Short-Term Interactions of Noctiluca scintillans with the Toxic Dinoflagellates Dinophysis acuminata and Alexandrium minutum: Growth, Toxins and Allelopathic Effects


Abstract
:1. Introduction
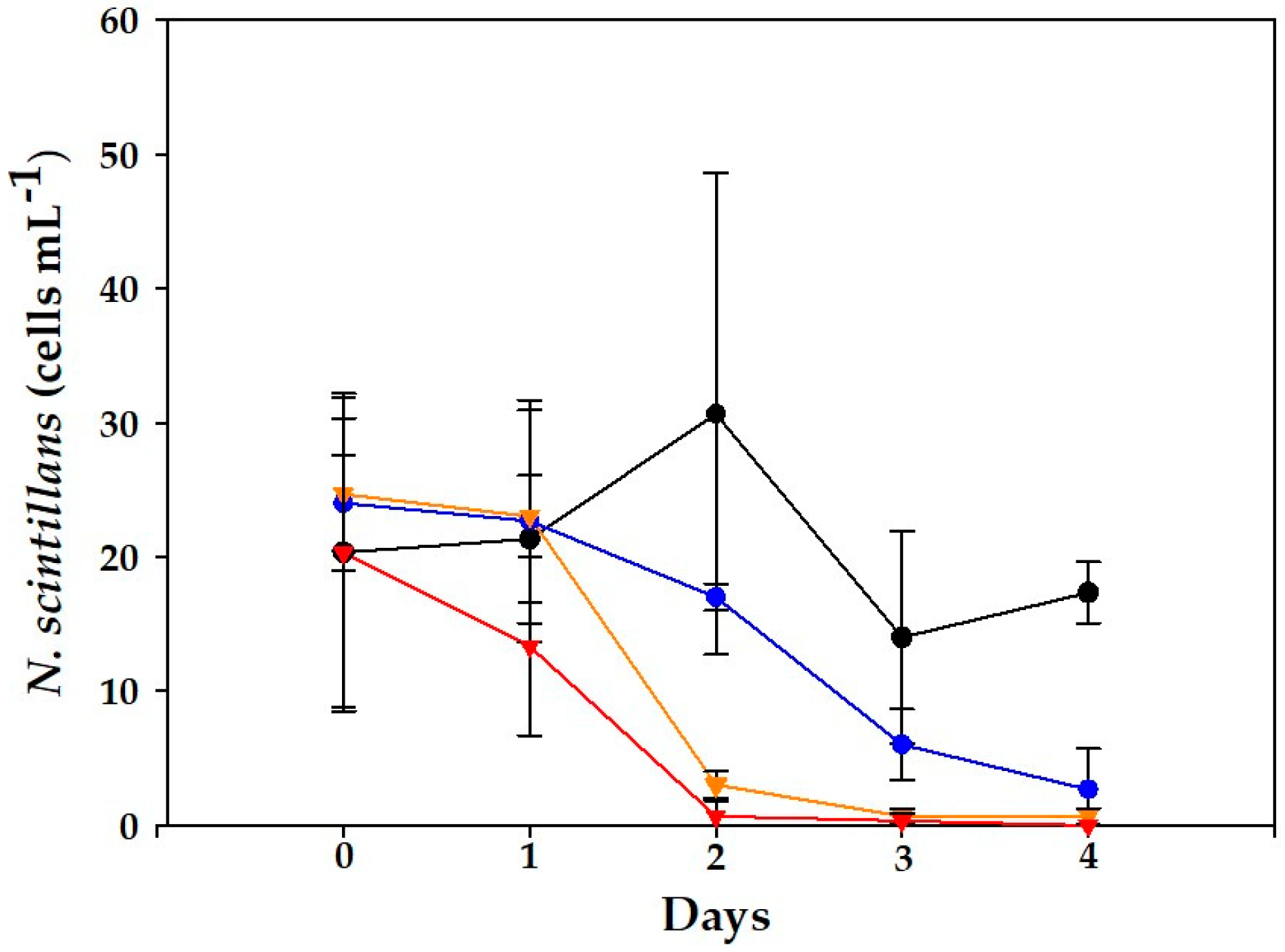
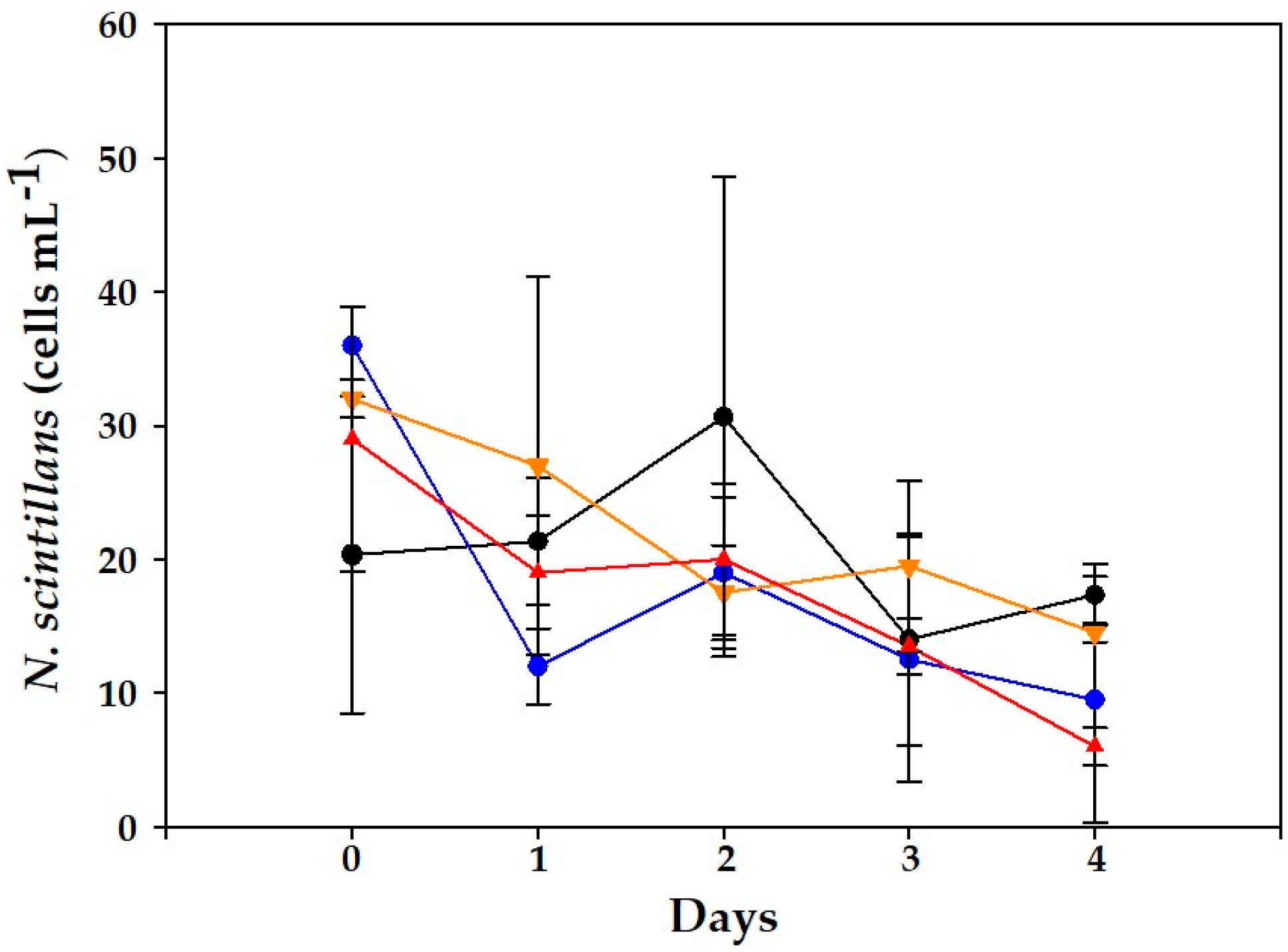
2. Results
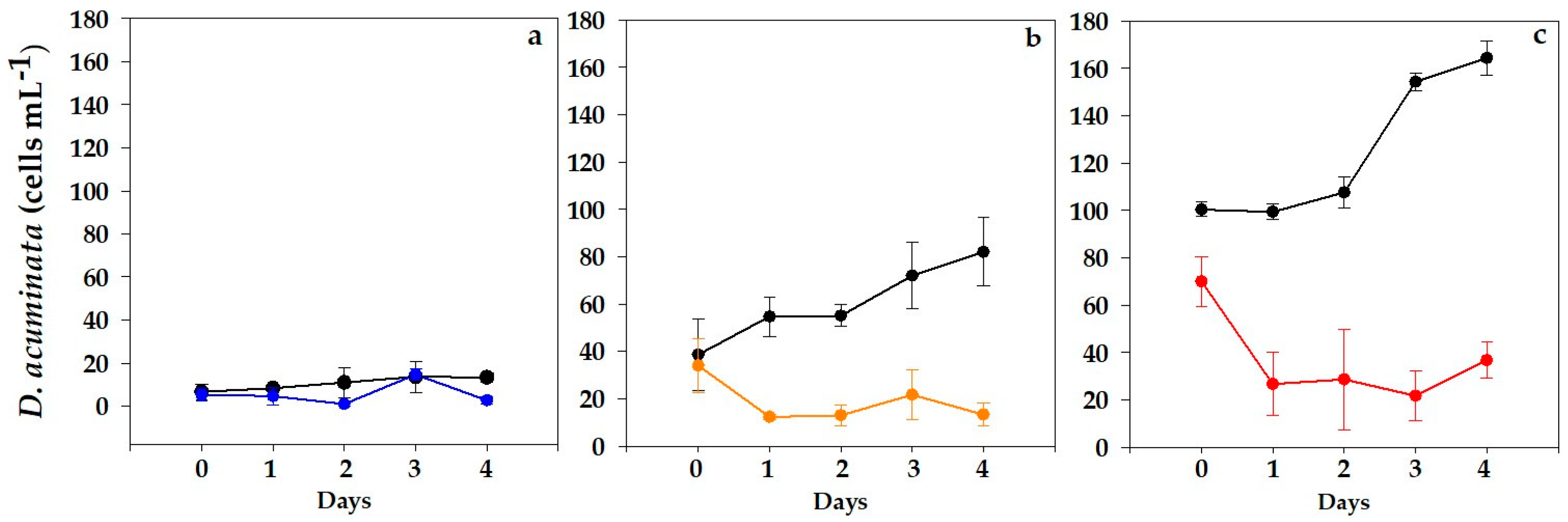
2.1. Dinophysis acuminata VGO1465 Assay
OA Analyses
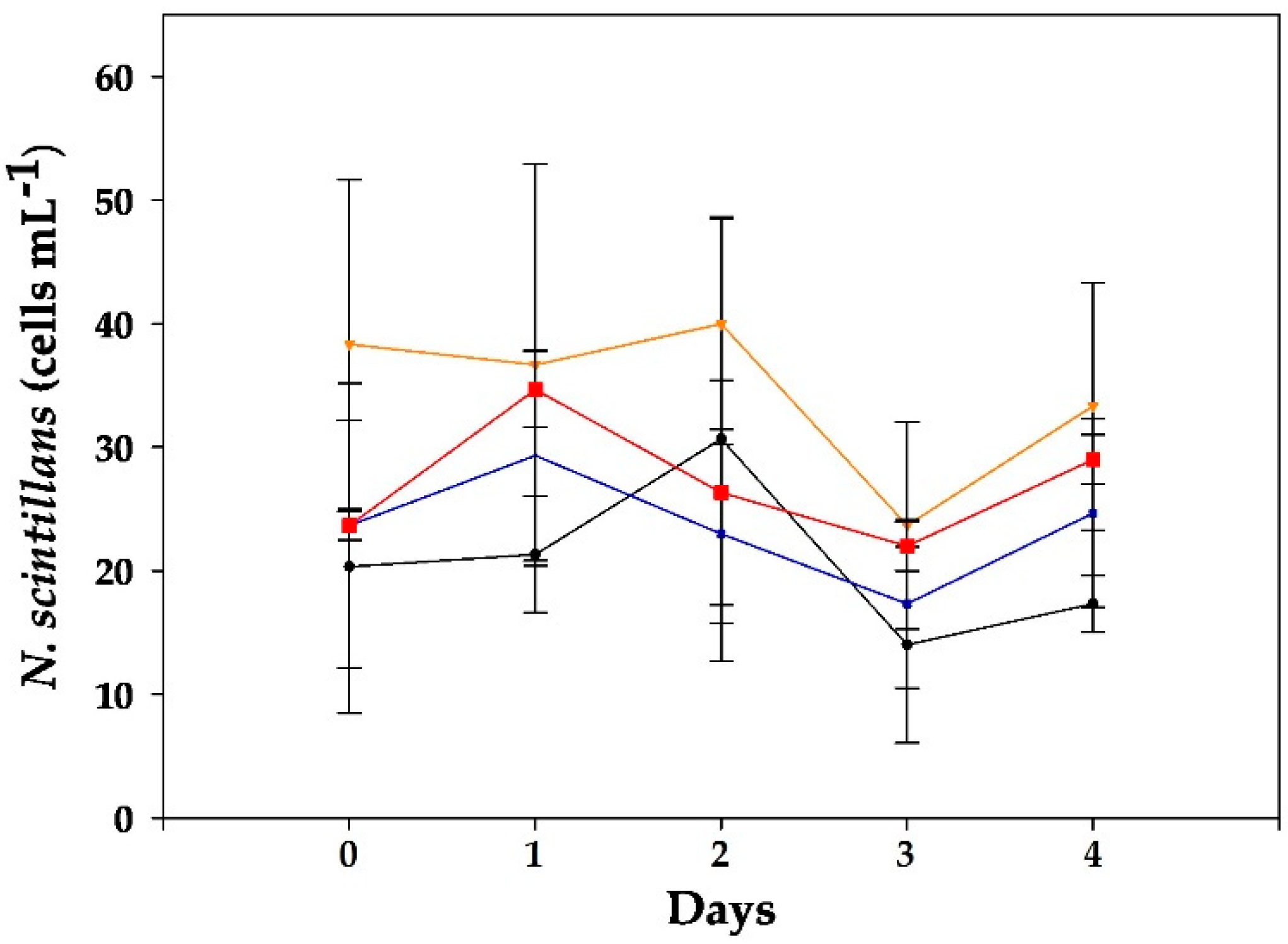
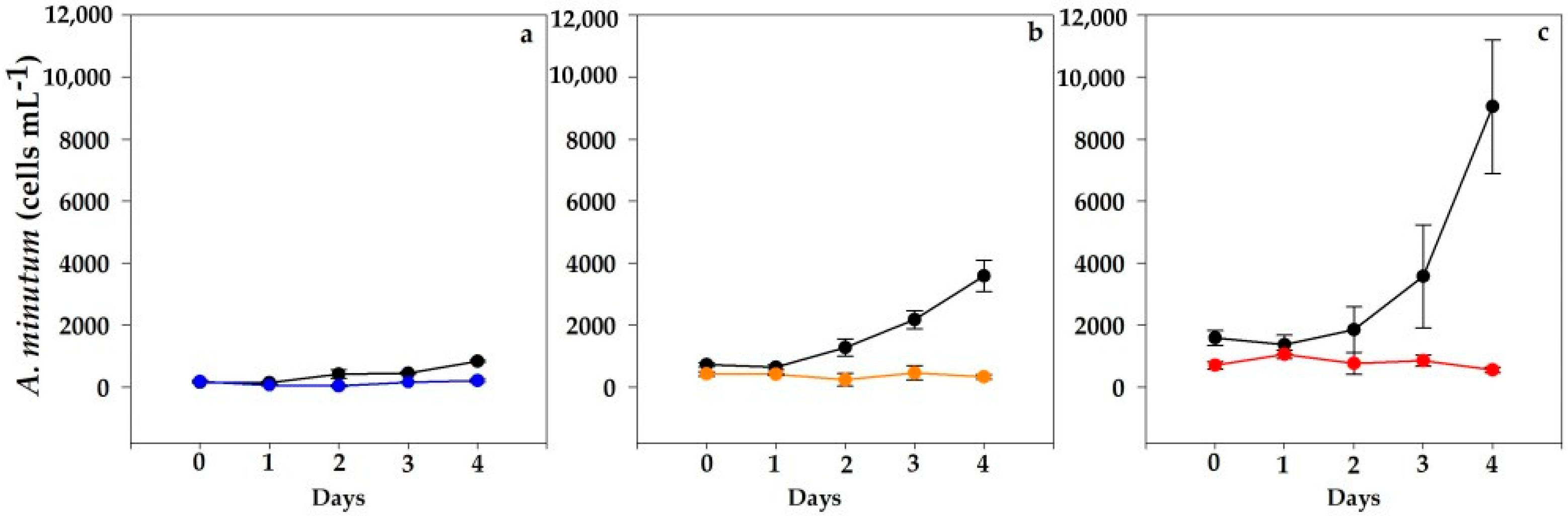
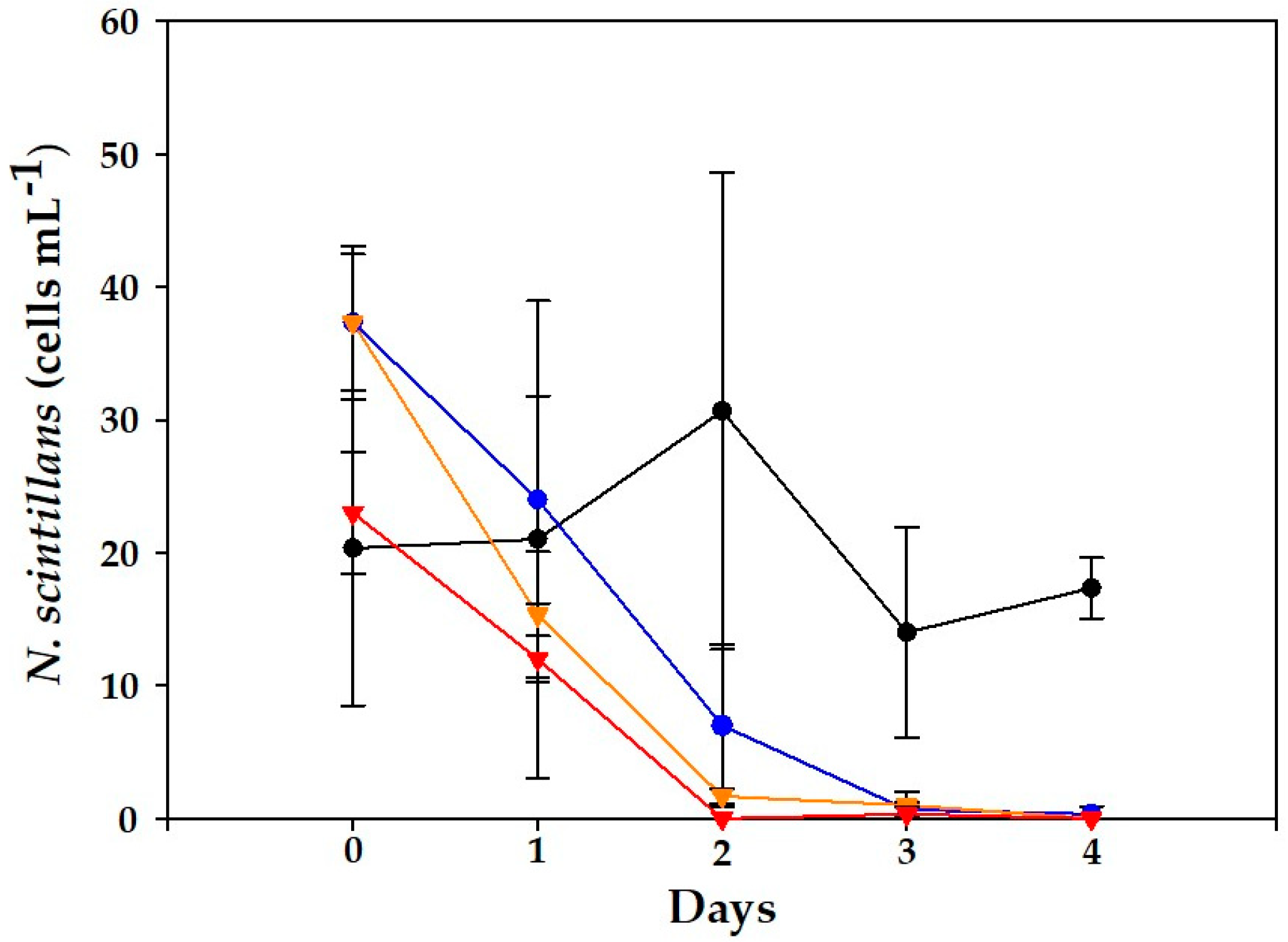
2.2. Alexandrium minutum Assay
2.2.1. VGO1435 Strain
PST Analyses
2.2.2. VGO 1439 Strain
PST Analyses
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Toxins
4.2.1. Dinophysis acuminata: OA Extraction and Detection
4.2.2. Alexandrium Minutum: PSP Toxins’ Extraction and Detection
4.3. Growth
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bolch, C.; Hallegraeff, G. Dinoflagellate Cysts in Recent Marine Sediments from Tasmania, Australia. Bot. Mar. 1990, 33, 173–192. [Google Scholar] [CrossRef]
- Fonda Umani, S.; Beran, A.; Parlato, S.; Virgilio, D.; Zollet, T.; De Olazabal, A.; Lazzarini, B.; Cabrini, M. Noctiluca scintillans MACARTNEY in the Northern Adriatic Sea: Long-Term Dynamics, Relationships with Temperature and Eutrophication, and Role in the Food Web. J. Plankton Res. 2004, 26, 545–561. [Google Scholar] [CrossRef]
- Harrison, P.J.; Furuya, K.; Glibert, P.M.; Xu, J.; Liu, H.B.; Yin, K.; Lee, J.H.W.; Anderson, D.M.; Gowen, R.; Al-Azri, A.R.; et al. Geographical Distribution of Red and Green Noctiluca Scintill. Chin. J. Ocean. Limnol. 2011, 29, 807–831. [Google Scholar] [CrossRef]
- Fukuda, Y.; Endoh, H. New Details from the Complete Life Cycle of the Red-Tide Dinoflagellate Noctiluca scintillans (Ehrenberg) McCartney. Eur. J. Protistol. 2006, 42, 209–219. [Google Scholar] [CrossRef]
- Sheng, L.; Jiang, Z.; Sun, Z.; Zhu, Y.; Zhai, H.; Ding, L.; Tong, M.; Chen, J.; Chen, Q.; Zeng, J. Noctiluca scintillans Distribution Largely Regulated by Phytoplankton Biomass in the East China Sea and Southern Yellow Sea. Front. Mar. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Smayda, T.J. What Is a Bloom? A Commentary. Limnol. Oceanogr. 1997, 42, 1132–1136. [Google Scholar] [CrossRef]
- Bravo, I.; Fraga, S.; Figueroa, R.I.; Pazos, Y.; Massanet, A.; Ramilo, I. Bloom Dynamics and Life Cycle Strategies of Two Toxic Dinoflagellates in a Coastal Upwelling System (NW Iberian Peninsula). Deep-Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 222–234. [Google Scholar] [CrossRef]
- Sato, N.E.; Hernández, D.; Viñas, M.D. Feeding Habits of Noctiluca scintillans in Coastal Waters off Buenos Aires Province, Argentina. Lat. Am. J. Aquat. Res. 2010, 38, 403–412. [Google Scholar] [CrossRef]
- Selander, E.; Thor, P.; Toth, G.; Pavia, H. Copepods Induce Paralytic Shellfish Toxin Production in Marine Dinoflagellates. Proc. R. Soc. B. 2006, 273, 1673–1680. [Google Scholar] [CrossRef]
- Harvey, E.; Menden-Deuer, S. Predator-Induced Fleeing Behaviors in Phytoplankton: A New Mechanism for Harmful Algal Bloom Formation? PLoS ONE 2012, 7, e46438. [Google Scholar] [CrossRef]
- Escalera, L.; Pazos, Y.; Moroño, Á.; Reguera, B. Noctiluca scintillans May Act as a Vector of Toxigenic Microalgae. Harmful Algae 2007, 6, 317–320. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, H.; Glibert, P.; Guo, C.; Ke, Y. Effects of Prey of Different Nutrient Quality on Elemental Nutrient Budgets in Noctiluca scintillans. Sci. Rep. 2017, 7, 7622. [Google Scholar] [CrossRef]
- Bustillos-Guzmán, J.J.; Band-Schmidt, C.J.; López-Cortés, D.J.; Hernández-Sandoval, F.E.; Núñez-Vázquez, E.; Gárate-Lizárraga, I. Grazing of the Dinoflagellate Noctiluca scintillans on the Paralytic Toxin-Producing Dinoflagellate Gymnodinium catenatum: Does Grazing Eliminate Cells during a Bloom? Cienc. Mar. 2013, 39, 291–302. [Google Scholar] [CrossRef]
- Arístegui, J.; Barton, E.D.; Álvarez-Salgado, X.A.; Santos, A.M.P.; Figueiras, F.G.; Kifani, S.; Hernández-León, S.; Mason, E.; Machú, E.; Demarcq, H. Sub-Regional Ecosystem Variability in the Canary Current Upwelling. Prog. Oceanogr. 2009, 83, 33–48. [Google Scholar] [CrossRef]
- Álvarez-Salgado, X.A.; Labarta, U.; Fernández-Reiriz, M.J.; Figueiras, F.G.; Rosón, G.; Piedracoba, S.; Filgueira, R.; Cabanas, J.M. Renewal Time and the Impact of Harmful Algal Blooms on the Extensive Mussel Raft Culture of the Iberian Coastal Upwelling System (SW Europe). Harmful Algae 2008, 7, 849–855. [Google Scholar] [CrossRef]
- Díaz, P.A.; Ruiz-Villarreal, M.; Pazos, Y.; Moita, T.; Reguera, B. Climate Variability and Dinophysis acuta Blooms in an Upwelling System. Harmful Algae 2016, 53, 145–159. [Google Scholar] [CrossRef]
- Blanco, J.; Moroño, A.; Pazos, Y.; Maneiro, J.; Mariño, J. Trends and Variations of the Abundance of Main PSP and DSP Producing Species in the Galician Rías: Environmental and Biological Influences. In Proceedings of the VIII International Conference on Harmful Algae, Vigo, Spain, 25–29 June 1997; pp. 204–207. [Google Scholar]
- Reguera, B.; Garcés, E.; Bravo, I.; Pazos, Y.; Ramilo-Rivero, M.I.; González-Gil, S. In Situ Division Rates of Several Species of Dinophysis Estimated by a Postmitotic Index. MEPS 2003, 249, 117–131. [Google Scholar] [CrossRef]
- Detoni, A.M.S.; Navarro, G.; Garrido, J.L.; Rodríguez, F.; Hernández-Urcera, J.; Caballero, I. Mapping Dinoflagellate Blooms (Noctiluca and Alexandrium) in Aquaculture Production Areas in the NW Iberian Peninsula with the Sentinel-2/3 Satellites. Sci. Total Environ. 2023, 868, 161579. [Google Scholar] [CrossRef]
- Datos Xerais de Acuicultura Mariña No 2021. Available online: https://www.pescadegalicia.gal/Publicaciones/AnuarioAcuicultura2021/Informes/5.1.1.html (accessed on 21 April 2023).
- Nogueira, E.; Bravo, I.; Montero, P.; Díaz-Tapia, P.; Calvo, S.; Ben-Gigirey, B.; Figueroa, R.I.; Garrido, J.L.; Ramilo, I.; Lluch, N.; et al. HABs in Coastal Upwelling Systems: Insights from an Exceptional Red Tide of the Toxigenic Dinoflagellate Alexandrium minutum. Ecol. Indic. 2022, 137, 108790. [Google Scholar] [CrossRef]
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in Understanding Harmful Algal Blooms: Paradigm Shifts and New Technologies for Research, Monitoring, and Management. Annu. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef]
- Chambouvet, A.; Laabir, M.; Sengco, M.; Vaquer, A.; Guillou, L. Genetic Diversity of Amoebophryidae (Syndiniales) during Alexandrium catenella/tamarense (Dinophyceae) Blooms in the Thau Lagoon (Mediterranean Sea, France). Res. Microbiol. 2011, 162, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Laabir, M.; Jauzein, C.; Genovesi, B.; Masseret, E.; Grzebyk, D.; Cecchi, P.; Vaquer, A.; Perrin, Y.; Collos, Y. Influence of Temperature, Salinity and Irradiance on the Growth and Cell Yield of the Harmful Red Tide Dinoflagellate Alexandrium catenella Colonizing Mediterranean Waters. J. Plankton Res. 2011, 33, 1550–1563. [Google Scholar] [CrossRef]
- Vale, P.; Botelho, M.J.; Rodrigues, S.M.; Gomes, S.S.; de M. Sampayo, M.A. Two Decades of Marine Biotoxin Monitoring in Bivalves from Portugal (1986–2006): A Review of Exposure Assessment. Harmful Algae 2008, 7, 11–25. [Google Scholar] [CrossRef]
- Long, M.; Krock, B.; Castrec, J.; Tillmann, U. Unknown Extracellular and Bioactive Metabolites of the Genus Alexandrium: A Review of Overlooked Toxins. Toxins 2021, 13, 905. [Google Scholar] [CrossRef]
- Cembella, A.D. Chemical Ecology of Eukaryotic Microalgae in Marine Ecosystems. Phycologia 2003, 42, 420–447. [Google Scholar] [CrossRef]
- Blanco, J.; Correa, J.; Muñíz, S.; Mariño, C. Revista Galega dos Recursos Mariños. Art. Inf. Tec. 2013, 1–55. Available online: https://www.researchgate.net/publication/236842103_Evaluacion_del_impacto_de_los_metodos_y_niveles_utilizados_para_el_control_de_toxinas_en_el_mejillon (accessed on 18 May 2023).
- Lewis, A.M.; Coates, L.N.; Turner, A.D.; Percy, L.; Lewis, J. A Review of the Global Distribution of Alexandrium minutum (Dinophyceae) and Comments on Ecology and Associated Paralytic Shellfish Toxin Profiles, with a Focus on Northern Europe. J. Phycol. 2018, 54, 581–598. [Google Scholar] [CrossRef]
- You, J.H.; Jeong, H.J.; Park, S.A.; Ok, J.H.; Kang, H.C.; Eom, S.H.; Lim, A.S. Development of an Automatic System for Cultivating the Bioluminescent Heterotrophic Dinoflagellate Noctiluca scintillans on a 100-Liter Scale. Algae 2022, 37, 149–161. [Google Scholar] [CrossRef]
- Nakamura, Y. Growth and Grazing of a Large Heterotrophic Dinoflagellate, Noctiluca scintillans, in Laboratory Cultures. J. Plankton Res. 1998, 20, 1711–1720. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Albinsson, M.E.; Dowdney, J.; Holmes, A.K.; Mansour, M.P.; Seger, A. Prey Preference, Environmental Tolerances and Ichthyotoxicity by the Red-Tide Dinoflagellate Noctiluca scintillans Cultured from Tasmanian Waters. J. Plankton Res. 2019, 41, 407–418. [Google Scholar] [CrossRef]
- Stauffer, B.; Gellene, A.; Rico, D.; Sur, C.; Caron, D. Grazing of the Heterotrophic Dinoflagellate Noctiluca scintillans on Dinoflagellate and Raphidophyte Prey. Aquat. Microb. Ecol. 2017, 80, 193–207. [Google Scholar] [CrossRef]
- Frangópulos, M.; Spyrakos, E.; Guisande, C. Ingestion and Clearance Rates of the Red Noctiluca scintillans Fed on the Toxic Dinoflagellate Alexandrium minutum (Halim). Harmful Algae 2011, 10, 304–309. [Google Scholar] [CrossRef]
- Lush, G.J.; Hallegraeff, G.M. High Toxic Potential of the Widespread Red Tide Dinoflagellate Alexandrium minutum to the Brineshrimp Artemia salina. In Proceedings of the International Conference on Toxic Phytoplankton, Sendai, Japan, 12–16 July 1995; pp. 389–392. [Google Scholar]
- Ryderheim, F.; Selander, E.; Kiørboe, T. Predator-Induced Defence in a Dinoflagellate Generates Benefits without Direct Costs. ISME J. 2021, 15, 2107–2116. [Google Scholar] [CrossRef] [PubMed]
- Granéli, E.; Hansen, P.J. Allelopathy in Harmful Algae: A Mechanism to Compete for Resources? In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2006; Volume 189, pp. 189–201. ISBN 978-3-540-32210-8. [Google Scholar]
- Wood, A.; Everroad, R.C.; Wingard, L.M. Measuring Growth Rates in Microalgal Cultures. In Algal Culturing Techniques; Elsevier Academic Press: New York, NY, USA, 2005; pp. 269–285. ISBN 978-0-12-088426-1. [Google Scholar]
- Azanza, R.V.; Cruz, L.J.; Cariño, F.A.; Blanco, A.G.; Butardo, V.M. Paralytic Shellfish Toxin Concentration and Cell Density Changes in Pyrodinium bahamense—Noctiluca scintillans Feeding Experiments. Toxicon 2010, 55, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Hamasaki, K.; Horie, M.; Tokimitsu, S.; Toda, T.; Taguchi, S. Variability in Toxicity of the Dinoflagellate Alexandrium tamarense Isolated from Hiroshima Bay, Western Japan, as a Reflection of Changing Environmental Conditions. J. Plankton Res. 2001, 23, 271–278. [Google Scholar] [CrossRef]
- Leong, S.C.Y.; Murata, A.; Nagashima, Y.; Taguchi, S. Variability in Toxicity of the Dinoflagellate Alexandrium tamarense in Response to Different Nitrogen Sources and Concentrations. Toxicon 2004, 43, 407–415. [Google Scholar] [CrossRef]
- Flynn, K.; Franco, J.M.; Fernández, P.; Reguera, B.; Zapata, M.; Wood, G.; Flynn, K.J. Changes in Toxin Content, Biomass and Pigments of the Dinoflagellate Alexandrium minutum during Nitrogen Refeeding and Growth into Nitrogen or Phosphorus Stress. Mar. Ecol. Prog. Ser. 1994, 111, 99–109. [Google Scholar] [CrossRef]
- Collos, Y.; Harrison, P.J. Acclimation and Toxicity of High Ammonium Concentrations to Unicellular Algae. Mar. Pollut. Bull. 2014, 80, 8–23. [Google Scholar] [CrossRef]
- Mafra, L.L., Jr.; Nagai, S.; Uchida, H.; Tavares, C.P.S.; Escobar, B.P.; Suzuki, T. Harmful Effects of Dinophysis to the Ciliate Mesodinium rubrum: Implications for Prey Capture. Harmful Algae 2016, 59, 82–90. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis Is a Diatom, Not a Chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Reguera, B.; Riobó, P.; Rodríguez, F.; Díaz, P.A.; Pizarro, G.; Paz, B.; Franco, J.M.; Blanco, J. Dinophysis Toxins: Causative Organisms, Distribution and Fate in Shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef] [PubMed]
- Velo-Suárez, L.; González-Gil, S.; Pazos, Y.; Reguera, B. The Growth Season of Dinophysis acuminata in an Upwelling System Embayment: A Conceptual Model Based on in Situ Measurements. Deep.-Sea Res. Part II Top. Stud. Oceanogr. 2014, 101, 141–151. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Anderson, D.M.; Cembella, A.D.; Enevoldsen, H.O. Monographs on Oceanographic Methodology. In Manual on Harmful Marine Microalgae, 2nd ed.; UNESCO: Paris, France, 2004; Volume 11, ISBN 9789231039485. [Google Scholar]
- Boundy, M.J.; Selwood, A.I.; Harwood, D.T.; McNabb, P.S.; Turner, A.D. Development of a Sensitive and Selective Liquid Chromatography–Mass Spectrometry Method for High Throughput Analysis of Paralytic Shellfish Toxins Using Graphitised Carbon Solid Phase Extraction. J. Chromatogr. A 2015, 1387, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, F.; Garrido, J.L.; Sobrino, C.; Johnsen, G.; Riobó, P.; Franco, J.; Aamot, I.; Ramilo, I.; Sanz, N.; Kremp, A. Divinyl Chlorophyll a in the Marine Eukaryotic Protist Alexandrium ostenfeldii (Dinophyceae). Environ. Microbiol. 2016, 18, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R. Culture Methods and Growth Measurements. In Division Rates in Handbook of Phycological Methods; Stein, J.R., Ed.; Chambridge University Press: Cambridge, UK, 1973; pp. 289–311. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| D. acuminata (VGO 1465) (Controls) | D. acuminata (VGO 1465) (Mixed Cultures) | |||||
|---|---|---|---|---|---|---|
| Initial abundance (cells mL−1) | 10 | 50 | 100 | 10 | 50 | 100 |
| Final total cells (day 4) | 13 ± 2.08 | 82 ± 14 | 164 ± 7 | 3 ± 2 | 13 ± 5 | 37 ± 8 |
| Intracellular OA (pg cell−1) | <LOD | 57.7 ± 9.94 | 46.6 ± 12.69 | <LOD | <LOD | 61.28 ± 9.46 |
| OA in the culture-filtrate (ng mL−1) | <LOD | 1.17 ± 0.00 | 1.93 ± 0.20 | <LOD | 1.73 ± 0.04 | 2.37 ± 0.31 |
| A. minutum (VGO1435) (Controls) | |||
|---|---|---|---|
| Initial abundance (cells mL−1) | 100 | 500 | 1000 |
| Final total cells (day 4) | 3.33 (±0.14) × 104 | 1.43 (±0.2) × 105 | 3.62 (±0.86) × 105 |
| pg STX equivalents cell−1 (day 4) | 2.27 | 2.64 | 1.89 |
| GTX4 (pg cell−1)/(molar %) | 2.75 ± 0.14/85 | 3.05 ± 0.34/81 | 2.35 ± 0.38/87 |
| GTX1 (pg cell−1)/(molar %) | 0.15 ± 0.05/7 | 0.20 ± 0.03/8 | 0.07 ± 0.04/ 4 |
| GTX3 (pg cell−1)/(molar %) | 0.29 ± 0.01/8 | 0.46 ± 0.05/11 | 0.25 ± 0.05/8 |
| GTX2 (pg cell−1)/(molar %) | 0.04 ± 0.02/1 | 0.07 ± 0.01/1 | 0.05 ± 0.00/1 |
| A. Minutum (VGO1439) (Controls) | A. Minutum (VGO1439) (Mixed Cultures) | |||
|---|---|---|---|---|
| Initial abundance (cells mL−1) | 500 | 1000 | 500 | 1000 |
| Final total cells (day 4) | 10.27 (±1.15) × 104 | 26.02 (±4.2) × 104 | 19.23 (±1.16) × 103 | 31.47 (±1.6) × 103 |
| pg STX equivalents cell−1 | 1.26 | 1.21 | 0.03 | 0.03 |
| GTX4 (pg cell−1)/(molar %) | 1.30 ± 0.46/71 | 1.24 ± 0.27/72 | 0.04 ± 0.00/ 100 | 0.02 ± 0.00/41 |
| GTX1 (pg cell−1)/(molar %) | 0.15 ± 0.03/12 | 0.15 ± 0.04/12 | <LOD | 0.02 ± 0.00/59 |
| GTX3 (pg cell−1)/(molar %) | 0.27 ± 0.06/13 | 0.26 ± 0.07/13 | <LOD | <LOD |
| GTX2 (pg cell−1)/(molar %) | 0.10 ± 0.01/ 3 | 0.08 ± 0.02/3 | <LOD | <LOD |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, S.; Riobó, P.; Rial, P.; Rodríguez, F. Short-Term Interactions of Noctiluca scintillans with the Toxic Dinoflagellates Dinophysis acuminata and Alexandrium minutum: Growth, Toxins and Allelopathic Effects. Toxins 2023, 15, 373. https://doi.org/10.3390/toxins15060373
Garrido S, Riobó P, Rial P, Rodríguez F. Short-Term Interactions of Noctiluca scintillans with the Toxic Dinoflagellates Dinophysis acuminata and Alexandrium minutum: Growth, Toxins and Allelopathic Effects. Toxins. 2023; 15(6):373. https://doi.org/10.3390/toxins15060373
Chicago/Turabian StyleGarrido, Soledad, Pilar Riobó, Pilar Rial, and Francisco Rodríguez. 2023. "Short-Term Interactions of Noctiluca scintillans with the Toxic Dinoflagellates Dinophysis acuminata and Alexandrium minutum: Growth, Toxins and Allelopathic Effects" Toxins 15, no. 6: 373. https://doi.org/10.3390/toxins15060373