Effects of Exposure to Low Zearalenone Concentrations Close to the EU Recommended Value on Weaned Piglets’ Colon


 , ,
, ,
Abstract
:1. Introduction
2. Results
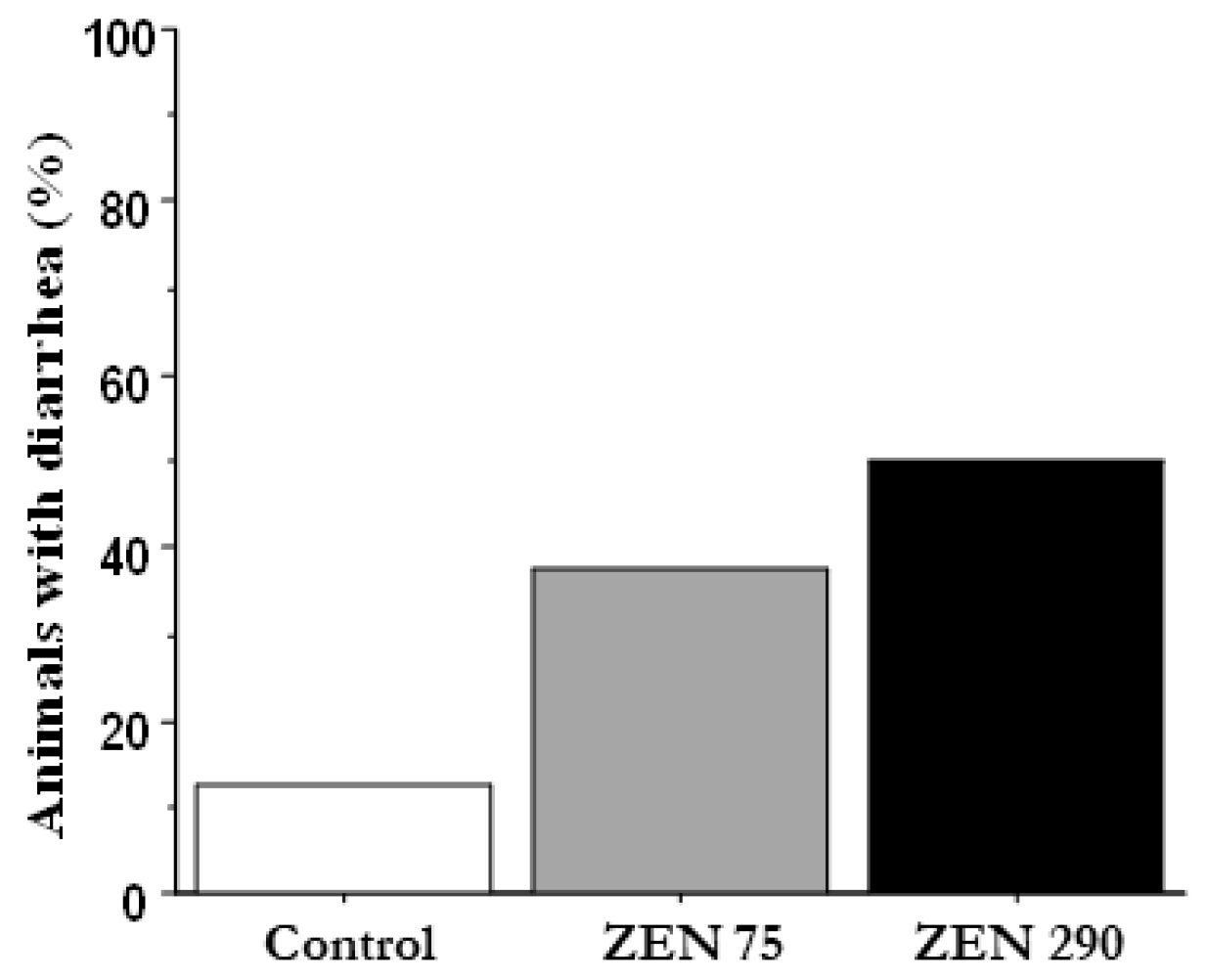
2.1. Effect of ZEN on the Percentage of Animals with Diarrhoea and on Animal Performances
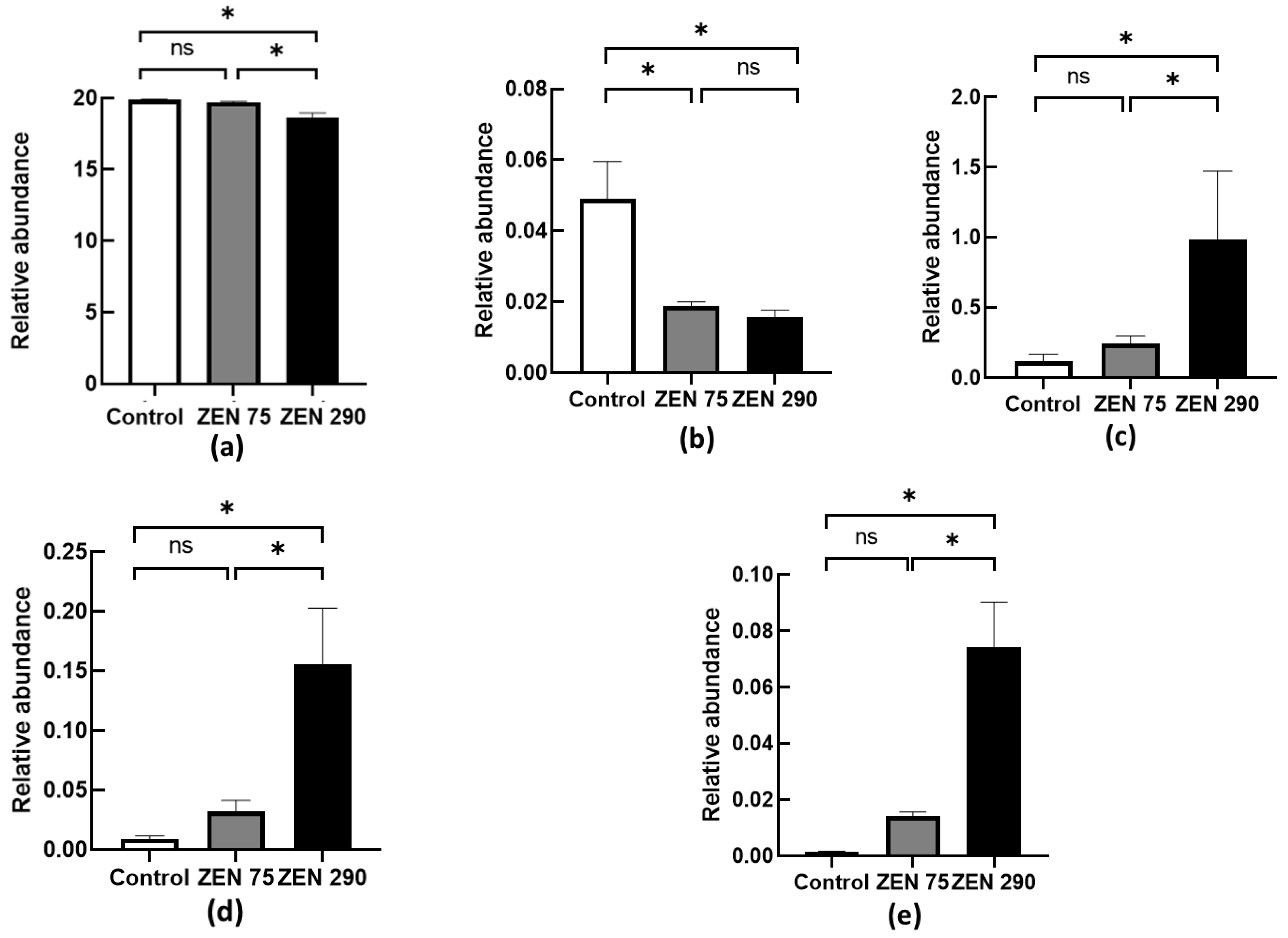
2.2. Relative Abundance of Several Important Microbial Populations under the Influence of ZEN
2.3. Effect of ZEN on the SCFA Colonic Content, pH Value and Ammonia Concentration
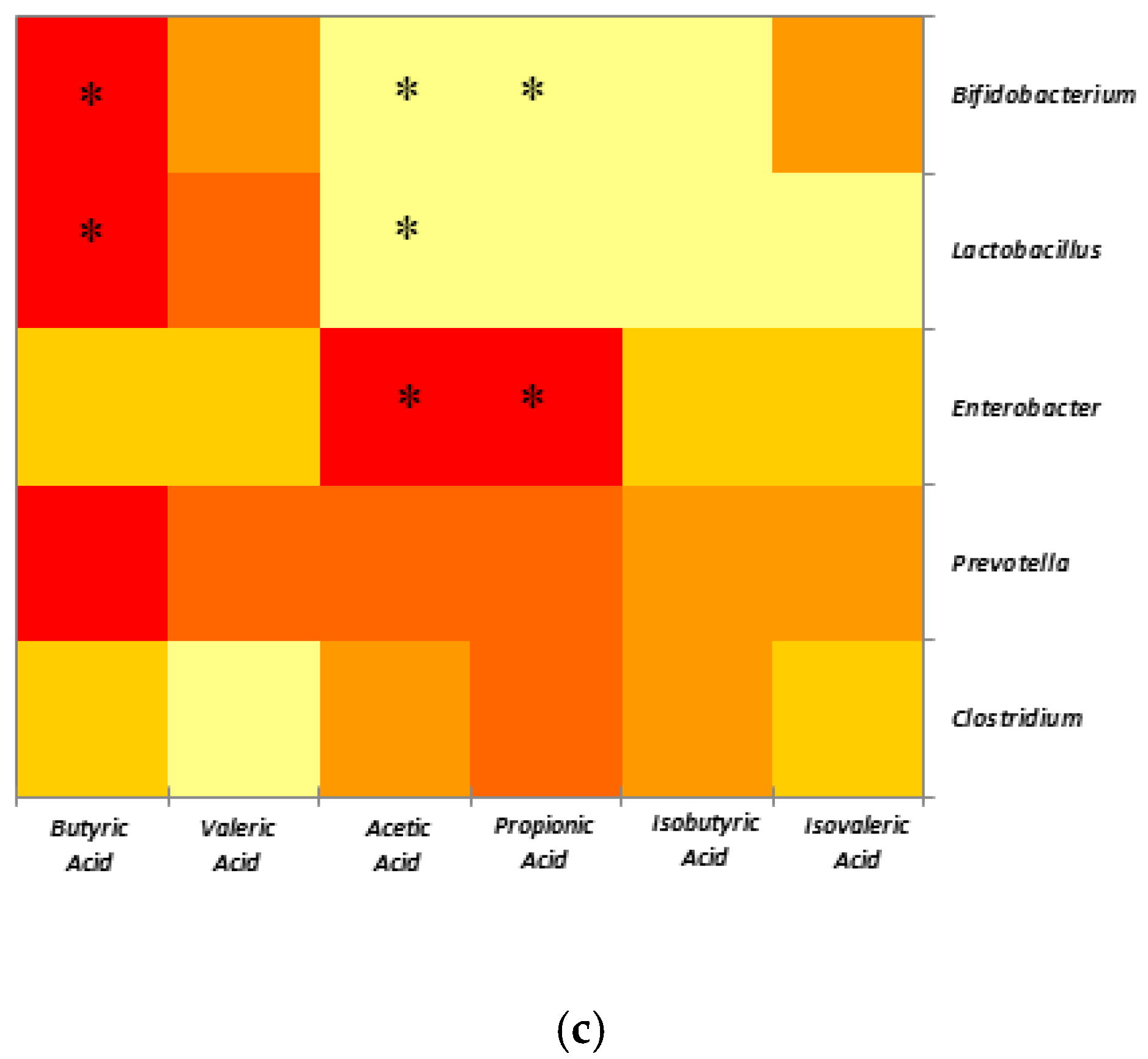
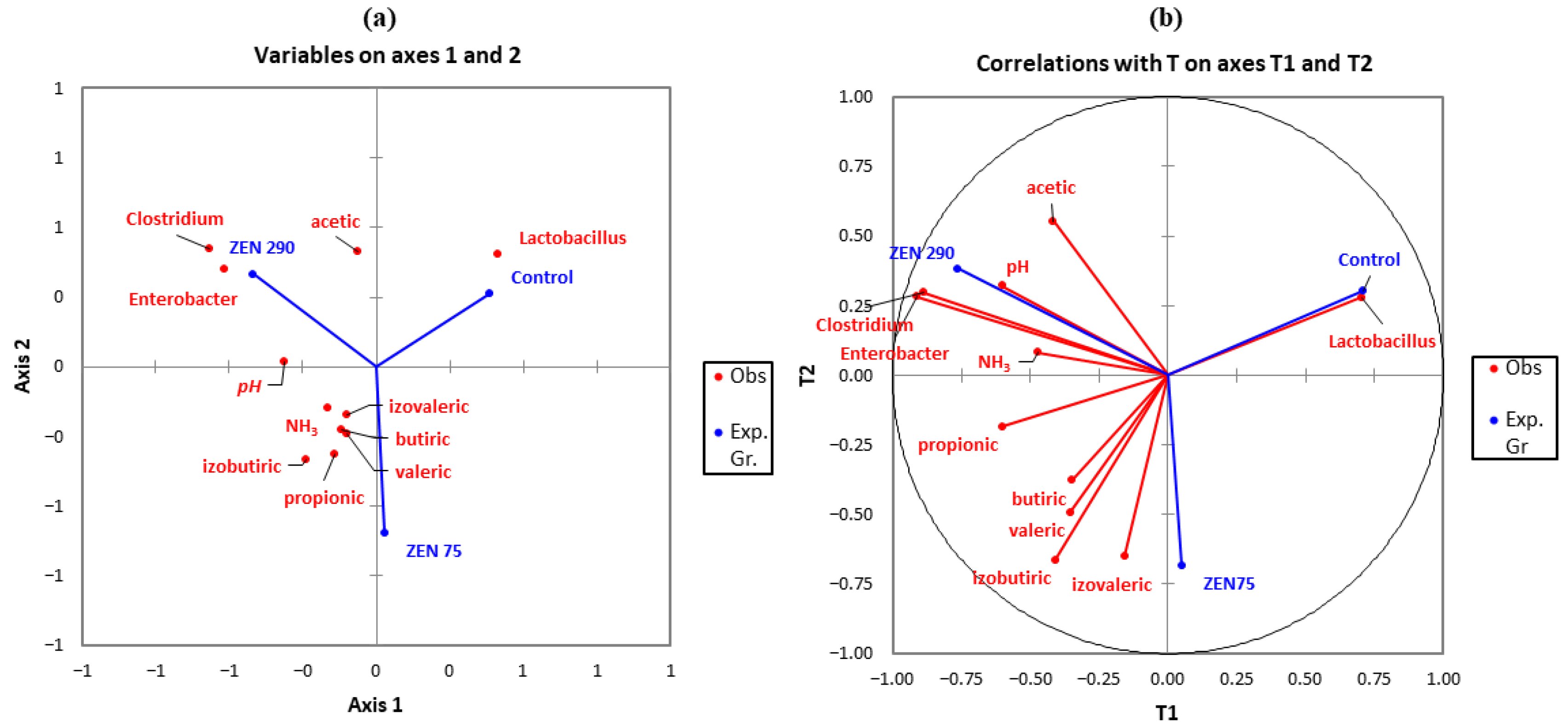
2.4. Correlations between SCFAs and Selected Microbial Populations
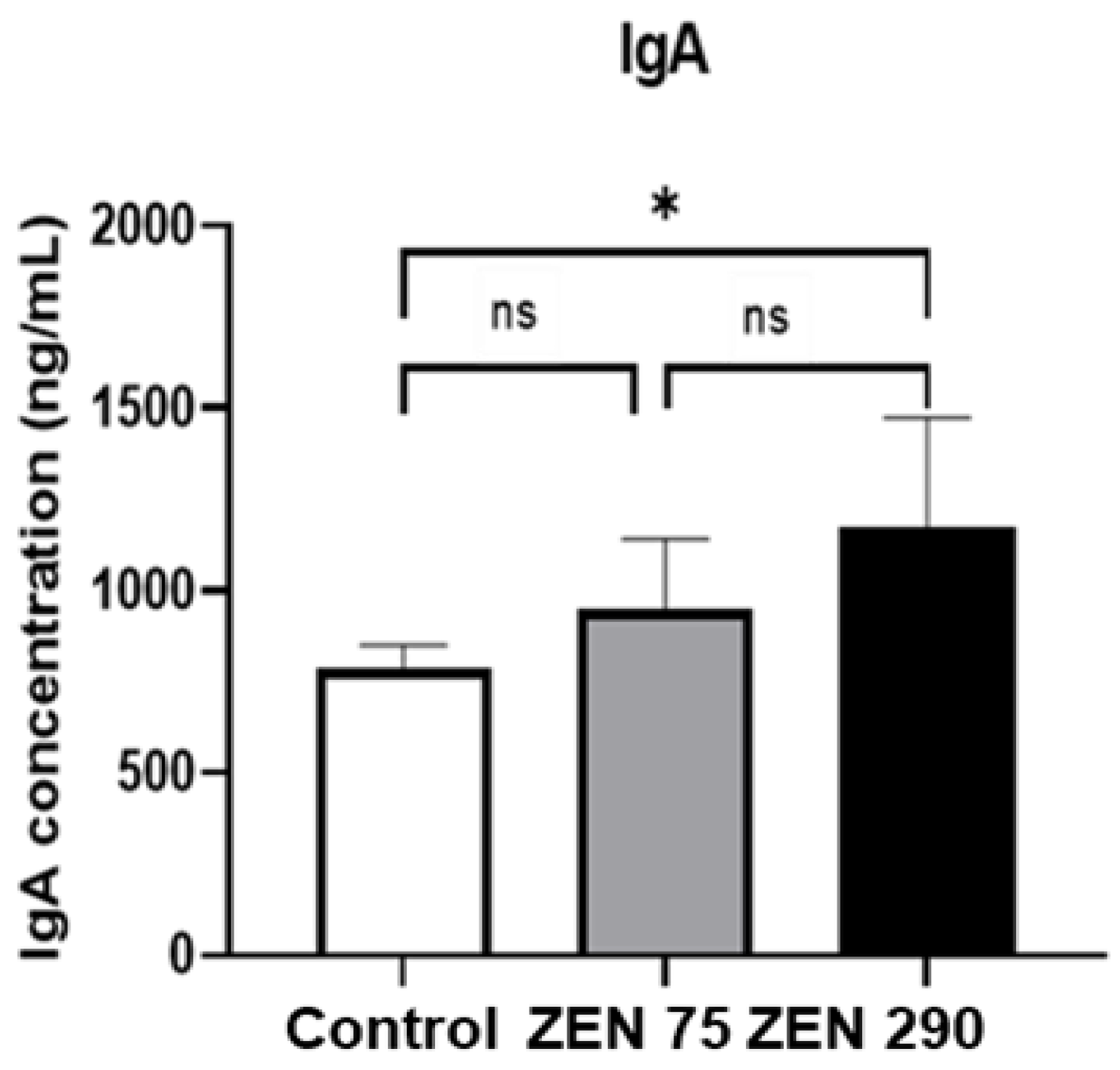
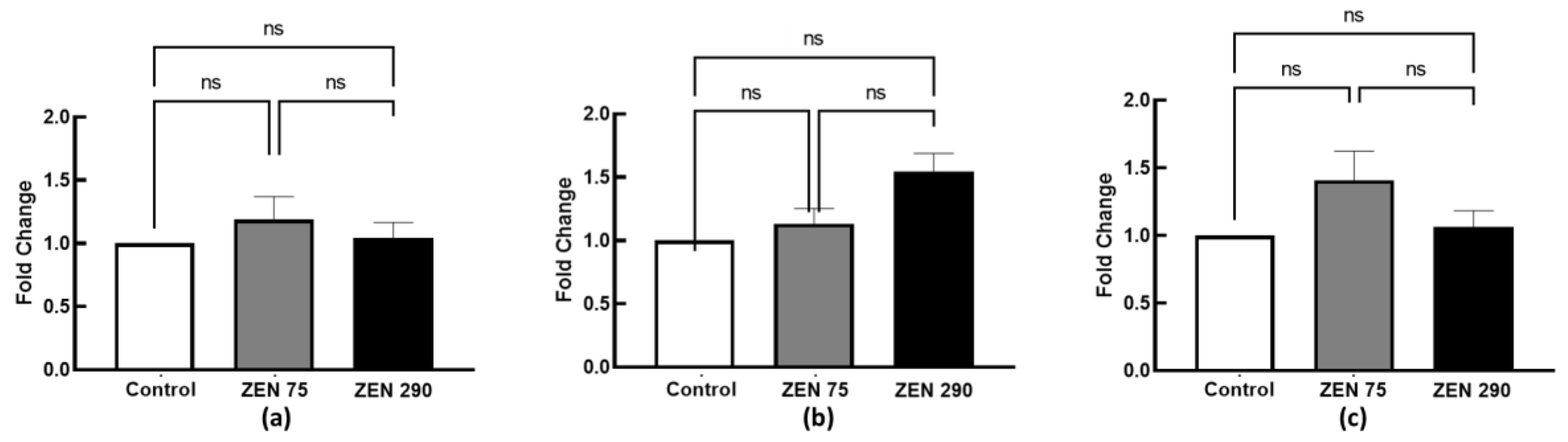
2.5. Effect of Zearalenone on Secretory IgA Synthesis
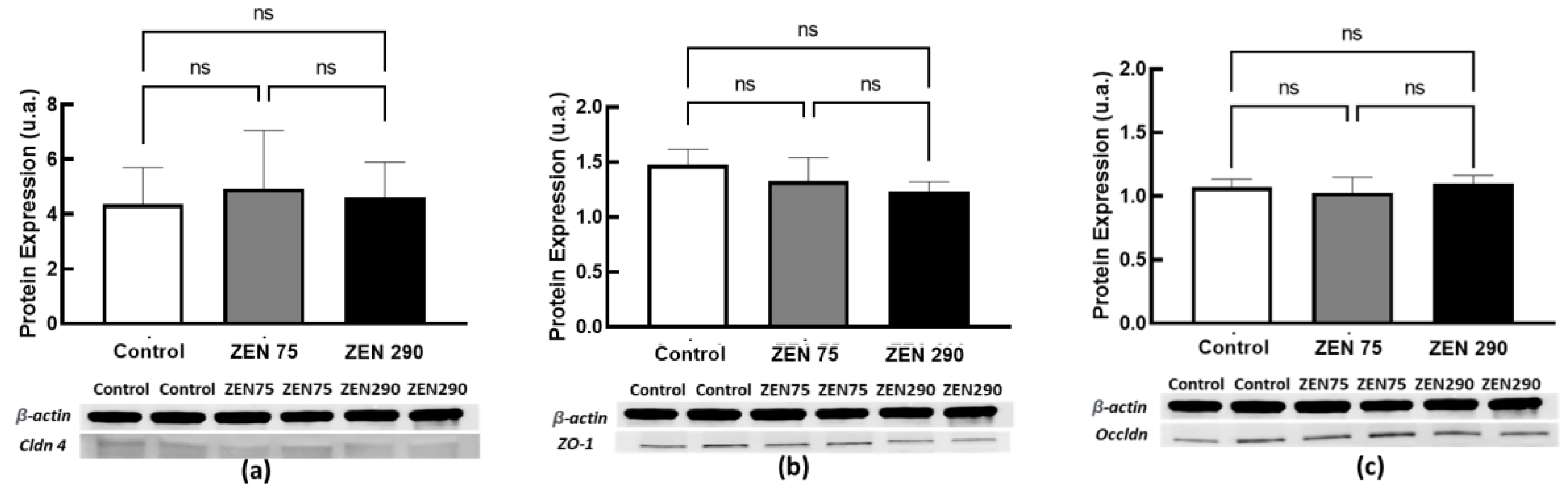
2.6. The Effect of Dietary Zearalenone on Tight Junction Proteins
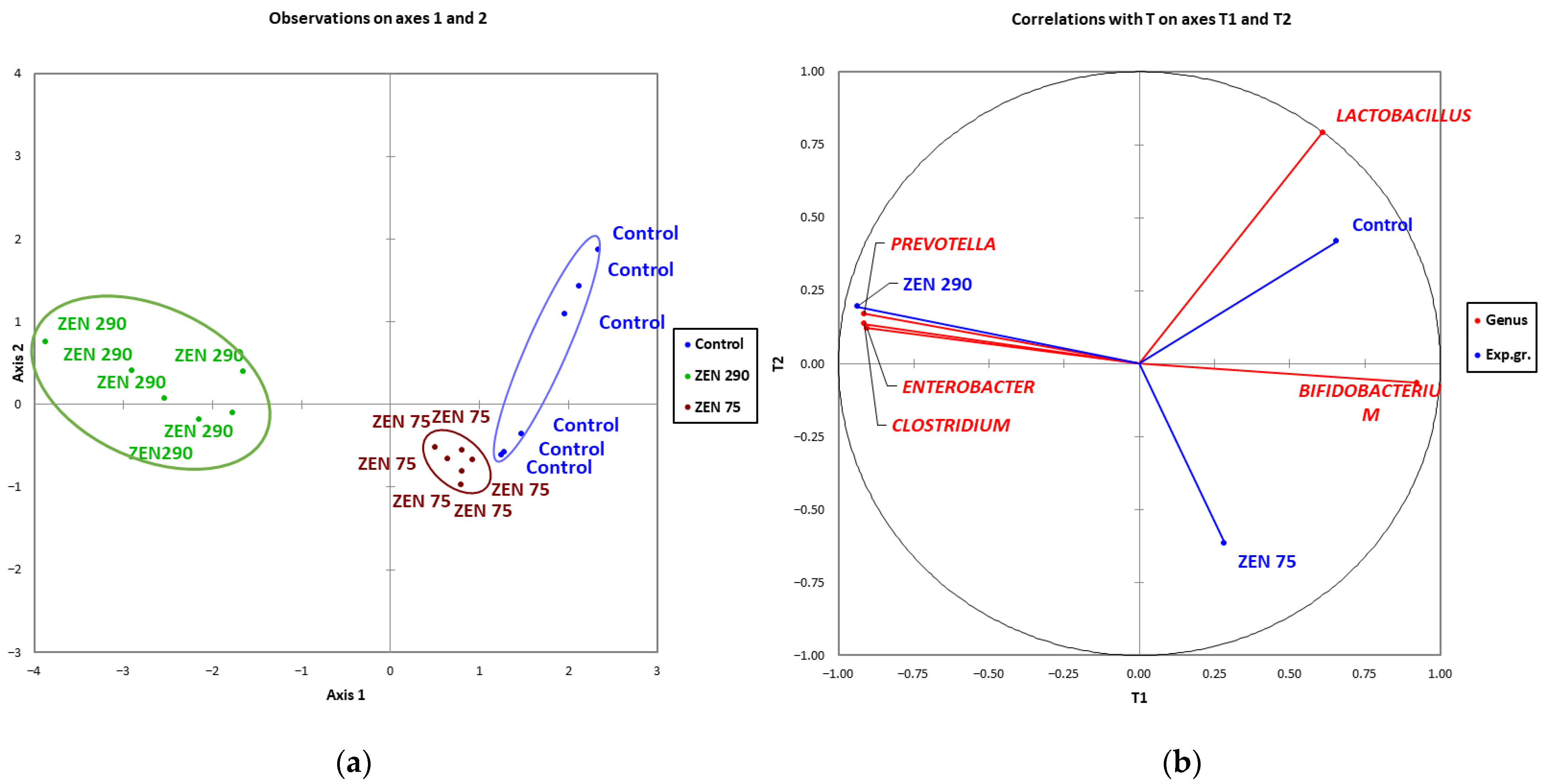
2.7. Microbial Community Clustering and Relationship between
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experimental Design
5.2. Toxins
5.3. Microbial DNA Extraction
5.4. Determination of SCFAs, Ammonia Content and pH Value
5.5. Quantification of Gene Expression by qPCR
5.6. Detection of Mucosal IgA in the Colonic Content
5.7. Immunoblotting Analysis
5.8. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, X.; Li, F.; Ning, H.; Zhang, W.; Niu, D.; Shi, Z.; Chai, S.; Shan, A. Screening of Pig-Derived Zearalenone-Degrading Bacteria through the Zearalenone Challenge Model, and Their Degradation Characteristics. Toxins 2022, 14, 224. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.E.; Jeong, J.Y.; Song, J.; Lee, Y.; Lee, H.-J.; Kim, D.-W.; Jung, H.J.; Kim, K.H.; Kim, M.; Oh, Y.K. Colon microbiome of pigs fed diet contaminated with commercial purified deoxynivalenol and zearalenone. Toxins 2018, 10, 347. [Google Scholar] [CrossRef] [Green Version]
- Le Sciellour, M.; Zemb, O.; Serviento, A.-M.; Renaudeau, D. Transient effect of single or repeated acute deoxynivalenol and zearalenone dietary challenge on fecal microbiota composition in female finishing pigs. Animal 2020, 14, 2277–2287. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Ren, C.; Yang, P.; Qi, D. Effects of Intestinal Microorganisms on Metabolism and Toxicity Mitigation of Zearalenone in Broilers. Animals 2022, 12, 1962. [Google Scholar] [CrossRef]
- Chen, X.; Yang, C.; Huang, L.; Niu, Q.; Jiang, S.; Chi, F. Zearalenone altered the serum hormones, morphologic and apoptotic measurements of genital organs in post-weaning gilts. Asian-Australas. J. Anim. Sci. 2015, 28, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Zhou, H.; Guo, T.; Zhang, Y.; Ma, L. Integrated multi-spectroscopic and molecular modeling techniques to study the formation mechanism of hidden zearalenone in maize. Food Chem. 2021, 351, 129286. [Google Scholar] [CrossRef]
- Cieplińska, K.; Gajęcka, M.; Dąbrowski, M.; Rykaczewska, A.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Zielonka, Ł.; Gajęcki, M.T. Time-dependent changes in the intestinal microbiome of gilts exposed to low zearalenone doses. Toxins 2019, 11, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, S.-H.; Koh, S.-E.; Oh, Y.; Cho, H.-S. Exposure to low concentrations of mycotoxins triggers unique responses from the pig gut microbiome. Korean J. Vet. Serv. 2020, 43, 39–44. [Google Scholar]
- Wang, T.; Ye, Y.; Ji, J.; Yang, X.; Xu, J.; Wang, J.-S.; Han, X.; Zhang, T.; Sun, X. Diet composition affects long-term zearalenone exposure on the gut–blood–liver axis metabolic dysfunction in mice. Ecotoxicol. Environ. Saf. 2022, 236, 113466. [Google Scholar] [CrossRef]
- Taranu, I.; Braicu, C.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Balacescu, L.; Neagoe, I.B.; Burlacu, R. Exposure to zearalenone mycotoxin alters in vitro porcine intestinal epithelial cells by differential gene expression. Toxicol. Lett. 2015, 232, 310–325. [Google Scholar] [CrossRef]
- Marin, D.E.; Motiu, M.; Taranu, I. Food contaminant zearalenone and its metabolites affect cytokine synthesis and intestinal epithelial integrity of porcine cells. Toxins 2015, 7, 1979–1988. [Google Scholar] [CrossRef]
- Guerre, P. Mycotoxin and gut microbiota interactions. Toxins 2020, 12, 769. [Google Scholar] [CrossRef]
- Wang, S.; Fu, W.; Zhao, X.; Chang, X.; Liu, H.; Zhou, L.; Li, J.; Cheng, R.; Wu, X.; Li, X. Zearalenone disturbs the reproductive-immune axis in pigs: The role of gut microbial metabolites. Microbiome 2022, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Gao, R.; Meng, Q.; Zhang, Y.; Bi, C.; Shan, A. Toxic effects of maternal zearalenone exposure on intestinal oxidative stress, barrier function, immunological and morphological changes in rats. PLoS ONE 2014, 9, e106412. [Google Scholar] [CrossRef] [Green Version]
- Malekinejad, H.; Maas-Bakker, R.; Fink-Gremmels, J. Species differences in the hepatic biotransformation of zearalenone. Vet. J. 2006, 172, 96–102. [Google Scholar] [CrossRef]
- Hartinger, T.; Grabher, L.; Pacífico, C.; Angelmayr, B.; Faas, J.; Zebeli, Q. Short-term exposure to the mycotoxins zearalenone or fumonisins affects rumen fermentation and microbiota, and health variables in cattle. Food Chem. Toxicol. 2022, 162, 112900. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J.; Malekinejad, H. Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed. Sci. Technol. 2007, 137, 326–341. [Google Scholar] [CrossRef]
- Hu, C.; Xiao, K.; Luan, Z.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taranu, I.; Arghir, A.; Pistol, G.; Marin, D.; Niculescu, A.-M. Effect of Fusarium mycotoxin Zearalenone on gut epithelium. Bull. UASVM Anim. Sci. Biotechnol. 2014, 71, 2. [Google Scholar] [CrossRef] [Green Version]
- Bulgaru, C.V.; Marin, D.E.; Pistol, G.C.; Taranu, I. Zearalenone and the immune response. Toxins 2021, 13, 248. [Google Scholar] [CrossRef]
- Malekinejad, H.; Maas-Bakker, R.; Fink-Gremmels, J. Bioactivation of zearalenone by porcine hepatic biotransformation. Vet. Res. 2005, 36, 799–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malekinejad, H.; Colenbrander, B.; Fink-Gremmels, J. Hydroxysteroid dehydrogenases in bovine and porcine granulosa cells convert zearalenone into its hydroxylated metabolites α-zearalenol and β-zearalenol. Vet. Res. Commun. 2006, 30, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Chang, J.; Wang, P.; Liu, C.; Yin, Q.; Song, A.; Gao, T.; Dang, X.; Lu, F. Effect of compound probiotics and mycotoxin degradation enzymes on alleviating cytotoxicity of swine jejunal epithelial cells induced by aflatoxin B1 and zearalenone. Toxins 2019, 11, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Qin, G.; Sun, Z.; Che, D.; Bao, N.; Zhang, X. Effects of soybean agglutinin on intestinal barrier permeability and tight junction protein expression in weaned piglets. Int. J. Mol. Sci. 2011, 12, 8502–8512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, W.; Lee, Y.-K.; Xie, J.; Zhang, H. Spatial heterogeneity and co-occurrence of mucosal and luminal microbiome across swine intestinal tract. Front. Microbiol. 2018, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Wang, J.; Gu, A.; Wang, T.; Li, J.; Shan, A. Zearalenone-Induced Mechanical Damage of Intestinal Barrier via the RhoA/ROCK Signaling Pathway in IPEC-J2 Cells. Int. J. Mol. Sci. 2022, 23, 12550. [Google Scholar] [CrossRef]
- Zhang, P.; Jing, C.; Liang, M.; Jiang, S.; Huang, L.; Jiao, N.; Li, Y.; Yang, W. Zearalenone Exposure Triggered Cecal Physical Barrier Injury through the TGF-β1/Smads Signaling Pathway in Weaned Piglets. Toxins 2021, 13, 902. [Google Scholar] [CrossRef]
- Skiepko, N.; Przybylska-Gornowicz, B.; Gajęcka, M.; Gajęcki, M.; Lewczuk, B. Effects of deoxynivalenol and zearalenone on the histology and ultrastructure of pig liver. Toxins 2020, 12, 463. [Google Scholar] [CrossRef]
- Zheng, W.; Ji, X.; Zhang, Q.; Yao, W. Intestinal microbiota ecological response to oral administrations of hydrogen-rich water and lactulose in female piglets fed a Fusarium toxin-contaminated diet. Toxins 2018, 10, 246. [Google Scholar] [CrossRef] [Green Version]
- Przybylska-Gornowicz, B.; Tarasiuk, M.; Lewczuk, B.; Prusik, M.; Ziółkowska, N.; Zielonka, Ł.; Gajęcki, M.; Gajęcka, M. The effects of low doses of two Fusarium toxins, zearalenone and deoxynivalenol, on the pig jejunum. A light and electron microscopic study. Toxins 2015, 7, 4684–4705. [Google Scholar] [CrossRef] [Green Version]
- Jia, R.; Liu, W.; Zhao, L.; Cao, L.; Shen, Z. Low doses of individual and combined deoxynivalenol and zearalenone in naturally moldy diets impair intestinal functions via inducing inflammation and disrupting epithelial barrier in the intestine of piglets. Toxicol. Lett. 2020, 333, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Taranu, I.; Nguyen, T.-T.; Pham, K.-D.; Gras, M.A.; Pistol, G.C.; Marin, D.E.; Rotar, C.; Habeanu, M.; Ho, P.-H.; Le, T.-M. Rice and cassava distillers dried grains in Vietnam: Nutritional values and effects of their dietary inclusion on blood chemical parameters and immune responses of growing pigs. Waste Biomass Valorization 2019, 10, 3373–3382. [Google Scholar] [CrossRef]
- Chethan, G.E.; Garkhal, J.; Sircar, S.; Malik, Y.P.S.; Mukherjee, R.; Sahoo, N.R.; Agarwal, R.K.; De, U.K. Immunomodulatory potential of β-glucan as supportive treatment in porcine rotavirus enteritis. Vet. Immunol. Immunopathol. 2017, 191, 36–43. [Google Scholar] [CrossRef]
- Tran, T.H.T.; Everaert, N.; Bindelle, J. Review on the effects of potential prebiotics on controlling intestinal enteropathogens Salmonella and Escherichia coli in pig production. J. Anim. Physiol. Anim. Nutr. 2018, 102, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Piotrowska, M.; Śliżewska, K.; Nowak, A.; Zielonka, Ł.; Żakowska, Z.; Gajęcka, M.; Gajęcki, M. The effect of experimental fusarium mycotoxicosis on microbiota diversity in porcine ascending colon contents. Toxins 2014, 6, 2064–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Lu, N.; Zhao, H.; Yuan, H.; Xia, D.; Lei, H. The microbiome–metabolome response in the colon of piglets under the status of weaning stress. Front. Microbiol. 2020, 11, 2055. [Google Scholar] [CrossRef]
- Zeng, Y.; Wang, Z.; Zou, T.; Chen, J.; Li, G.; Zheng, L.; Li, S.; You, J. Bacteriophage as an alternative to antibiotics promotes growth performance by regulating intestinal inflammation, intestinal barrier function and gut microbiota in weaned piglets. Front. Vet. Sci. 2021, 8, 623899. [Google Scholar] [CrossRef] [PubMed]
- Holman, D.B.; Brunelle, B.W.; Trachsel, J.; Allen, H.K. Meta-analysis to define a core microbiota in the swine gut. MSystems 2017, 2, e00004-17. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Ren, W.; Smidt, H.; Wright, A.-D.G.; Yu, B.; Schyns, G.; McCormack, U.M.; Cowieson, A.J.; Yu, J.; He, J. Dynamic distribution of gut microbiota in pigs at different growth stages: Composition and contribution. Microbiol. Spectr. 2022, 10, e00688-621. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xiao, Y.; Wang, J.; Xiang, Y.; Gong, Y.; Wen, X.; Li, D. Core gut microbiota in Jinhua pigs and its correlation with strain, farm and weaning age. J. Microbiol. 2018, 56, 346–355. [Google Scholar] [CrossRef]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut microbiota dysbiosis in postweaning piglets: Understanding the keys to health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Y.I.; Zhou, T.; Bullerman, L.B. Sourdough lactic acid bacteria as antifungal and mycotoxin-controlling agents. Food Sci. Technol. Int. 2016, 22, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Yang, S.; Zhang, X.; Huang, S.; Wang, N.; Wang, M.; Long, M.; He, J. Zearalenone changes the diversity and composition of caecum microbiota in weaned rabbit. BioMed Research International 2018, 2018, 3623274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.; Ge, W.; Wang, J.; Liu, W.; Zhao, Y.; Shen, W.; Li, L. Zearalenone-induced aberration in the composition of the gut microbiome and function impacts the ovary reserve. Chemosphere 2020, 244, 125493. [Google Scholar] [CrossRef]
- Saenz, J.S.; Kurz, A.; Ruczizka, U.; Bünger, M.; Dippel, M.; Nagl, V.; Grenier, B.; Ladinig, A.; Seifert, J.; Selberherr, E. Metaproteomics Reveals Alteration of the Gut Microbiome in Weaned Piglets Due to the Ingestion of the Mycotoxins Deoxynivalenol and Zearalenone. Toxins 2021, 13, 583. [Google Scholar] [CrossRef] [PubMed]
- Riwes, M.; Reddy, P. Short chain fatty acids: Postbiotics/metabolites and graft versus host disease colitis. Semin. Hematol. 2020, 57, 1–6. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Lee, J.H.; Lee, S.H.; Seok, M.-J.; Kim, D.W.; Kang, B.N.; Johnson, T.J.; Isaacson, R.E.; Kim, H.B. Piglet gut microbial shifts early in life: Causes and effects. J. Anim. Sci. Biotechnol. 2019, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Hurst, N.R.; Kendig, D.M.; Murthy, K.S.; Grider, J.R. The short chain fatty acids, butyrate and propionate, have differential effects on the motility of the guinea pig colon. Neurogastroenterol. Motil. 2014, 26, 1586–1596. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y. Fatty acids, inflammation and intestinal health in pigs. J. Anim. Sci. Biotechnol. 2015, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Bernad-Roche, M.; Bellés, A.; Grasa, L.; Casanova-Higes, A.; Mainar-Jaime, R.C. Effects of Dietary Supplementation with Protected Sodium Butyrate on Gut Microbiota in Growing-Finishing Pigs. Animals 2021, 11, 2137. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Zhu, D.; Guo, T.; Zhang, Y.; Shi, B.; Shan, A.; Chen, Z. Toxicity of zearalenone on the intestines of pregnant sows and their offspring and alleviation with modified halloysite nanotubes. J. Sci. Food Agric. 2018, 98, 698–706. [Google Scholar] [CrossRef]
- Liu, G.; Yan, T.; Wang, J.; Huang, Z.; Chen, X.; Jia, G.; Wu, C.; Zhao, H.; Xue, B.; Xiao, L. Biological system responses to zearalenone mycotoxin exposure by integrated metabolomic studies. J. Agric. Food Chem. 2013, 61, 11212–11221. [Google Scholar] [CrossRef]
- Henze, L.J.; Koehl, N.J.; Bennett-Lenane, H.; Holm, R.; Grimm, M.; Schneider, F.; Weitschies, W.; Koziolek, M.; Griffin, B.T. Characterization of gastrointestinal transit and luminal conditions in pigs using a telemetric motility capsule. Eur. J. Pharm. Sci. 2021, 156, 105627. [Google Scholar] [CrossRef]
- Pieper, R.; Tudela, C.V.; Taciak, M.; Bindelle, J.; Pérez, J.F.; Zentek, J. Health relevance of intestinal protein fermentation in young pigs. Anim. Health Res. Rev. 2016, 17, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Holzer, P. Acid-sensing ion channels in gastrointestinal function. Neuropharmacology 2015, 94, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K.; Aschenbach, J.R. Ureases in the gastrointestinal tracts of ruminant and monogastric animals and their implication in urea-N/ammonia metabolism: A review. J. Adv. Res. 2018, 13, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Mansilla, W.D.; Silva, K.E.; Zhu, C.L.; Nyachoti, C.M.; Htoo, J.K.; Cant, J.P.; de Lange, C.F. Ammonia nitrogen added to diets deficient in dispensable amino acid nitrogen is poorly utilized for urea production in growing pigs. J. Nutr. 2017, 147, 2228–2234. [Google Scholar] [CrossRef] [Green Version]
- Xi, P.; Jiang, Z.; Dai, Z.; Li, X.; Yao, K.; Zheng, C.; Lin, Y.; Wang, J.; Wu, G. Regulation of protein turnover by L-glutamine in porcine intestinal epithelial cells. J. Nutr. Biochem. 2012, 23, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Rist, V.; Weiss, E.; Eklund, M.; Mosenthin, R. Impact of dietary protein on microbiota composition and activity in the gastrointestinal tract of piglets in relation to gut health: A review. Animal 2013, 7, 1067–1078. [Google Scholar] [CrossRef] [Green Version]
- Metzler-Zebeli, B.U.; Mann, E.; Schmitz-Esser, S.; Wagner, M.; Ritzmann, M.; Zebeli, Q. Changing dietary calcium-phosphorus level and cereal source selectively alters abundance of bacteria and metabolites in the upper gastrointestinal tracts of weaned pigs. Appl. Environ. Microbiol. 2013, 79, 7264–7272. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Liu, H.; Wang, S.; Zhang, W.; Wang, J.; Tian, H.; Wang, Y.; Ji, H. Fecal microbiota and its correlation with fatty acids and free amino acids metabolism in piglets after a Lactobacillus strain oral administration. Front. Microbiol. 2019, 10, 785. [Google Scholar] [CrossRef]
- Hutabarat, D.J.C.; Zakaria, F.R.; Purwani, E.Y.; Suhartono, M.T. SCFA profile of rice RS fermentation by colonic microbiota, Clostridium butyricum BCC B2571, and Eubacterium rectale DSM 17629. Adv. Biosci. Biotechnol. 2018, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Barszcz, M.; Taciak, M.; Tuśnio, A.; Święch, E.; Skomiał, J.; Čobanová, K.; Grešáková, Ľ. The effect of organic and inorganic zinc source, used with lignocellulose or potato fiber, on microbiota composition, fermentation, and activity of enzymes involved in dietary fiber breakdown in the large intestine of pigs. Livest. Sci. 2021, 245, 104429. [Google Scholar] [CrossRef]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogere, P.; Choi, Y.; Heo, J. Probiotics as alternatives to antibiotics in treating post-weaning diarrhoea in pigs. S. Afr. J. Anim. Sci. 2019, 49, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Canibe, N.; Højberg, O.; Kongsted, H.; Vodolazska, D.; Lauridsen, C.; Nielsen, T.S.; Schönherz, A.A. Review on Preventive Measures to Reduce Post-Weaning Diarrhoea in Piglets. Animals 2022, 12, 2585. [Google Scholar] [CrossRef] [PubMed]
- Dell’Anno, M.; Callegari, M.L.; Reggi, S.; Caprarulo, V.; Giromini, C.; Spalletta, A.; Coranelli, S.; Sgoifo Rossi, C.A.; Rossi, L. Lactobacillus plantarum and Lactobacillus reuteri as Functional Feed Additives to Prevent Diarrhoea in Weaned Piglets. Animals 2021, 11, 1766. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, Q.; Qi, R.; Wang, J.; Qiu, X.; Liu, Z.; Huang, J. Effects of Lactobacillus plantarum on the intestinal morphology, intestinal barrier function and microbiota composition of suckling piglets. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1908–1918. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Koo, B.; Nyachoti, M.; Columbus, D.A. Formulating Diets for Improved Health Status of Pigs: Current Knowledge and Perspectives. Animals 2022, 12, 2877. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, S.; Tsiloyiannis, V.; Vlemmas, J.; Sarris, K.; Tsinas, A.; Alexopoulos, C.; Jansegers, L. The effect of probiotic LSP 122 on the control of post-weaning diarrhoea syndrome of piglets. Res. Vet. Sci. 1999, 67, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jin, L.; Chen, T. The effects of secretory IgA in the mucosal immune system. BioMed Res. Int. 2020, 2020, 2032057. [Google Scholar] [CrossRef] [Green Version]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yu, H.; Shan, A.; Jin, Y.; Fang, H.; Zhao, Y.; Shen, J.; Zhou, C.; Zhou, Y.; Fu, Y. Toxic effects of Zearalenone on intestinal microflora and intestinal mucosal immunity in mice. Food Agric. Immunol. 2018, 29, 1002–1011. [Google Scholar] [CrossRef]
- León, E.D.; Francino, M.P. Roles of secretory immunoglobulin A in host-microbiota interactions in the gut ecosystem. Front. Microbiol. 2022, 13, 880484. [Google Scholar] [CrossRef]
- Yang, C.; Mogno, I.; Contijoch, E.J.; Borgerding, J.N.; Aggarwala, V.; Li, Z.; Siu, S.; Grasset, E.K.; Helmus, D.S.; Dubinsky, M.C. Fecal IgA levels are determined by strain-level differences in Bacteroides ovatus and are modifiable by gut microbiota manipulation. Cell Host Microbe 2020, 27, 467–475.e466. [Google Scholar] [CrossRef] [Green Version]
- Pietrzak, B.; Tomela, K.; Olejnik-Schmidt, A.; Mackiewicz, A.; Schmidt, M. Secretory IgA in intestinal mucosal secretions as an adaptive barrier against microbial cells. Int. J. Mol. Sci. 2020, 21, 9254. [Google Scholar] [CrossRef]
- Marin, D.E.; Anghel, A.C.; Bulgaru, C.V.; Grosu, I.; Pistol, G.C.; Cismileanu, A.E.; Taranu, I. The Use of Agro-Industrial Waste Rich in Omega-3 PUFA during the Weaning Stress Improves the Gut Health of Weaned Piglets. Agriculture 2022, 12, 1142. [Google Scholar] [CrossRef]
- Grosu, I.A.; Cismileanu, A.E.; Marin, D.E.; Taranu, I. The effects of a low dose OTA exposure on weanling piglet gut microbiota. Arch. Zootech. 2021, 24, 31–43. [Google Scholar] [CrossRef]
- Cişmileanu, A.; Oancea, A.; Dragomir, C. Effect of 2 Types of Yeast on Rumen Fermentation in Carpatina Crossbred Goats-Short Term Study. Sci. Pap. Anim. Sci. Biotechnol./Lucr. Stiintifice Zooteh. Si Biotehnol. 2022, 55, 13–18. [Google Scholar]
- Sivieri, K.; Morales, M.L.V.; Adorno, M.A.T.; Sakamoto, I.K.; Saad, S.M.I.; Rossi, E.A. Lactobacillus acidophilus CRL 1014 improved “gut health” in the SHIME® reactor. BMC Gastroenterol. 2013, 13, 100. [Google Scholar] [CrossRef] [PubMed]
- Pistol, G.C.; Bulgaru, C.V.; Marin, D.E.; Oancea, A.G.; Taranu, I. Dietary Grape Seed Meal Bioactive Compounds Alleviate Epithelial Dysfunctions and Attenuates Inflammation in Colon of DSS-Treated Piglets. Foods 2021, 10, 530. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters * | Control | ZEN 75 | ZEN 290 | p Value |
|---|---|---|---|---|
| Initial BW (kg) | 11.7 ± 0.9 | 11 ± 1 | 11 ± 1.4 | 0.577 |
| Final BW (kg) | 21.5 ± 2.8 | 19.6 ± 3.5 | 19.5 ± 1 | 0.465 |
| Average Daily Gain (kg) | 0.42 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.656 |
| Average Daily Feed Intake (kg) | 1.1 ± 0.2 | 1.0 ± 0.2 | 1.0 ± 0.2 | 0.177 |
| Feed Efficiency (kg/kg) | 2.6 ± 0.2 | 2.7 ± 0.3 | 2.8 ± 0.3 | 0.988 |
| SCFA (mM/L) /Experimental Group * | Acetic Acid (mM/L) | Propionic Acid (mM/L) | Isobutyric Acid (mM/L) | Butyric Acid (mM/L) | Isovaleric Acid (mM/L) | Valeric Acid (mM/L) | Total SCFAs (mM/L) | pH | NH3 (µM/g) |
|---|---|---|---|---|---|---|---|---|---|
| Control | 22.58 ± 0.6 | 7.45 ± 0.4 | 0.67 ± 0.1 | 4.35 ± 0.3 | 0.70 ± 0.1 | 1.14 ± 0.1 | 38.67 ± 1.3 | 6.69 ± 0.0 | 11.10 ± 0.9 |
| ZEN 75 | 19.50 ± 1.9 | 8.89 ± 0.8 | 0.90 ± 0.1 | 5.33 ± 0.8 | 0.86 ± 0.2 | 1.40 ± 0.3 | 39.46 ± 3.8 | 6.73 ± 0.0 | 12.70 ± 1.9 |
| ZEN 290 | 23.42 ± 2.6 | 8.60 ± 1.1 | 0.88 ± 0.1 | 5.15 ± 0.9 | 0.84 ± 0.1 | 1.33 ± 0.2 | 42.52 ± 4.4 | 6.78 ± 0.0 | 13.07 ± 0.7 |
| Experimental Group | SCFA/ Bacterial Genus | Acetic Acid | Propionic Acid | Isobutyric Acid | Butyric Acid | Isovaleric Acid | Valeric Acid |
|---|---|---|---|---|---|---|---|
| Control | Clostridium | 0.434 | 0.723 (p = 0.0288) | 0.553 | −0.015 | −0.444 | −0.438 |
| Prevotella | 0.469 | 0.378 | 0.361 | 0.317 | −0.096 | −0.074 | |
| Enterobacter | −0.371 | 0.538 | 0.426 | 0.250 | 0.727 | 0.724 | |
| Lactobacillus | −0.070 | −0.006 | 0.682 (p = 0.0399) | 0.075 | 0.422 | 0.724 (p = 0.0214) | |
| Bifidobacterium | 0.570 | 0.076 | −0.366 | 0.094 | −0.630 | −0.114 | |
| ZEN 75 | Clostridium | 0.470 | 0.430 | 0.520 | 0.451 | 0.515 | 0.313 |
| Prevotella | 0.410 | 0.680 (p = 0.0156) | 0.343 | 0.481 | 0.307 | 0.485 | |
| Enterobacter | 0.430 (p = 0.0439) | 0.435 (p = 0.0346) | 0.262 | 0.256 | 0.230 | 0.196 | |
| Lactobacillus | 0.244 | 0.223 | 0.074 | 0.755 (p = 0.0353) | 0.220 | 0.420 | |
| Bifidobacterium | 0.394 | 0.112 | 0.100 | 0.744 (p = 0.0180) | 0.744 (p = 0.0189) | 0.225 | |
| ZEN 290 | Clostridium | 0.370 | 0.581 | 0.265 | 0.120 | 0.246 | −0.482 |
| Prevotella | 0.480 | 0.620 | 0.277 | 0.650 | 0.400 | 0.574 | |
| Enterobacter | 0.630 (p = 0.0144) | 0.660 (p = 0.0416) | 0.149 | 0.140 | 0.157 | 0.193 | |
| Lactobacillus | −0.770 (p = 0.0267) | −0.446 | −0.410 | 0.652 (p = 0.0155) | −0.482 | 0.539 | |
| Bifidobacterium | −0.800 (p = 0.0124) | −0.640 (p = 0.0302) | −0.420 | 0.700 (p = 0.0244) | 0.295 | 0.380 |
| Ingredients (%) | Control | ZEN 75 | ZEN 290 |
|---|---|---|---|
| Corn | 68.46 | 68.46 | 68.46 |
| Soybean meal | 19.00 | 19.00 | 19.00 |
| Milk replacement | 5.00 | 5.00 | 5.00 |
| Corn gluten | 4.00 | 4.00 | 4.00 |
| l-Lysine | 0.31 | 0.31 | 0.31 |
| Methionine | 0.10 | 0.10 | 0.10 |
| CaCo3 | 1.57 | 1.57 | 1.57 |
| Ca(H2PO4)2 | 0.35 | 0.35 | 0.35 |
| NaCl | 0.10 | 0.10 | 0.10 |
| Mineral-Vitamin Premix | 1.00 | 1.00 | 1.00 |
| Choline Premix | 0.10 | 0.10 | 0.10 |
| Phytase | 0.01 | 0.01 | 0.01 |
| ZEN (mg/kg) | - | 75 | 290 |
| Metabolizable Energy (kcal/kg) | 3282.60 | 3282.60 | 3282.60 |
| Crude protein % | 18.70 | 18.70 | 18.70 |
| Lysine % | 1.20 | 1.20 | 1.20 |
| (Methionine + Cysteine) % | 0.72 | 0.72 | 0.72 |
| Ca % | 0.90 | 0.90 | 0.90 |
| P % | 0.72 | 0.72 | 0.72 |
| Fats % | 2.40 | 2.40 | 2.40 |
| Cellulose % | 4.24 | 4.24 | 4.24 |
| Dry Matter % | 89.44 | 89.44 | 89.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grosu, I.A.; Bulgaru, C.V.; Pistol, G.C.; Cismileanu, A.; Marin, D.E.; Taranu, I. Effects of Exposure to Low Zearalenone Concentrations Close to the EU Recommended Value on Weaned Piglets’ Colon. Toxins 2023, 15, 206. https://doi.org/10.3390/toxins15030206
Grosu IA, Bulgaru CV, Pistol GC, Cismileanu A, Marin DE, Taranu I. Effects of Exposure to Low Zearalenone Concentrations Close to the EU Recommended Value on Weaned Piglets’ Colon. Toxins. 2023; 15(3):206. https://doi.org/10.3390/toxins15030206
Chicago/Turabian StyleGrosu, Iulian Alexandru, Cristina Valeria Bulgaru, Gina Cecilia Pistol, Ana Cismileanu, Daniela Eliza Marin, and Ionelia Taranu. 2023. "Effects of Exposure to Low Zearalenone Concentrations Close to the EU Recommended Value on Weaned Piglets’ Colon" Toxins 15, no. 3: 206. https://doi.org/10.3390/toxins15030206