Functional and Proteomic Insights into Aculeata Venoms

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Transcriptome
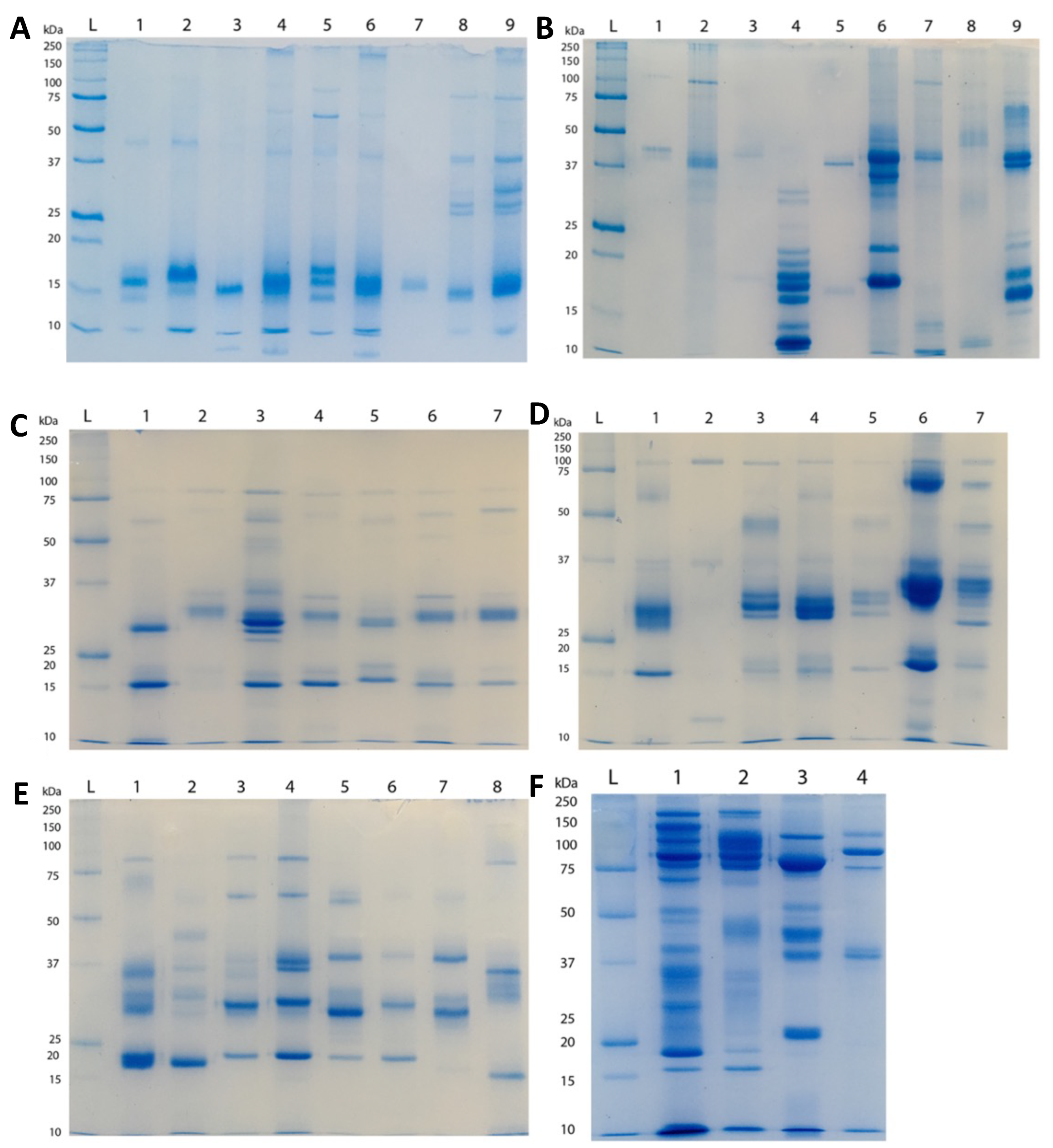
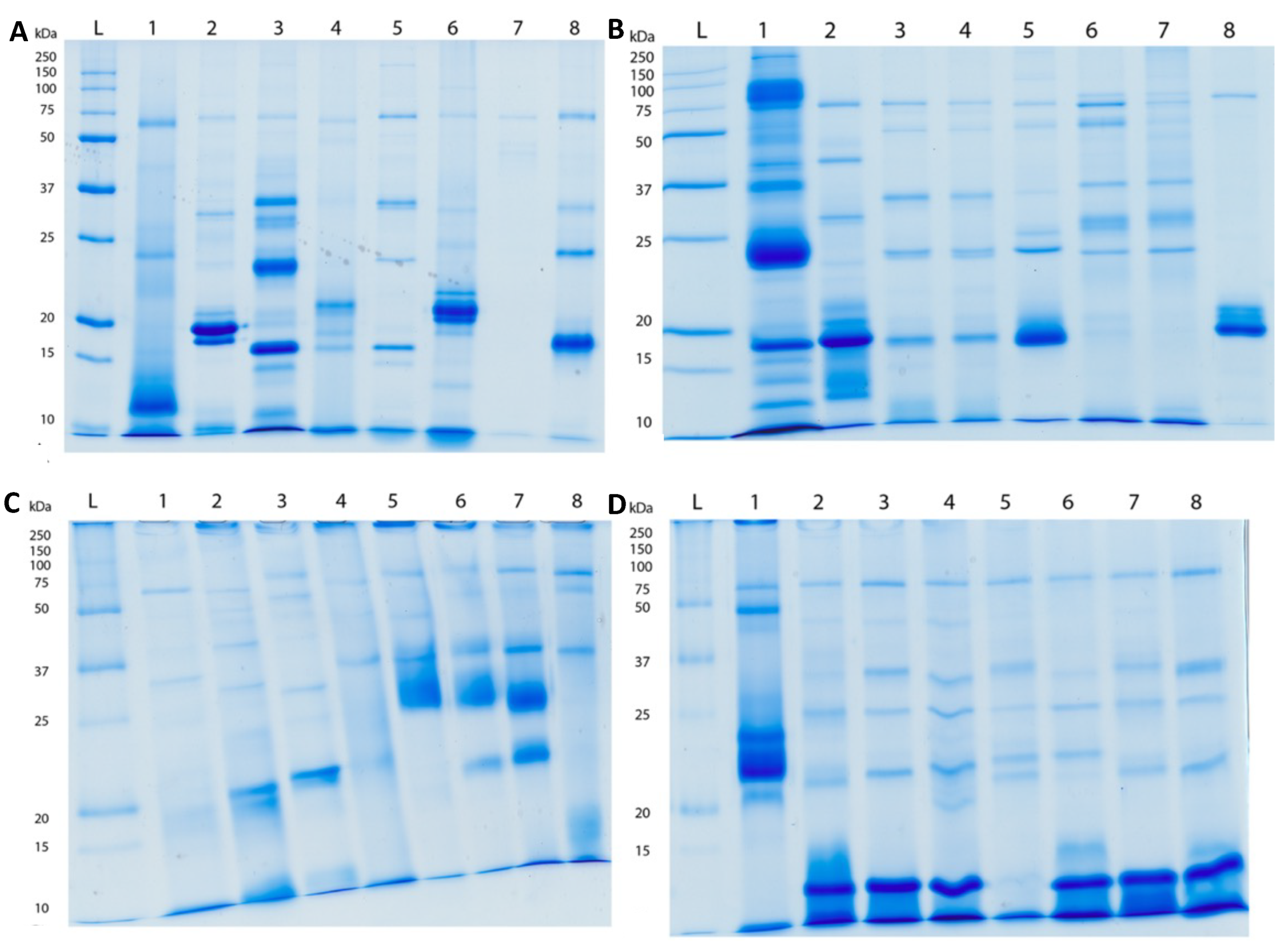
2.2. Proteomics
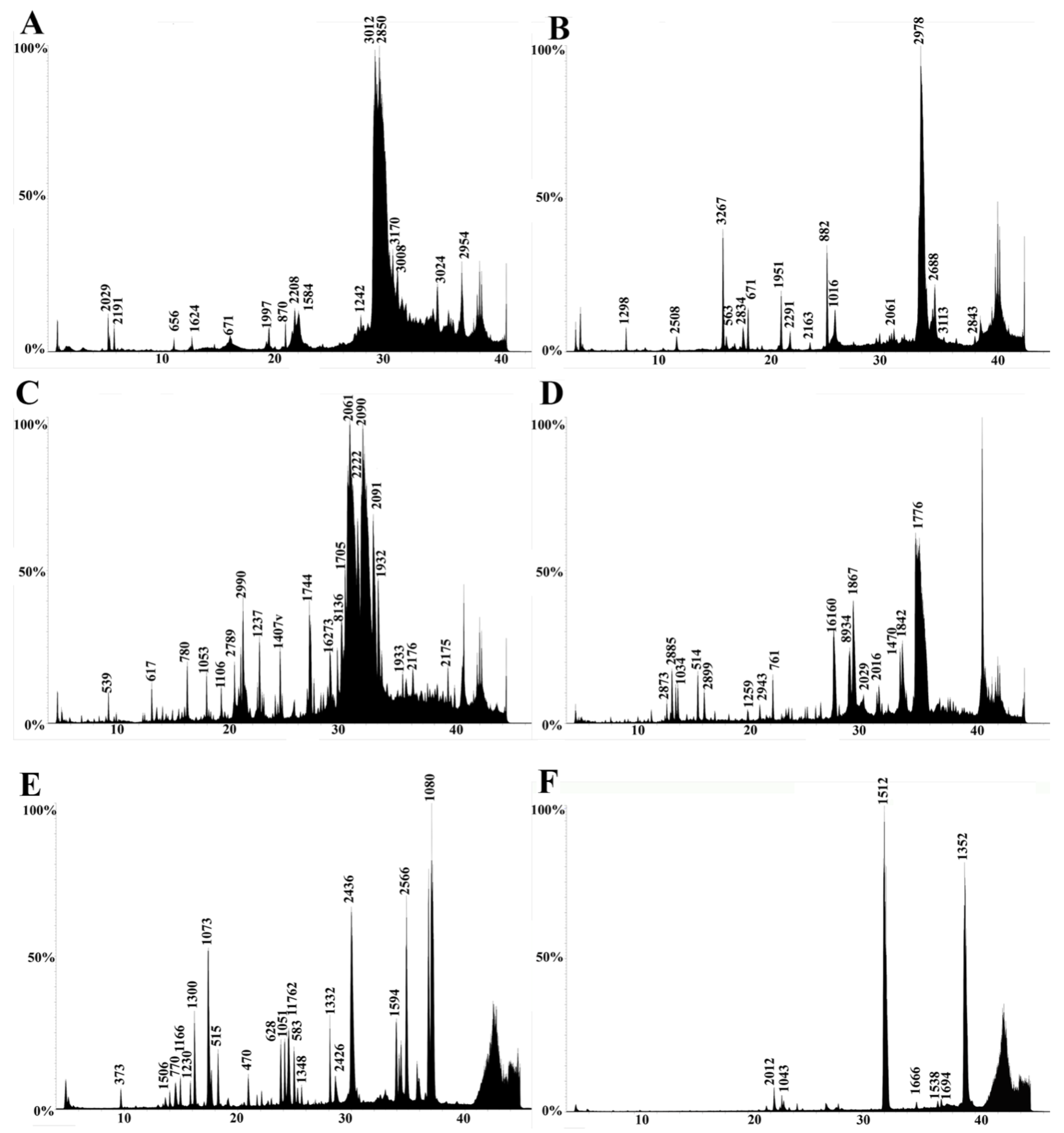
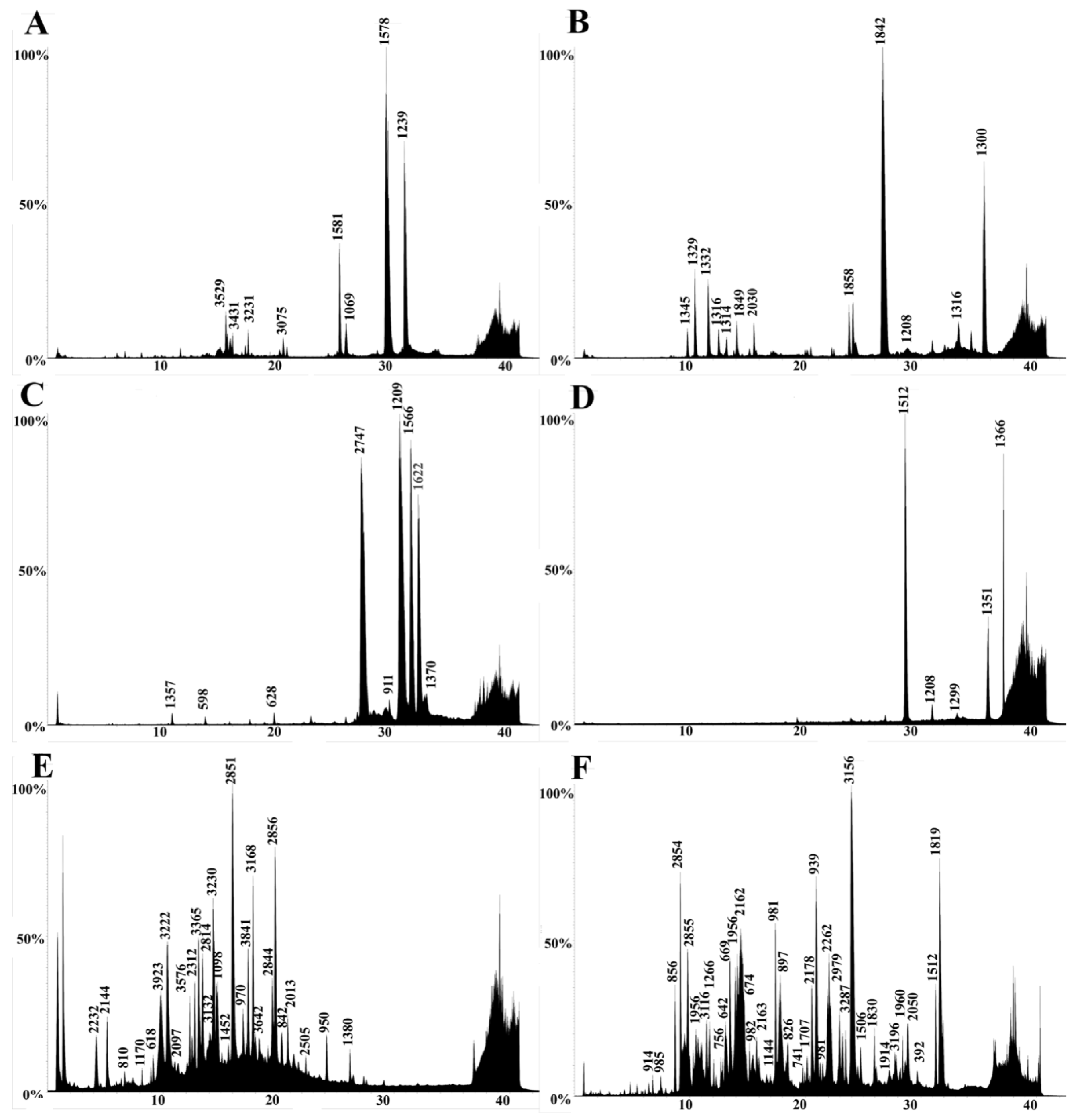
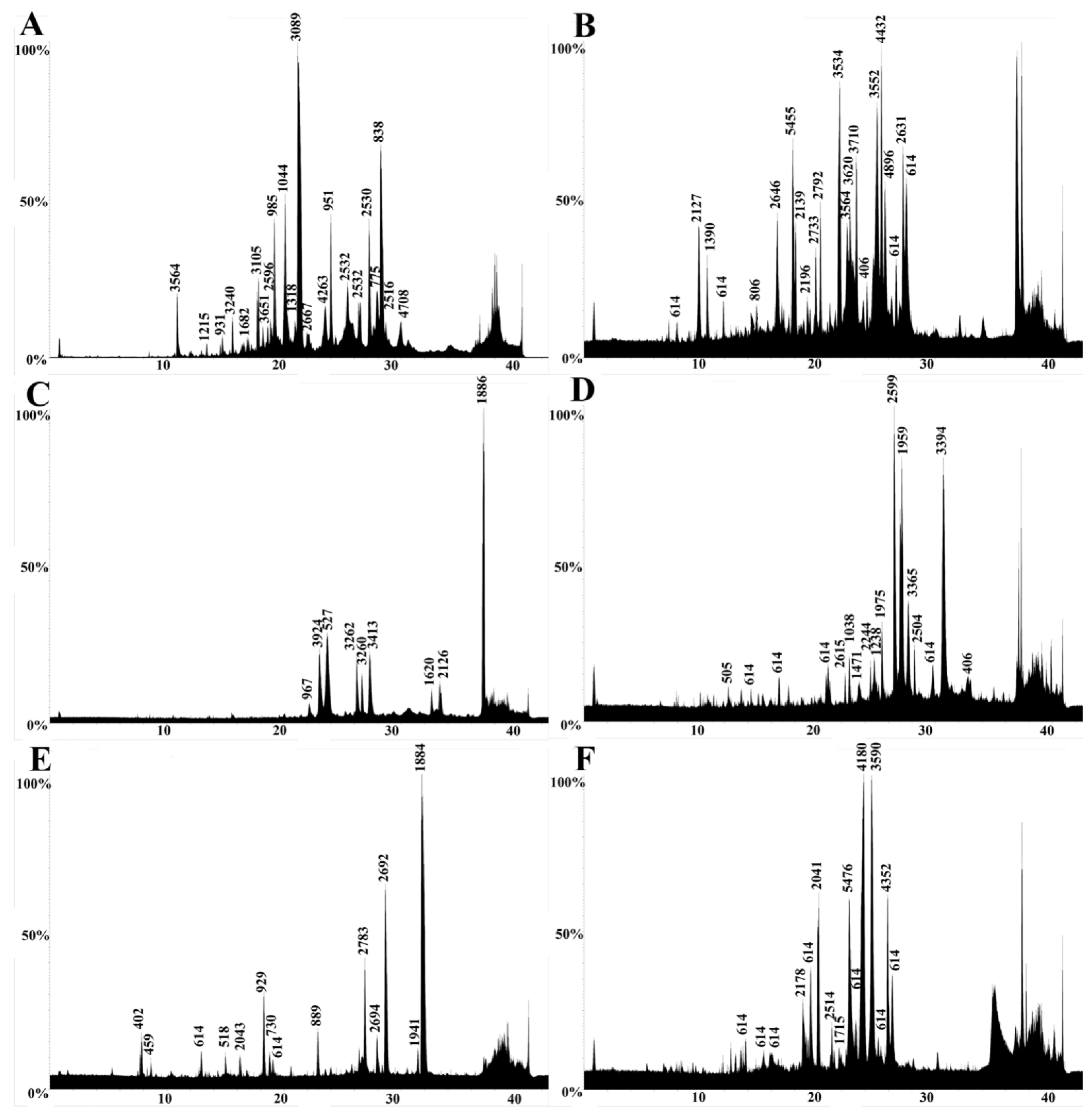
2.3. LC-MS
2.4. LC-MS/MS
2.5. Enzymatic Assays
2.6. Cytotoxicity Assays
3. Discussion
4. Conclusions
5. Materials and Methods
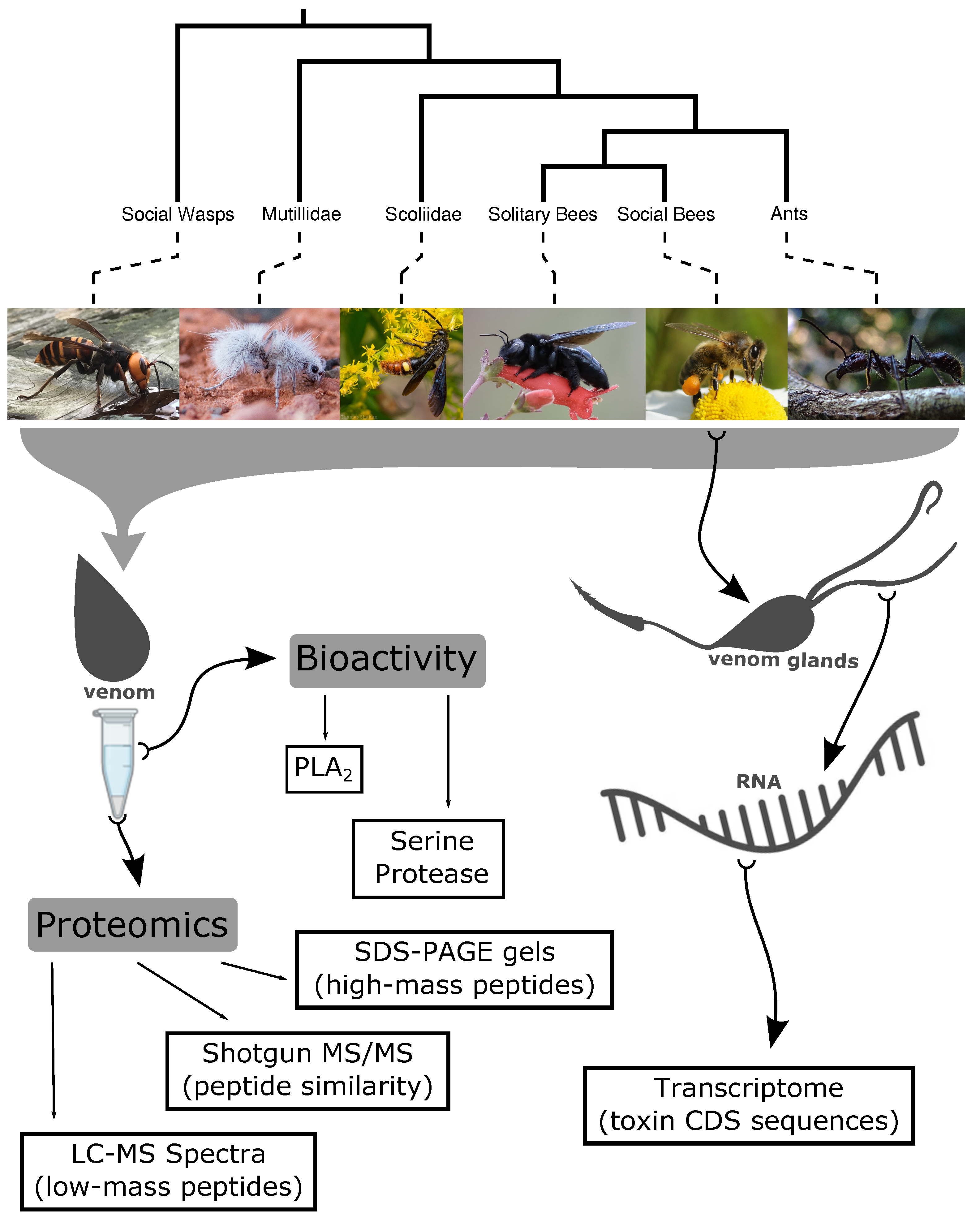
5.1. Taxonomic Selection
5.2. Venom Collection
5.3. Proteomics
5.3.1. SDS-PAGE
5.3.2. Liquid Chromatography–Mass Spectrometry (LC-MS)
5.3.3. Tandem Mass Spectrometry (LC-MS/MS)
5.4. Transcriptomics
5.4.1. RNA Extraction and Library Preparation
5.4.2. Sequence Data Pre-Processing and Transcriptome Assembly
5.4.3. Transcriptome Annotation
5.5. Bioactivity Activity Testing
5.5.1. Enzymatic Activity Studies
5.5.2. Cytotoxicity Studies
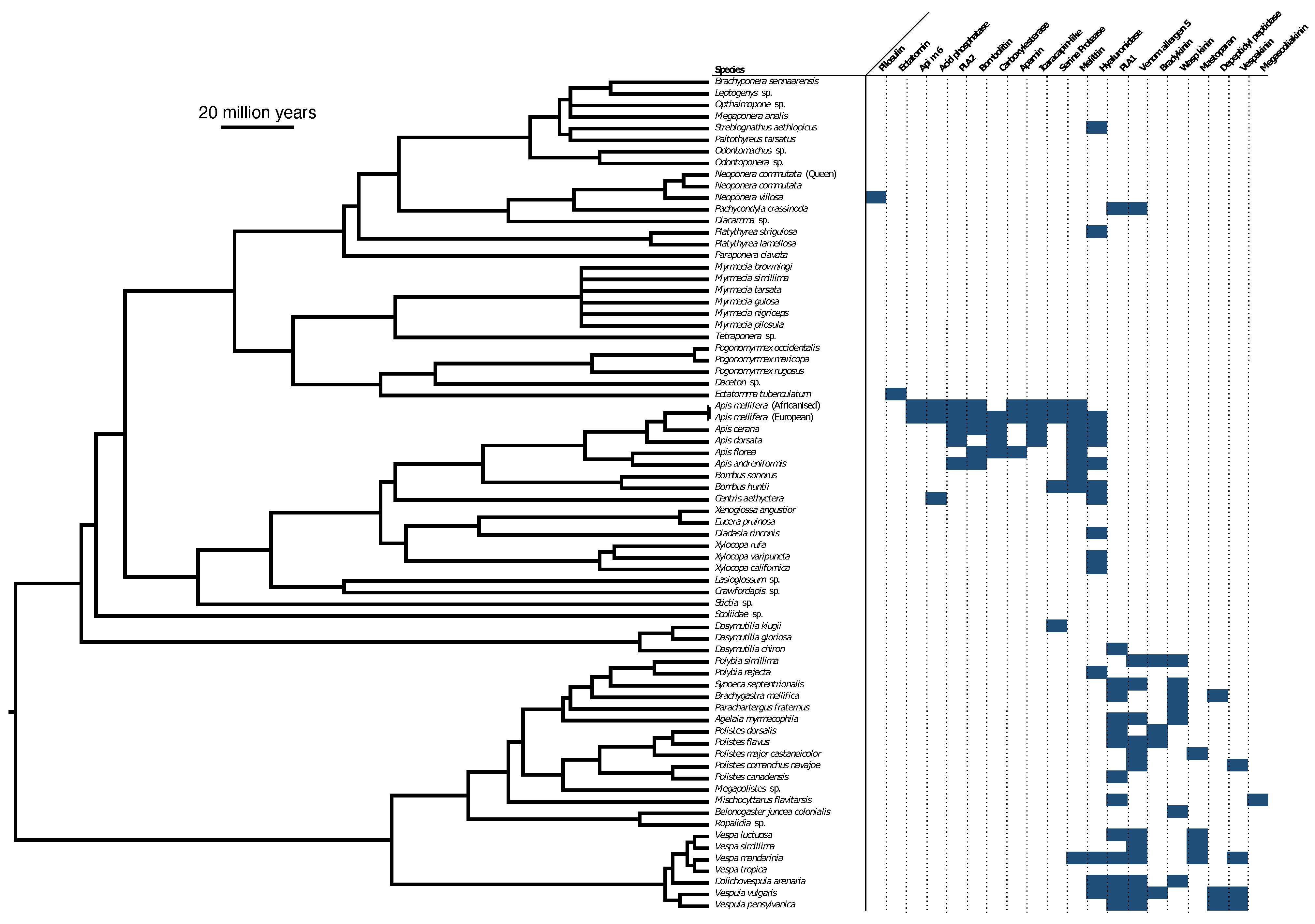
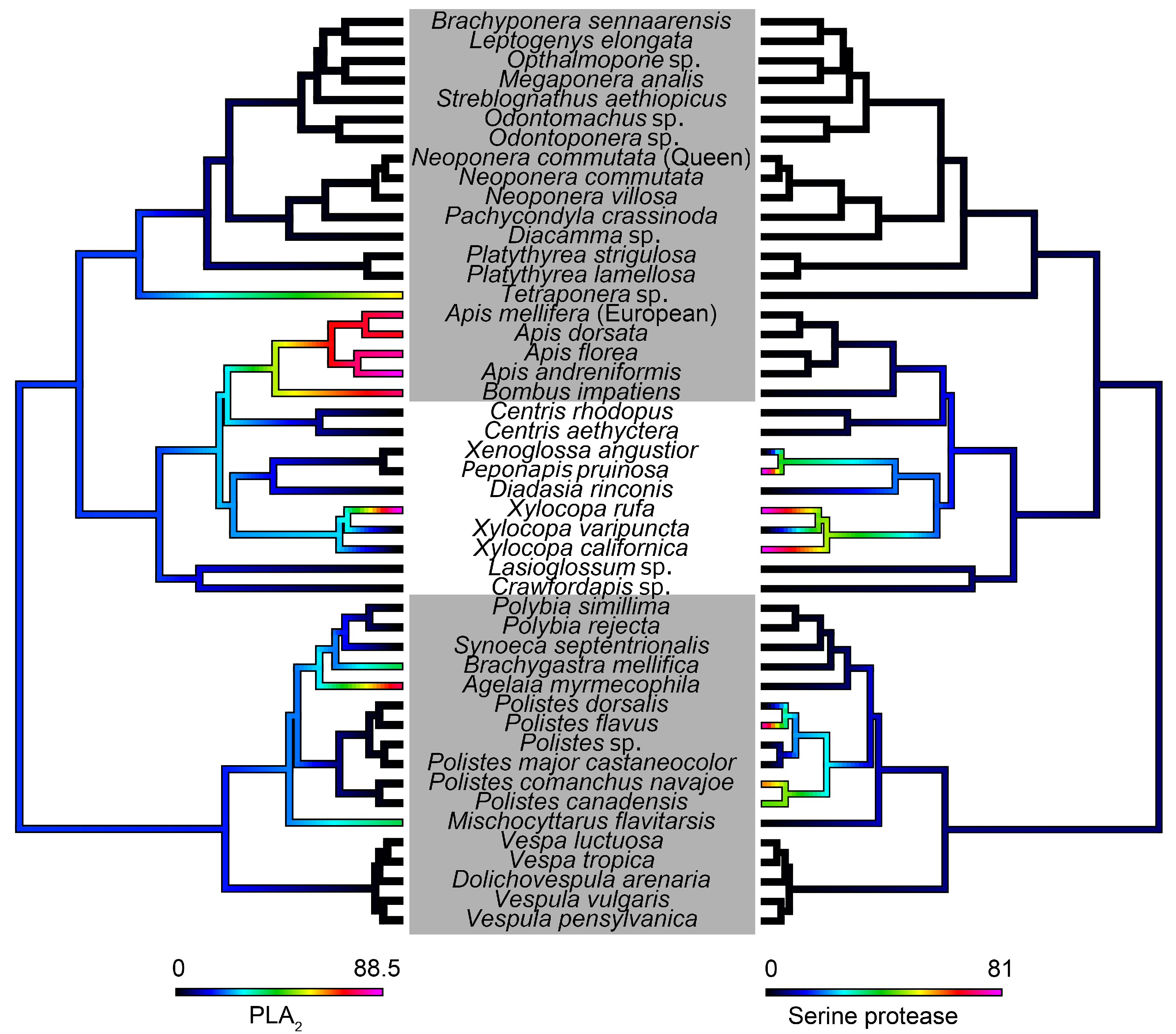
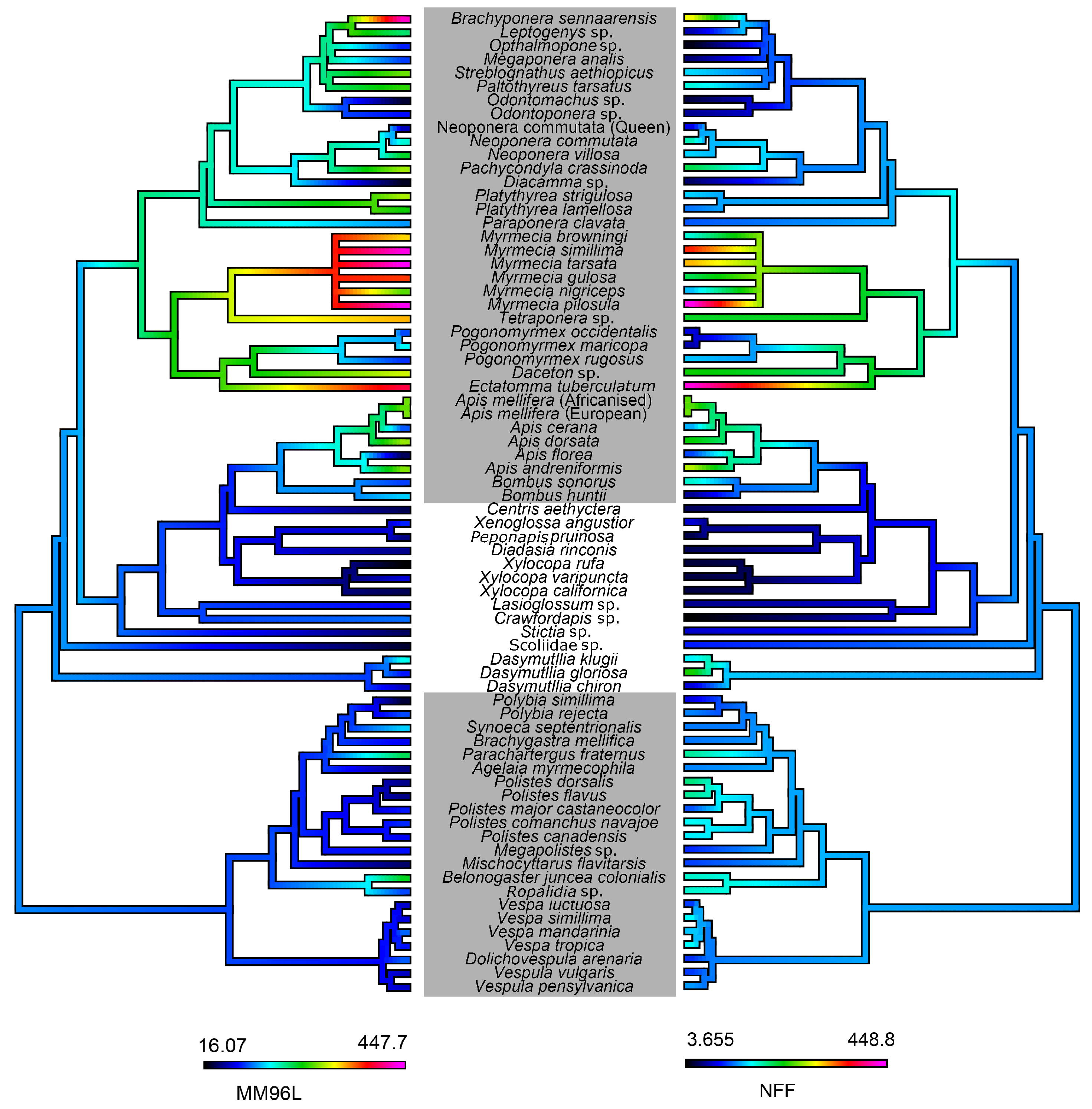
5.6. Ancestral State Reconstruction
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PLA1 | Phospholipase A1 |
| PLA2 | Phospholipase A2 |
| 1D-SDS PAGE | One-dimensional sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| LC-MS | Liquid chromatography-mass spectrometry |
| MS/MS | Tandem mass spectrometry |
| PGLS | Phylogenetic generalized least squares |
References
- Piek, T. Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Academic Press: London, UK, 1986. [Google Scholar]
- Gaston, K.J. The magnitude of global insect species richness. Conserv. Biol. 1991, 5, 283–296. [Google Scholar] [CrossRef]
- Stork, N.E. How many species of insects and other terrestrial arthropods are there on Earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, A.A.; Bagley, R.K.; Beer, M.A.; Hippee, A.C.; Widmayer, H.A. Quantifying the unquantifiable: Why Hymenoptera, not Coleoptera, is the most speciose animal order. BMC Ecol. 2018, 18, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, A.A.; Robinson, S.D.; Yeates, D.K.; Jin, J.; Baumann, K.; Dobson, J.; Fry, B.G.; King, G.F. Entomo-venomics: The evolution, biology and biochemistry of insect venoms. Toxicon 2018, 154, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, L.W.; Korzukhin, M.D.; Porter, S.D. Predicted range expansion of the invasive fire ant, Solenopsis invicta, in the eastern United States based on the VEMAP global warming scenario. Divers. Distrib. 2005, 11, 199–204. [Google Scholar] [CrossRef]
- Kenis, M.; Auger-Rozenberg, M.A.; Roques, A.; Timms, L.; Péré, C.; Cock, M.J.; Settele, J.; Augustin, S.; Lopez-Vaamonde, C. Ecological effects of invasive alien insects. Biol. Invasions 2009, 11, 21–45. [Google Scholar] [CrossRef]
- Dos Santos Pinto, J.; Fox, E.; Saidemberg, D.; Santos, L.; Silva Menegasso, A.; Costa-Manso, E.; Machado, E.; Bueno, O.; Palma, M. Proteomic View of the Venom from the Fire Ant Solenopsis invicta Buren. J. Proteome Res. 2012, 11, 4643–4653. [Google Scholar] [CrossRef]
- Lei, W.; Xu, Y.-J.; Ling, Z.; Lu, Y.-Y. Impact of the red imported fire ant Solenopsis invicta Buren on biodiversity in South China: A review. J. Integr. Agric. 2019, 18, 788–796. [Google Scholar]
- Orr, D.B. Scelionid wasps as biological control agents: A review. Fla. Entomol. 1988, 71, 506–528. [Google Scholar] [CrossRef]
- Southon, R.J.; Fernandes, O.A.; Nascimento, F.S.; Sumner, S. Social wasps are effective biocontrol agents of key lepidopteran crop pests. Proc. R. Soc. B 2019, 286, 20191676. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.Z.; Liu, Y.Q.; Min, S.H.I.; Huang, J.H.; Chen, X.X. Parasitoid wasps as effective biological control agents. J. Integr. Agric. 2019, 18, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, J.A. Biotic pollination mechanisms in the Australian flora—A review. N. Z. J. Bot. 1979, 17, 467–508. [Google Scholar] [CrossRef]
- Hein, L. The Economic Value of the Pollination Service, a Review Across Scales. Open Ecol. J. 2009, 2, 74–82. [Google Scholar] [CrossRef]
- Schumacher, M.J.; Egen, N.B. Significance of Africanized Bees for Public Health: A Review. Arch. Intern. Med. 1995, 155, 2038–2043. [Google Scholar] [CrossRef]
- Vetter, R.S.; Visscher, P.K.; Camazine, S. Mass envenomations by honey bees and wasps. West. J. Med. 1999, 170, 223–227. [Google Scholar]
- Schmidt, J.O. Clinical consequences of toxic envenomations by Hymenoptera. Toxicon 2018, 150, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Cerni, F.A.; Oliveira, I.S.; Jenkins, T.P.; Argemí, L.; Sørensen, C.V.; Ahmadi, S.; Barbosa, J.E.; Laustsen, A.H. Bee Updated: Current Knowledge on Bee Venom and Bee Envenoming Therapy. Front. Immunol. 2019, 10, 2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, W.; Oldroyd, B.; Beekman, M.; Ratnieks, F. Ancestral Monogamy Shows Kin Selection Is Key to the Evolution of Eusociality. Science 2008, 320, 1213–1216. [Google Scholar] [CrossRef]
- Bourke, A. The validity and value of inclusive fitness theory. Proceedings. Biol. Sci. 2011, 278, 3313–3320. [Google Scholar] [CrossRef]
- Hamilton, W. The genetical evolution of social behaviour. I. J. Theor. Biol. 1964, 7, 1–16. [Google Scholar] [CrossRef]
- Strachecka, A.; Chobotow, J.; Paleolog, J.; Łoś, A.; Schulz, M.; Teper, D.; Kucharczyk, H.; Grzybek, M. Insights into the biochemical defence and methylation of the solitary bee Osmia rufa L: A foundation for examining eusociality development. PLoS ONE 2017, 12, e0176539. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, J.B. Phylogeny and evolution of host-parasitoid interactions in Hymenoptera. Annu. Rev. Entomol. 1998, 43, 129–151. [Google Scholar] [CrossRef]
- Heraty, J. Parasitoid biodiversity and insect pest management. In Insect Biodiversity: Science and Society; John Wiley & Sons, Ltd.: West Sussex, UK, 2009; pp. 445–462. [Google Scholar]
- Poirié, M.; Carton, Y.; Dubuffet, A. Virulence strategies in parasitoid Hymenoptera as an example of adaptive diversity. Comptes Rendus Biol. 2009, 332, 311–320. [Google Scholar] [CrossRef]
- Dashevsky, D.; Rodriguez, J. A short review of the venoms and toxins of spider wasps (Hymenoptera: Pompilidae). Toxins 2021, 13, 744. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.O. Hymenopteran venoms: Striving toward the ultimate defense against vertebrates. In Insect Defense: Adaptations and Strategies of Prey and Predators; Evans, D.L., Schmidt, J.O., Eds.; SUNY Press: Albany, NY, USA, 1990; pp. 387–419. [Google Scholar]
- De Lima, P.R.; Brochetto-Braga, M.R. Hymenoptera venom review focusing on Apis mellifera. J. Venom. Anim. Toxins Incl. Trop. Dis. 2003, 9, 149–162. [Google Scholar]
- Bland, R.G.; Jaques, H.E. How to Know the Insects; Waveland Press: Long Grove, IL, USA, 2010. [Google Scholar]
- Schmidt, J.O. The Sting of the Wild; Johns Hopkins University Press: Baltimore, MD, USA, 2016. [Google Scholar]
- Lee, S.H.; Baek, J.H.; Yoon, K.A. Differential Properties of Venom Peptides and Proteins in Solitary vs. Social Hunting Wasps. Toxins 2016, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, D.; Jacobson, R. Allergens in Hymenoptera venom. XXVII: Bumblebee venom allergy and allergens. J. Allergy Clin. Immunol. 1996, 97, 812–821. [Google Scholar] [CrossRef]
- Hoffman, D. Hymenoptera Venom Allergens. Clin. Rev. Allergy Immunol. 2006, 30, 109–128. [Google Scholar] [CrossRef]
- Son, D.; Lee, J.; Lee, Y.; Song, H.; Lee, C.; Hong, J. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 2007, 115, 246–270. [Google Scholar] [CrossRef] [PubMed]
- Winningham, K.; Fitch, C.; Schmidt, M.; Hoffman, D. Hymenoptera venom protease allergens. J. Allergy Clin. Immunol. 2004, 114, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Arbuckle, K. Evolutionary Context of Venom in Animals; Springer: Dordrecht, The Netherlands, 2017. [Google Scholar]
- dos Santos-Pinto, J.R.A.; Perez-Riverol, A.; Lasa, A.M.; Palma, M.S. Diversity of peptidic and proteinaceous toxins from social Hymenoptera venoms. Toxicon 2018, 148, 172–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batra, S. Solitary Bees. Sci. Am. 1984, 250, 120–127. [Google Scholar] [CrossRef]
- Gauld, I.; Bolton, B. The Aculeate Apocritans; British Museum (Natural History): London, UK, 1988. [Google Scholar]
- Gauld, I.; Bolton, B. The Biology of the Hymenoptera; British Museum (Natural History): London, UK, 1988. [Google Scholar]
- Aili, S.; Touchard, A.; Escoubas, P.; Padula, M.; Orivel, J.; Dejean, A.; Nicholson, G. Diversity of peptide toxins from stinging ant venoms. Toxicon 2014, 92, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Touchard, A.; Aili, S.; Fox, E.; Escoubas, P.; Orivel, J.; Nicholson, G.; Dejean, A. The Biochemical Toxin Arsenal from Ant Venoms. Toxins 2016, 8, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biló, B.M.; Rueff, F.; Mosbech, H.; Bonifazi, F.; Oude-Elberink, J.N.G.; EAACI Interest Group on Insect Venom Hypersensitivity. Diagnosis of Hymenoptera venom allergy. Allergy 2005, 60, 1339–1349. [Google Scholar] [CrossRef]
- Alfaya Arias, T.; Soriano Gómis, V.; Soto Mera, T.; Vega Castro, A.; Vega Gutiérrez, J.; Alonso Llamazares, A.; Antolín Amérigo, D.; Carballada Gonzalez, F.; Dominguez Noche, C.; Gutierrez Fernandez, D.; et al. Key Issues in Hymenoptera Venom Allergy: An Update. J. Investig. Allergol. Clin. Immunol. 2017, 27, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Nešuta, O.; Hexnerová, R.; Buděšínský, M.; Slaninová, J.; Bednárová, L.; Hadravová, R.; Straka, J.; Veverka, V.; Čeřovský, V. Antimicrobial Peptide from the Wild Bee Hylaeus signatus Venom and Its Analogues: Structure–Activity Study and Synergistic Effect with Antibiotics. J. Nat. Prod. 2016, 79, 1073–1083. [Google Scholar] [CrossRef]
- Čujová, S.; Slaninová, J.; Monincová, L.; Fučík, V.; Bednárová, L.; Štokrová, J.; Hovorka, O.; Voburka, Z.; Straka, J.; Čeřovský, V. Panurgines, novel antimicrobial peptides from the venom of communal bee Panurgus calcaratus (Hymenoptera: Andrenidae. Amino Acids 2013, 45, 143–157. [Google Scholar] [CrossRef]
- Čujová, S.; Bednárová, L.; Slaninová, J.; Straka, J.; Čeřovský, V. Interaction of a novel antimicrobial peptide isolated from the venom of solitary bee Colletes daviesanus with phospholipid vesicles and Escherichia coli cells. J. Pept. Sci. 2014, 20, 885–895. [Google Scholar] [CrossRef]
- Monincová, L.; Buděšínský, M.; Slaninová, J.; Hovorka, O.; Cvačka, J.; Voburka, Z.; Fučík, V.; Borovičková, L.; Bednárová, L.; Straka, J.; et al. Novel antimicrobial peptides from the venom of the eusocial bee Halictus sexcinctus (Hymenoptera: Halictidae) and their analogs. Amino Acids 2010, 39, 763–775. [Google Scholar] [CrossRef]
- Monincová, L.; Veverka, V.; Slaninová, J.; Buděšínský, M.; Fučík, V.; Bednárová, L.; Straka, J.; Čeřovský, V. Structure-activity study of macropin, a novel antimicrobial peptide from the venom of solitary bee Macropis fulvipes (Hymenoptera: Melittidae). J. Pept. Sci. 2014, 20, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Stöcklin, R.; Favreau, P.; Thai, R.; Pflugfelder, J.; Bulet, P.; Mebs, D. Structural identification by mass spectrometry of a novel antimicrobial peptide from the venom of the solitary bee Osmia rufa (Hymenoptera: Megachilidae). Toxicon 2010, 55, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Diniz-Sousa, R.; Kayano, A.; Caldeira, C.; Simões-Silva, R.; Monteiro, M.; Moreira-Dill, L.; Grabner, F.; Calderon, L.; Zuliani, J.; Stábeli, R.; et al. Biochemical characterization of a phospholipase A2 homologue from the venom of the social wasp Polybia occidentalis. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos Cabrera, M.; Souza, B.; Fontana, R.; Konno, K.; Palma, M.; Azevedo, W.; Ruggiero Neto, J. Conformation and lytic activity of eumenine mastoparan: A new antimicrobial peptide from wasp venom. J. Pept. Res. 2004, 64, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Palma, M. Peptides as toxins/defensins. Amino Acids 2011, 40, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Burzyńska, M.; Piasecka-Kwiatkowska, D. A Review of Honeybee Venom Allergens and Allergenicity. Int. J. Mol. Sci. 2021, 22, 8371. [Google Scholar] [CrossRef]
- Moreau, S.; Asgari, S. Venom Proteins from Parasitoid Wasps and Their Biological Functions. Toxins 2015, 7, 2385–2412. [Google Scholar] [CrossRef] [Green Version]
- Donovan, G.R.; Street, M.D.; Baldo, B.A.; Alewood, D.; Alewood, P.; Sutherland, S. Identification of an IgE-binding determinant of the major allergen Myr pI from the venom of the Australian jumper ant Myrmecia pilosula. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1994, 1204, 48–52. [Google Scholar] [CrossRef]
- Donovan, G.R.; Street, M.D.; Tetaz, T.; Smith, A.I.; Alewood, D.; Alewood, P.; Sutherland, S.K.; Baldo, B.A. Expression of jumper ant (Myrmecia pilosula) venom allergens: Post-translational processing of allergen gene products. IUBMB Life 1996, 39, 877–885. [Google Scholar] [CrossRef]
- Wu, Q.X.; King, M.; Donovan, G.; Alewood, D.; Alewood, P.; Sawyer, W.; Baldo, B. Cytotoxicity of pilosulin 1, a peptide from the venom of the jumper ant Myrmecia pilosula. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1998, 1425, 74–80. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Fontana, R.; Lorenzi, C.C.B.; Naoki, H.; Itagaki, Y.; Miwa, A.; Kawai, N.; Nakata, Y.; Yasuhara, T.; et al. Anoplin, a novel antimicrobial peptide from the venom of the solitary wasp Anoplius samariensis. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 2001, 1550, 70–80. [Google Scholar] [CrossRef]
- Davies, N.W.; Wiese, M.D.; Brown, S.G.A. Characterisation of major peptides in ‘jack jumper’ ant venom by mass spectrometry. Toxicon 2004, 43, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos Cabrera, M.P.; Arcisio-Miranda, M.; Broggio Costa, S.T.; Konno, K.; Ruggiero, J.R.; Procopio, J.; Ruggiero Neto, J. Study of the mechanism of action of anoplin, a helical antimicrobial decapeptide with ion channel-like activity, and the role of the amidated C-terminus. J. Pept. Sci. 2008, 14, 661–669. [Google Scholar] [CrossRef]
- Robinson, S.; Mueller, A.; Clayton, D.; Starobova, H.; Hamilton, B.; Payne, R.; Vetter, I.; King, G.; Undheim, E.B. A comprehensive portrait of the venom of the giant red bull ant, Myrmecia gulosa, reveals a hyperdiverse hymenopteran toxin gene family. Sci. Adv. 2018, 4, eaau4640. [Google Scholar] [CrossRef] [Green Version]
- Moon, D.O.; Park, S.Y.; Heo, M.S.; Kim, K.C.; Park, C.; Ko, W.; Choi, Y.; Kim, G.Y. Key regulators in bee venom-induced apoptosis are Bcl-2 and caspase-3 in human leukemic U937 cells through downregulation of ERK and Akt. Int. Immunopharmacol. 2006, 6, 1796–1807. [Google Scholar] [CrossRef] [PubMed]
- Shiassi Arani, F.; Karimzadeh, L.; Ghafoori, S.M.; Nabiuni, M. Antimutagenic and Synergistic Cytotoxic Effect of Cisplatin and Honey Bee Venom on 4T1 Invasive Mammary Carcinoma Cell Line. Adv. Pharmacol. Sci. 2019, 2019, e7581318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajski, G.; Čimbora Zovko, T.; Rak, S.; Rožman, M.; Osmak, M.; Garaj-Vrhovac, V. Combined antitumor effects of bee venom and cisplatin on human cervical and laryngeal carcinoma cells and their drug resistant sublines. J. Appl. Toxicol. 2014, 34, 1332–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.W.; Chaturvedi, P.; Chun, S.; Lee, Y.; Ahn, W. Honeybee venom possesses anticancer and antiviral effects by differential inhibition of HPV E6 and E7 expression on cervical cancer cell line. Oncol. Rep. 2015, 33, 1675–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Kang, S.; Kim, B.; Kim, Y.; Woo, H.; Chung, H. Cytotoxicity of honeybee (Apis mellifera) venom in normal human lymphocytes and HL-60 cells. Chem.-Biol. Interact. 2007, 169, 189–197. [Google Scholar] [CrossRef]
- Hoshina, M.; Santos, L.; Palma, M.; Marin-Morales, M. Cytotoxic, genotoxic/antigenotoxic and mutagenic/antimutagenic effects of the venom of the wasp Polybia paulista. Toxicon 2013, 72, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Tamimi, J.; Semlali, A.; Hassan, I.; Ebaid, H.; Alhazza, I.; Mehdi, S.; Al-Khalifa, M.; Alanazi, M. Samsum Ant Venom Exerts Anticancer Activity Through Immunomodulation In Vitro and In Vivo. Cancer Biother. Radiopharm. 2018, 33, 65–73. [Google Scholar] [CrossRef]
- Leite, N.; Aufderhorst-Roberts, A.; Palma, M.; Connell, S.; Ruggiero Neto, J.; Beales, P. PE and PS Lipids Synergistically Enhance Membrane Poration by a Peptide with Anticancer Properties. Biophys. J. 2015, 109, 936–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dkhil, M.; Abdel-Baki, A.; Al-Quraishi, S.; Al-Khalifa, M. Anti-inflammatory activity of the venom from samsum ants Pachycondyla sennaarensis. Afr. J. Pharm. Pharmacol. 2010, 4, 115–118. [Google Scholar]
- Danneels, E.L.; Gerlo, S.; Heyninck, K.; Craenenbroeck, K.V.; Bosscher, K.D.; Haegeman, G.; Graaf, D.C.d. How the Venom from the Ectoparasitoid Wasp Nasonia vitripennis Exhibits Anti-Inflammatory Properties on Mammalian Cell Lines. PLoS ONE 2014, 9, e96825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.; Bae, H. Anti-Inflammatory Applications of Melittin, a Major Component of Bee Venom: Detailed Mechanism of Action and Adverse Effects. Molecules 2016, 21, 616. [Google Scholar] [CrossRef] [Green Version]
- Kocyigit, A.; Guler, E.M.; Kaleli, S. Anti-inflammatory and antioxidative properties of honey bee venom on Freund’s Complete Adjuvant-induced arthritis model in rats. Toxicon 2019, 161, 4–11. [Google Scholar] [CrossRef]
- Khalil, A.; Elesawy, B.H.; Ali, T.M.; Ahmed, O.M. Bee Venom: From Venom to Drug. Molecules 2021, 26, 4941. [Google Scholar] [CrossRef]
- Yun, H.S.; Oh, J.; Lim, J.S.; Kim, H.J.; Kim, J.S. Anti-Inflammatory Effect of Wasp Venom in BV-2 Microglial Cells in Comparison with Bee Venom. Insects 2021, 12, 297. [Google Scholar] [CrossRef]
- Koludarov, I.; Velasque, M.; Timm, T.; Lochnit, G.; Heinzinger, M.; Vilcinskas, A.; Gloag, R.; Harpur, B.A.; Podsiadlowski, L.; Rost, B.; et al. Bee core venom genes predominantly originated before aculeate stingers evolved. bioRxiv 2022. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annesley, T. Ion suppression in mass spectrometry. Clin. Chem. 2003, 49, 1041–1044. [Google Scholar] [CrossRef] [Green Version]
- Cardinal, S.; Danforth, B.N. Bees diversified in the age of eudicots. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122686. [Google Scholar] [CrossRef] [Green Version]
- Perrard, A.; Pickett, K.; Villemant, C.; Kojima, J.i.; Carpenter, J. Phylogeny of hornets: A total evidence approach (Hymenoptera, Vespidae, Vespinae, Vespa). J. Hymenopt. Res. 2013, 32, 1–15. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]
- Peters, R.S.; Krogmann, L.; Mayer, C.; Donath, A.; Gunkel, S.; Meusemann, K.; Kozlov, A.; Podsiadlowski, L.; Petersen, M.; Lanfear, R.; et al. Evolutionary history of the Hymenoptera. Curr. Biol. 2017, 27, 1013–1018. [Google Scholar] [CrossRef] [Green Version]
- Piekarski, P.K.; Carpenter, J.M.; Lemmon, A.R.; Moriarty Lemmon, E.; Sharanowski, B.J. Phylogenomic Evidence Overturns Current Conceptions of Social Evolution in Wasps (Vespidae). Mol. Biol. Evol. 2018, 35, 2097–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossert, S.; Murray, E.A.; Almeida, E.A.B.; Brady, S.G.; Blaimer, B.B.; Danforth, B.N. Combining transcriptomes and ultraconserved elements to illuminate the phylogeny of Apidae. Mol. Phylogenetics Evol. 2019, 130, 121–131. [Google Scholar] [CrossRef]
- Borowiec, M.L.; Moreau, C.S.; Rabeling, C. Ants: Phylogeny and Classification. In Encyclopedia of Social Insects; Starr, C.K., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–18. [Google Scholar] [CrossRef]
- Menezes, R.S.T.; Lloyd, M.W.; Brady, S.G. Phylogenomics indicates Amazonia as the major source of Neotropical swarm-founding social wasp diversity. Proc. R. Soc. B Biol. Sci. 2020, 287, 20200480. [Google Scholar] [CrossRef] [PubMed]
- Somavilla, A.; Santos, B.F.; Carpenter, J.M.; Andena, S.R.; Oliveira, M.L. Total-Evidence Phylogeny of the New World Polistes Lepeletier, 1836, Paper Wasps (Vespidae, Polistinae, Polistini). Am. Mus. Novit. 2021, 2021, 1–42. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.70. 2021. Available online: https://www.mesquiteproject.org (accessed on 24 February 2022).
- Yoon, K.; Kim, K.; Nguyen, P.; Seo, J.; Park, Y.; Kim, K.G.; Seo, H.Y.; Koh, Y.; Lee, S. Comparative functional venomics of social hornets Vespa crabro and Vespa analis. J. -Asia-Pac. Entomol. 2015, 18, 815–823. [Google Scholar] [CrossRef]
- Kazuma, K.; Masuko, K.; Konno, K.; Inagaki, H. Combined Venom Gland Transcriptomic and Venom Peptidomic Analysis of the Predatory Ant Odontomachus monticola. Toxins 2017, 9, 323. [Google Scholar] [CrossRef] [Green Version]
- Özbek, R.; Wielsch, N.; Vogel, H.; Lochnit, G.; Foerster, F.; Vilcinskas, A.; von Reumont, B.M. Proteo-Transcriptomic Characterization of the Venom from the Endoparasitoid Wasp Pimpla turionellae with Aspects on Its Biology and Evolution. Toxins 2019, 11, 721. [Google Scholar] [CrossRef] [Green Version]
- Alberto-Silva, C.; Vieira Portaro, F.C.; Kodama, R.T.; Pantaleão, H.Q.; Inagaki, H.; Nihei, K.i.; Konno, K. Comprehensive Analysis and Biological Characterization of Venom Components from Solitary Scoliid Wasp Campsomeriella annulata annulata. Toxins 2021, 13, 885. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Ren, L.; Wang, M.; Wang, Z.; Fu, N.; Wang, H.; Wang, X.; Ao, T.; Du, W.; Zheng, Z.; et al. Proteo-Transcriptomic Characterization of Sirex nitobei (Hymenoptera: Siricidae) Venom. Toxins 2021, 13, 562. [Google Scholar] [CrossRef]
- Gatti, J.L.; Belghazi, M.; Legeai, F.; Ravallec, M.; Frayssinet, M.; Robin, S.; Aboubakar-Souna, D.; Srinivasan, R.; Tamò, M.; Poirié, M.; et al. Proteo-Trancriptomic Analyses Reveal a Large Expansion of Metalloprotease-Like Proteins in Atypical Venom Vesicles of the Wasp Meteorus pulchricornis (Braconidae). Toxins 2021, 13, 502. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.; Walker, A.A.; Nguyen, S.H.; Jin, A.H.; Deuis, J.R.; Vetter, I.; King, G.F.; Schmidt, J.O.; Robinson, S.D. Venom chemistry underlying the painful stings of velvet ants (Hymenoptera: Mutillidae). Cell. Mol. Life Sci. 2021, 78, 5163–5177. [Google Scholar] [CrossRef] [PubMed]
- Pinto, C.P.G.; Walker, A.A.; Robinson, S.D.; Chin, Y.K.Y.; King, G.F.; Rossi, G.D. Venom composition of the endoparasitoid wasp Cotesia flavipes (Hymenoptera: Braconidae) and functional characterization of a major venom peptide. Toxicon 2021, 202, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Quicke, D.L.J.; Butcher, B.A. Review of Venoms of Non-Polydnavirus Carrying Ichneumonoid Wasps. Biology 2021, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Scieuzo, C.; Salvia, R.; Franco, A.; Pezzi, M.; Cozzolino, F.; Chicca, M.; Scapoli, C.; Vogel, H.; Monti, M.; Ferracini, C.; et al. An integrated transcriptomic and proteomic approach to identify the main Torymus sinensis venom components. Sci. Rep. 2021, 11, 5032. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ye, X.; Dang, C.; Cao, Y.; Hong, R.; Sun, Y.H.; Xiao, S.; Mei, Y.; Xu, L.; Fang, Q.; et al. Genome of the pincer wasp Gonatopus flavifemur reveals unique venom evolution and a dual adaptation to parasitism and predation. BMC Biol. 2021, 19, 145. [Google Scholar] [CrossRef]
- Barassé, V.; Téné, N.; Klopp, C.; Paquet, F.; Tysklind, N.; Troispoux, V.; Lalägue, H.; Orivel, J.; Lefranc, B.; Leprince, J.; et al. Venomics survey of six myrmicine ants provides insights into the molecular and structural diversity of their peptide toxins. Insect Biochem. Mol. Biol. 2022, 151, 103876. [Google Scholar] [CrossRef] [PubMed]
- Hurka, S.; Brinkrolf, K.; Özbek, R.; Förster, F.; Billion, A.; Heep, J.; Timm, T.; Lochnit, G.; Vilcinskas, A.; Lüddecke, T. Venomics of the Central European Myrmicine Ants Myrmica rubra and Myrmica ruginodis. Toxins 2022, 14, 358. [Google Scholar] [CrossRef]
- von Reumont, B.M.; Dutertre, S.; Koludarov, I. Venom profile of the European carpenter bee Xylocopa violacea: Evolutionary and applied considerations on its toxin components. Toxicon X 2022, 14, 100117. [Google Scholar] [CrossRef]
- Chapman, N.C.; Lim, J.; Oldroyd, B.P. Population Genetics of Commercial and Feral Honey Bees in Western Australia. J. Econ. Entomol. 2008, 101, 272–277. [Google Scholar] [CrossRef]
- Oxley, P.R.; Oldroyd, B.P. Mitochondrial Sequencing Reveals Five Separate Origins of ‘Black’ Apis mellifera (Hymenoptera: Apidae) in Eastern Australian Commercial Colonies. J. Econ. Entomol. 2009, 102, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Harpur, B.A.; Minaei, S.; Kent, C.F.; Zayed, A. Management increases genetic diversity of honey bees via admixture. Mol. Ecol. 2012, 21, 4414–4421. [Google Scholar] [CrossRef]
- Magnus, R.M.; Tripodi, A.D.; Szalanski, A.L. Mitochondrial DNA Diversity of Honey Bees (Apis mellifera) from Unmanaged Colonies and Swarms in the United States. Biochem. Genet. 2014, 52, 245–257. [Google Scholar] [CrossRef]
- Wallberg, A.; Han, F.; Wellhagen, G.; Dahle, B.; Kawata, M.; Haddad, N.; Simões, Z.L.P.; Allsopp, M.H.; Kandemir, I.; De la Rúa, P.; et al. A worldwide survey of genome sequence variation provides insight into the evolutionary history of the honeybee Apis mellifera. Nat. Genet. 2014, 46, 1081–1088. [Google Scholar] [CrossRef] [Green Version]
- Espregueira Themudo, G.; Rey-Iglesia, A.; Robles Tascón, L.; Bruun Jensen, A.; da Fonseca, R.R.; Campos, P.F. Declining genetic diversity of European honeybees along the twentieth century. Sci. Rep. 2020, 10, 10520. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.; Wüster, W.; Vonk, F.; Harrison, R.; Fry, B. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Moran, Y. The Rise and Fall of an Evolutionary Innovation: Contrasting Strategies of Venom Evolution in Ancient and Young Animals. PLoS Genet. 2015, 11, e1005596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, A.A. The evolutionary dynamics of venom toxins made by insects and other animals. Biochem. Soc. Trans. 2020, 48, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Danneels, E.; Vaerenbergh, M.; Debyser, G.; Devreese, B.; Graaf, D. Honeybee Venom Proteome Profile of Queens and Winter Bees as Determined by a Mass Spectrometric Approach. Toxins 2015, 7, 4468–4483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konno, K.; Kazuma, K.; Nihei, K.I. Peptide Toxins in Solitary Wasp Venoms. Toxins 2016, 8, 114. [Google Scholar] [CrossRef] [Green Version]
- Nihei, K.i.; Peigneur, S.; Tytgat, J.; Lange, A.B.; Konno, K. Isolation and characterization of FMRFamide-like peptides in the venoms of solitary sphecid wasps. Peptides 2021, 142, 170575. [Google Scholar] [CrossRef]
- Kazuma, K.; Ando, K.; Nihei, K.I.; Wang, X.; Rangel, M.; Franzolin, M.; Mori-Yasumoto, K.; Sekita, S.; Kadowaki, M.; Satake, M.; et al. Peptidomic analysis of the venom of the solitary bee Xylocopa appendiculata circumvolans. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čeřovský, V.; Hovorka, O.; Cvačka, J.; Voburka, Z.; Bednárová, L.; Borovičková, L.; Slaninová, J.; Fučík, V. Melectin: A Novel Antimicrobial Peptide from the Venom of the Cleptoparasitic Bee Melecta albifrons. ChemBioChem 2008, 9, 2815–2821. [Google Scholar] [CrossRef]
- Čeřovský, V.; Buděšínský, M.; Hovorka, O.; Cvačka, J.; Voburka, Z.; Slaninová, J.; Borovičková, L.; Fučík, V.; Bednárová, L.; Votruba, I.; et al. Lasioglossins: Three Novel Antimicrobial Peptides from the Venom of the Eusocial Bee Lasioglossum laticeps (Hymenoptera: Halictidae). ChemBioChem 2009, 10, 2089–2099. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J. Toxinology of venoms from the honeybee genus Apis. Toxicon 1995, 33, 917–927. [Google Scholar] [CrossRef]
- Choo, Y.; Lee, K.; Yoon, H.; Kim, B.; Sohn, M.; Roh, J.; Je, Y.; Kim, N.; Kim, I.; Woo, S.; et al. Dual function of a bee venom serine protease: Prophenoloxidase-activating factor in arthropods and fibrin(ogen)olytic enzyme in mammals. PLoS ONE 2010, 5, 10393. [Google Scholar] [CrossRef]
- Kastin, A. Handbook of Biologically Active Peptides; Academic Press: Boston, MA, USA, 2013. [Google Scholar]
- Habermann, E. Bee and Wasp Venoms: The biochemistry and pharmacology of their peptides and enzymes are reviewed. Science 1972, 177, 314–322. [Google Scholar] [CrossRef]
- King, T.; Spangfort, M. Structure and biology of stinging insect venom allergens. Int. Arch. Allergy Immunol. 2000, 123, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Sobral, F.; Sampaio, A.; Falcão, S.; Queiroz, M.; Calhelha, R.; Vilas-Boas, M.; Ferreira, I. Chemical characterization, antioxidant, anti-inflammatory and cytotoxic properties of bee venom collected in Northeast Portugal. Food Chem. Toxicol. 2016, 94, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.M.; Jeong, Y.J.; Cho, H.J.; Park, K.K.; Chung, I.K.; Lee, I.K.; Kwak, J.Y.; Chang, H.W.; Kim, C.H.; Moon, S.K.; et al. Melittin Suppresses HIF-1α/VEGF Expression through Inhibition of ERK and mTOR/p70S6K Pathway in Human Cervical Carcinoma Cells. PLoS ONE 2013, 8, e69380. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, D.; Gudz, T.; Novgorodov, S.; Erdahl, W. The Peptide Mastoparan Is a Potent Facilitator of the Mitochondrial Permeability Transition. J. Biol. Chem. 1995, 270, 4923–4932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, Y.; Mangetsu, M.; Yang, P.; Kofujita, H.; Suzuki, K.; Ohfune, Y.; Shinada, T. A Quinone Isolated from the Nest of Vespa simillima and Its Growth-Inhibitory Effect on Rat Liver Cancer Cells. Biol. Pharm. Bull. 2008, 31, 722–725. [Google Scholar] [CrossRef] [Green Version]
- Alvares, D.; Ruggiero Neto, J.; Ambroggio, E. Phosphatidylserine lipids and membrane order precisely regulate the activity of Polybia-MP1 peptide. Biochim. Biophys. Acta (BBA)-Biomembr. 2017, 1859, 1067–1074. [Google Scholar] [CrossRef]
- Heinen, T.; Veiga, A. Arthropod venoms and cancer. Toxicon 2011, 57, 497–511. [Google Scholar] [CrossRef]
- Schmidt, J.; Blum, M.; Overal, W. Comparative enzymology of venoms from stinging Hymenoptera. Toxicon 1986, 24, 907–921. [Google Scholar] [CrossRef]
- Ali, S.; Baumann, K.; Jackson, T.; Wood, K.; Mason, S.; Undheim, E.B.; Nouwens, A.; Koludarov, I.; Hendrikx, I.; Jones, A.; et al. Proteomic comparison of Hypnale hypnale (Hump-Nosed Pit-Viper) and Calloselasma rhodostoma (Malayan Pit-Viper) venoms. J. Proteom. 2013, 91, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Yang, D.; Jackson, T.; Undheim, E.B.; Koludarov, I.; Wood, K.; Jones, A.; Hodgson, W.; Mccarthy, S.; Ruder, T.; et al. Venom proteomic characterization and relative antivenom neutralization of two medically important Pakistani elapid snakes (Bungarus sindanus and Naja naja. J. Proteom. 2013, 89, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Jackson, T.; Casewell, N.; Low, D.; Rossi, S.; Baumann, K.; Fathinia, B.; Visser, J.; Nouwens, A.; Hendrikx, I.; et al. Extreme venom variation in Middle Eastern vipers: A proteomics comparison of Eristicophis macmahonii, Pseudocerastes fieldi and Pseudocerastes persicus. J. Proteom. 2015, 116, 106–113. [Google Scholar] [CrossRef]
- Bolger, A.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.; Haas, B.; Yassour, M.; Levin, J.; Thompson, D.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.; Lipman, D. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- The uniprot consortium UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, 158–169. [CrossRef] [PubMed] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Baumann, K.; Vicenzi, E.P.; Lam, T.; Douglas, J.; Arbuckle, K.; Cribb, B.; Brady, S.G.; Fry, B.G. Harden up: Metal acquisition in the weaponized ovipositors of aculeate hymenoptera. Zoomorphology 2018, 137, 389–406, Correction in Zoomorphology 2018, 137, 407–408. [Google Scholar] [CrossRef] [Green Version]

 , and amino acids are colored according to the default settings of AliView [79]. Toxin families include: (A) icarapins, (B) phospholipase A2, (C) anthophilins such as apamin [78], (D) carboxylesterases. (E) Relative length-normalized expression of these toxin families in the transcriptome, measured as total RPK for each family.
, and amino acids are colored according to the default settings of AliView [79]. Toxin families include: (A) icarapins, (B) phospholipase A2, (C) anthophilins such as apamin [78], (D) carboxylesterases. (E) Relative length-normalized expression of these toxin families in the transcriptome, measured as total RPK for each family.
, and amino acids are colored according to the default settings of AliView [79]. Toxin families include: (A) icarapins, (B) phospholipase A2, (C) anthophilins such as apamin [78], (D) carboxylesterases. (E) Relative length-normalized expression of these toxin families in the transcriptome, measured as total RPK for each family.
, and amino acids are colored according to the default settings of AliView [79]. Toxin families include: (A) icarapins, (B) phospholipase A2, (C) anthophilins such as apamin [78], (D) carboxylesterases. (E) Relative length-normalized expression of these toxin families in the transcriptome, measured as total RPK for each family.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Family | Subfamily | Species |
|---|---|---|---|
| Social Bees | Apidae | Apinae | Apis andreniformis |
| Apidae | Apinae | Apis cerana | |
| Apidae | Apinae | Apis dorsata | |
| Apidae | Apinae | Apis florea | |
| Apidae | Apinae | Apis mellifera ligustica (European) | |
| Apidae | Apinae | Apis mellifera scutellata (Africanised) | |
| Apidae | Apinae | Bombus huntii | |
| Apidae | Apinae | Bombus sonorus | |
| Solitary Bees | Apidae | Apinae | Centris aethyctera |
| Apidae | Apinae | Diadasia rinconis | |
| Apidae | Apinae | Xenoglossa angustior | |
| Apidae | Xylocopinae | Xylocopa rufa | |
| Apidae | Xylocopinae | Xylocopa californica | |
| Apidae | Xylocopinae | Xylocopa varipuncta | |
| Colletidae | Diphaglossinae | Crawfordapis sp. | |
| Halictidae | Halictinae | Lasioglossum sp. | |
| Social Wasps | Vespidae | Polistinae | Agelaia myrmecophila |
| Vespidae | Polistinae | Belonogaster juncea colonialis | |
| Vespidae | Polistinae | Brachygastra mellifica | |
| Vespidae | Polistinae | Mischocyttarus flavitarsus | |
| Vespidae | Polistinae | Parachartergus fraternus | |
| Vespidae | Polistinae | Polistes canadensis | |
| Vespidae | Polistinae | Polistes comanchus navajoe | |
| Vespidae | Polistinae | Polistes dorsalis | |
| Vespidae | Polistinae | Polistes flavus | |
| Vespidae | Polistinae | Polistes major castaneocolor | |
| Vespidae | Polistinae | Polybia rejecta | |
| Vespidae | Polistinae | Polybia simillima | |
| Vespidae | Polistinae | Ropalidia sp. | |
| Vespidae | Polistinae | Synoeca septentrionalis | |
| Vespidae | Vespinae | Dolichovespula arenaria | |
| Vespidae | Vespinae | Vespa luctuosa | |
| Vespidae | Vespinae | Vespa mandarinia | |
| Vespidae | Vespinae | Vespa simillima | |
| Vespidae | Vespinae | Vespa tropica | |
| Vespidae | Vespinae | Vespula pensylvanica | |
| Vespidae | Vespinae | Vespula vulgaris | |
| Solitary Wasps | Mutillidae | Sphaeropthalminae | Dasymutilla chiron |
| Mutillidae | Sphaeropthalminae | Dasymutilla gloriosa | |
| Mutillidae | Sphaeropthalminae | Dasymutilla klugii | |
| Scoliidae | Scoliinae | Scoliidae sp. | |
| Crabronidae | Bembicinae | Stictia sp. | |
| Ants | Formicidae | Ectatomminae | Ectatomma tuberculatum |
| Formicidae | Mymicinae | Pogonomyrmex maricopa | |
| Formicidae | Mymicinae | Pogonomyrmex occidentalis | |
| Formicidae | Mymicinae | Pogonomyrmex rugosus | |
| Formicidae | Myrmeciinae | Myrmecia browningi | |
| Formicidae | Myrmeciinae | Myrmecia gulosa | |
| Formicidae | Myrmeciinae | Myrmecia nigriceps | |
| Formicidae | Myrmeciinae | Myrmecia pilosula | |
| Formicidae | Myrmeciinae | Myrmecia simillima | |
| Formicidae | Myrmeciinae | Myrmecia tarsata | |
| Formicidae | Myrmicinae | Daceton sp. | |
| Formicidae | Paraponerinae | Paraponera clavata | |
| Formicidae | Ponerinae | Brachyponera sennaarensis | |
| Formicidae | Ponerinae | Diacamma sp. | |
| Formicidae | Ponerinae | Leptogenys sp. | |
| Formicidae | Ponerinae | Neoponera commutata | |
| Formicidae | Ponerinae | Neoponera commutata (Queen) | |
| Formicidae | Ponerinae | Neoponera villosa | |
| Formicidae | Ponerinae | Odontomachus sp. | |
| Formicidae | Ponerinae | Opthalmopone sp. | |
| Formicidae | Ponerinae | Megaponera analis | |
| Formicidae | Ponerinae | Pachycondyla crassinoda | |
| Formicidae | Ponerinae | Paltothyreus tarsatus | |
| Formicidae | Ponerinae | Platythyrea lamellosa | |
| Formicidae | Ponerinae | Platythyrea strigulosa | |
| Formicidae | Ponerinae | Streblognathus aethiopicus | |
| Formicidae | Ponerinae | Tetraponera sp. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dashevsky, D.; Baumann, K.; Undheim, E.A.B.; Nouwens, A.; Ikonomopoulou, M.P.; Schmidt, J.O.; Ge, L.; Kwok, H.F.; Rodriguez, J.; Fry, B.G. Functional and Proteomic Insights into Aculeata Venoms. Toxins 2023, 15, 224. https://doi.org/10.3390/toxins15030224
Dashevsky D, Baumann K, Undheim EAB, Nouwens A, Ikonomopoulou MP, Schmidt JO, Ge L, Kwok HF, Rodriguez J, Fry BG. Functional and Proteomic Insights into Aculeata Venoms. Toxins. 2023; 15(3):224. https://doi.org/10.3390/toxins15030224
Chicago/Turabian StyleDashevsky, Daniel, Kate Baumann, Eivind A. B. Undheim, Amanda Nouwens, Maria P. Ikonomopoulou, Justin O. Schmidt, Lilin Ge, Hang Fai Kwok, Juanita Rodriguez, and Bryan G. Fry. 2023. "Functional and Proteomic Insights into Aculeata Venoms" Toxins 15, no. 3: 224. https://doi.org/10.3390/toxins15030224