
Convolvulus pluricaulis Choisy’s Extraction, Chemical Characterization and Evaluation of the Potential Effects on Glycaemic Balance in a 3T3-L1 Adipocyte Cell Model

 , , , , ,
, , , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Convolvulus pluricaulis Extraction
2.3. Flash Chromatographic Separation
2.4. GC-MS and GC-FID Analyses
2.5. Cell Cultures and Induction of Adipocyte Differentiation
2.6. Extract and Fractions’ Dissolution for Cell Treatments
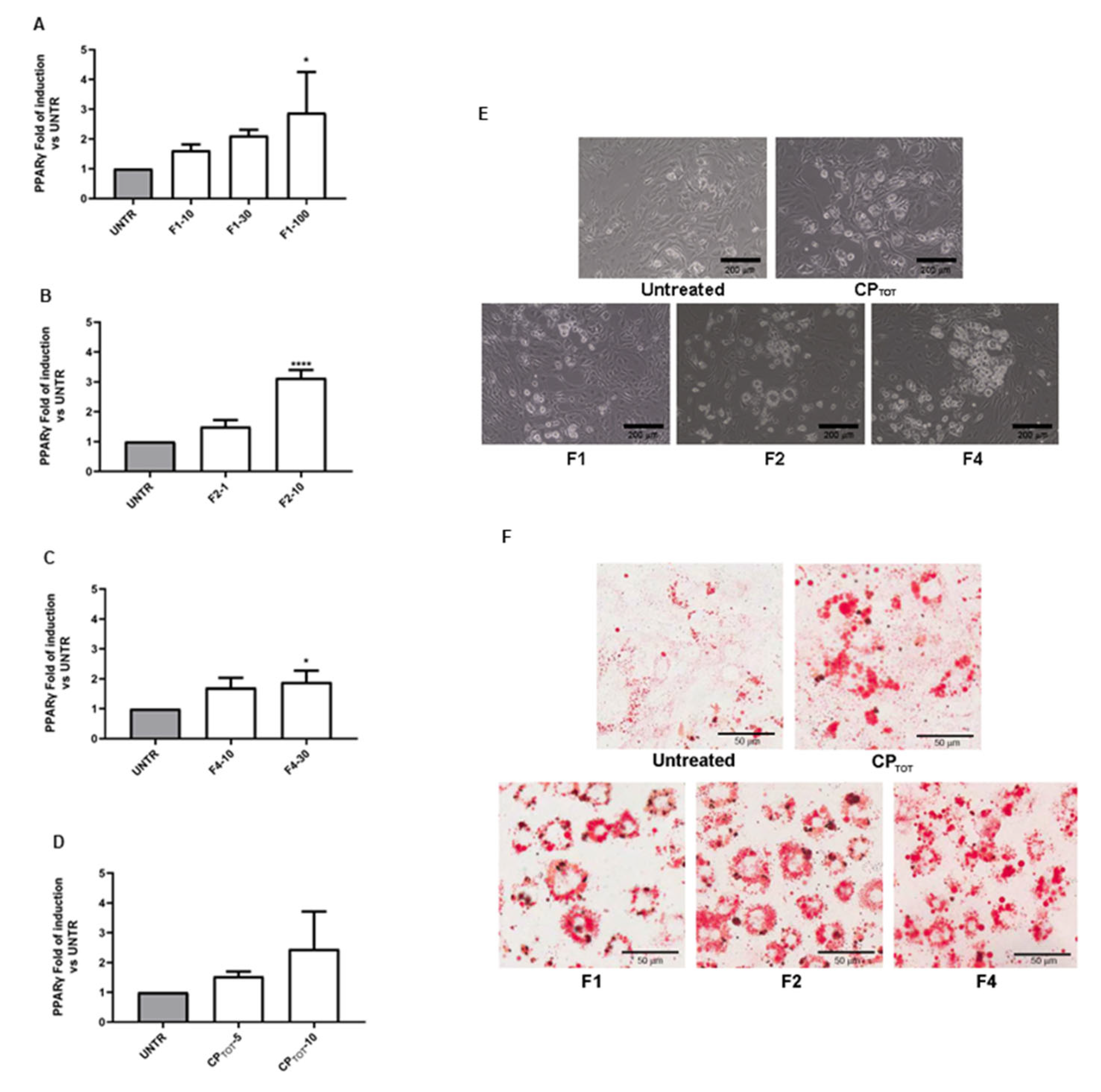
2.7. Morphological Analysis and Oil-Red O Staining
2.8. RT-PCR Analysis
2.9. Cytokines Analysis
2.10. Statistical Analysis
3. Results
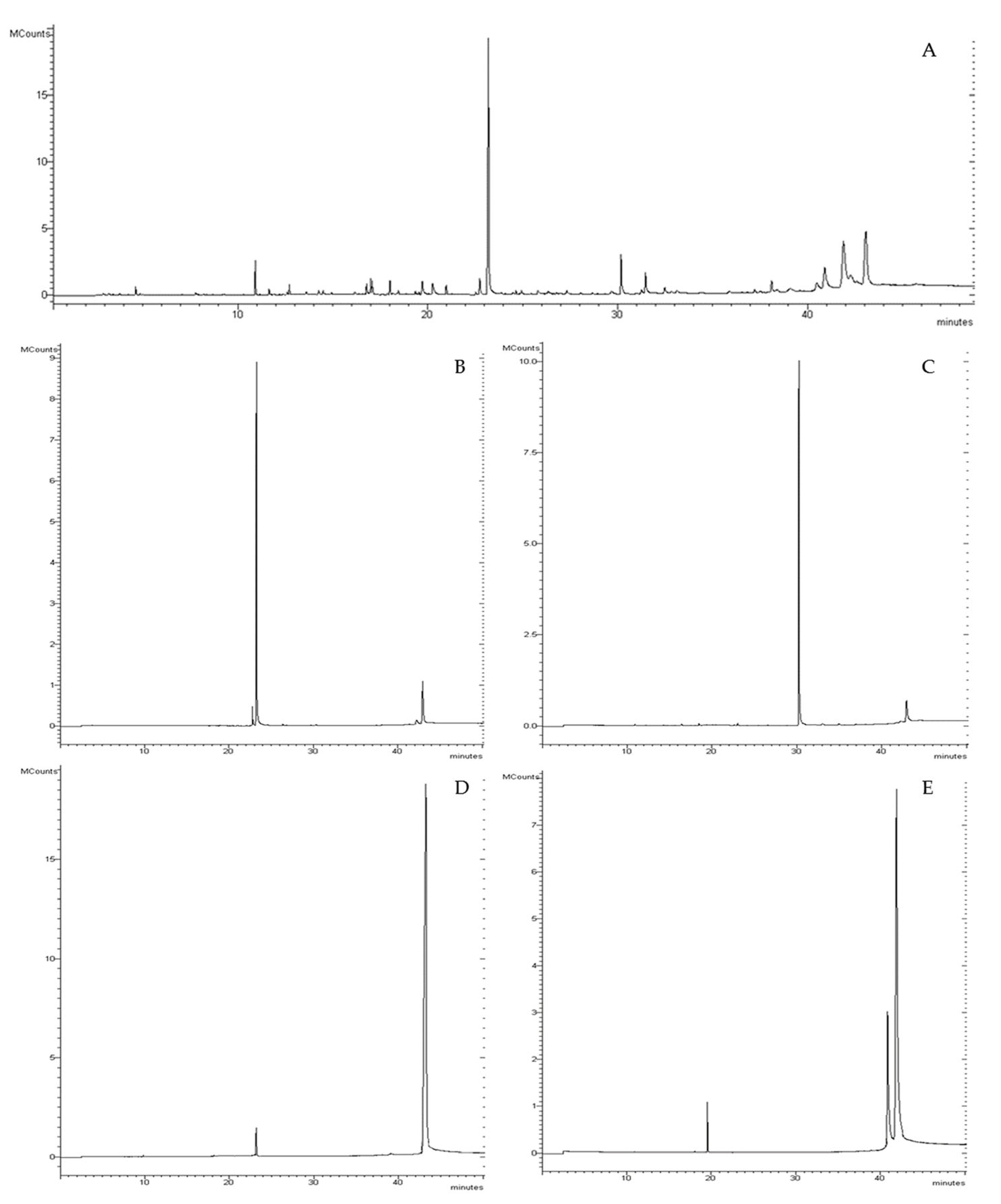
3.1. Convolvulus pluricaulis Fractions Chemical Characterization
3.2. Effects of Convolvulus pluricaulis Extract on 3T3-L1 Adipocyte Differentiation
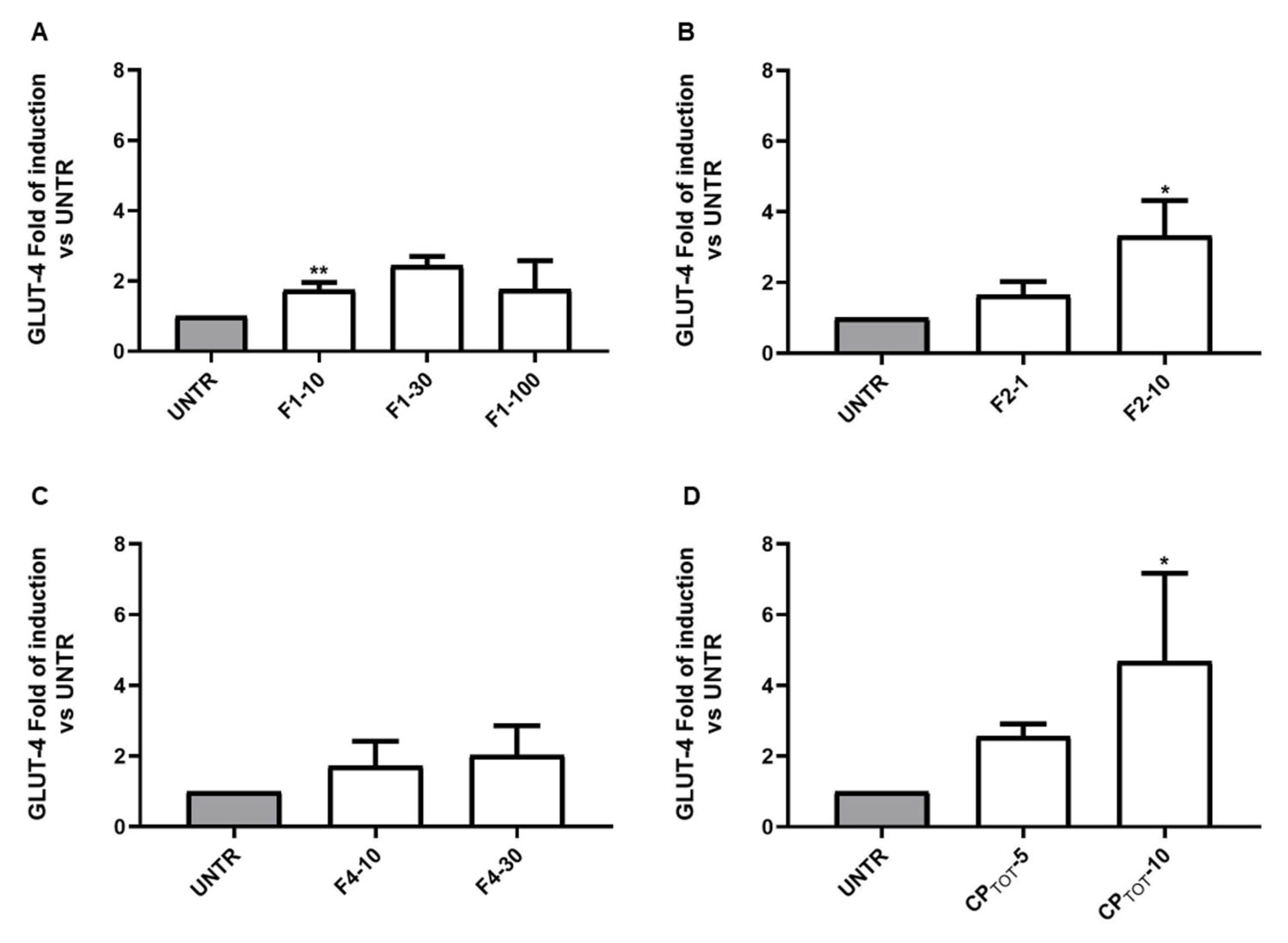
3.3. Convolvulus pluricaulis Fractions Induced a Differential Modulation on the Insulin-Inducible Gene GLUT-4
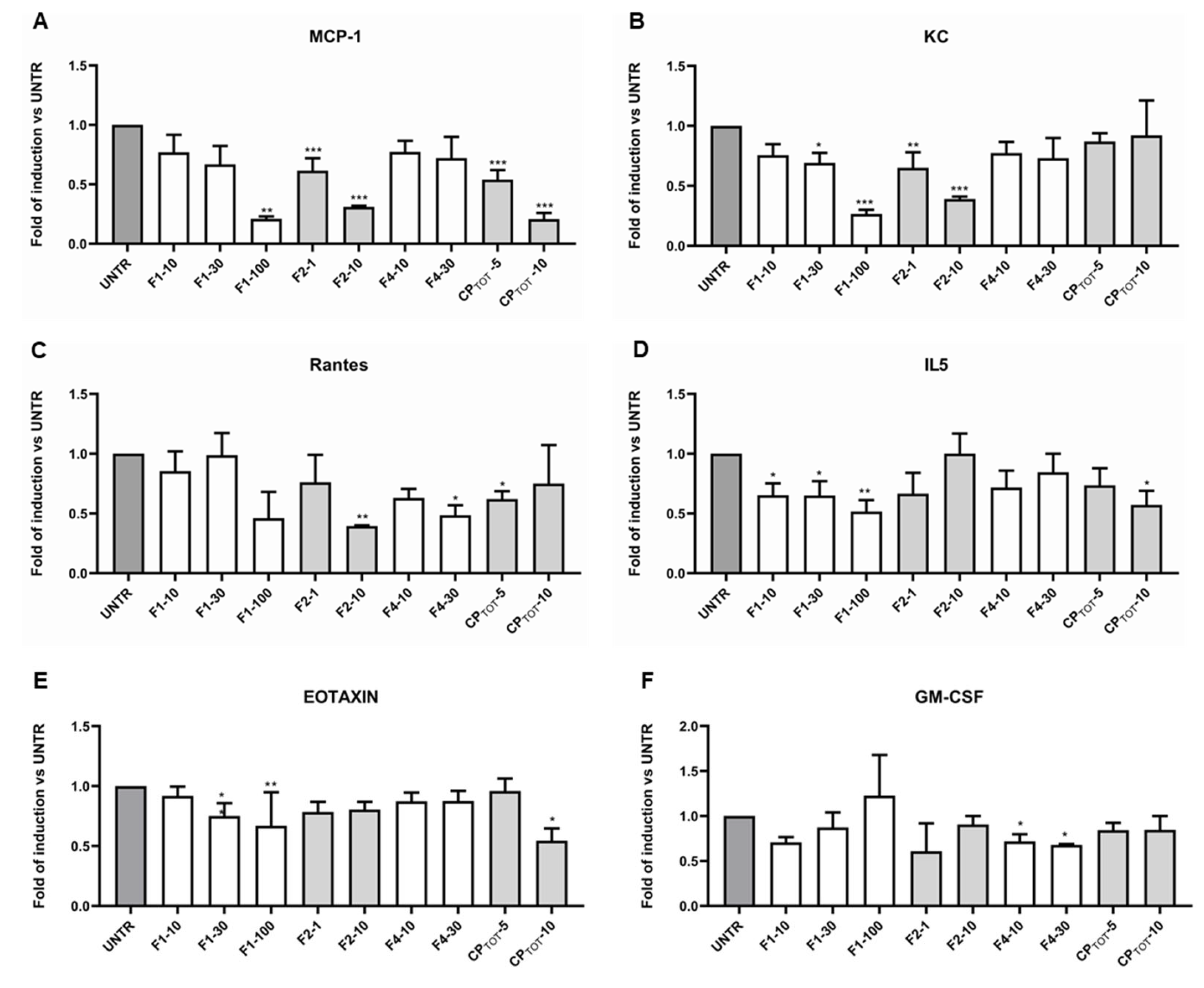
3.4. Convolvulus pluricaulis Extract Downmodulated the Release of Specific Inflammatory Cytokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Agarwa, P.; Sharma, B.; Fatima, A.; Jain, S.K. An update on Ayurvedic herb Convolvulus pluricaulis Choisy. Asian Pac. J. Trop. Biomed. 2014, 4, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, G.L.; Fernandes, J. Protective effect of Convolvulus pluricaulis against neuroinflammation associated depressive behavior induced by chronic unpredictable mild stress in rat. Biomed. Pharmacother. 2019, 109, 1698–1708. [Google Scholar] [CrossRef] [PubMed]
- Shalavadi, M.H.; Chandrashekhar, V.; Muchchandi, I.S. Neuroprotective effect of Convolvulus pluricaulis Choisy in oxidative stress model of cerebral ischemia reperfusion injury and assessment of MAP2 in rats. J. Ethnopharmacol. 2020, 249, 112393. [Google Scholar] [CrossRef] [PubMed]
- Mehla, J.; Gupta, P.; Pahuja, M.; Diwan, D.; Diksha, D. Indian Medicinal Herbs and Formulations for Alzheimer’s Disease, from Traditional Knowledge to Scientific Assessment. Brain Sci. 2020, 10, 964. [Google Scholar] [CrossRef]
- Gayathri, S.; Chandrashekar, H.R.; Fayaz, S.M. Phytotherapeutics Against Alzheimer’s Disease: Mechanism, Molecular Targets and Challenges for Drug Development. CNS Neurol. Disord. Drug Targets 2022, 21, 409–426. [Google Scholar] [CrossRef]
- Garg, G.; Patil, A.N.; Kumar, R.; Bhatia, A.; Kasudhan, K.S.; Pattanaik, S. Protective role of Convolvulus pluricaulis on lipid abnormalities in high-fat diet with low dose streptozotocin-induced experimental rat model. J. Ayurveda Integr. Med. 2020, 11, 426–431. [Google Scholar] [CrossRef]
- Garg, G.; Patil, A.; Singh, J.; Kaushik, N.; Praksah, A.; Pal, A.; Chakrabarti, A. Pharmacological evaluation of Convolvulus pluricaulis as hypolipidaemic agent in Triton WR-1339-induced hyperlipidaemia in rats. J. Pharm. Pharmacol. 2018, 70, 1572–1580. [Google Scholar] [CrossRef]
- Rezq, A.M.; Al-Malki, A.L.; Refai, M.Y.; Kumosani, T.A.; Moselhy, S.S. Phytochemical analysis of Convolvulus hystrix Vahl and its biological effects in rats. Toxicol. Ind. Health. 2015, 31, 304–318. [Google Scholar] [CrossRef]
- Nelson, R.H. Hyperlipidemia as a risk factor for cardiovascular disease. Prim. Care Clin. Off. Pract. 2013, 40, 195–211. [Google Scholar] [CrossRef] [Green Version]
- Nandha, R.; Singh, H.; Moudgill, P.; Kular, G. A pilot study to clinically evaluate the role of herbomineral compound “Rakatchap Har” in the management of essential hypertension. Ayu 2011, 32, 329–332. [Google Scholar] [CrossRef]
- González-Becerra, K.; Ramos-Lopez, O.; Barrón-Cabrera, E.; Riezu-Boj, J.I.; Milagro, F.I.; Martínez-López, E.; Martínez, J.A. Fatty acids, epigenetic mechanisms and chronic diseases: A systematic review. Lipids Health Dis. 2019, 18, 178. [Google Scholar] [CrossRef] [Green Version]
- Martin, H. Role of PPAR-gamma in inflammation. Prospects for therapeutic intervention by food components. Mutat. Res. 2010, 690, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.M.; Staels, B. Peroxisome proliferator-activated receptor γ and adipose tissue–understanding obesity-related changes in regulation of lipid and glucose metabolism. J. Clin. Endocrinol. Metab. 2007, 92, 386–395. [Google Scholar] [CrossRef]
- Kapadia, R.; Yi, J.H.; Vemuganti, R. Mechanisms of anti-inflammatory and neuroprotective actions of PPAR-gamma agonists. Front. Biosci. 2008, 13, 1813–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirza, A.Z.; Althagafi, I.I.; Shamshad, H. Role of PPAR receptor in different diseases and their ligands: Physiological importance and clinical implications. Eur. J. Med. Chem. 2019, 166, 502–513. [Google Scholar] [CrossRef] [PubMed]
- Villarroel-Vicente, C.; Gutiérrez-Palomo, S.; Ferri, J.; Cortes, D.; Cabedo, N. Natural products and analogs as preventive agents for metabolic syndrome via peroxisome proliferator-activated receptors: An overview. Eur. J. Med. Chem. 2021, 221, 113535. [Google Scholar] [CrossRef]
- Francisqueti-Ferron, F.V.; Garcia, J.L.; Ferron, A.J.T.; Nakandakare-Maia, E.T.; Gregolin, C.S.; Silva, J.P.D.C.; Dos Santos, K.C.; Lo, Â.T.C.; Siqueira, J.S.; de Mattei, L.; et al. Gamma-oryzanol as a potential modulator of oxidative stress and inflammation via PPAR-y in adipose tissue: A hypothetical therapeutic for cytokine storm in COVID-19? Mol. Cell. Endocrinol. 2021, 520, 111095. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Lee, D.H.; Ahn, J.; Lee, H.; Choi, W.H.; Jang, Y.J.; Ha, T.Y. γ-Oryzanol Enhances Adipocyte Differentiation and Glucose Uptake. Nutrients 2015, 7, 4851–4861. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.R.; Barrick, C.; Kim, K.A.; Lindner, J.; Blondeau, B.; Fujimoto, Y.; Shiota, M.; Kesterson, R.A.; Kahn, B.B.; Magnuson, M.A. Deletion of PPARgamma in adipose tissues of mice protects against high fat diet-induced obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2005, 102, 6207–6212. [Google Scholar] [CrossRef] [Green Version]
- Kintscher, U.; Law, R.E. PPARgamma-mediated insulin sensitization: The importance of fat versus muscle. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E287–E291. [Google Scholar] [CrossRef] [Green Version]
- Deng, D.; Yan, N. GLUT, SGLT, and SWEET: Structural and mechanistic investigations of the glucose transporters. Protein Sci. 2016, 25, 546–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Asp. Med. 2013, 34, 121–138. [Google Scholar] [CrossRef] [Green Version]
- Pujimulyani, D.; Yulianto, W.A.; Setyowati, A.; Arumwardana, S.; Kusuma, H.S.W.; Sholihah, I.A.; Rizal, R.; Widowati, W.; Maruf, A. Hypoglycemic Activity of Curcuma mangga Val. Extract via Modulation of GLUT4 and PPAR-γ mRNA Expression in 3T3-L1 Adipocytes. J. Exp. Pharmacol. 2020, 12, 363–369. [Google Scholar] [CrossRef]
- Rozjan, A.; Shan, W.; Yao, Q. Metabolic Consequences of Neuronal HIF1α-Deficiency in Mediobasal Hypothalamus in Mice. Front. Endocrinol. 2021, 12, 668193. [Google Scholar] [CrossRef]
- Ruze, R.; Xu, Q.; Liu, G.; Li, Y.; Chen, W.; Cheng, Z.; Xiong, Y.; Liu, S.; Zhang, G.; Hu, S.; et al. Central GLP-1 contributes to improved cognitive function and brain glucose uptake after duodenum-jejunum bypass on obese and diabetic rats. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E392–E409. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Y.; Meng, X.; Li, M.; Cao, W.; Yang, J.; Xu, X.; Liu, W.; Li, W.; Cai, Q.; et al. A novel DPP-4 inhibitor Gramcyclin A attenuates cognitive deficits in APP/PS1/tau triple transgenic mice via enhancing brain GLP-1-dependent glucose uptake. Phytother. Res. 2022, 36, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Reno, C.M.; Puente, E.C.; Sheng, Z.; Daphna-Iken, D.; Bree, A.J.; Routh, V.H.; Kahn, B.B.; Fisher, S.J. Brain GLUT4 Knockout Mice Have Impaired Glucose Tolerance, Decreased Insulin Sensitivity, and Impaired Hypoglycemic Counterregulation. Diabetes 2017, 66, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Leguisamo, N.M.; Lehnen, A.M.; Machado, U.F.; Okamoto, M.; Markoski, M.M.; Pinto, G.H.; Schaan, B.D. GLUT4 content decreases along with insulin resistance and high levels of inflammatory markers in rats with metabolic syndrome. Cardiovasc. Diabetol. 2012, 11, 100. [Google Scholar] [CrossRef] [Green Version]
- Hadrich, F.; Mahmoudi, A.; Chamkha, M.; Isoda, H.; Sayadi, S. Olive Leaves Extract and Oleuropein Improve Insulin Sensitivity in 3T3-L1 Cells and in High-Fat Diet-Treated Rats via PI3K/AkT Signaling Pathway. Oxid. Med. Cell. Longev. 2023, 2023, 6828230. [Google Scholar] [CrossRef]
- Huang, R.; Wang, B.; He, J.; Zhang, Z.; Xie, R.; Li, S.; Li, Q.; Tian, C.; Tuo, Y.; Zheng, R.; et al. Lian-Qu formula treats metabolic syndrome via reducing fat synthesis, insulin resistance and inflammation. J. Ethnopharmacol. 2023, 306, 116060. [Google Scholar] [CrossRef]
- Yadav, S.S.; Jain, N.K.; Singhal, R. In-Vitro Evaluation of Ethanolic Extract of Convolvulus pluricaulis Choisy for AntiInflammation & Memory Enhancing Activity. Int. J. Pharm. Life Sci. 2022, 13, 19–26. [Google Scholar]
- Siddiqui, N.A.; Ahmad, N.; Musthaq, N.; Chattopadhyaya, I.; Kumria, R.; Gupta, S. Neuropharmacological Profile of Extracts of Aerial Parts of Convolvulus pluricaulis Choisy in Mice Model. Open Neurol. J. 2011, 8, 11–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, R.; Sengupta, T.; Roy, S.; Chattarji, S.; Ray, J. Convolvulus pluricaulis extract can modulate synaptic plasticity in rat brain hippocampus. Neuroreport 2020, 31, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Kaur, I.P.; Malik, J. Development and Validation of a Novel, Rapid Gradient HPLC Method for Simultaneous Estimation of Bioactive Marker Compounds in a Mixture of Convolvulus pluricaulis, Withania somnifera and Bacopa monnieri Extracts. J. Chromatogr. Sci. 2020, 57, 920–930. [Google Scholar] [CrossRef]
- Belsito, E.L.; Chidichimo, G.; Di Gioia, M.L.; Leggio, A.; Liguori, A.; Perri, F.; Siciliano, C. Extraction of Quinolizidine Alkaloids in Non Aqueous Basic Conditions: The Case of Spartium Junceum Flowers. Chromatographia 2008, 68, 345–349. [Google Scholar] [CrossRef]
- Guerrini, A.; Sacchetti, G.; Guevara, M.P.E.; Paganetto, G.; Grandini, A.; Maresca, I.; Menghini, L.; Di Martino, L.; Marengo, A.; Tacchini, M. Wild Italian Hyssopus officinalis subsp. aristatus (Godr.) Nyman: From Morphological and Phytochemical Evidences to Biological Activities. Plants 2021, 10, 631. [Google Scholar] [CrossRef]
- Melloni, E.; Zauli, G.; Celeghini, C.; Volpi, I.; Secchiero, P. Release of a specific set of proinflammatory adipokines by differentiating 3T3-L1 cells. Nutrition 2013, 29, 32–37. [Google Scholar] [CrossRef]
- Zauli, G.; Tisato, V.; Melloni, E.; Volpato, S.; Cervellati, C.; Bonaccorsi, G.; Radillo, O.; Marci, R.; Secchiero, P. Inverse correlation between circulating levels of TNF-related apoptosis-inducing ligand and 17β-estradiol. J. Clin. Endocrinol. Metab. 2014, 99, 659–664. [Google Scholar] [CrossRef] [Green Version]
- Sethiya, N.K.; Mishra, S.H. Review on ethnomedicinal uses and phytopharmacology of memory boosting herb Convolvulus pluricaulis Choisy. Aust. J. Herb. Med. 2010, 22, 19–25. [Google Scholar]
- Rachitha, P.; Krupashree, K.; Jayashree, G.V.; Kandikattu, H.K.; Amruta, N.; Gopalan, N.; Rao, M.K.; Khanum, F. Chemical composition, antioxidant potential, macromolecule damage and neuroprotective activity of Convolvulus pluricaulis. Tradit. Complement. Med. 2018, 8, 483–496. [Google Scholar] [CrossRef]
- Elmazar, M.M.; El-Abhar, H.S.; Schaalan, M.F.; Farag, N.A. Phytol/Phytanic acid and insulin resistance: Potential role of phytanic acid proven by docking simulation and modulation of biochemical alterations. PLoS ONE 2013, 8, e45638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraman, S.; Devarajan, N.; Rajagopal, P.; Babu, S.; Ganesan, S.K.; Veeraraghavan, V.P.; Palanisamy, C.P.; Cui, B.; Periyasamy, V.; Chandrasekar, K. β-Sitosterol Circumvents Obesity Induced Inflammation and Insulin Resistance by down-Regulating IKKβ/NF-κB and JNK Signaling Pathway in Adipocytes of Type 2 Diabetic Rats. Molecules 2021, 26, 2101. [Google Scholar] [CrossRef] [PubMed]
- Indu, S.; Vijayalakshmi, P.; Selvaraj, J.; Rajalakshmi, M. Novel Triterpenoids from Cassia fistula Stem Bark Depreciates STZ-Induced Detrimental Changes in IRS-1/Akt-Mediated Insulin Signaling Mechanisms in Type-1 Diabetic Rats. Molecules 2021, 26, 6812. [Google Scholar] [CrossRef]
- Reddy, K.P.; Singh, A.B.; Puri, A.; Srivastava, A.K.; Narender, T. Synthesis of novel triterpenoid (lupeol) derivatives and their in vivo antihyperglycemic and antidyslipidemic activity. Bioorg. Med. Chem. Lett. 2009, 19, 4463–4466. [Google Scholar] [CrossRef] [PubMed]
- Khanra, R.; Bhattacharjee, N.; Dua, T.K.; Nandy, A.; Saha, A.; Kalita, J.; Manna, P.; Dewanjee, S. Taraxerol, a pentacyclic triterpenoid, from Abroma augusta leaf attenuates diabetic nephropathy in type 2 diabetic rats. Biomed. Pharmacother. 2017, 94, 726–741. [Google Scholar] [CrossRef] [PubMed]
- Sangeetha, K.N.; Shilpa, K.; Kumari, P.J.; Lakshmi, B.S. Reversal of dexamethasone induced insulin resistance in 3T3L1 adipocytes by 3β-taraxerol of Mangifera indica. Phytomedicine 2013, 20, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, F.; Qi, J.; Song, X.C.; Hu, Z.F.; Zhu, D.N.; Yu, B.Y. Homoisoflavonoids from the fibrous roots of Polygonatum odoratum with glucose uptake-stimulatory activity in 3T3-L1 adipocytes. J. Nat. Prod. 2010, 73, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Park, J.Y.; Seo, Y.; Han, J.S. A new chromanone isolated from Portulaca oleracea L. increases glucose uptake by stimulating GLUT4 translocation to the plasma membrane in 3T3-L1 adipocytes. Int. J. Biol. Macromol. 2019, 123, 26–34. [Google Scholar] [CrossRef]
- Way, J.M.; Harrington, W.W.; Brown, K.K.; Gottschalk, W.K.; Sundseth, S.S.; Mansfield, T.A.; Ramachandran, R.K.; Willson, T.M.; Kliewer, S.A. Comprehensive messenger ribonucleic acid profiling reveals that peroxisome proliferator-activated receptor gamma activation has coordinate effects on gene expression in multiple insulin-sensitive tissues. Endocrinology 2001, 142, 1269–1277. [Google Scholar] [CrossRef]
- Wu, C.W.; Chu, E.S.H.; Lam, C.N.Y.; Cheng, A.S.L.; Lee, C.W.; Wong, V.W.S.; Sung, J.J.Y.; Yu, J. PPARgamma is essential for protection against nonalcoholic steatohepatitis. Gene Ther. 2010, 17, 790–798. [Google Scholar] [CrossRef] [Green Version]
- Azhar, S. Peroxisome proliferator-activated receptors, metabolic syndrome and cardiovascular disease. Future Cardiol. 2010, 6, 657–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pais, R.; Zietek, T.; Hauner, H.; Daniel, H.; Skurk, T. RANTES (CCL5) reduces glucose-dependent secretion of glucagon-like peptides 1 and 2 and impairs glucose-induced insulin secretion in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G330–G337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueba, T.; Nomura, S.; Inami, N.; Yokoi, T.; Inoue, T. Elevated RANTES level is associated with metabolic syndrome and correlated with activated platelets associated markers in healthy younger men. Clin. Appl. Thromb. Hemost. 2014, 20, 813–818. [Google Scholar] [CrossRef] [Green Version]
- Dworacka, M.; Krzyżagórska, E.; Iskakova, S.; Bekmukhambetov, Y.; Urazayev, O.; Dworacki, G. Increased circulating RANTES in type 2 diabetes. Eur. Cytokine Netw. 2014, 25, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K.; et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef]
- Sartipy, P.; Loskutoff, D.J. Monocyte chemoattractant protein 1 in obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2003, 100, 7265–7270. [Google Scholar] [CrossRef] [Green Version]
- Neels, J.G.; Badeanlou, L.; Hester, K.D.; Samad, F. Keratinocyte-derived chemokine in obesity: Expression, regulation, and role in adipose macrophage infiltration and glucose homeostasis. J. Biol. Chem. 2009, 284, 20692–20698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okdahl, T.; Wegeberg, A.M.; Pociot, F.; Brock, B.; Størling, J.; Brock, C. Low-grade inflammation in type 2 diabetes: A cross-sectional study from a Danish diabetes outpatient clinic. BMJ Open 2022, 12, e062188. [Google Scholar] [CrossRef]
- Balogh, A.; Milibák, T.; Szabó, V.; Nagy, Z.Z.; Kaarniranta, K.; Resch, M.D. Immunological biomarkers of the vitreous responsible for proliferative alteration in the different forms of retinal detachment. BMC Ophthalmol. 2020, 20, 491. [Google Scholar] [CrossRef]
- Gong, Q.Y.; Hu, G.Y.; Yu, S.Q.; Qian, T.W.; Xu, X. Comprehensive assessment of growth factors, inflammatory mediators, and cytokines in vitreous from patients with proliferative diabetic retinopathy. Int. J. Ophthalmol. 2022, 15, 1736–1742. [Google Scholar] [CrossRef]
- Mason, R.H.; Minaker, S.A.; Luna, G.L.; Bapat, P.; Farahvash, A.; Garg, A.; Bhambra, N.; Muni, R.H. Changes in aqueous and vitreous inflammatory cytokine levels in proliferative diabetic retinopathy: A systematic review and meta-analysis. Eye 2022. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, M.; Mitani, T. 25-Hydroxyvitamin D Increases Insulin-Stimulated Glucose Uptake by Enhancing Adipocyte Differentiation. J. Nutr. Sci. Vitaminol. 2022, 68, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Nies, V.J.M.; Struik, D.; Liu, S.; Liu, W.; Kruit, J.K.; Downes, M.; van Zutphen, T.; Verkade, H.J.; Evans, R.M.; Jonker, J.W. Autocrine FGF1 signaling promotes glucose uptake in adipocytes. Proc. Natl. Acad. Sci. USA 2022, 119, e2122382119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cui, C.; Lin, Y.; Cai, J. Ameliorating effect on glycolipid metabolism and chemical profile of Millettia speciosa champ. extract. J. Ethnopharmacol. 2021, 279, 114360. [Google Scholar] [CrossRef] [PubMed]
- Kohn, A.D.; Summers, S.A.; Birnbaum, M.J.; Roth, R.A. Expression of a constitutively active Akt Ser/Thr kinase in 3T3-L1 adipocytes stimulates glucose uptake and glucose transporter 4 translocation. J. Biol. Chem. 1996, 271, 31372–31378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shyur, L.F.; Varga, V.; Chen, C.M.; Mu, S.C.; Chang, Y.C.; Li, S.C. Extract of white sweet potato tuber against TNF-α-induced insulin resistance by activating the PI3K/Akt pathway in C2C12 myotubes. Bot. Stud. 2021, 62, 7. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT (min) | Compounds a | CPTOT | Fraction 1 | Fraction 2 | Fraction 3 | Fraction 4 |
|---|---|---|---|---|---|---|
| Area % b | Area % b | Area % b | Area % b | Area % b | ||
| 4.63 | Glyceraldehyde diethylacetal | 0.38 ± 0.02 | ||||
| 7.77 | Eugenol | 0.26 ± 0.01 | ||||
| 10.92 | 2,4-bis(1,1-dimethylethyl)-phenol | 1.69 ± 0.01 | ||||
| 11.65 | Dihydroactinidiolide | 0.38 ± 0.02 | ||||
| 12.72 | 1-Hexadecene | 0.55 ± 0.05 | ||||
| 16.99 | 1-Octadecene | 0.81 ± 0.06 | ||||
| 17.08 | 5,5,8a-Trimethyl-3,5,6,7,8,8a-hexahydro-2H-chromene | 0.95 ± 0.05 | ||||
| 18.02 | 2-Pentadecanone, 6,10,14-trimethyl- | 0.83 ± 0.06 | ||||
| 19.35 | 7,9-Di-tert-butyl-1-oxaspiro[4,5]deca-6,9-diene-2,8-dione | 0.27 ± 0.01 | ||||
| 19.55 | 8-Hexadecenal, 14-methyl-, (Z)- | 0.19 ± 0.02 | 1.57 ± 0.14 | |||
| 19.70 | Methyl ferulate | 1.56 ± 0.07 | ||||
| 20.06 | Isophytol | 0.26 ± 0.02 | ||||
| 20.26 | Ethyl ferulate | 1.57 ± 0.02 | ||||
| 20.98 | 1-Heptadecanol | 0.62 ± 0.01 | ||||
| 22.74 | 1-Octadecanol | 1.21 ± 0.03 | 3.42 ± 0.09 | |||
| 23.20 | trans-Phytol | 20.09 ± 1.00 | 72.88 ± 3.62 | 1.62 ± 0.08 | ||
| 24.65 | 1-Nonadecanol | 0.34 ±0.01 | ||||
| 25.81 | E,E,Z-1,3,12-Nonadecatriene-5,14-diol | 0.69 ± 0.02 | ||||
| 27.33 | 4,8,12,16-Tetramethylheptadecan-4-olide | 0.53 ± 0.01 | ||||
| 29.68 | 13-Tetradecen-1-ol acetate | 0.77 ± 0.01 | ||||
| 30.18 | 7-Hydroxy-3-(3,4-methylenedioxyphenyl)-4-chromanone | 3.51 ± 0.02 | 89.44 ± 0.58 | |||
| 31.24 | Demethylhomopterocarpin | 0.81 ± 0.01 | ||||
| 31.47 | 3,7,3′,4′-Tetrahydroxyflavone | 2.49 ± 0.02 | ||||
| 32.48 | 3-(2,4-Dimethoxyphenyl)-7-chromanol | 1.34 ± 0.04 | ||||
| 37.23 | Stigmasta-4,7,22-trien-3-ol | 0.56 ± 0.01 | ||||
| 38.11 | 9,19-Cyclolanost-24-en-3-ol, acetate | 2.05 ± 0.02 | ||||
| 38.39 | Stigmastan-3,5-diene | 1.22 ± 0.06 | ||||
| 40.48 | Campesterol | 4.22 ± 0.07 | 4.66 ± 0.08 | |||
| 40.90 | Stigmasterol | 8.92 ± 0.33 | 22.25 ± 0.81 | |||
| 41.89 | β-Sitosterol | 14.12 ± 0.23 | 65.13 ± 1.06 | |||
| 42.26 | α-Amyrin | 6.15 ± 0.35 | 2.79 ± 0.16 | |||
| 42.58 | Taraxerol | 3.38 ± 0.12 | 3.61 ± 0.13 | |||
| 42.95 | 22,23-dihydro-Stigmasterol | trace | 6.40 ± 0.04 | |||
| 43.07 | Lupeol | 17.28 ± 0.17 | 19.15 ± 0.19 | 87.10 ± 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melloni, E.; Grassilli, S.; Romani, A.; Rimondi, E.; Marcuzzi, A.; Zauli, E.; Secchiero, P.; Paganetto, G.; Guerrini, A.; Sacchetti, G.; et al. Convolvulus pluricaulis Choisy’s Extraction, Chemical Characterization and Evaluation of the Potential Effects on Glycaemic Balance in a 3T3-L1 Adipocyte Cell Model. Nutrients 2023, 15, 1727. https://doi.org/10.3390/nu15071727
Melloni E, Grassilli S, Romani A, Rimondi E, Marcuzzi A, Zauli E, Secchiero P, Paganetto G, Guerrini A, Sacchetti G, et al. Convolvulus pluricaulis Choisy’s Extraction, Chemical Characterization and Evaluation of the Potential Effects on Glycaemic Balance in a 3T3-L1 Adipocyte Cell Model. Nutrients. 2023; 15(7):1727. https://doi.org/10.3390/nu15071727
Chicago/Turabian StyleMelloni, Elisabetta, Silvia Grassilli, Arianna Romani, Erika Rimondi, Annalisa Marcuzzi, Enrico Zauli, Paola Secchiero, Guglielmo Paganetto, Alessandra Guerrini, Gianni Sacchetti, and et al. 2023. "Convolvulus pluricaulis Choisy’s Extraction, Chemical Characterization and Evaluation of the Potential Effects on Glycaemic Balance in a 3T3-L1 Adipocyte Cell Model" Nutrients 15, no. 7: 1727. https://doi.org/10.3390/nu15071727