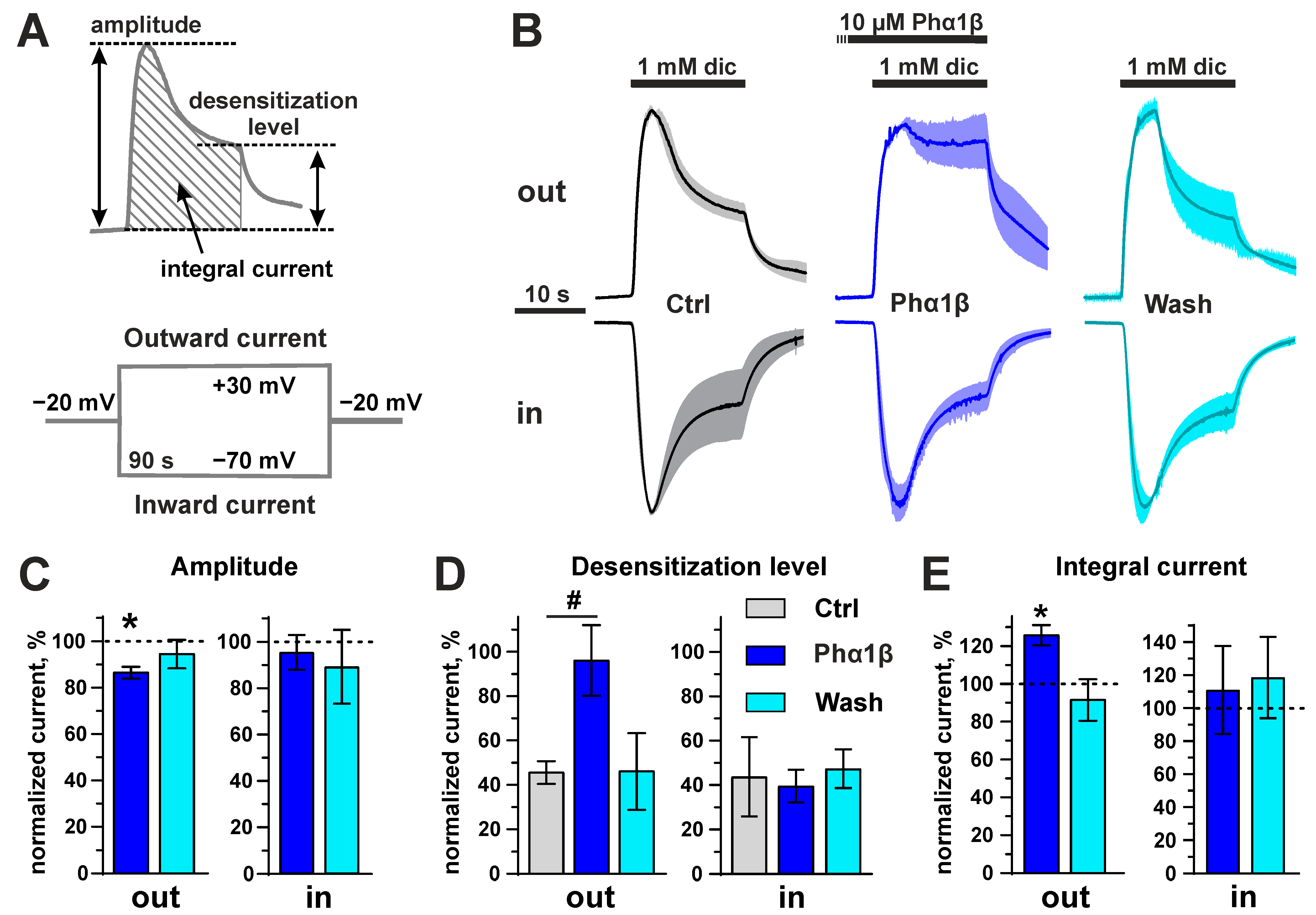
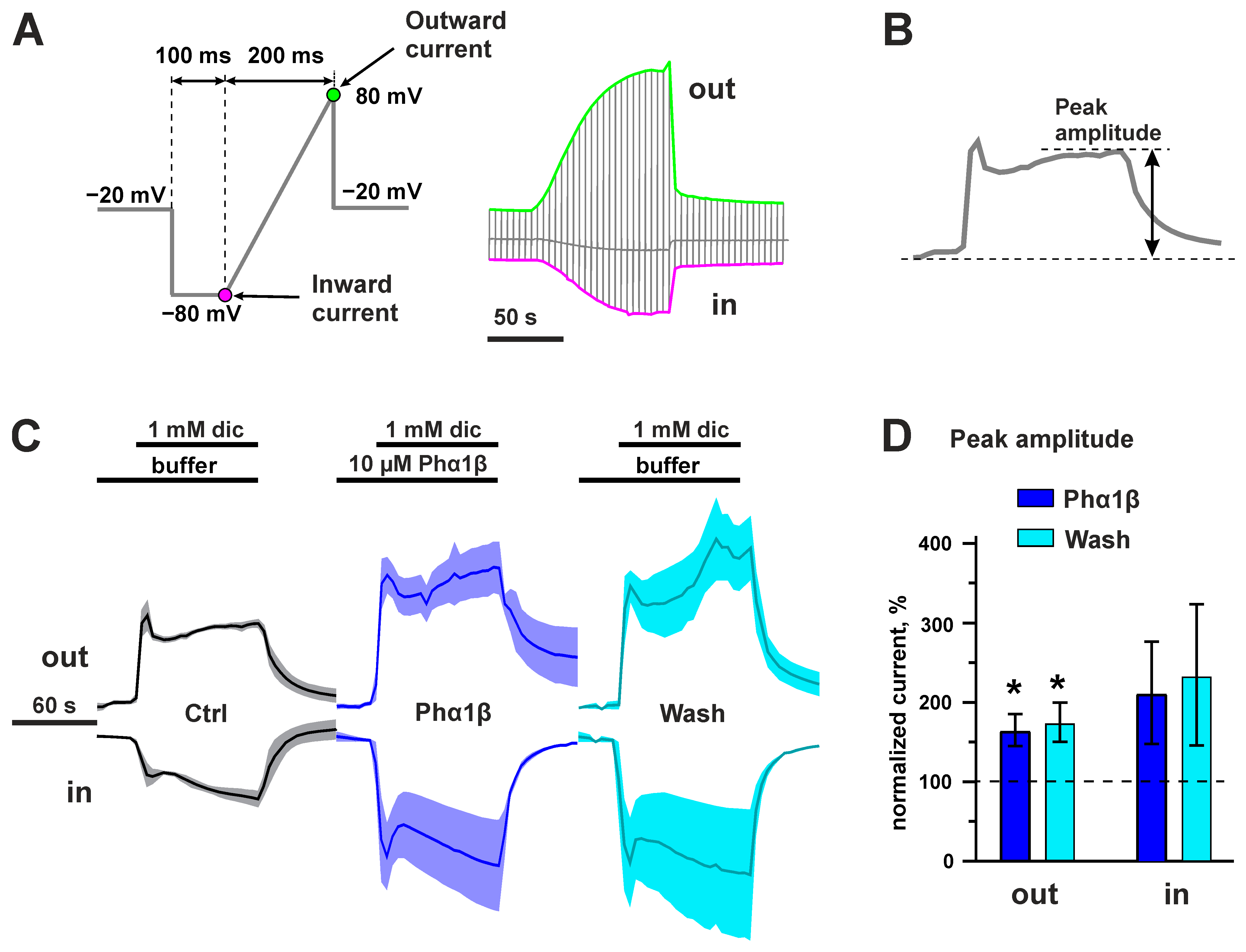
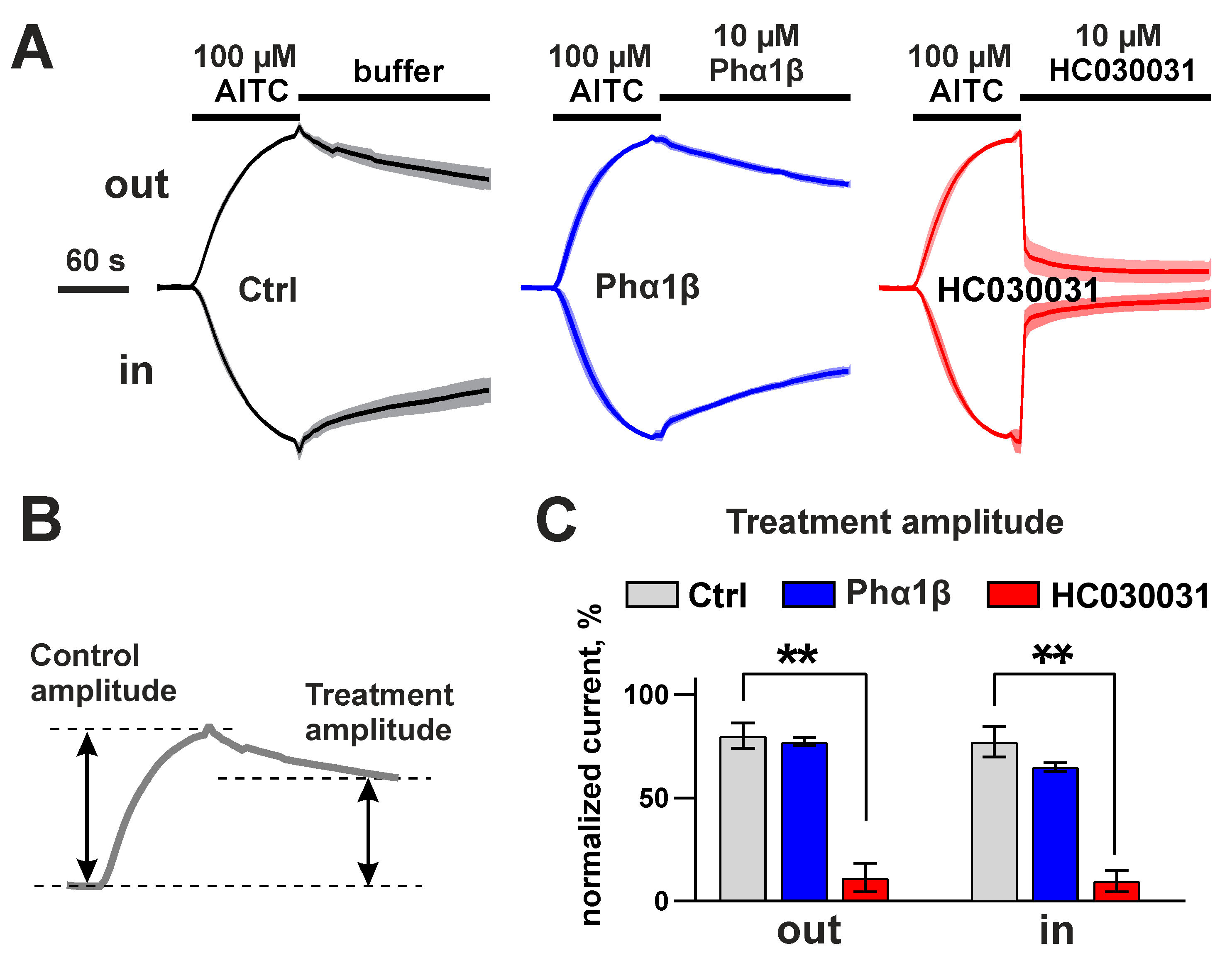
2.6. Interaction of Phα1β with Rat TRPA1 Expressed in Xenopus Oocytes
3. Conclusions
4. Materials and Methods
4.1. Bacterial Expression of Phα1β
4.2. Liquid Chromatography and Mass Spectrometry
4.3. NMR Experiments and Spatial Structure Calculation
4.4. Liposome Titration
4.5. Electrophysiological Recordings
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mouhat, S.; Jouirou, B.; Mosbah, A.; De Waard, M.; Sabatier, J.-M. Diversity of Folds in Animal Toxins Acting on Ion Channels. Biochem. J. 2004, 378, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Herzig, V.; Cristofori-Armstrong, B.; Israel, M.R.; Nixon, S.A.; Vetter, I.; King, G.F. Animal Toxins—Nature’s Evolutionary-Refined Toolkit for Basic Research and Drug Discovery. Biochem. Pharmacol. 2020, 181, 114096. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Beeton, C.; Galea, C.A.; Smith, B.J.; Chi, V.; Monaghan, K.P.; Garcia, A.; Rangaraju, S.; Giuffrida, A.; Plank, D.; et al. Engineering a Stable and Selective Peptide Blocker of the Kv1.3 Channel in T Lymphocytes. Mol. Pharmacol. 2009, 75, 762–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morsy, M.A.; Gupta, S.; Dora, C.P.; Jhawat, V.; Dhanawat, M.; Mehta, D.; Gupta, K.; Nair, A.B.; El-Daly, M. Venoms Classification and Therapeutic Uses: A Narrative Review. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 1633–1653. [Google Scholar] [CrossRef] [PubMed]
- Lauria, P.S.S.; Villarreal, C.F.; Casais-e-Silva, L.L. Pain Modulatory Properties of Phoneutria Nigriventer Crude Venom and Derived Peptides: A Double-Edged Sword. Toxicon 2020, 185, 120–128. [Google Scholar] [CrossRef]
- Maatuf, Y.; Geron, M.; Priel, A. The Role of Toxins in the Pursuit for Novel Analgesics. Toxins 2019, 11(2), 131. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro, M.d.N.; de Figueiredo, S.G.; Valentim, A.d.C.; Diniz, C.R.; von Eickstedt, V.R.; Gilroy, J.; Richardson, M. Purification and Amino Acid Sequences of Six Tx3 Type Neurotoxins from the Venom of the Brazilian “armed” Spider Phoneutria Nigriventer (Keys). Toxicon 1993, 31, 35–42. [Google Scholar] [CrossRef]
- Gomez, M.V.; Kalapothakis, E.; Guatimosim, C.; Prado, M.A.M. Phoneutria Nigriventer Venom: A Cocktail of Toxins That Affect Ion Channels. Cell Mol. Neurobiol. 2002, 22, 579–588. [Google Scholar] [CrossRef]
- Souza, A.H.; Ferreira, J.; Cordeiro, M.d.N.; Vieira, L.B.; De Castro, C.J.; Trevisan, G.; Reis, H.; Souza, I.A.; Richardson, M.; Prado, M.A.M.; et al. Analgesic Effect in Rodents of Native and Recombinant Ph Alpha 1beta Toxin, a High-Voltage-Activated Calcium Channel Blocker Isolated from Armed Spider Venom. Pain 2008, 140, 115–126. [Google Scholar] [CrossRef]
- de Souza, A.H.; Lima, M.C.; Drewes, C.C.; da Silva, J.F.; Torres, K.C.L.; Pereira, E.M.R.; de Castro Junior, C.J.; Vieira, L.B.; Cordeiro, M.N.; Richardson, M.; et al. Antiallodynic Effect and Side Effects of Phα1β, a Neurotoxin from the Spider Phoneutria Nigriventer: Comparison with ω-Conotoxin MVIIA and Morphine. Toxicon 2011, 58, 626–633. [Google Scholar] [CrossRef]
- Rigo, F.K.; Trevisan, G.; Rosa, F.; Dalmolin, G.D.; Otuki, M.F.; Cueto, A.P.; de Castro Junior, C.J.; Romano-Silva, M.A.; Cordeiro, M.d.N.; Richardson, M.; et al. Spider Peptide Phα1β Induces Analgesic Effect in a Model of Cancer Pain. Cancer Sci. 2013, 104, 1226–1230. [Google Scholar] [CrossRef]
- do Silva, J.F.; Binda, N.S.; Pereira, E.M.R.; de Lavor, M.S.L.; Vieira, L.B.; de Souza, A.H.; Rigo, F.K.; Ferrer, H.T.; de Castro Júnior, C.J.; Ferreira, J.; et al. Analgesic Effects of Phα1β Toxin: A Review of Mechanisms of Action Involving Pain Pathways. J. Venom. Anim. Toxins incl. Trop. Dis. 2021, 27, e20210001. [Google Scholar] [CrossRef]
- Vieira, L.B.; Kushmerick, C.; Reis, H.J.; Diniz, C.R.; Cordeiro, M.N.; Prado, M.A.M.; Kalapothakis, E.; Romano-Silva, M.A.; Gomez, M.V. PnTx3-6 a Spider Neurotoxin Inhibits K+-Evoked Increase in [Ca2+]i and Ca2+-Dependent Glutamate Release in Synaptosomes. Neurochem. Int. 2003, 42, 277–282. [Google Scholar] [CrossRef]
- Vieira, L.B.; Kushmerick, C.; Hildebrand, M.E.; Garcia, E.; Stea, A.; Cordeiro, M.N.; Richardson, M.; Gomez, M.V.; Snutch, T.P. Inhibition of High Voltage-Activated Calcium Channels by Spider Toxin PnTx3-6. J. Pharmacol. Exp. Ther. 2005, 314, 1370–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonello, R.; Fusi, C.; Materazzi, S.; Marone, I.M.; De Logu, F.; Benemei, S.; Gonçalves, M.C.; Coppi, E.; Castro-Junior, C.J.; Gomez, M.V.; et al. The Peptide Phα1β, from Spider Venom, Acts as a TRPA1 Channel Antagonist with Antinociceptive Effects in Mice. Br. J. Pharmacol. 2017, 174, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Julius, D. TRP Channels and Pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef] [Green Version]
- Logashina, Y.A.; Korolkova, Y.V.; Kozlov, S.A.; Andreev, Y.A. TRPA1 Channel as a Regulator of Neurogenic Inflammation and Pain: Structure, Function, Role in Pathophysiology, and Therapeutic Potential of Ligands. Biochem. Mosc. 2019, 84, 101–118. [Google Scholar] [CrossRef]
- Manolache, A.; Babes, A.; Madalina Babes, R. Mini-Review: The Nociceptive Sensory Functions of the Polymodal Receptor Transient Receptor Potential Ankyrin Type 1 (TRPA1). Neurosci. Lett. 2021, 764, 136286. [Google Scholar] [CrossRef]
- Koivisto, A.-P.; Belvisi, M.G.; Gaudet, R.; Szallasi, A. Advances in TRP Channel Drug Discovery: From Target Validation to Clinical Studies. Nat. Rev. Drug. Discov. 2022, 21, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, S.; Nikolaeva-Koleva, M.; Alarcón-Alarcón, D.; Butrón, L.; González-Rodríguez, S. Is TRPA1 Burning Down TRPV1 as Druggable Target for the Treatment of Chronic Pain? Int. J. Mol. Sci. 2019, 20(12), 2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza Monteiro de Araujo, D.; Nassini, R.; Geppetti, P.; De Logu, F. TRPA1 as a Therapeutic Target for Nociceptive Pain. Expert Opin. Ther. Targets 2020, 24, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Rigo, F.K.; Dalmolin, G.D.; Trevisan, G.; Tonello, R.; Silva, M.A.; Rossato, M.F.; Klafke, J.Z.; Cordeiro, M.d.N.; Castro Junior, C.J.; Montijo, D.; et al. Effect of ω-Conotoxin MVIIA and Phα1β on Paclitaxel-Induced Acute and Chronic Pain. Pharmacol. Biochem. Behav. 2013, 114–115, 16–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, J.; Liu, B.; Cao, G.; Lipchik, A.M.; Perez, M.; Dekan, Z.; Mobli, M.; Daly, N.L.; Alewood, P.F.; Parker, L.L.; et al. A Tarantula-Venom Peptide Antagonizes the TRPA1 Nociceptor Ion Channel by Binding to the S1-S4 Gating Domain. Curr. Biol. 2014, 24, 473–483. [Google Scholar] [CrossRef] [Green Version]
- Agwa, A.J.; Henriques, S.T.; Schroeder, C.I. Gating Modifier Toxin Interactions with Ion Channels and Lipid Bilayers: Is the Trimolecular Complex Real? Neuropharmacology 2017, 127, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Männikkö, R.; Shenkarev, Z.O.; Thor, M.G.; Berkut, A.A.; Myshkin, M.Y.; Paramonov, A.S.; Kulbatskii, D.S.; Kuzmin, D.A.; Sampedro Castañeda, M.; King, L.; et al. Spider Toxin Inhibits Gating Pore Currents Underlying Periodic Paralysis. Proc. Natl. Acad. Sci. USA 2018, 115, 4495–4500. [Google Scholar] [CrossRef] [Green Version]
- Milescu, M.; Vobecky, J.; Roh, S.H.; Kim, S.H.; Jung, H.J.; Kim, J.I.; Swartz, K.J. Tarantula Toxins Interact with Voltage Sensors within Lipid Membranes. J. Gen. Physiol. 2007, 130, 497–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milescu, M.; Bosmans, F.; Lee, S.; Alabi, A.A.; Kim, J.I.; Swartz, K.J. Interactions between Lipids and Voltage Sensor Paddles Detected with Tarantula Toxins. Nat. Struct. Mol. Biol. 2009, 16, 1080–1085. [Google Scholar] [CrossRef]
- Berkut, A.A.; Peigneur, S.; Myshkin, M.Y.; Paramonov, A.S.; Lyukmanova, E.N.; Arseniev, A.S.; Grishin, E.V.; Tytgat, J.; Shenkarev, Z.O.; Vassilevski, A.A. Structure of Membrane-Active Toxin from Crab Spider Heriaeus Melloteei Suggests Parallel Evolution of Sodium Channel Gating Modifiers in Araneomorphae and Mygalomorphae. J. Biol. Chem. 2015, 290, 492–504. [Google Scholar] [CrossRef] [Green Version]
- Shenkarev, Z.O.; Lyukmanova, E.N.; Paramonov, A.S.; Panteleev, P.V.; Balandin, S.V.; Shulepko, M.A.; Mineev, K.S.; Ovchinnikova, T.V.; Kirpichnikov, M.P.; Arseniev, A.S. Lipid–Protein Nanodiscs Offer New Perspectives for Structural and Functional Studies of Water-Soluble Membrane-Active Peptides. Acta Nat. 2014, 6, 84. [Google Scholar] [CrossRef]
- Redaelli, E.; Cassulini, R.R.; Silva, D.F.; Clement, H.; Schiavon, E.; Zamudio, F.Z.; Odell, G.; Arcangeli, A.; Clare, J.J.; Alagón, A.; et al. Target Promiscuity and Heterogeneous Effects of Tarantula Venom Peptides Affecting Na+ and K+ Ion Channels. J. Biol. Chem. 2010, 285, 4130–4142. [Google Scholar] [CrossRef] [Green Version]
- Middleton, R.E.; Warren, V.A.; Kraus, R.L.; Hwang, J.C.; Liu, C.J.; Dai, G.; Brochu, R.M.; Kohler, M.G.; Gao, Y.-D.; Garsky, V.M.; et al. Two Tarantula Peptides Inhibit Activation of Multiple Sodium Channels. Biochemistry 2002, 41, 14734–14747. [Google Scholar] [CrossRef] [PubMed]
- Kornilov, P.; Peretz, A.; Lee, Y.; Son, K.; Lee, J.H.; Refaeli, B.; Roz, N.; Rehavi, M.; Choi, S.; Attali, B. Promiscuous Gating Modifiers Target the Voltage Sensor of K(v)7.2, TRPV1, and H(v)1 Cation Channels. FASEB J. 2014, 28, 2591–2602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadezhdin, K.D.; Romanovskaia, D.D.; Sachkova, M.Y.; Oparin, P.B.; Kovalchuk, S.I.; Grishin, E.V.; Arseniev, A.S.; Vassilevski, A.A. Modular Toxin from the Lynx Spider Oxyopes Takobius: Structure of Spiderine Domains in Solution and Membrane-Mimicking Environment. Protein Sci. 2017, 26, 611–616. [Google Scholar] [CrossRef] [Green Version]
- Wormwood, K.L.; Ngounou Wetie, A.G.; Gomez, M.V.; Ju, Y.; Kowalski, P.; Mihasan, M.; Darie, C.C. Structural Characterization and Disulfide Assignment of Spider Peptide Phα1β by Mass Spectrometry. J. Am. Soc. Mass. Spectrom. 2018, 29, 827–841. [Google Scholar] [CrossRef] [PubMed]
- Lyukmanova, E.N.; Shulepko, M.A.; Shenkarev, Z.O.; Dolgikh, D.A.; Kirpichnikov, M.P. In Vitro Production of Three-Finger Neurotoxins from Snake Venoms, a Disulfide Rich Proteins. Problems and Their Solutions (Review). Russ. J. Bioorganic Chem. 2010, 36, 137–145. [Google Scholar] [CrossRef]
- Krishnarjuna, B.; Sunanda, P.; Villegas-Moreno, J.; Csoti, A.; Morales, R.A.V.; Wai, D.C.C.; Panyi, G.; Prentis, P.; Norton, R.S. A Disulfide-Stabilised Helical Hairpin Fold in Acrorhagin I: An Emerging Structural Motif in Peptide Toxins. J. Struct. Biol. 2021, 213, 107692. [Google Scholar] [CrossRef] [PubMed]
- Platzer, G.; Okon, M.; McIntosh, L.P. PH-Dependent Random Coil (1)H, (13)C, and (15)N Chemical Shifts of the Ionizable Amino Acids: A Guide for Protein PK a Measurements. J. Biomol. NMR 2014, 60, 109–129. [Google Scholar] [CrossRef]
- Schubert, M.; Labudde, D.; Oschkinat, H.; Schmieder, P. A Software Tool for the Prediction of Xaa-Pro Peptide Bond Conformations in Proteins Based on 13C Chemical Shift Statistics. J. Biomol. NMR 2002, 24, 149–154. [Google Scholar] [CrossRef]
- Shen, Y.; Bax, A. Protein Backbone and Sidechain Torsion Angles Predicted from NMR Chemical Shifts Using Artificial Neural Networks. J. Biomol. NMR 2013, 56, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Frishman, D.; Argos, P. Knowledge-Based Protein Secondary Structure Assignment. Proteins 1995, 23, 566–579. [Google Scholar] [CrossRef]
- Schmidt, E.; Güntert, P. Automated Structure Determination from NMR Spectra. Methods Mol. Biol. 2015, 1261, 303–329. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Bohlen, C.J.; Priel, A.; Zhou, S.; King, D.; Siemens, J.; Julius, D. A Bivalent Tarantula Toxin Activates the Capsaicin Receptor, TRPV1, by Targeting the Outer Pore Domain. Cell 2010, 141, 834–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chassagnon, I.R.; McCarthy, C.A.; Chin, Y.K.-Y.; Pineda, S.S.; Keramidas, A.; Mobli, M.; Pham, V.; De Silva, T.M.; Lynch, J.W.; Widdop, R.E.; et al. Potent Neuroprotection after Stroke Afforded by a Double-Knot Spider-Venom Peptide That Inhibits Acid-Sensing Ion Channel 1a. Proc. Natl. Acad. Sci. USA 2017, 114, 3750–3755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.H.; Jung, H.J.; Konishi, S.; Kim, H.H.; Park, Z.-Y.; Kim, J.I. Structure-Activity Relationships of ω-Agatoxin IVA in Lipid Membranes. Biochem. Biophys. Res. Commun. 2017, 482, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Liang, S.; Ding, J.; Liu, X.; Zhang, R.; Gu, X. Proton Nuclear Magnetic Resonance Studies on Huwentoxin-I from the Venom of the Spider Selenocosmia Huwena: 2. Three-Dimensional Structure in Solution. J. Protein. Chem. 1997, 16, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Dai, J.; Dai, L.; Deng, M.; Hu, Z.; Hu, W.; Liang, S. Function and Solution Structure of Huwentoxin-X, a Specific Blocker of N-Type Calcium Channels, from the Chinese Bird Spider Ornithoctonus Huwena*. J. Biol. Chem. 2006, 281, 8628–8635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyte, J.; Doolittle, R.F. A Simple Method for Displaying the Hydropathic Character of a Protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Rong, M.; Xiao, Y.; Liang, S. The Effects of Huwentoxin-I on the Voltage-Gated Sodium Channels of Rat Hippocampal and Cockroach Dorsal Unpaired Median Neurons. Peptides 2012, 34, 19–25. [Google Scholar] [CrossRef]
- Winterfield, J.R.; Swartz, K.J. A Hot Spot for the Interaction of Gating Modifier Toxins with Voltage-Dependent Ion Channels. J. Gen. Physiol. 2000, 116, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Salari, A.; Vega, B.S.; Milescu, L.S.; Milescu, M. Molecular Interactions between Tarantula Toxins and Low-Voltage-Activated Calcium Channels. Sci. Rep. 2016, 6, 23894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.J.; Lee, J.Y.; Kim, S.H.; Eu, Y.J.; Shin, S.Y.; Milescu, M.; Swartz, K.J.; Kim, J.I. Solution Structure and Lipid Membrane Partitioning of VSTx1, an Inhibitor of the KvAP Potassium Channel. Biochemistry 2005, 44, 6015–6023. [Google Scholar] [CrossRef]
- Ingólfsson, H.I.; Carpenter, T.S.; Bhatia, H.; Bremer, P.-T.; Marrink, S.J.; Lightstone, F.C. Computational Lipidomics of the Neuronal Plasma Membrane. Biophys. J. 2017, 113, 2271–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logashina, Y.A.; Mosharova, I.V.; Korolkova, Y.V.; Shelukhina, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Kozlov, S.A.; Stensvåg, K.; et al. Peptide from Sea Anemone Metridium Senile Affects Transient Receptor Potential Ankyrin-Repeat 1 (TRPA1) Function and Produces Analgesic Effect. J. Biol. Chem. 2017, 292, 2992–3004. [Google Scholar] [CrossRef] [Green Version]
- Logashina, Y.A.; Solstad, R.G.; Mineev, K.S.; Korolkova, Y.V.; Mosharova, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Arseniev, A.S.; et al. New Disulfide-Stabilized Fold Provides Sea Anemone Peptide to Exhibit Both Antimicrobial and TRPA1 Potentiating Properties. Toxins 2017, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Andreev, Y.A.; Kozlov, S.A.; Vassilevski, A.A.; Grishin, E.V. Cyanogen Bromide Cleavage of Proteins in Salt and Buffer Solutions. Anal. Biochem. 2010, 407, 144–146. [Google Scholar] [CrossRef]
- Kovalchuk, S.I.; Jensen, O.N.; Rogowska-Wrzesinska, A. FlashPack: Fast and Simple Preparation of Ultrahigh-Performance Capillary Columns for LC-MS. Mol. Cell Proteom. 2019, 18, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bax, A.; Vuister, G.W.; Grzesiek, S.; Delaglio, F.; Wang, A.C.; Tschudin, R.; Zhu, G. Measurement of Homo- and Heteronuclear J Couplings from Quantitative J Correlation. Meth. Enzymol. 1994, 239, 79–105. [Google Scholar] [CrossRef]
- Kazimierczuk, K.; Orekhov, V.Y. Accelerated NMR Spectroscopy by Using Compressed Sensing. Angew. Chem. Int. Ed. Engl. 2011, 50, 5556–5559. [Google Scholar] [CrossRef]
- Koradi, R.; Billeter, M.; Wüthrich, K. MOLMOL: A Program for Display and Analysis of Macromolecular Structures. J. Mol. Graph. 1996, 14, 51–55. [Google Scholar] [CrossRef]
- Cole, R.; Loria, J.P. FAST-Modelfree: A Program for Rapid Automated Analysis of Solution NMR Spin-Relaxation Data. J. Biomol. NMR 2003, 26, 203–213. [Google Scholar] [CrossRef]
- Bystrov, V.F.; Dubrovina, N.I.; Barsukov, L.I.; Bergelson, L.D. NMR Differentiation of the Internal and External Phospholipid Membrane Surfaces Using Paramagnetic Mn2+ and Eu3+ Ions. Chem. Phys. Lipids 1971, 6, 343–350. [Google Scholar] [CrossRef]
- Shenkarev, Z.O.; Shulepko, M.A.; Bychkov, M.L.; Kulbatskii, D.S.; Shlepova, O.V.; Vasilyeva, N.A.; Andreev-Andrievskiy, A.A.; Popova, A.S.; Lagereva, E.A.; Loktyushov, E.V.; et al. Water-Soluble Variant of Human Lynx1 Positively Modulates Synaptic Plasticity and Ameliorates Cognitive Impairment Associated with A7-NAChR Dysfunction. J. Neurochem. 2020, 155, 45–61. [Google Scholar] [CrossRef]
- Hu, H.; Tian, J.; Zhu, Y.; Wang, C.; Xiao, R.; Herz, J.M.; Wood, J.D.; Zhu, M.X. Activation of TRPA1 Channels by Fenamate Nonsteroidal Anti-Inflammatory Drugs. Pflugers Arch. 2010, 459, 579–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raisinghani, M.; Zhong, L.; Jeffry, J.A.; Bishnoi, M.; Pabbidi, R.M.; Pimentel, F.; Cao, D.-S.; Evans, M.S.; Premkumar, L.S. Activation Characteristics of Transient Receptor Potential Ankyrin 1 and Its Role in Nociception. Am. J. Physiol. Cell Physiol. 2011, 301, C587–C600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kneller, J.M.; Lu, M.; Bracken, C. An Effective Method for the Discrimination of Motional Anisotropy and Chemical Exchange. J. Am. Chem. Soc. 2002, 124, 1852–1853. [Google Scholar] [CrossRef]
- Pyrkov, T.V.; Chugunov, A.O.; Krylov, N.A.; Nolde, D.E.; Efremov, R.G. PLATINUM: A Web Tool for Analysis of Hydrophobic/Hydrophilic Organization of Biomolecular Complexes. Bioinformatics 2009, 25, 1201–1202. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipids | Partition Equilibrium a Kp | Langmuir Isotherm b | |
|---|---|---|---|
| KN | N | ||
| ×103·M−1 | ×106·M−1 | ||
| POPC/POPG (3:1) | 5.1 ± 0.1 | 2.8 ± 0.7 | 41 ± 1 |
| POPC/POPG (3:1), 150 mM NaCl | 0.20 ± 0.02 | 0.0087 ± 0.0046 | 41 c |
| POPC, 150 mM NaCl | 0.14 ± 0.01 | 0.0060 ± 0.0013 | 41 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyukmanova, E.N.; Mironov, P.A.; Kulbatskii, D.S.; Shulepko, M.A.; Paramonov, A.S.; Chernaya, E.M.; Logashina, Y.A.; Andreev, Y.A.; Kirpichnikov, M.P.; Shenkarev, Z.O. Recombinant Production, NMR Solution Structure, and Membrane Interaction of the Phα1β Toxin, a TRPA1 Modulator from the Brazilian Armed Spider Phoneutria nigriventer. Toxins 2023, 15, 378. https://doi.org/10.3390/toxins15060378
Lyukmanova EN, Mironov PA, Kulbatskii DS, Shulepko MA, Paramonov AS, Chernaya EM, Logashina YA, Andreev YA, Kirpichnikov MP, Shenkarev ZO. Recombinant Production, NMR Solution Structure, and Membrane Interaction of the Phα1β Toxin, a TRPA1 Modulator from the Brazilian Armed Spider Phoneutria nigriventer. Toxins. 2023; 15(6):378. https://doi.org/10.3390/toxins15060378
Chicago/Turabian StyleLyukmanova, Ekaterina N., Pavel A. Mironov, Dmitrii S. Kulbatskii, Mikhail A. Shulepko, Alexander S. Paramonov, Elizaveta M. Chernaya, Yulia A. Logashina, Yaroslav A. Andreev, Mikhail P. Kirpichnikov, and Zakhar O. Shenkarev. 2023. "Recombinant Production, NMR Solution Structure, and Membrane Interaction of the Phα1β Toxin, a TRPA1 Modulator from the Brazilian Armed Spider Phoneutria nigriventer" Toxins 15, no. 6: 378. https://doi.org/10.3390/toxins15060378