In-Vitro and In-Silico Investigation for the Spent-Coffee Bioactive Phenolics as a Promising Aflatoxins Production Inhibitor



Abstract
:1. Introduction
2. Results
2.1. Characterization of SCGs Phenolic and Flavonoids
2.2. Cytotoxic Impact of SCGs Methanol Extract
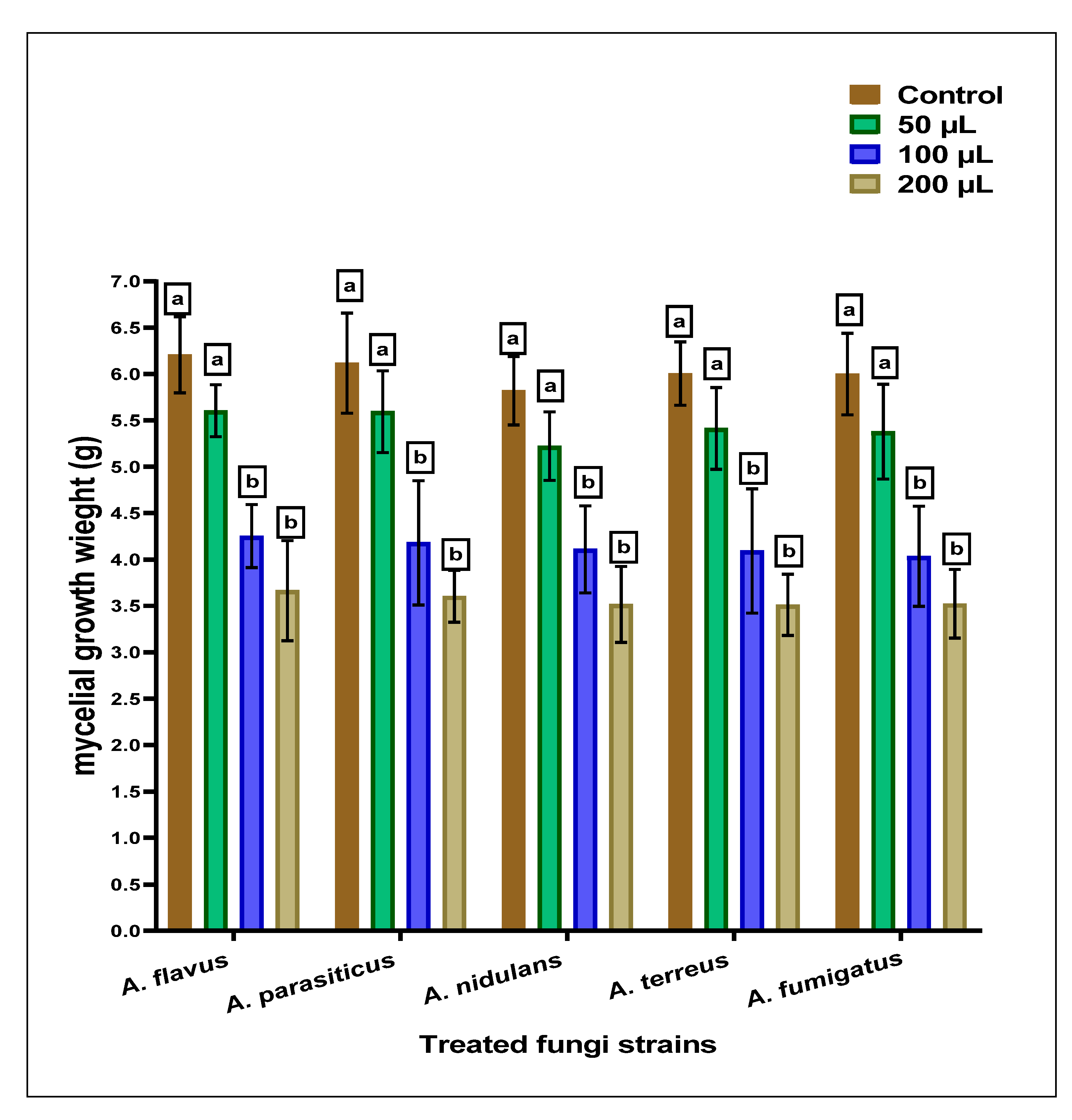
2.3. Antifungal of the SCG-Extract using Diffusion Assays
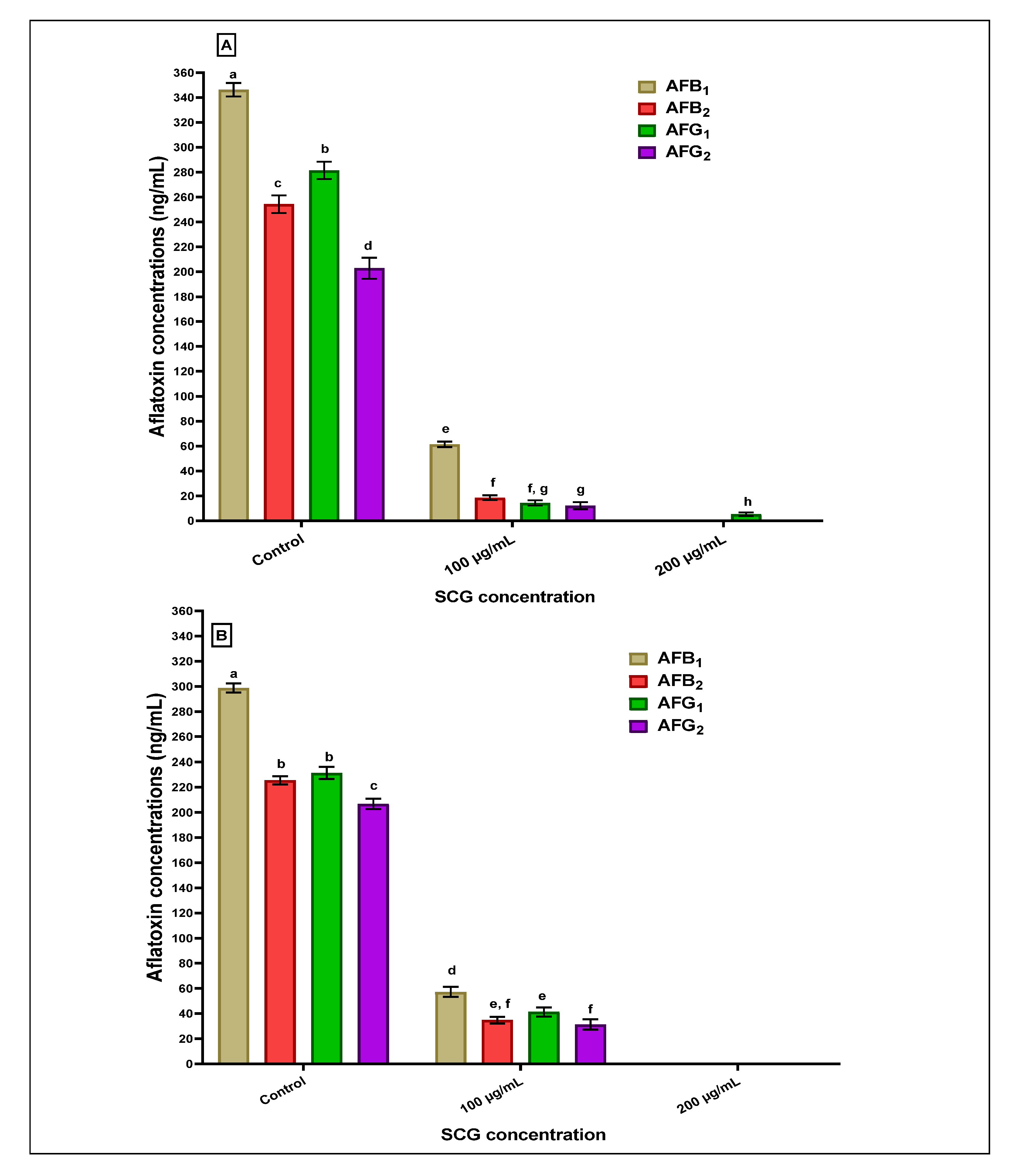
2.4. Estimation of the Anti-Aflatoxigenic SCG-Extract Effect
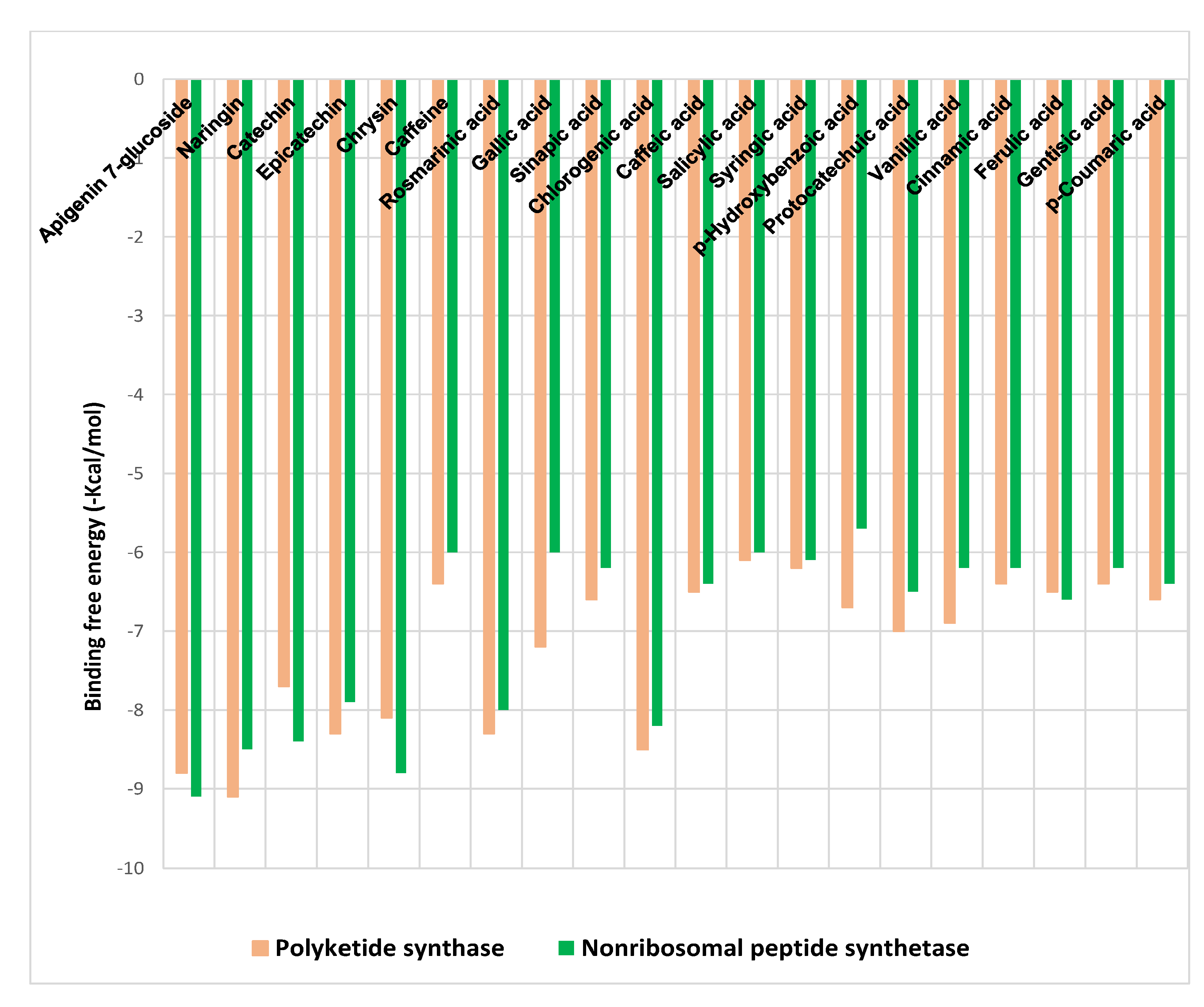
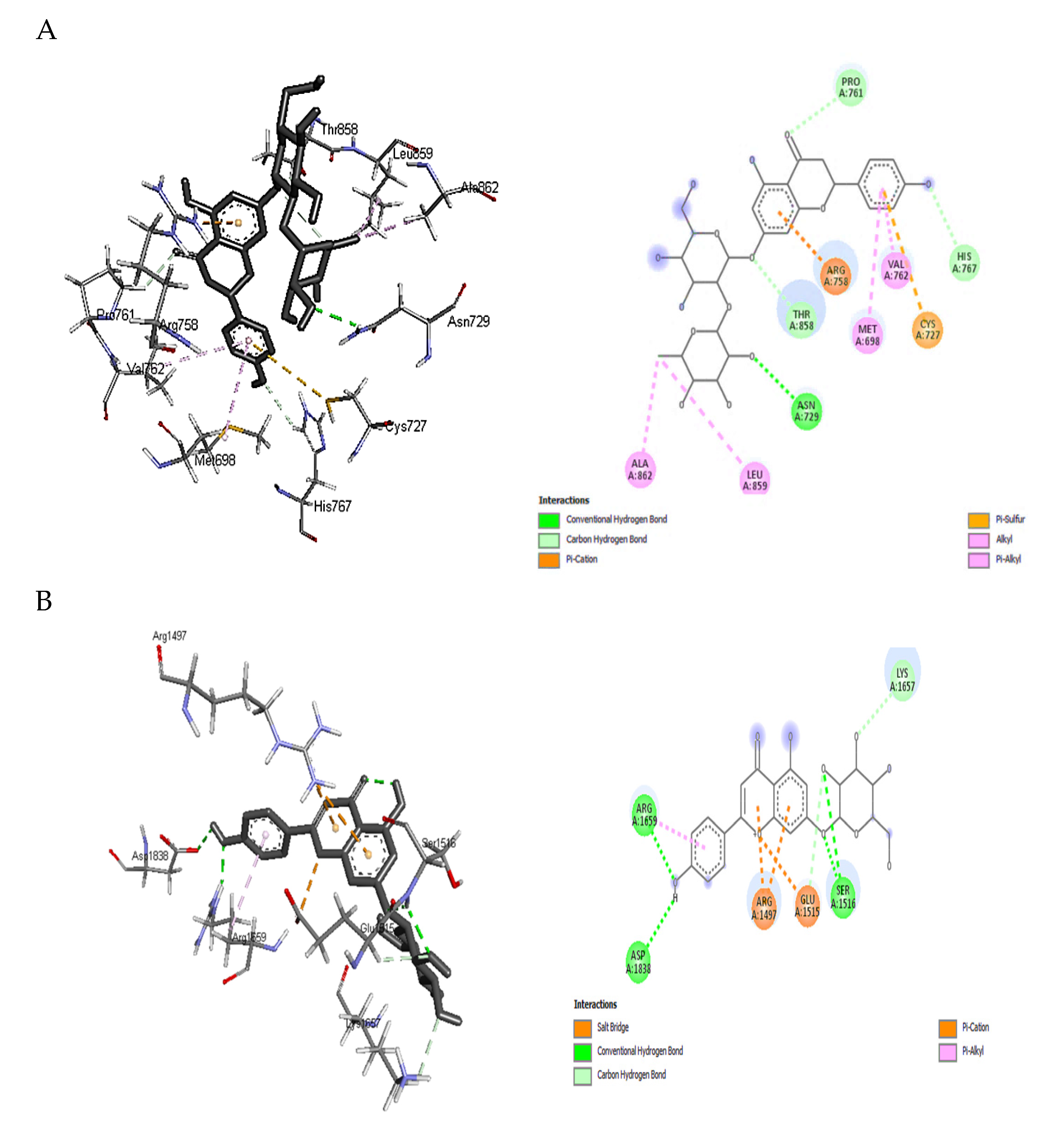
2.5. Molecular Docking Analysis
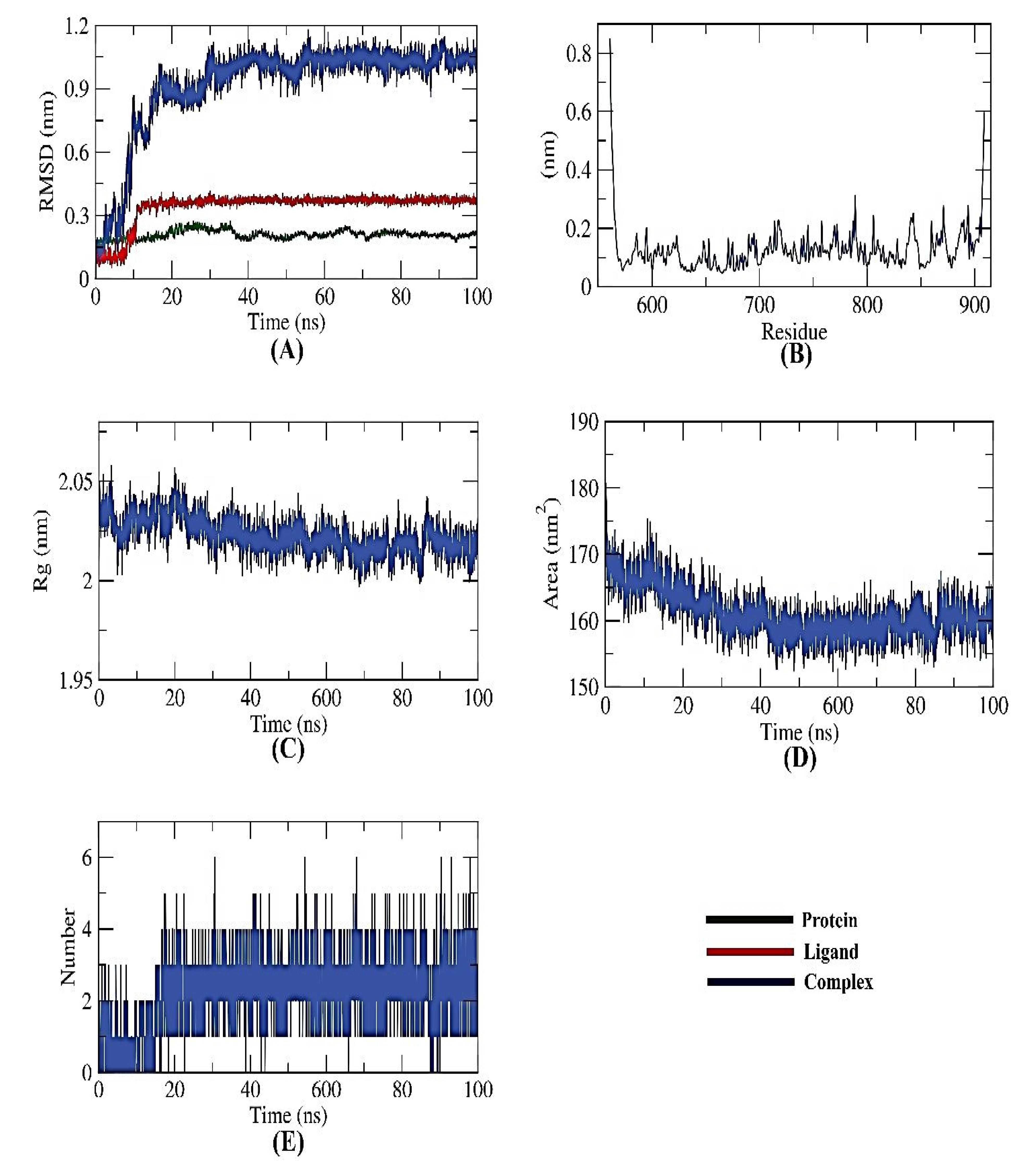
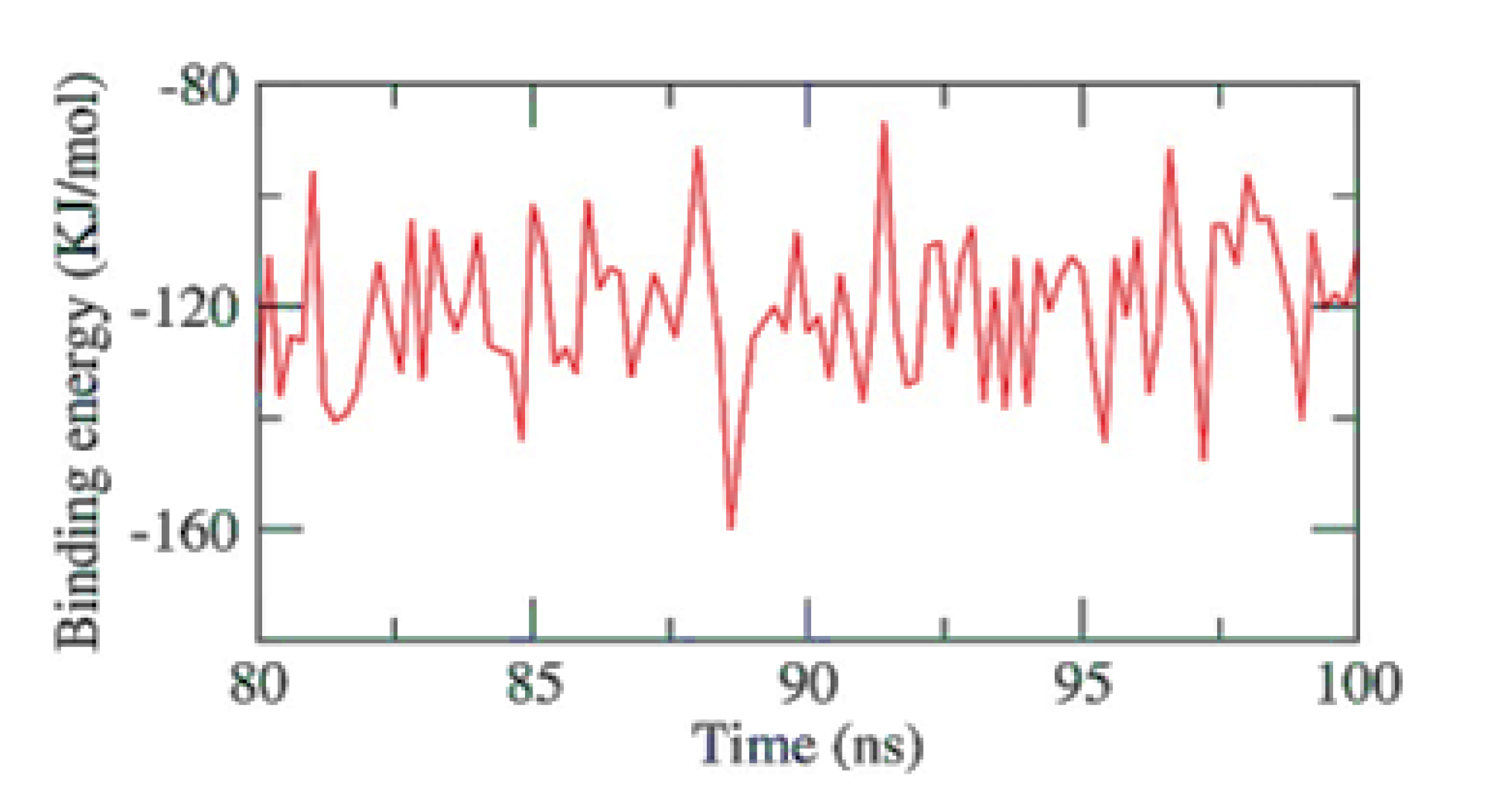
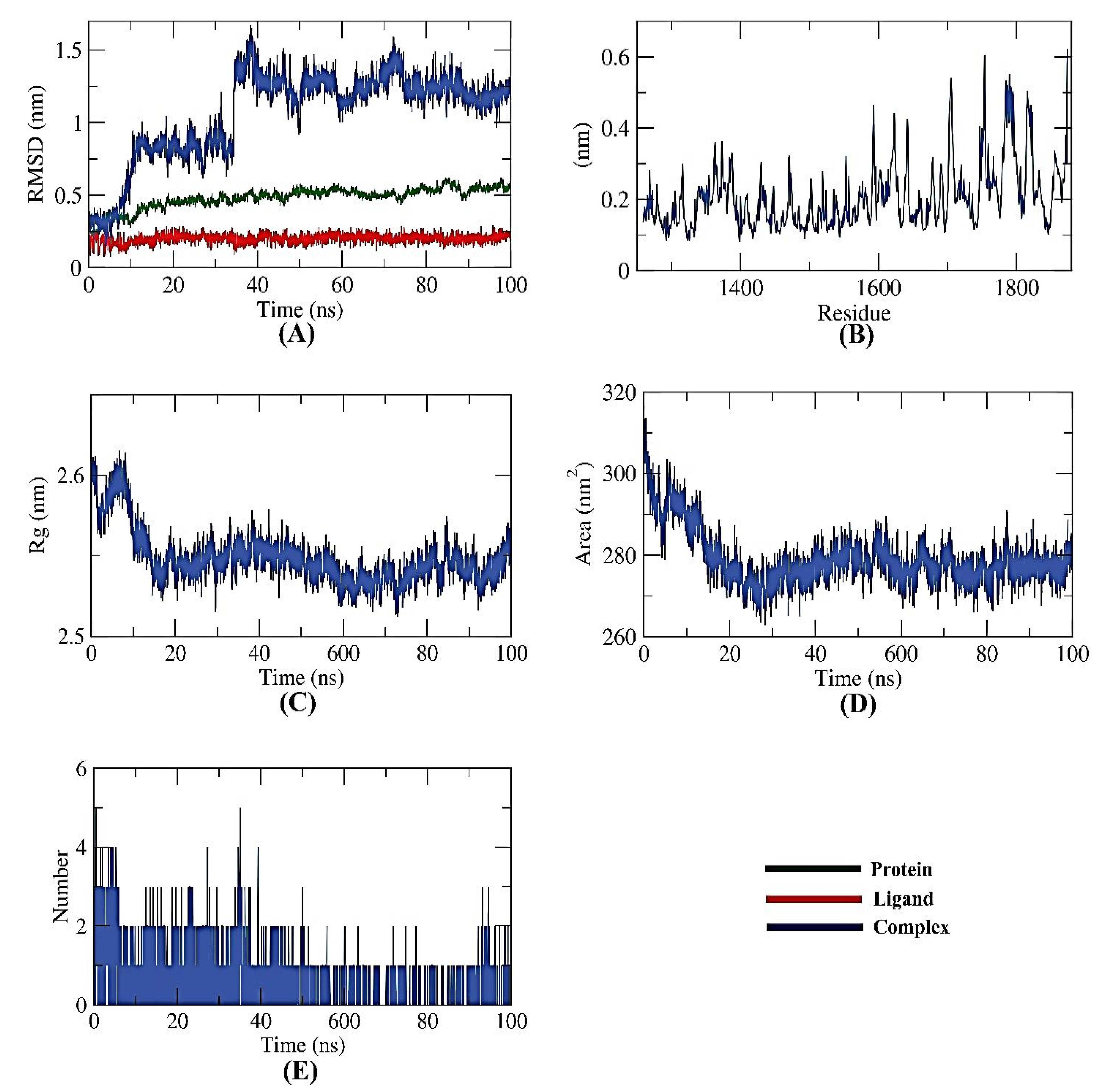
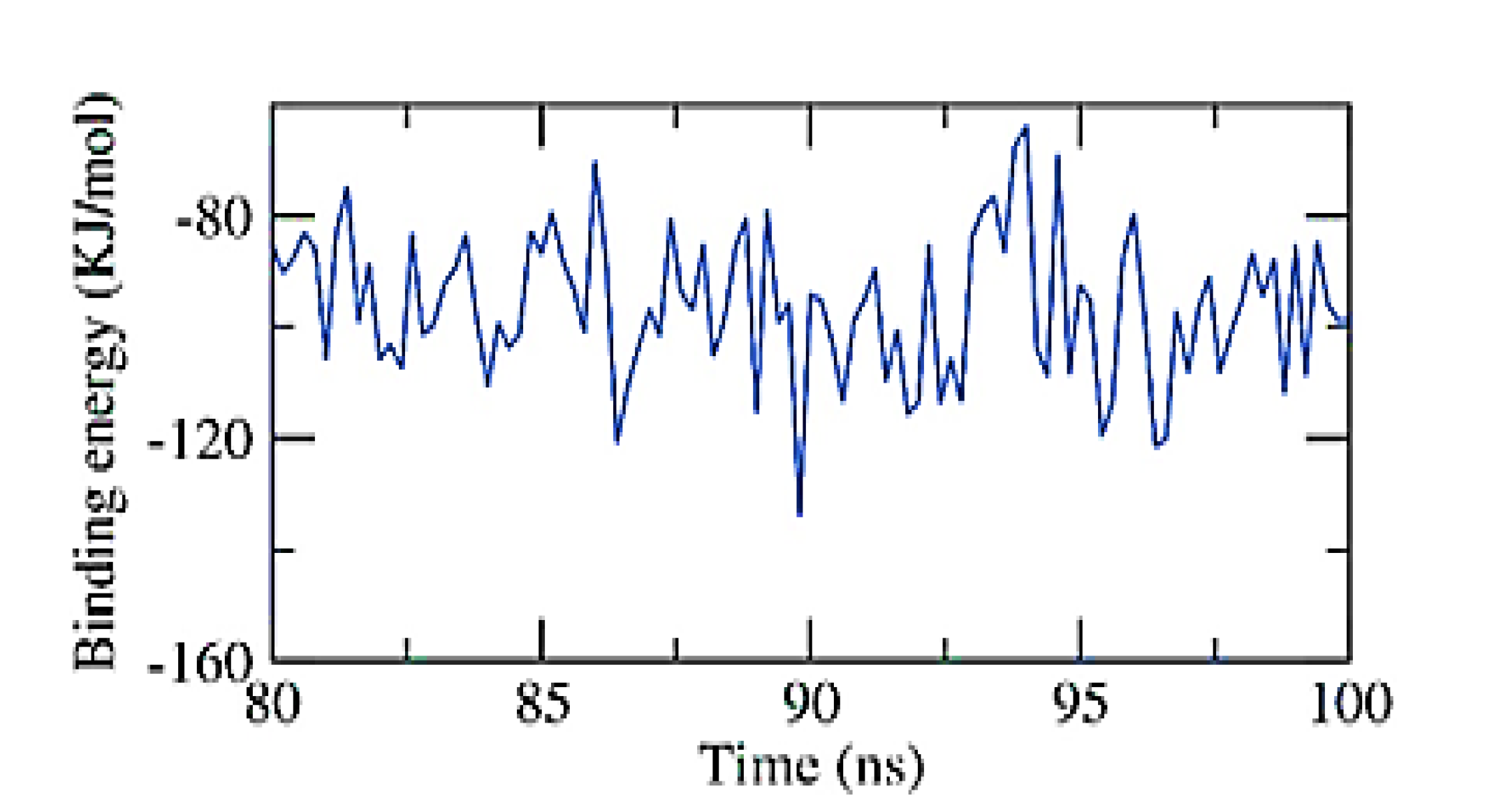
2.6. Validating Analysis of Receptor-Ligand Complexes Using MD Simulation
2.6.1. PKS-Naringin
2.6.2. NPS-Apigenin 7-Glucoside
3. Conclusions
4. Materials and Methods
4.1. Materials, Chemicals, and Microorganisms
4.2. Preparation of Spent Coffee Extract
4.3. Determination of Phenolic Acids and Flavonoids
4.4. Determination of the Cytotoxicity of SCGs
4.4.1. Determination of the Cytotoxic Effect Using the Tetrazolium-Based (MTT) Assay
4.4.2. Determination of the Cytotoxic Effect Using Sulforhodamine B (SRB) Assay
4.4.3. Determination of Selectivity Index (SI)
4.5. Determination of Antifungal Effects
4.5.1. Determination of SCG Minimal Inhibition Concentration
4.5.2. Determination of Disk and Well Diffusion Assays
4.6. The Antifungal SCG Influence Using Simulated Liquid Media
4.7. Estimation of the Changes in Aflatoxin Production
4.8. Mycotoxin Determination
4.9. Molecular Docking
4.10. MD Simulation Methodology
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liew, W.-P.-P.; Mohd-Redzwan, S. Mycotoxin: Its Impact on Gut Health and Microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretti, A.; Pascale, M.; Logrieco, A.F. Mycotoxin risks under a climate change scenario in Europe. Trends Food Sci. Technol. 2019, 84, 38–40. [Google Scholar] [CrossRef]
- Muhammad, I.; Sun, X.; Wang, H.; Li, W.; Wang, X.; Cheng, P.; Li, S.; Zhang, X.; Hamid, S. Curcumin Successfully Inhibited the Computationally Identified CYP2A6 Enzyme-Mediated Bioactivation of Aflatoxin B1 in Arbor Acres broiler. Front. Pharmacol. 2017, 8, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohd-Redzwan, S.; Jamaluddin, R.; Abd.-Mutalib, M.S.; Ahmad, Z. A mini review on aflatoxin exposure in Malaysia: Past, present and future. Front. Microbiol. 2013, 4, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damiano, S.; Jarriyawattanachaikul, W.; Girolami, F.; Longobardi, C.; Nebbia, C.; Andretta, E.; Lauritano, C.; Dabbou, S.; Avantaggiato, G.; Schiavone, A.; et al. Curcumin Supplementation Protects Broiler Chickens Against the Renal Oxidative Stress Induced by the Dietary Exposure to Low Levels of Aflatoxin B1. Front. Veter.-Sci. 2022, 8, 1659. [Google Scholar] [CrossRef]
- Jouany, J.P. Methods for preventing, decontaminating and minimizing the toxicity of mycotoxins in feeds. Anim. Feed. Sci. Technol. 2007, 137, 342–362. [Google Scholar] [CrossRef]
- Makhuvele, R.; Naidu, K.; Gbashi, S.; Thipe, V.C.; Adebo, O.A.; Njobeh, P.B. The use of plant extracts and their phytochemicals for control of toxigenic fungi and mycotoxins. Heliyon 2020, 6, e05291. [Google Scholar] [CrossRef]
- Møller, C.O.D.A.; Freire, L.; Rosim, R.E.; Margalho, L.P.; Balthazar, C.F.; Franco, L.T.; Sant’Ana, A.D.S.; Corassin, C.H.; Rattray, F.P.; de Oliveira, C.A.F. Effect of Lactic Acid Bacteria Strains on the Growth and Aflatoxin Production Potential of Aspergillus parasiticus, and Their Ability to Bind Aflatoxin B1, Ochratoxin A, and Zearalenone in vitro. Front. Microbiol. 2021, 12, 655386. [Google Scholar] [CrossRef]
- Leyva-López, N.; Lizárraga-Velázquez, C.E.; Hernández, C.; Sánchez-Gutiérrez, E.Y. Exploitation of Agro-Industrial Waste as Potential Source of Bioactive Compounds for Aquaculture. Foods 2020, 9, 843. [Google Scholar] [CrossRef]
- Badr, A.; Ali, H.; Abdel-Razek, A.; Shehata, M.; Albaridi, N. Bioactive Components of Pomegranate Oil and Their Influence on Mycotoxin Secretion. Toxins 2020, 12, 748. [Google Scholar] [CrossRef]
- Abdel-Razek, A.G.; Badr, A.N.; Alharthi, S.S.; Selim, K.A. Efficacy of Bottle Gourd Seeds’ Extracts in Chemical Hazard Reduction Secreted as Toxigenic Fungi Metabolites. Toxins 2021, 13, 789. [Google Scholar] [CrossRef]
- Abu-Sree, Y.H.; Abdel-Fattah, S.M.; Abdel-Razek, A.G.; Badr, A.N. Neoteric approach for peanuts biofilm using the merits of Moringa extracts to control aflatoxin contamination. Toxicol. Rep. 2021, 8, 1685–1692. [Google Scholar] [CrossRef] [PubMed]
- Getachew, A.T.; Chun, B.S. Influence of pretreatment and modifiers on subcritical water liquefaction of spent coffee grounds: A green waste valorization approach. J. Clean. Prod. 2017, 142, 3719–3727. [Google Scholar] [CrossRef]
- Abrahão, F.R.; Rocha, L.C.R.; Santos, T.A.; Carmo, E.L.D.; Pereira, L.A.S.; Borges, S.V.; Pereira, R.G.F.A.; Botrel, D.A. Microencapsulation of bioactive compounds from espresso spent coffee by spray drying. LWT 2019, 103, 116–124. [Google Scholar] [CrossRef]
- Kovalcik, A.; Obruca, S.; Marova, I. Valorization of spent coffee grounds: A review. Food Bioprod. Process. 2018, 110, 104–119. [Google Scholar] [CrossRef]
- Loi, M.; Paciolla, C.; Logrieco, A.F.; Mulè, G. Plant Bioactive Compounds in Pre- and Postharvest Management for Aflatoxins Reduction. Front. Microbiol. 2020, 11, 243. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.P.; Jaiswal, A.K.; Kumar, A.; Gupta, V.; Prakash, B. Untangling the multi-regime molecular mechanism of verbenol-chemotype Zingiber officinale essential oil against Aspergillus flavus and aflatoxin B1. Sci. Rep. 2021, 11, 1–20. [Google Scholar] [CrossRef]
- Choi, B.; Koh, E. Spent coffee as a rich source of antioxidative compounds. Food Sci. Biotechnol. 2017, 26, 921–927. [Google Scholar] [CrossRef]
- Ho, K.-V.; Schreiber, K.L.; Park, J.; Vo, P.; Lei, Z.; Sumner, L.W.; Brown, C.R.; Lin, C.-H. Identification and Quantification of Bioactive Molecules Inhibiting Pro-inflammatory Cytokine Production in Spent Coffee Grounds Using Metabolomics Analyses. Front. Pharmacol. 2020, 11, 229. [Google Scholar] [CrossRef]
- Zengin, G.; Sinan, K.I.; Mahomoodally, M.F.; Angeloni, S.; Mustafa, A.M.; Vittori, S.; Maggi, F.; Caprioli, G. Chemical Composition, Antioxidant and Enzyme Inhibitory Properties of Different Extracts Obtained from Spent Coffee Ground and Coffee Silverskin. Foods 2020, 9, 713. [Google Scholar] [CrossRef]
- Gigliobianco, M.R.; Campisi, B.; Peregrina, D.V.; Censi, R.; Khamitova, G.; Angeloni, S.; Caprioli, G.; Zannotti, M.; Ferraro, S.; Giovannetti, R.; et al. Optimization of the Extraction from Spent Coffee Grounds Using the Desirability Approach. Antioxidants 2020, 9, 370. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, S.; Freschi, M.; Marrazzo, P.; Hrelia, S.; Beghelli, D.; Juan-García, A.; Juan, C.; Caprioli, G.; Sagratini, G.; Angeloni, C. Antioxidant and Anti-Inflammatory Profiles of Spent Coffee Ground Extracts for the Treatment of Neurodegeneration. Oxidative Med. Cell. Longev. 2021, 2021, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Vajrabhaya, L.-O.; Korsuwannawong, S. Cytotoxicity evaluation of a Thai herb using tetrazolium (MTT) and sulforhodamine B (SRB) assays. J. Anal. Sci. Technol. 2018, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Prakash, B.; Kumar, A.; Singh, P.P.; Songachan, L.S. Antimicrobial and antioxidant properties of phytochemicals: Current status and future perspective. In Functional and Preservative Properties of Phytochemicals; Academic Press: Cambridge, MA, USA, 2020; pp. 1–45. [Google Scholar]
- Makhuvele, R.; Matshoga, R.; Antonissen, R.; Pieters, L.; Verschaeve, L.; Elgorashi, E. Genotoxicity and Antigenotoxicity of selected South African indigenous plants. S. Afr. J. Bot. 2018, 114, 89–99. [Google Scholar] [CrossRef]
- Redondo-Blanco, S.; Fernández, J.; López-Ibáñez, S.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Plant Phytochemicals in Food Preservation: Antifungal Bioactivity: A Review. J. Food Prot. 2020, 83, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Anjorin, T.S.; Salako, E.A.; Makun, H.A. Control of toxigenic fungi and mycotoxins with phytochemicals: Potentials and challenges. In Mycotoxin and Food Safety in Developing Countries; BoD–Books on Demand: Paris, France, 2013; p. 181. [Google Scholar]
- Giorni, P.; Rastelli, S.; Fregonara, S.; Bertuzzi, T. Monitoring Phenolic Compounds in Rice during the Growing Season in Relation to Fungal and Mycotoxin Contamination. Toxins 2020, 12, 341. [Google Scholar] [CrossRef]
- Samapundo, S.; De Meulenaer, B.; Osei-Nimoh, D.; Lamboni, L.Y.; Debevere, J.; Devlieghere, F. Can phenolic compounds be used for the protection of corn from fungal invasion and mycotoxin contamination during storage? Food Microbiol. 2007, 24, 465–473. [Google Scholar] [CrossRef]
- Nesci, A.V.; Etcheverry, M.G. Control ofAspergillus growth and aflatoxin production using natural maize phytochemicals under different conditions of water activity. Pest Manag. Sci. 2006, 62, 775–784. [Google Scholar] [CrossRef]
- Ferruz, E.; Atanasova-Pénichon, V.; Bonnin-Verdal, M.; Marchegay, G.; Pinson-Gadais, L.; Ducos, C.; Lorán, S.; Ariño, A.; Barreau, C.; Richard-Forget, F. Effects of Phenolic Acids on the Growth and Production of T-2 and HT-2 Toxins by Fusarium langsethiae and F. sporotrichioides. Molecules 2016, 21, 449. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, O.S.; Tardif, C.; Rouger, C.; Atanasova, V.; Richard-Forget, F.; Waffo-Téguo, P. Naturally occurring phenolic compounds as promising antimycotoxin agents: Where are we now? Compr. Rev. Food Sci. Food Saf. 2022, 21, 1161–1197. [Google Scholar] [CrossRef]
- Bavaro, S.; D’Antuono, I.; Cozzi, G.; Haidukowski, M.; Cardinali, A.; Logrieco, A. Inhibition of aflatoxin B1 production by verbascoside and other olive polyphenols. World Mycotoxin J. 2016, 9, 545–553. [Google Scholar] [CrossRef]
- Moon, Y.-S.; Kim, H.-M.; Chun, H.S.; Lee, S.-E. Organic acids suppress aflatoxin production via lowering expression of aflatoxin biosynthesis-related genes in Aspergillus flavus. Food Control 2018, 88, 207–216. [Google Scholar] [CrossRef]
- Pizzolitto, R.P.; Barberis, C.L.; Dambolena, J.S.; Herrera, J.M.; Zunino, M.P.; Magnoli, C.E.; Rubinstein, H.R.; Zygadlo, J.A.; Dalcero, A.M. Inhibitory Effect of Natural Phenolic Compounds on Aspergillus parasiticusGrowth. J. Chem. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.; Zhang, Y.Q.; Muend, S.; Rao, R. Mechanism of Antifungal Activity of Terpenoid Phenols Resembles Calcium Stress and Inhibition of the TOR Pathway. Antimicrob. Agents Chemother. 2010, 54, 5062–5069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Larraínzar, M.; Rúa, J.; Caro, I.; de Castro, C.; de Arriaga, D.; García-Armesto, M.R.; del Valle, P. Evaluation of antimicrobial and antioxidant activities of natural phenolic compounds against foodborne pathogens and spoilage bacteria. Food Control 2012, 26, 555–563. [Google Scholar] [CrossRef]
- de Freitas, R.F.; Schapira, M. A systematic analysis of atomic protein–ligand interactions in the PDB. Med. Chem. Commun. 2017, 8, 1970–1981. [Google Scholar] [CrossRef] [Green Version]
- Gallivan, J.P.; Dougherty, D.A. A Computational Study of Cation−π Interactions vs Salt Bridges in Aqueous Media: Implications for Protein Engineering. J. Am. Chem. Soc. 2000, 122, 870–874. [Google Scholar] [CrossRef]
- Kumar, M. In silico efficacy of [S]-8-gingerol (a derivative) with 6-gingerol against PT-domain of Polyketide synthase A (PksA). IP Int. J. Med. Microbiol. Trop. Dis. 2021, 7, 62–64. [Google Scholar] [CrossRef]
- Labib, M.M.; Amin, M.; Alzohairy, A.; Elashtokhy, M.; Samir, O.; Saleh, I.; Arif, I.; Osman, G.; Hassanein, S. In silico Targeting, inhibition and analysis of polyketide synthase enzyme in Aspergillus ssp. Saudi J. Biol. Sci. 2020, 27, 3187–3198. [Google Scholar] [CrossRef]
- Das, S.; Singh, V.K.; Dwivedy, A.K.; Chaudhari, A.K.; Deepika; Dubey, N.K. Eugenol loaded chitosan nanoemulsion for food protection and inhibition of Aflatoxin B1 synthesizing genes based on molecular docking. Carbohydr. Polym. 2021, 255, 117339. [Google Scholar] [CrossRef] [PubMed]
- Farouk, A.; Abdel-Razek, A.G.; Gromadzka, K.; Badr, A.N. Prevention of Aflatoxin Occurrence Using Nuts-Edible Coating of Ginger Oil Nanoemulsions and Investigate the Molecular Docking Strategy. Plants 2022, 11, 2228. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mao, H.; Hu, C.; Tron, T.; Lin, J.; Wang, J.; Sun, B. Molecular docking studies and in vitro degradation of four aflatoxins (AFB1, AFB2, AFG1, and AFG2) by a recombinant laccase from Saccharomyces cerevisiae. J. Food Sci. 2020, 85, 1353–1360. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Ballesteros, L.F.; Martins, S.; Teixeira, J. Extraction of antioxidant phenolic compounds from spent coffee grounds. Sep. Purif. Technol. 2011, 83, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Stuper-Szablewska, K.; Kurasiak-Popowska, D.; Nawracała, J.; Perkowski, J. Response of non-enzymatic antioxidative mechanisms to stress caused by infection with Fusarium fungi and chemical protection in different wheat genotypes. Chem. Ecol. 2017, 33, 949–962. [Google Scholar] [CrossRef]
- Berkow, E.L.; Lockhart, S.R.; Ostrosky-Zeichner, L. Antifungal Susceptibility Testing: Current Approaches. Clin. Microbiol. Rev. 2020, 33, e00069-19. [Google Scholar] [CrossRef] [PubMed]
- Shehata, M.G.; Badr, A.N.; El Sohaimy, S.A.; Asker, D.; Awad, T.S. Characterization of antifungal metabolites produced by novel lactic acid bacterium and their potential application as food biopreservatives. Ann. Agric. Sci. 2019, 64, 71–78. [Google Scholar] [CrossRef]
- Alastruey-Izquierdo, A.; Melhem, M.S.; Bonfietti, L.X.; Rodriguez-Tudela, J.L. Susceptibility test for fungi: Clinical and laboratorial correlations in medical mycology. Rev. Inst. Med. Trop. 2015, 57 (Suppl. 19), 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badr, A.N.; Stepien, L.; Drzewiecka, K.; Alharthi, S.S.; Selim, K.; Abdel-Razek, A.G. Synergistic Impact of Bioactive Byproduct Extract Leads to Anti-Fusarium and Anti-Mycotoxin Secretion. J. Fungi 2022, 8, 30. [Google Scholar] [CrossRef]
- Hafez, E.; El-Aziz, N.M.A.; Darwish, A.M.G.; Shehata, M.G.; Ibrahim, A.A.; Elframawy, A.M.; Badr, A.N. Validation of New ELISA Technique for Detection of Aflatoxin B1 Contamination in Food Products versus HPLC and VICAM. Toxins 2021, 13, 747. [Google Scholar] [CrossRef] [PubMed]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Grimm, M.; Dai, W.-T.; Hou, M.-C.; Xiao, Z.-X.; Cao, Y. CB-Dock: A web server for cavity detection-guided protein–ligand blind docking. Acta Pharmacol. Sin. 2020, 41, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Iskineyeva, A.; Fazylov, S.; Bakirova, R.; Sarsenbekova, A.; Pustolaikina, I.; Seilkhanov, O.; Alsfouk, A.A.; Elkaeed, E.B.; Eissa, I.H.; Metwaly, A.M. Combined In Silico and Experimental Investigations of Resveratrol Encapsulation by Beta-Cyclodextrin. Plants 2022, 11, 1678. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Acid Contents of the SCG Extract | |||
|---|---|---|---|
| Compound | Quantities (µg/g) | Compound | Quantities (µg/g) |
| Gallic acid | 34.83 ± 1.05 | Sinapic acid | 11.9 ± 0.94 |
| Protocatechuic acid | 2.75 ± 0.41 | (S)-(−)-Rosmarinic acid | 0.7 ± 0.11 |
| p-Hydroxybenzoic acid | 5.42 ± 0.67 | Ferulic acid | 0.45 ± 0.08 |
| Gentisic acid | 0.38 ± 0.06 | Salicylic acid | 8.16 ± 0.73 |
| Chlorogenic acid | 9.31 ± 0.94 | p-coumaric acid | 0.22 ± 0.03 |
| Caffeic acid | 8.58 ± 0.56 | Cinnamic acid | 1.073 ± 0.55 |
| Syringic acid | 3.41 ± 0.41 | (R)-(+)-Rosmarinic acid | 176.43 ± 2.41 |
| Vanillic acid | 2.26 ± 0.04 | - | - |
| Flavonoids contents of the SCG extract | |||
| Compound | Quantities (µg/g) | Compound | Quantities (µg/g) |
| Catechin | 18.78 ± 0.89 | ||
| Epicatechin | 9.49 ± 1.02 | Apigenin-7-glucoside | 1717.05 ± 5.76 |
| Naringin | 97.27 ± 1.97 | Chrysin | 1.06 ± 0.03 |
| Alkaloid contents of the SCG extract | |||
| Compound | (µg/g) | ||
| Caffeine | 1592.82 ± 3.77 | ||
| Extract | Cell Lines | IC50 (μg/mL) | SI |
|---|---|---|---|
| Cisplatin | HepG2 | 66.69 | 1.31 |
| HCT-116 | 58.85 | 1.49 | |
| OEC | 87.67 | - | |
| SCG extract (MTT) | HepG2 | 341.3 | 2.30 |
| HCT-116 | 250.4 | 3.12 | |
| OEC | 784.3 | - | |
| SCG extract (SRB) | HepG2 | 347.1 | 1.94 |
| HCT-116 | 327.2 | 2.05 | |
| OEC | 672.3 | - |
| Fungi Strains | Disk Diffusion (ZID; mm) | Well Diffusion (ZID; mm) | MFC (µg/mL) |
|---|---|---|---|
| Aspergillus flavus ITEM 698 | 12.81 ± 1.71 c | 13.63 ± 2.05 b | 460 |
| Aspergillus parasiticus ATCC 15517 | 13.57 ± 1.54 b,c | 14.21 ± 1.46 a,b | 420 |
| Aspergillus nidulans ATCC 26209 | 14.05 ± 1.12 a,b | 14.34 ± 1.27 a,b | 380 |
| Aspergillus terreus ATCC 1012 | 13.88 ± 1.41 b,c | 14.23 ± 1.02 a,b | 390 |
| Aspergillus fumigatus ATCC 1022 | 15.64 ± 1.08 a | 15.02 ± 1.14 a | 380 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farouk, A.; Alsulami, T.; Ali, H.S.; Badr, A.N. In-Vitro and In-Silico Investigation for the Spent-Coffee Bioactive Phenolics as a Promising Aflatoxins Production Inhibitor. Toxins 2023, 15, 225. https://doi.org/10.3390/toxins15030225
Farouk A, Alsulami T, Ali HS, Badr AN. In-Vitro and In-Silico Investigation for the Spent-Coffee Bioactive Phenolics as a Promising Aflatoxins Production Inhibitor. Toxins. 2023; 15(3):225. https://doi.org/10.3390/toxins15030225
Chicago/Turabian StyleFarouk, Amr, Tawfiq Alsulami, Hatem S. Ali, and Ahmed Noah Badr. 2023. "In-Vitro and In-Silico Investigation for the Spent-Coffee Bioactive Phenolics as a Promising Aflatoxins Production Inhibitor" Toxins 15, no. 3: 225. https://doi.org/10.3390/toxins15030225