Acontia, a Specialised Defensive Structure, Has Low Venom Complexity in Calliactis polypus

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Acontia Sampling and Observations
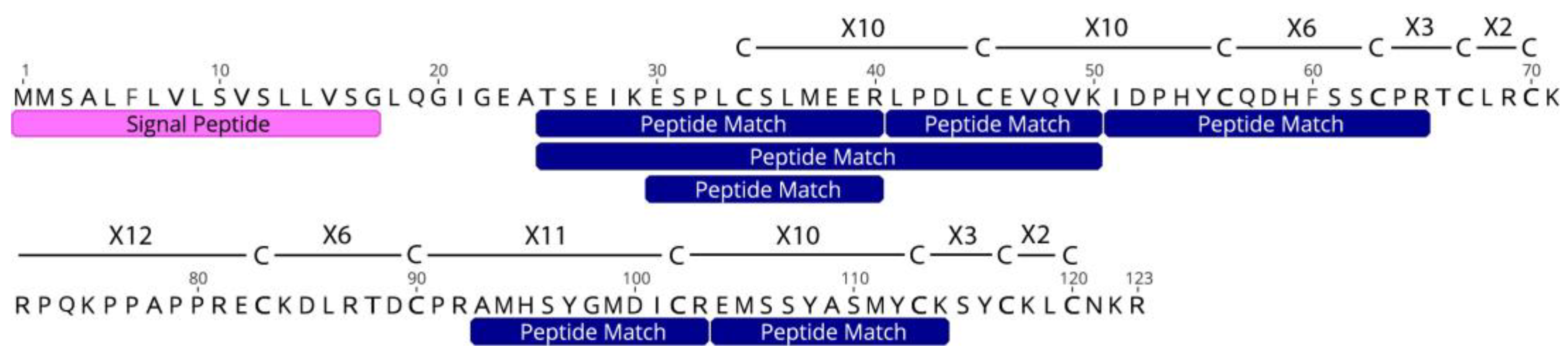
2.2. Identification of Toxins in the Proteome
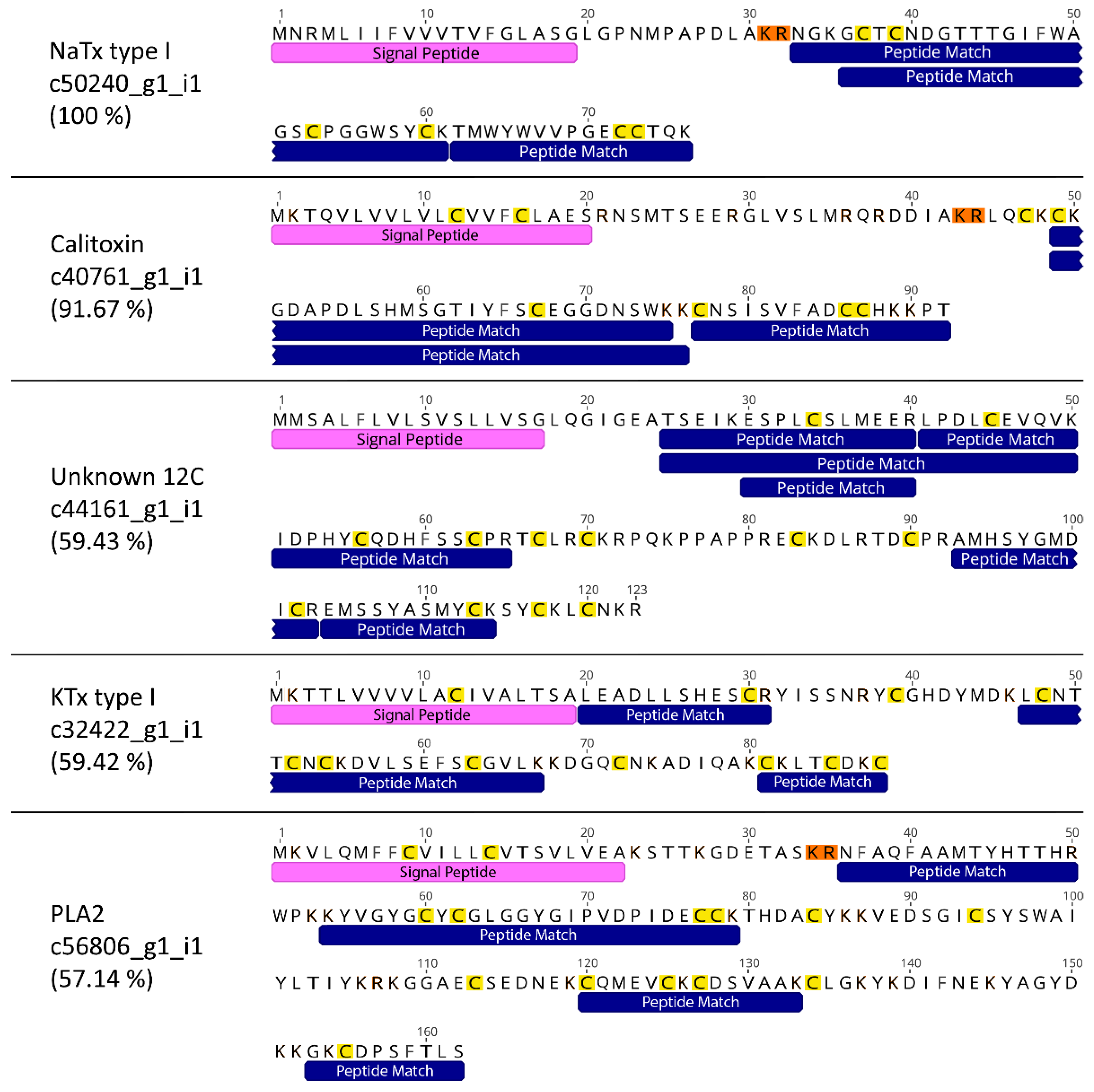
2.3. Comparison of Five Toxins of Interest in Sea Anemone Species
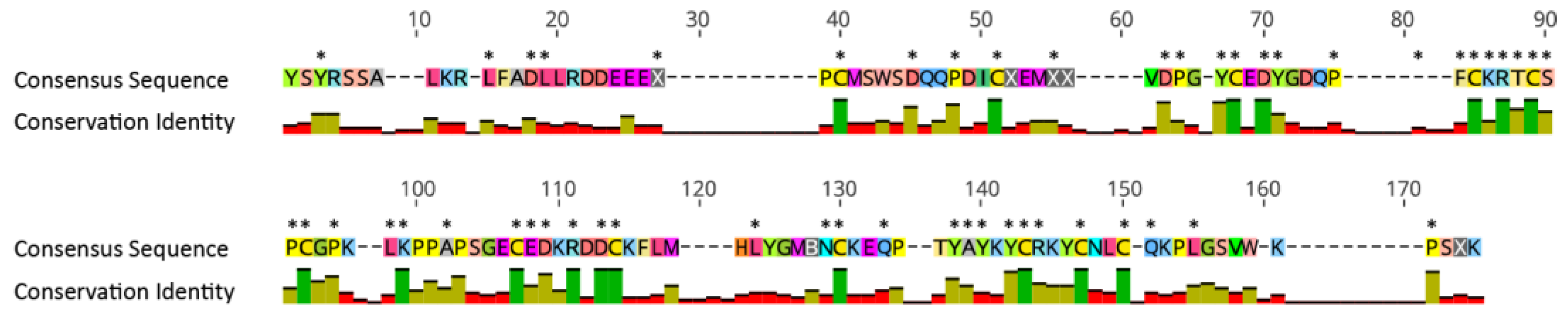
2.4. Phylogenetic Tree Analysis
2.5. Selection Analysis for Unknown 12C
3. Discussion
3.1. Limited Toxin Diversity in Acontia Venom Supports the Dominant Toxin Hypothesis
3.2. Venom in the Acontia Supports a Defensive Role
3.3. Discovery of a Novel Peptide in Acontia
4. Materials and Methods
4.1. Sea Anemone Collection and Acontia Sampling
4.2. Proteome Extraction and Generation
4.3. Proteome Annotation
4.4. Toxin and Toxin-like Gene Identification
4.5. Phylogenetic Analyses
4.6. Selection Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Macrander, J.; Broe, M.; Daly, M. Tissue-Specific Venom Composition and Differential Gene Expression in Sea Anemones. Genome Biol. Evol. 2016, 8, 2358–2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, C.C. Hexacorals 1: Sea Anemones and Anemone-like Animals (Actiniaria, Zoanthidea, Corallimorpharia, Ceriantharia and Antipatharia). In The Great Barrier Reef: Biology, Environment and Management; Hutchings, P., Kingsford, M., Hoegh-Guldberg, O., Eds.; CSIRO: Collingwood, VIC, Australia, 2008; pp. 198–207. ISBN 978-148-630-820-0. [Google Scholar]
- Schendel, V.; Rash, D.L.; Jenner, A.R.; Undheim, A.B. The Diversity of Venom: The Importance of Behavior and Venom System Morphology in Understanding Its Ecology and Evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prentis, P.J.; Pavasovic, A.; Norton, R.S. Sea anemones: Quiet achievers in the field of peptide toxins. Toxins 2018, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Ashwood, L.M.; Norton, R.S.; Undheim, E.A.; Hurwood, D.A.; Prentis, P.J. Characterising Functional Venom Profiles of Anthozoans and Medusozoans within Their Ecological Context. Mar. Drugs 2020, 18, 202. [Google Scholar] [CrossRef] [Green Version]
- Fautin, D.G. Structural diversity, systematics, and evolution of cnidae. Toxicon 2009, 54, 1054–1064. [Google Scholar] [CrossRef]
- Watson, G.M.; Wood, R.L. Colloquium on Terminology. In The Biology of Nematocysts; Hessinger, D.A., Lenhoff, H.M., Eds.; Academic Press: San Diego, CA, USA, 1988; pp. 21–23. ISBN 0-12-345320-8. [Google Scholar]
- Ashwood, L.M.; Mitchell, M.L.; Madio, B.; Hurwood, D.A.; King, G.F.; Undheim, E.A.B.; Norton, R.S.; Prentis, P.J. Tentacle morphological variation coincides with differential expression of toxins in sea anemones. Toxins 2021, 13, 452. [Google Scholar] [CrossRef]
- Columbus-Shenkar, Y.Y.; Sachkova, M.Y.; Macrander, J.; Fridrich, A.; Modepalli, V.; Reitzel, A.M.; Sunagar, K.; Moran, Y. Dynamics of venom composition across a complex life cycle. eLife 2018, 7, e35014. [Google Scholar] [CrossRef]
- Madio, B.; Peigneur, S.; Chin, Y.K.; Hamilton, B.R.; Henriques, S.T.; Smith, J.J.; Cristofori-Armstrong, B.; Dekan, Z.; Boughton, B.A.; Alewood, P.F.; et al. PHAB toxins: A unique family of predatory sea anemone toxins evolving via intra-gene concerted evolution defines a new peptide fold. Cell Mol. Life Sci. 2018, 75, 4511–4524. [Google Scholar] [CrossRef] [Green Version]
- Surm, J.M.; Smith, H.L.; Madio, B.; Undheim, E.A.; King, G.F.; Hamilton, B.R.; van der Burg, C.A.; Pavasovic, A.; Prentis, P.J. A process of convergent amplification and tissue-specific expression dominates the evolution of toxin and toxin-like genes in sea anemones. Mol. Ecol. 2019, 28, 2272–2289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashwood, L.M.; Undheim, E.A.B.; Madio, B.; Hamilton, B.R.; Daly, M.; Hurwood, D.A.; King, G.F.; Prentis, P.P. Venoms for all occasions: The functional toxin profiles of different anatomical regions in sea anemones are related to their ecological function. Mol. Ecol. 2022, 31, 866–883. [Google Scholar] [CrossRef]
- Chen, X.; Leahy, D.; Van Haeften, J.; Hartfield, P.; Prentis, P.J.; van der Burg, C.A.; Hamilton, B.R. A versatile and robust serine protease inhibitor scaffold from Actinia tenebrosa. Mar. Drugs 2019, 17, 701. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, M.L.; Hamilton, B.R.; Madio, B.; Morales, R.A.V.; Tonkin-Hill, G.Q.; Papenfuss, A.T.; Purcell, A.W.; King, G.F.; Undheim, E.A.B.; Norton, R.S. The use of imaging mass spectrometry to study peptide toxin distribution in Australian sea anemones. Aust. J. Chem. 2017, 70, 1235–1237. [Google Scholar] [CrossRef] [Green Version]
- Lam, J.; Cheng, Y.-W.; Chen, W.-N.U.; Li, H.-H.; Chen, C.-S.; Peng, S.-E. A detailed observation of the ejection and retraction of defense tissue acontia in sea anemone (Exaiptasia pallida). PeerJ 2017, 5, e2996. [Google Scholar] [CrossRef] [Green Version]
- Östman, C.; Kultima, J.R.; Roat, C.; Rundblom, K. Acontia and mesentery nematocysts of the sea anemone Metridium senile (Linnaeus, 1761) (Cnidaria: Anthozoa). Sci. Mar. 2010, 74, 483–497. [Google Scholar] [CrossRef] [Green Version]
- Arulvasu, C.; Dhana Sekaran, G.; Prabhakaran, B.; Kalaiselvi, V.; Kathirvel, N.; Lakshmanan, V. Cytotoxic effect of crude venom isolated from Sea anemone Calliactis tricolor on human cancer cell lines. Indian J. Geo. Mar. Sci. 2020, 49, 601–609. [Google Scholar]
- Conklin, E.J.; Bigger, C.H.; Mariscal, R.N. The Formation and Taxonomic Status of the Microbasic Q-Mastigophore Nematocyst of Sea Anemones. Biol. Bull. 1977, 152, 159–168. [Google Scholar] [CrossRef]
- Edmunds, M.; Potts, G.W.; Swinfen, R.C.; Waters, V.L. Defensive behaviour of sea anemones in response to predation by the opisthobranch mollusc Aeolidia papillosa (L.). J. Mar. Biol. Assoc. UK 1976, 56, 65–83. [Google Scholar] [CrossRef]
- Hidaka, M.; Afuso, K. Effects of Cations on the Volume and Elemental Composition of Nematocysts Isolated from Acontia of the Sea Anemone Calliactis polypus. Biol. Bull. 1993, 184, 97–104. [Google Scholar] [CrossRef]
- Hidaka, M.; Mariscal, R.N. Effects of Ions on Nematocysts Isolated from Acontia of the Sea Anemone Calliactis tricolor by different methods. J. Exp. Biol. 1987, 136, 23–24. [Google Scholar] [CrossRef]
- Schlesinger, A.; Zlotkin, E.; Kramarsky-Winter, E.; Loya, Y. Cnidarian internal stinging mechanism. Proc. R. Soc. B. 2009, 276, 1063–1067. [Google Scholar] [CrossRef] [Green Version]
- Ross, D.M. Protection of Hermit Crabs (Dardanus spp.) from Octopus by Commensal Sea Anemones (Calliactis spp.). Nature 1971, 230, 401–402. [Google Scholar] [CrossRef] [PubMed]
- McLean, R.B.; Mariscal, R.N. Protection of a Hermit Crab by its Symbiotic Sea Anemone CaIliactis tricolor. Experientia 1973, 29, 128–130. [Google Scholar] [CrossRef]
- Hessinger, D.A.; Lenhoff, H.M. Assay and Properties of the Hemolysis Activity of Pure Venom from the Nematocysts of the Acontia of the Sea Anemone Aiptasia pallida. Arch. Biochem. Biophys. 1973, 159, 629–638. [Google Scholar] [CrossRef]
- Grotendorst, G.R.; Hessinger, D.A. Purification and partial characterization of the phospholipase A2 and co-lytic factor from sea anemone (Aiptasia pallida) nematocyst venom. Toxicon 1999, 37, 1779–1796. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Genikhovich, G.; Gordon, D.; Wienkoop, S.; Zenkert, C.; Özbek, S.; Technau, U.; Gurevitz, M. Neurotoxin localization to ectodermal gland cells uncovers an alternative mechanism of venom delivery in sea anemones. Proc. R. Soc. B Biol. Sci. 2012, 279, 1351–1358. [Google Scholar] [CrossRef] [Green Version]
- Klinger, A.B.; Eberhardt, M.; Link, A.S.; Namer, B.; Kutsche, L.K.; Schuy, E.T.; Sittl, R.; Hoffmann, T.; Alzheimer, C.; Huth, T.; et al. Sea-anemone toxin ATX-II elicits A-fiber dependent pain and enhances resurgent and persistent sodium currents in large sensory neurons. Mol. Pain 2012, 8, 69. [Google Scholar] [CrossRef] [Green Version]
- van der Burg, C.A.; Prentis, P.J.; Surm, J.M.; Pavasovic, A. Insights into the innate immunome of actiniarians using a comparative genomic approach. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daly, M. Functional and genetic diversity of toxins in sea anemones. In Evolution of Venomous Animals and Their Toxins; Gopalakrishnakone, P., Malhotra, A., Eds.; Springer: Dordrecht, The Netherlands, 2020; ISBN 94-007-6727-7. [Google Scholar]
- Madio, B.; King, G.F.; Undheim, E.A. Sea anemone toxins: A structural overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, M.L.; Tonkin-Hill, G.Q.; Morales, R.A.; Purcell, A.W.; Papenfuss, A.T.; Norton, R.S. Tentacle transcriptomes of the speckled anemone (Actiniaria: Actiniidae: Oulactis sp.): Venom-related components and their domain structure. Mar. Biotech. 2020, 22, 207–219. [Google Scholar] [CrossRef]
- Moran, Y.; Gordon, D.; Gurevitz, M. Sea anemone toxins affecting voltage-gated sodium channels—Molecular and evolutionary features. Toxicon 2009, 54, 1089–1101. [Google Scholar] [CrossRef] [Green Version]
- Sachkova, M.Y.; Singer, S.A.; Macrander, J.; Reitzel, A.M.; Peigneur, S.; Tytgat, J.; Moran, Y. The Birth and Death of Toxins with Distinct Functions: A Case Study in the Sea Anemone Nematostella. Mol. Biol. Evol. 2019, 36, 2001–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachkova, M.Y.; Landau, M.; Surm, J.M.; Macrander, J.; Singer, S.A.; Reitzel, A.M.; Moran, Y. Toxin-like neuropeptides in the sea anemone Nematostella unravel recruitment from the nervous system to venom. PNAS 2020, 117, 27481–27492. [Google Scholar] [CrossRef] [PubMed]
- Kasheverov, I.E.; Logashina, Y.A.; Kornilov, F.D.; Lushpa, V.A.; Maleeva, E.E.; Korolkova, Y.V.; Yu, J.; Zhu, X.; Zhangsun, D.; Luo, S.; et al. Peptides from the Sea Anemone Metridium senile with Modified Inhibitor Cystine Knot (ICK) Fold Inhibit Nicotinic Acetylcholine Receptors. Toxins 2022, 15, 28. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.; Fautin, D.G.; Cappola, V.A. Systematics of the hexacorallia (Cnidaria: Anthozoa). Zoo. J. Linn. Soc. 2003, 139, 419–437. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.G.; Surm, J.M.; Macrander, J.; Simhi, A.; Amir, G.; Sachkova, M.Y.; Lewandowska, M.; Reitzel, A.M.; Moran, Y. Micro and macroevolution of sea anemone venom phenotype. Nat. Comm. 2023, 14, 249. [Google Scholar] [CrossRef] [PubMed]
- Cariello, L.; De Santis, A.; Fiore, F.; Piccoli, R.; Spagnuolo, A.; Zanetti, L.; Parente, A. Calitoxin, a neurotoxic peptide from the sea anemone Calliactis parasitica: Amino-acid sequence and electrophysiological properties. Biochemistry 1989, 28, 2484–2489. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.S.; Redaelli, E.; Zaharenko, A.J.; Cassulini, R.R.; Konno, K.; Pimenta, D.C.; Freitas, J.C.; Clare, J.J.; Wanke, E. Binding Specificity of Sea Anemone Toxins to Nav 1.1–1.6 Sodium Channels. J. Biol. Chem. 2004, 279, 33323–33335. [Google Scholar] [CrossRef] [Green Version]
- Kalman, K.; Pennington, M.W.; Lanigan, M.D.; Nguyen, A.; Rauer, H.; Mahnir, V.; Paschetto, K.; Kem, W.R.; Grissmer, S.; Gutman, G.A.; et al. ShK-Dap22, a Potent Kv1.3-specific Immunosuppressive Polypeptide. J. Biol. Chem. 1998, 273, 32697–32707. [Google Scholar] [CrossRef] [Green Version]
- Sunanda, P.; Krishnarjuna, B.; Peigneur, S.; Mitchell, M.L.; Estrada, R.; Villegas-Moreno, J.; Pennington, M.W.; Tytgat, J.; Norton, R.S. Identification, chemical synthesis, structure, and function of a new KV1 channel blocking peptide from Oulactis sp. Pep. Sci. 2018, 110, e24073. [Google Scholar] [CrossRef]
- Norton, R.S.; Chandy, K.G. Venom-derived peptide inhibitors of voltage-gated potassium channels. Neuropharmacology 2017, 127, 124–138. [Google Scholar] [CrossRef]
- Nevalainen, T.J.; Peuravuori, H.J.; Quinn, R.J.; Llewellyn, L.E.; Benzie, J.A.H.; Fenner, P.J.; Winkel, K.D. Phospholipase A2 in Cnidaria. Comp. Biochem. Physiol. B. 2004, 139, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Madio, B.; Undheim, E.A.; King, G.F. Revisiting venom of the sea anemone Stichodactyla haddoni: Omics techniques reveal the complete toxin arsenal of a well-studied sea anemone genus. J. Proteom. 2017, 166, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, S.; Grishin, E. Convenient nomenclature of cysteine-rich polypeptide toxins from sea anemones. Peptides 2012, 33, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, O.; Sotolongo, V.; Amor, A.M.; Stöcklin, R.; Anderson, A.J.; Harvey, A.L.; Engström, Å.; Wernstedt, C.; Karlsson, E. Characterization of a Potassium Channel Toxin from the Carribbean Sea Anemone Stichodactyla helianthus. Toxicon 1995, 33, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Ashwood, L.M.; Elnahriry, K.A.; Stewart, Z.K.; Shafee, T.; Naseem, M.U.; Szanto, T.G.; van der Burg, C.A.; Smith, H.L.; Surm, J.M.; Undheim, E.A.B.; et al. Genomic, Functional and Structural Analyses Reveal Mechanisms of Evolutionary Innovation within the Sea Anemone 8 Toxin Family. bioRxiv 2022. [Google Scholar] [CrossRef]
- Anderluh, G.; Podlesek, Z.; Maček, P. A common motif in proparts of Cnidarian toxins and nematocyst collagens and its putative role. Biochim. Biophys. Acta 2000, 1476, 372–376. [Google Scholar] [CrossRef]
- Moran, Y.; Gurevitz, M. When positive selection of neurotoxin genes is missing: The riddle of the sea anemone Nematostella vectensis. FEBS J. 2006, 273, 3886–3892. [Google Scholar] [CrossRef]
- Zang, T.; Cuttle, L.; Broszczak, D.A.; Broadbent, J.A.; Tanzer, C.; Parker, T.J. Characterization of the Blister Fluid Proteome for Pediatric Burn Classification. J. Proteome Res. 2019, 18, 69–85. [Google Scholar] [CrossRef]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2020, 49, D458–D460. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifinopoulos, J.; Nguyen, L.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimova, M.; Gil, M.; Dufayard, J.-F.; Dessimoz, C.; Gascuel, O. Survey of Branch Support Methods Demonstrates Accuracy, Power, and Robustness of Fast Likelihood-based Approximation Schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murrell, B.; Wertheim, J.O.; Moola, S.; Weighill, T.; Scheffler, K.; Kosakovsky Pond, S.L. Detecting individual sites subject to episodic diversifying selection. PLoS Genet. 2012, 8, e1002764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosakovsky Pond, S.L.; Frost, S.D. Not so different after all: A comparison of methods for detecting amino acid sites under selection. Mol. Biol. Evol. 2005, 22, 1208–1222. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin Family | Contig | Peptide Hits (Conf ≥ 95) | Sequence Coverage (%) | Mature Protein Coverage (%) |
|---|---|---|---|---|
| Disintegrin and Metalloproteinase | c59360_g1_i1 | 15 | 14.69 | 14.96 |
| Disintegrin and Metalloproteinase | c62072_g1_i1 | 10 | 13.07 | 13.40 |
| Ficolin-type lectin | c60639_g1_i1 | 3 | 10.42 | 11.11 |
| Jellyfish type II | c60596_g1_i1 | 4 | 7.60 | 7.83 |
| KTx type I (ShK) | c32422_g1_i1 | 4 | 46.59 | 59.42 |
| KTx type I (ShK) | c50551_g1_i1 | 3 | 42.16 | 52.44 |
| KTx type III | c8939_g1_i1 | 2 | 26.83 | 46.81 |
| Multicopper oxidase | c56947_g1_i1 | 5 | 21.80 | 23.39 |
| NaTx (Calitoxin) | c40761_g1_i1 | 4 | 52.17 | 91.67 |
| NaTx type I | c50240_g1_i1 | 5 | 57.89 | 100.00 |
| Peptidase M12A | c26461_g1_i1 | 7 | 11.97 | 12.40 |
| Peptidase M12A | c49341_g1_i3 | 4 | 2.47 | 2.55 |
| Peptidase M13 | c63393_g1_i1 | 13 | 5.83 | 5.99 |
| Phospholipase A2 | c56806_g1_i1 | 7 | 49.38 | 57.14 |
| Sea anemone type 8 | c47095_g1_i1 | 2 | 24.39 | 31.75 |
| Unknown | c44161_g1_i1 | 7 | 51.22 | 59.43 |
| Unknown | c56815_g1_i2 | 6 | 37.65 | 42.66 |
| Unknown | c58770_g1_i1 | 17 | 47.61 | 50.56 |
| Functional Category | Toxin Family | Toxin Subtype | Transcript Copy Number | Transcripts Observed at Peptide Level |
|---|---|---|---|---|
| Enzyme | Lectin | C-Type | 4 | 0 |
| Enzyme | Lectin | Ficolin | 2 | 1 |
| Enzyme | Lipase | AB hydrolase | 2 | 0 |
| Enzyme | Lipase | Phospholipase A2 | 4 | 1 |
| Enzyme | Lipase | Type B carboxylesterase | 1 | 0 |
| Enzyme | Metalloprotease | Disintegrin and metalloprotease | 2 | 2 |
| Enzyme | Metalloprotease | Peptidase M12A | 8 | 2 |
| Enzyme | Protease | Multicopper oxidase | 4 | 1 |
| Neurotoxin | Potassium channel toxin | Kazal | 2 | 0 |
| Neurotoxin | Potassium channel toxin | Type I (ShK) | 4 | 2 |
| Neurotoxin | Potassium channel toxin | Type II (venom kunitz) | 5 | 0 |
| Neurotoxin | Potassium channel toxin | Type III | 1 | 1 |
| Neurotoxin | Sodium channel toxin | Calitoxin | 1 | 1 |
| Neurotoxin | Sodium channel toxin | Sea anemone sodium channel toxin | 2 | 1 |
| Neurotoxin | Sodium channel toxin | Type I | 1 | 1 |
| Unknown | Structural class peptide | Sea anemone type 8 | 6 | 1 |
| Unknown | Structural class peptide | Sea anemone type 9 | 1 | 0 |
| Unknown | Unknown | Unknown (12C) | 1 | 1 |
| Unknown | Unknown | Cephalotoxin | 1 | 0 |
| Unknown | Unknown | VP302 | 4 | 0 |
| Total | 56 | 15 |
| Superfamily | Species | Transcript Copy Number | ||||
|---|---|---|---|---|---|---|
| KTx Type III | NaTx type I | PLA2 (12C) | Sea Anemone 8 | Unknown 12C | ||
| Actinioidea | Actinia tenebrosa | 1 | 1 | 1 | 6 | 1 |
| Actinioidea | Actinodendron plumosum | 0 | 0 | 1 | 0 | 1 |
| Actinioidea | Anemonia sulcata | 3 | 1 | 1 | 6 | 1 |
| Actinioidea | Anthopleura buddemeieri | 2 | 1 | 1 | 5 | 1 |
| Actinioidea | Aulactinia veratra | 1 | 0 | 1 | 6 | 2 |
| Actinioidea | Megalactis griffithsi | 0 | 0 | 1 | 2 | 1 |
| Actinioidea | Stichodactyla mertensii | 1 | 0 | 3 | 2 | 2 |
| Edwardsioidea | Edwardsiella carnea | 0 | 0 | 1 | 4 | 1 |
| Edwardsioidea | Nematostella vectensis | 0 | 0 | 1 | 3 | 1 |
| Metridioidea | Calliactis polypus | 1 | 3 | 4 | 9 | 2 |
| Metridioidea | Exaiptasia diaphana | 1 | 0 | 2 | 4 | 2 |
| Metridioidea | Nemanthus annamensis | 1 | 6 | 2 | 5 | 2 |
| Metridioidea | Telmatactis stephensoni | 2 | 0 | 3 | 7 | 2 |
| Metridioidea | Triactis producta | 2 | 0 | 1 | 3 | 2 |
| Total | 15 | 12 | 23 | 62 | 21 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, H.L.; Prentis, P.J.; Bryan, S.E.; Norton, R.S.; Broszczak, D.A. Acontia, a Specialised Defensive Structure, Has Low Venom Complexity in Calliactis polypus. Toxins 2023, 15, 218. https://doi.org/10.3390/toxins15030218
Smith HL, Prentis PJ, Bryan SE, Norton RS, Broszczak DA. Acontia, a Specialised Defensive Structure, Has Low Venom Complexity in Calliactis polypus. Toxins. 2023; 15(3):218. https://doi.org/10.3390/toxins15030218
Chicago/Turabian StyleSmith, Hayden L., Peter J. Prentis, Scott E. Bryan, Raymond S. Norton, and Daniel A. Broszczak. 2023. "Acontia, a Specialised Defensive Structure, Has Low Venom Complexity in Calliactis polypus" Toxins 15, no. 3: 218. https://doi.org/10.3390/toxins15030218