Detoxification of Aflatoxins in Fermented Cereal Gruel (Ogi) by Probiotic Lactic Acid Bacteria and Yeasts with Differences in Amino Acid Profiles


Abstract
:1. Introduction
2. Results
2.1. LAB and Yeast Populations
2.2. LAB Isolates and Their Identification
2.3. Yeast Isolates and Their Identification
2.4. Molecular Characterization
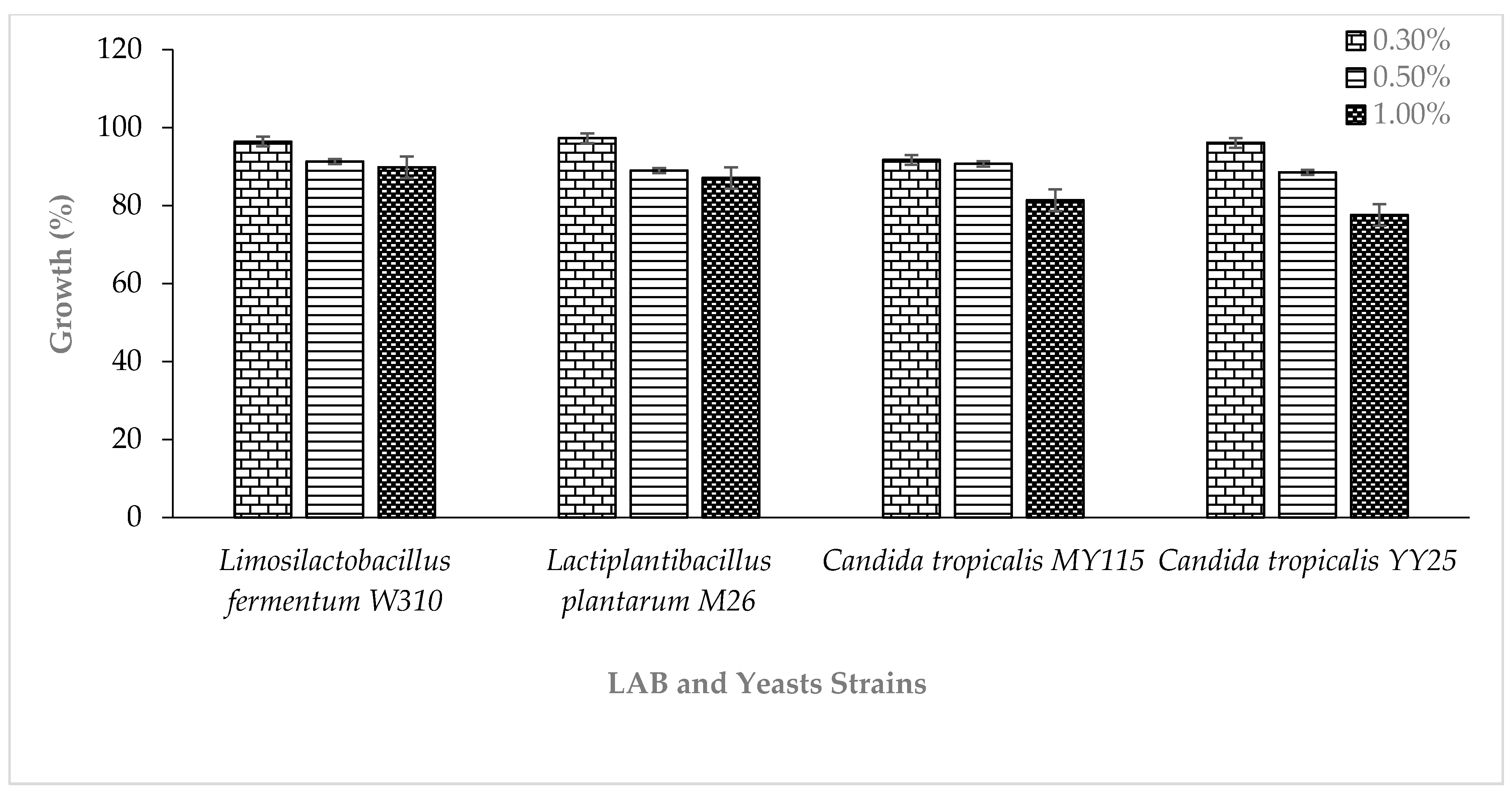
2.5. Probiotic Potentials and Safety Assessment of the LAB and Yeasts Strains
2.6. Aflatoxin Concentration and Amino Acid Changes Due to LAB and Yeast Fermentation
2.6.1. Aflatoxin Concentration Changes
2.6.2. Amino Acid Profiles without the Inclusion of Probiotics
2.6.3. Amino Acid Concentrations with the Inclusion of LAB and Aspergillus
2.6.4. Amino Acid Concentrations with the Inclusion of Yeasts and Aspergillus
2.6.5. Amino Acid Concentrations with the Inclusion of Probiotics, Excluding Aspergillus
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Samples Collection
5.2. Isolation and Characterization of Lactic Acid Bacteria
5.3. Isolation and Characterization of Yeasts
5.4. Determination of Probiotic Potentials of the LAB and Yeast Isolates
5.5. Safety Assessment of the Pre-Selected LAB and Yeasts
5.6. Molecular Characterization of the Selected LAB and Yeast Isolates
5.7. Controlled Fermentation with LAB and Yeasts in the Presence of Toxigenic and Atoxigenic A. flavus Strains
5.8. Aflatoxin Quantification
5.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ajani, J.; Pasha, K.V. Screening of Millets for the Natural Occurrence of Aflatoxin B1 and their Susceptibility in Samples Collected during Pre-harvesting, Harvesting and Post harvesting Conditions. Indian J. Adv. Chem. Sci. 2017, 5, 340–343. [Google Scholar]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. Sorghum Grain: From Genotype, Nutrition and Phenolic Profile to its Health Benefits and Food Applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2025–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayitesi, E.; Onojakpor, O.; Moyo, S.M. Highlighting the Impact of Lactic-Acid-Bacteria-Derived Flavours or Aromas on Sensory Perception of African Fermented Cereals. Fermentation 2023, 9, 111. [Google Scholar] [CrossRef]
- Nyanzi, R.; Jooste, P.J. Cereal-Based Functional Foods, Probiotics; Rigobelo, E., Ed.; InTech: London, UK, 2012; ISBN 978-953-510776-7. Available online: https://www.intechopen.com/chapters/39605 (accessed on 30 January 2023).
- Chaves-Lopez, C.; Rossi, C.; Maggio, F.; Paparella, A.; Serio, A. Changes Occurring in Spontaneous Maize Fer-mentation: An Overview. Fermentation 2020, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Bolaji, O.T.; Adepoju, P.A.; Olalusi, A.P. Economic implication of industrialization of a popular weaning food ogi production in Nigeria: A review. Afr. J. Food Sci. 2015, 9, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Yang, H.; Jiao, Y.; Pang, Q.; Wang, Y.; Wang, M.; Shan, A.; Feng, X. Dietary Curcumin Alleviated Acute Ileum Damage of Ducks (Anas platyrhynchos) Induced by AFB1 through Regulating Nrf2-ARE and NF-κB Signaling Pathways. Foods 2021, 10, 1370. [Google Scholar] [CrossRef]
- Pickova, D.; Ostry, V.; Toman, J.; Malir, F. Aflatoxins: History, Significant Milestones, Recent Data on Their Toxicity and Ways to Mitigation. Toxins 2021, 13, 399. [Google Scholar] [CrossRef] [PubMed]
- Jalili, M.A. A Review on Aflatoxins Reduction in Food. Iran J. Health Saf. Environ. 2016, 3, 445–459. [Google Scholar]
- Alshannaq, A.; Yu, J.H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [Green Version]
- Chavez, R.A.; Cheng, X.; Herrman, T.J.; Stasiewicz, M.J. Single kernel aflatoxin and fumonisin contamination distribution and spectral classification in commercial corn. Food Control. 2021, 131, 108393. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Ortega-Beltran, A.; Akande, A.; Mutegi, C.; Atehnkeng, J.; Kaptoge, L.; Senghor, A.; Adhikari, B.; Cotty, P. Biological control of aflatoxins in Africa: Current status and potential challenges in the face of climate change. World Mycotoxin J. 2016, 9, 771–789. [Google Scholar] [CrossRef] [Green Version]
- Santini, A.; Ritieni, A. Aflatoxins: Risk, Exposure and Remediation. In Aflatoxins Recent Advances and Future Prospects; Razzaghi-Abyaneh, M., Ed.; InTechOpen: London, UK, 2013; pp. 143–265. [Google Scholar]
- Ortega-Beltran, A.; Agbetiameh, D.; Atehnkeng, J.; Falade, T.D.O.; Bandyopadhyay, R. Does Use of Atoxigenic Biocontrol Products to Mitigate Aflatoxin in Maize Increase Fumonisin Content in Grains? Plant Dis. 2021, 105, 2196–2201. [Google Scholar] [CrossRef] [PubMed]
- Reid, G. Probiotics and prebiotics—Progress and challenges. Int. Dairy J. 2008, 18, 969–975. [Google Scholar] [CrossRef]
- Ukwuru, M.U.; Ohaegbu, C.G. Local cereal fermented foods with probiotic potentials. Res. J. Food Nutr. 2018, 2, 1–13. [Google Scholar]
- Banwo, K.; Asogwa, F.C.; Ogunremi, O.R.; Adesulu-Dahunsi, A.; Sanni, A. Nutritional profile and antioxidant capacities of fermented millet and sorghum gruels using lactic acid bacteria and yeasts. Food Biotechnol. 2021, 35, 199–220. [Google Scholar] [CrossRef]
- Byakika, S.; Mukisa, I.M.; Wacoo, A.P.; Kort, R.; Byaruhanga, Y.B.; Muyanja, C. Potential application of lactic acid starters in the reduction of aflatoxin contamination in fermented sorghum-millet beverages. Int. J. Food Contam. 2019, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Adebo, J.A.; Njobeh, P.B.; Gbashi, S.; Oyedeji, A.B.; Ogundele, O.M.; Oyeyinka, S.A.; Adebo, O.A. Fermentation of Cereals and Legumes: Impact on Nutritional Constituents and Nutrient Bioavailability. Fermentation 2022, 8, 63. [Google Scholar] [CrossRef]
- Okara, J.O.; Lokoyi, O.O. Developing an Efficient Method for Ogi Production: Towards Educating the Rural Women. Niger. J. Res. Prod. 2012, 20, 1–7. [Google Scholar]
- Steve, I.O.; David, O.T.; Oluwadunsin, O.A.; Bolawa, M.S. Comparative Study on Nutrient Composition, Functional Property and Glycaemic Index of “Ogi” in Healthy Rats Prepared from Selected Cereal Grains. J. Future Foods 2022, 2, 2022. [Google Scholar] [CrossRef]
- Martinez-Miranda, M.M.; Rosero-Moreano, M.; Taborda-Ocampo, G. Occurrence, Dietary Exposure and Risk As-sessment of Aflatoxins in Arepa, Bread and Rice. Food Control 2019, 98, 359–366. [Google Scholar] [CrossRef]
- Wacoo, A.P.; Mukisa, I.M.; Meeme, R.; Byakika, S.; Wendiro, D.; Sybesma, W.; Kort, R. Probiotic Enrichment and Reduction of Aflatoxins in a Traditional African Maize-Based Fermented Food. Nutrients 2019, 11, 265. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A Global Concern for Food Safety, Human Health and Their Management. Front. Microbiol. 2017, 07, 2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, F.; Zhou, X.; Liu, M.; Zang, H.; Liu, X.; Shan, A.; Feng, X. Alleviation of Oral Exposure to Aflatoxin B1-Induced Renal Dysfunction, Oxidative Stress, and Cell Apoptosis in Mice Kidney by Curcumin. Antioxidants 2022, 11, 1082. [Google Scholar] [CrossRef] [PubMed]
- FAO; WHO. Guidelines for the Evaluation of Probiotics in Food; Food and Agriculture Organization of the United Nations: Rome, Italy; World Health Organization: London, ON, Canada, 2002. Available online: https://www.fao.org/3/a0512e/a0512e.pdf (accessed on 30 January 2023).
- Vinderola, C.G.; Reinheimer, J.A. Lactic acid starter and probiotic bacteria, a comparative ‘in vitro’ study of probiotic characteristics and biological barrier resistance. Food Res. Int. 2003, 36, 895–904. [Google Scholar] [CrossRef]
- Kostinek, M.; Specht, I.; Edward, V.A.; Pinto, C.; Egounlety, M.; Sossa, C.; Mbugua, S.; Dorti, C.; Thonart, P.; Taljaard, L.; et al. Characterization and biochemical properties of predominant lactic acid bacteria from fermenting cassava for selection as starter cultures. Int. J. Food Microbiol. 2007, 114, 342–351. [Google Scholar] [CrossRef]
- Banwo, K.; Sanni, A.; Tan, H. Technological properties and probiotic potential of Enterococcus faecium strains isolated from cow milk. J. Appl. Microbiol. 2012, 114, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Olatunde, O.O.; Obadina, A.O.; Omemu, A.M.; Oyewole, O.B.; Olugbile, A.; Olukomaiya, O.O. Screening and molecular identification of potential probiotic lactic acid bacteria in effluents generated during Ogi production. Ann. Microbiol. 2018, 68, 433–443. [Google Scholar] [CrossRef]
- Ogunremi, O.; Sanni, A.; Agrawal, R. Probiotic potentials of yeasts isolated from some cereal-based Nigerian traditional fermented food products. J. Appl. Microbiol. 2015, 119, 797–808. [Google Scholar] [CrossRef]
- Falade, T.D.O.; Chrysanthopoulos, P.K.; Hodson, M.P.; Sultanbawa, Y.; Fletcher, M.; Darnell, R.; Korie, S.; Fox, G. Metabolites Identified during Varied Doses of Aspergillus Species in Zea mays Grains, and Their Correlation with Aflatoxin Levels. Toxins 2018, 10, 187. [Google Scholar] [CrossRef] [Green Version]
- Petrova, P.; Arsov, A.; Tsvetanova, F.; Parvanova-Mancheva, T.; Vasileva, E.; Tsigoriyna, L.; Petrov, K. The Complex Role of Lactic Acid Bacteria in Food Detoxification. Nutrients 2022, 14, 2038. [Google Scholar] [CrossRef]
- Akin-Osanaiye, B.C.; Kamalu, I.O. Evaluation of ogi (corn caramel) from maize and sorghum for isolation and characterization of lactic acid bacteria (LAB). Biochem. Mol. Biol. 2019, 4, 28–34. [Google Scholar]
- Fawole, M.O.; Oso, B.A. Characterization of Bacteria: Laboratory Manual of Microbiology, 4th ed.; Spectrum Book Limited: Ibadan, Nigeria, 2004. [Google Scholar]
- Alakeji, T.P.; Banwo, K.; Ogunremi, O.R.; Sanni, A.I. Functional properties of yeasts isolated from some Nigerian tradi-tional fermented foods. J. Microbiol. Biotechnol. Food Sci. 2015, 4, 437–441. [Google Scholar] [CrossRef]
- Yoon, M.Y.; Kim, Y.J.; Hwang, H.-J. Properties and safety aspects of Enterococcus faecium strains isolated from Chungkukjang, a fermented soy product. LWT 2008, 41, 925–933. [Google Scholar] [CrossRef]
- Gupta, H.; Malik, R.K. Incidence of virulence in bacteriocin-producing enterococcal isolates. Le Lait 2007, 87, 587–601. [Google Scholar] [CrossRef]
- Omar, N.B.; Castro, A.; Lucas, R.; Abriouel, H.; Yousif, N.M.; Franz, C.M.; Holzapfel, W.H.; Rubén, P.P.; Martínez-Canãmero, M.; Gálvez, A. Functional and safety aspects of enterococci isolated from different Spanish foods. Syst. Appl. Microbiol. 2004, 27, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Dela-Cruz, T.E. Gelatin Hydrolysis Test Protocol; American Society for Microbiology: Washington, DC, USA, 2012; Available online: https://asm.org/ASM/media/Protocol-Images/Gelatin-Hydrolysis-Test-Protocol.pdf?ext=.pdf (accessed on 30 January 2023).
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungi Ribosomal RNA Genes for Phy-Logenetics. In PCR Protocols. A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Kostinek, M.; Specht, I.; Edward, V.A.; Schillinger, U.; Hertel, C.; Holzapfel, W.H.; Franz, C.M. Diversity and technological properties of predominant lactic acid bacteria from fermented cassava used for the preparation of Gari, a traditional African food. Syst. Appl. Microbiol. 2005, 28, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Banwo, K.; Sanni, A.; Tan, H.; Tian, Y. Phenotypic and Genotypic Characterization of Lactic Acid Bacteria Isolated from Some Nigerian Traditional Fermented Foods. Food Biotechnol. 2012, 26, 124–142. [Google Scholar] [CrossRef]
- Agbetiameh, D.; Awuah, R.T.; Atehnkeng, J.; Islam, M.; Callicott, K.A.; Cotty, P.J.; Bandyopadhyay, R. Potential of Atoxigenic Aspergillus flavus Vegetative Compatibility Groups Associated With Maize and Groundnut in Ghana as Biocontrol Agents for Aflatoxin Management. Front. Microbiol. 2019, 10, 2069. [Google Scholar] [CrossRef]
- Alamu, E.O.; Menkir, A.; Adesokan, M.; Fawole, S.; Maziya-Dixon, B. Near-Infrared Reflectance Spectrophotometry (NIRS) Application in the Amino Acid Profiling of Quality Protein Maize (QPM). Foods 2022, 11, 2779. [Google Scholar] [CrossRef]
- Atehnkeng, J.; Ojiambo, P.S.; Donner, M.; Ikotun, T.; Sikora, R.A.; Cotty, P.J.; Bandyopadhyay, R. Distribution and toxigenicity of Aspergillus species isolated from maize kernels from three agro-ecological zones in Nigeria. Int. J. Food Microbiol. 2008, 122, 74–84. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| LAB | Compounds Able to Utilize | Compounds Unable to Utilize | Occurrence (%) | Types of Ogi Isolated from |
|---|---|---|---|---|
| Lb. acidophilus | Glucose, galactose, fructose, d-sorbitol, and lactose | Sucrose and mannitol | 5% | Maize (yellow and white) |
| Lb. brevis | Glucose, galactose, fructose, d-sorbitol, and lactose | Sucrose and mannitol | 8% | Millet, sorghum, and yellow maize |
| Lc. casei | Glucose, sucrose, galactose, fructose, mannitol, lactose, and d-sorbitol (Y16 only) | d-sorbitol, sucrose (Y16 only) and mannitol (Y16 and Y42 only), d-sorbitol (Y42 only), and galactose (Y42 only) | 10% | Maize (yellow and white), millet, and sorghum |
| Lb. delbrueckii | Glucose, fructose, d-sorbitol, and lactose | Sucrose, galactose, and mannitol | 17% | Maize (yellow and white), millet, and sorghum |
| Lm. Fermentum | Glucose, sucrose, galactose, fructose, lactose, and mannitol (Y310) | d-sorbitol and mannitol | 15% | Maize (yellow and white), millet, and sorghum |
| Lb. leichmanii | Glucose, sucrose, fructose, and lactose | Galactose, d-sorbitol, and mannitol | 3% | Millet and sorghum |
| Lb. paraplantarum | Glucose, galactose, d-sorbitol, Lactose (W44 only), and fructose (W44 only) | Sucrose, fructose, mannitol, and lactose | 5% | Maize (yellow and white) |
| Lb. pentosus | Glucose, galactose, fructose, d-sorbitol, and lactose | Sucrose and mannitol | 8% | Millet and sorghum |
| Lp. plantarum | Glucose, sucrose, galactose, fructose, and mannitol | d-sorbitol, lactose, and mannitol (W37) | 12% | Maize (yellow and white), millet, and sorghum |
| Leuc. mesenteroides | Glucose, sucrose, galactose, and lactose | Fructose, D-sorbitol, and mannitol | 17% | Maize (yellow and white), millet, and sorghum |
| Yeasts | Compounds Able to Utilize | Compounds Unable to Utilize | Occurrence (%) |
|---|---|---|---|
| Candida krusei | Glucose | Sucrose, galactose, fructose, d-sorbitol, mannitol, and lactose | 35% |
| C. tropicalis | Glucose and sucrose, galactose and fructose, | Mannitol and lactose | 35% |
| Geotrichum candidum | Glucose and fructose | Sucrose, galactose, d-sorbitol, mannitol, and lactose | 30% |
| LAB/Yeast Strains | 0 h | 6 h | 24 h | 48 h | Hemolysis | Gelatinase | DNAse |
|---|---|---|---|---|---|---|---|
| Limosilactobacillus fermentum W310 | 6.54 | 5.92 | 3.78 | 3.70 | γ | - | - |
| Lactiplantibacillus plantarum M26 | 6.54 | 5.48 | 4.26 | 3.65 | γ | - | - |
| Candida tropicalis MY115 | 6.54 | 5.85 | 4.27 | 4.26 | γ | - | - |
| Candida tropicalis YY25 | 6.54 | 5.90 | 5.00 | 4.60 | γ | - | - |
| Aflatoxin Conc in ng/g | Perc. Reduction | |||||
|---|---|---|---|---|---|---|
| Treatments | B1 | B2 | G1 | G2 | B1 | B2 |
| Ogi alone | 0 | 0 | 0 | 0 | NA | NA |
| Ogi and L. plantarum M26 | 0 | 0 | 0 | 0 | NA | NA |
| Ogi and L. fermentum W310 | 0 | 0 | 0 | 0 | NA | NA |
| Ogi and C. tropicalis MY115 | 0 | 0 | 0 | 0 | NA | NA |
| Ogi and C. tropicalis YY25 | 0 | 0 | 0 | 0 | NA | NA |
| Ogi and A. flavus 3279 (atoxigenic) | 0 | 0 | 0 | 0 | NA | NA |
| Ogi and A. flavus 3228 (toxigenic) | 7048 | 309 | 0 | 0 | NA | NA |
| Ogi, Lp. plantarum M26, and A. flavus 3279 | 0 | 0 | 0 | 0 | NA | NA |
| Ogi, Lp. plantarum M26, and A. flavus 3228 | 2704 | 113 | 0 | 0 | 62 | 63 |
| Ogi, Lm. fermentum W310, and A. flavus 3279 | 0 | 0 | 0 | 0 | NA | NA |
| Ogi, Lm. fermentum W310, and A. flavus 3228 | 965 | 77 | 0 | 0 | 86 | 75 |
| Ogi, C. tropicalis MY115, and A. flavus 3279 | 0 | 0 | 0 | 0 | NA | NA |
| Ogi, C. tropicalis MY115 and A. flavus 3228 | 2827 | 70 | 0 | 0 | 60 | 77 |
| Ogi, C. tropicalis YY25, and A. flavus 3279 | 0 | 0 | 0 | 0 | NA | NA |
| Ogi, C. tropicalis YY25, and A. flavus 3228 | 2835 | 212 | 0 | 0 | 60 | 31 |
| Treatments | Asp | Ser | Gln | Gly | His | Thr | Arg | Ala | Pro | Cys | Tyr | Val | Met | Lys | Ile | Leu | Phe |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ogi alone | 0.65 C | 0.18 DE | 0.74 CD | 0.32 D | 0.32 BCD | 0.1 DE | 0.05 D | 0.3 CDE | 0.39 C | 0.06 D | 0.11 DEF | 0.15 BC | 0.01 DE | 0.49 C | 0.14 B | 0.14 D | 0.08 CD |
| Ogi + 3228 (toxigenic) | 1.99 B | 1.61 B | 8.15 B | 1.12 B | 0.77 AB | 1.4 A | 1.77 AB | 2.34 B | 2.68 A | 7.27 A | 0.15 DE | 1.65 A | 0.08 C | 0.49 C | 1.06 B | 3.63 A | 1.28 B |
| Ogi + 3279 (atoxigenic) | 0.69 C | 0.12 E | 0.72 CD | 0.12 EF | 0.13 CD | 0.36 CD | 0.02 D | 0.39 CD | 0.09 D | 0.15 C | 0.09 DEF | 0.11 BC | 0.01 DE | 0.46 C | 0.17 B | 0.02 E | 0.01 D |
| Ogi + Candida tropicalis (millet ogi) | 0.7 C | 0.18 DE | 0.75 CD | 0.15 E | 0.19 BCD | 0.22 DE | 0.08 D | 0.27 CDE | 0.22 CD | 0.08 D | 0.12 DEF | 0.08 BC | 0.09 C | 0.49 C | 0.15 B | 0.1 D | 0.15 B |
| Ogi + Candida tropicalis (millet ogi) + 3228 | 2.78 A | 2.00 A | 9.45 A | 1.51 A | 0.82 AB | 1.64 A | 2.05 A | 2.98 A | 3.09 A | 0.68 B | 1.73 A | 2.05 A | 0.35 B | 0.85 B | 4.4 A | 0.1 D | 1.4 B |
| Ogi + Candida tropicalis (millet ogi) + 3279 | 0.64 C | 0.05 FG | 0.54 D | 0.08 A | 1.23 A | 0.61 BC | 1.12 C | 0.24 DEF | 0.39 C | 0.17 C | 0.04 F | 0.04 C | 0.02 DE | 0.59 C | 0.24 B | 0.13 D | 0.14 CD |
| Ogi + Candida tropicalis (yellow maize ogi) | 0.55 C | 0.17 DE | 0.74 CD | 0.02 G | 0.02 D | 0.03 E | 0.03 D | 0.12 F | 0.2 CD | 0.06 D | 0.09 DEF | 0.08 BC | 0.01 DE | 0.51 C | 0.13 B | 0.14 D | 0.54 C |
| Ogi + Candida tropicalis (yellow maize ogi) + 3228 | 2.78 A | 1.94 A | 8.89 AB | 1.34 AB | 0.71 AB | 1.59 A | 1.97 A | 2.66 AB | 2.71 A | 0.07 D | 1.32 B | 0.14 BC | 0.14 C | 1.22 A | 4.48 A | 0.02 E | 1.56 B |
| Ogi + Candida tropicalis (yellow maize ogi) + 3279 | 1.92 B | 1.43 C | 7.96 B | 1.13 AB | 0.72 AB | 0.93 B | 1.59 B | 2.43 B | 2.61 A | 0.06 D | 1.22 B | 1.43 A | 0.03 D | 0.46 C | 1.79 B | 1.07 B | 1.2 B |
| Ogi + Limosilactobacillus fermentum | 0.54 C | 0.15 DE | 0.97 C | 0.7 C | 0.57 ABC | 2.03 A | 1.15 C | 2.68 AB | 0.93 B | 0.06 D | 0.06 EF | 0.41 B | 0.49 A | 0.46 C | 0.1 B | 0.26 C | 3.83 A |
| Ogi + Limosilactobacillus fermentum (yellow maize ogi) + 3228 | 0.68 C | 0.08 F | 0.65 CD | 0.01 G | 0.03 D | 0.68 BC | 0.02 D | 0.36 CDE | 0.08 D | 0.06 D | 0.3 C | 0.07 BC | 0.13 C | 0.07 D | 0.07 B | 0.09 D | 0.02 CD |
| Ogi + Limosilactobacillus fermentum (yellow maize ogi) + 3279 | 0.7 C | 0.18 DE | 0.75 CD | 0.03 G | 0.04 CD | 0.05 E | 0.05 D | 0.45 C | 0.29 CD | 0.06 D | 0.11 DEF | 0.16 BC | 0.0 E | 0.59 C | 0.14 B | 0.14 D | 0.12 CD |
| Ogi + Lactiplantibacillus plantarum | 0.71 C | 0.23 D | 0.76 CD | 0.07 EFG | 0.13 CD | 0.03 E | 0.01 D | 0.2 EF | 0.27 CD | 0.06 D | 0.11 DEF | 0.14 BC | 0.01 DE | 0.45 C | 0.14 B | 0.12 D | 0.11 CD |
| Ogi + Lactiplantibacillus plantarum (millet ogi) + 3228 | 0.68 C | 0.02 F | 0.68 CD | 0.01 G | 0.03 D | 0.11 DE | 0.03 D | 0.43 C | 0.25 CD | 0.06 D | 0.19 D | 0.13 BC | 0.01 DE | 0.58 C | 0.1 B | 0.13 D | 0.1 CD |
| Ogi + Lactiplantibacillus plantarum (millet ogi) + 3279 | 0.6 C | 0.06 F | 0.47 CD | 0.05 FG | 0.14 CD | 0.15 ED | 0.09 D | 0.15 F | 0.36 CD | 0.06 D | 0.09 DEF | 0.1 BC | 0.01 DE | 0.57 C | 0.09 B | 1.09 B | 0.09 CD |
| Treatments |
|---|
| Ogi alone |
| Ogi + Lactiplantibacillus plantarum M26 Ogi + Limosilactobacillus fermentum W310 |
| Ogi + Candida tropicalis MY115 Ogi + Candida tropicalis YY25 |
| Ogi + La3279 Ogi + La3228 |
| Ogi + Lactiplantibacillus plantarum M26 + La3279 Ogi + Limosilactobacillus fermentum W310 + La3279 |
| Ogi + Candida tropicalis MY115 + La3279 Ogi + Candida tropicalis YY25 + La3279 |
| Ogi + Lactiplantibacillus plantarum M26 + La3228 Ogi + Limosilactobacillus fermentum W310 + La3228 |
| Ogi + Candida tropicalis MY115 + La3228 Ogi + Candida tropicalis YY25 + La3228 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banwo, K.; Adesina, T.; Aribisala, O.; Falade, T.D.O. Detoxification of Aflatoxins in Fermented Cereal Gruel (Ogi) by Probiotic Lactic Acid Bacteria and Yeasts with Differences in Amino Acid Profiles. Toxins 2023, 15, 210. https://doi.org/10.3390/toxins15030210
Banwo K, Adesina T, Aribisala O, Falade TDO. Detoxification of Aflatoxins in Fermented Cereal Gruel (Ogi) by Probiotic Lactic Acid Bacteria and Yeasts with Differences in Amino Acid Profiles. Toxins. 2023; 15(3):210. https://doi.org/10.3390/toxins15030210
Chicago/Turabian StyleBanwo, Kolawole, Taiwo Adesina, Olubunmi Aribisala, and Titilayo D. O. Falade. 2023. "Detoxification of Aflatoxins in Fermented Cereal Gruel (Ogi) by Probiotic Lactic Acid Bacteria and Yeasts with Differences in Amino Acid Profiles" Toxins 15, no. 3: 210. https://doi.org/10.3390/toxins15030210