
Fish Oil Supplementation Improves the Repeated-Bout Effect and Redox Balance in 20–30-Year-Old Men Submitted to Strength Training

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Participants
2.2. A Single Bout of a Strength Exercise Protocol
2.3. Strength-Training Protocol
2.4. Supplementation with Fish Oil
2.5. Blood Collection and Plasma Separation
2.6. Measurements of Plasma Cytokines and C-Reactive Protein
2.7. Plasma Activities of Creatine Kinase and Lactate Dehydrogenase
2.8. Measurements of Cortisol and Testosterone
2.9. Determination of Redox State Parameters
2.9.1. Total Iron Determination
2.9.2. Heme Iron Determination
2.9.3. Plasma Trolox Equivalent Antioxidant Capacity (TEAC)
2.9.4. Reduced and Oxidized Glutathione Measurements
2.10. Statistical Analysis
3. Results
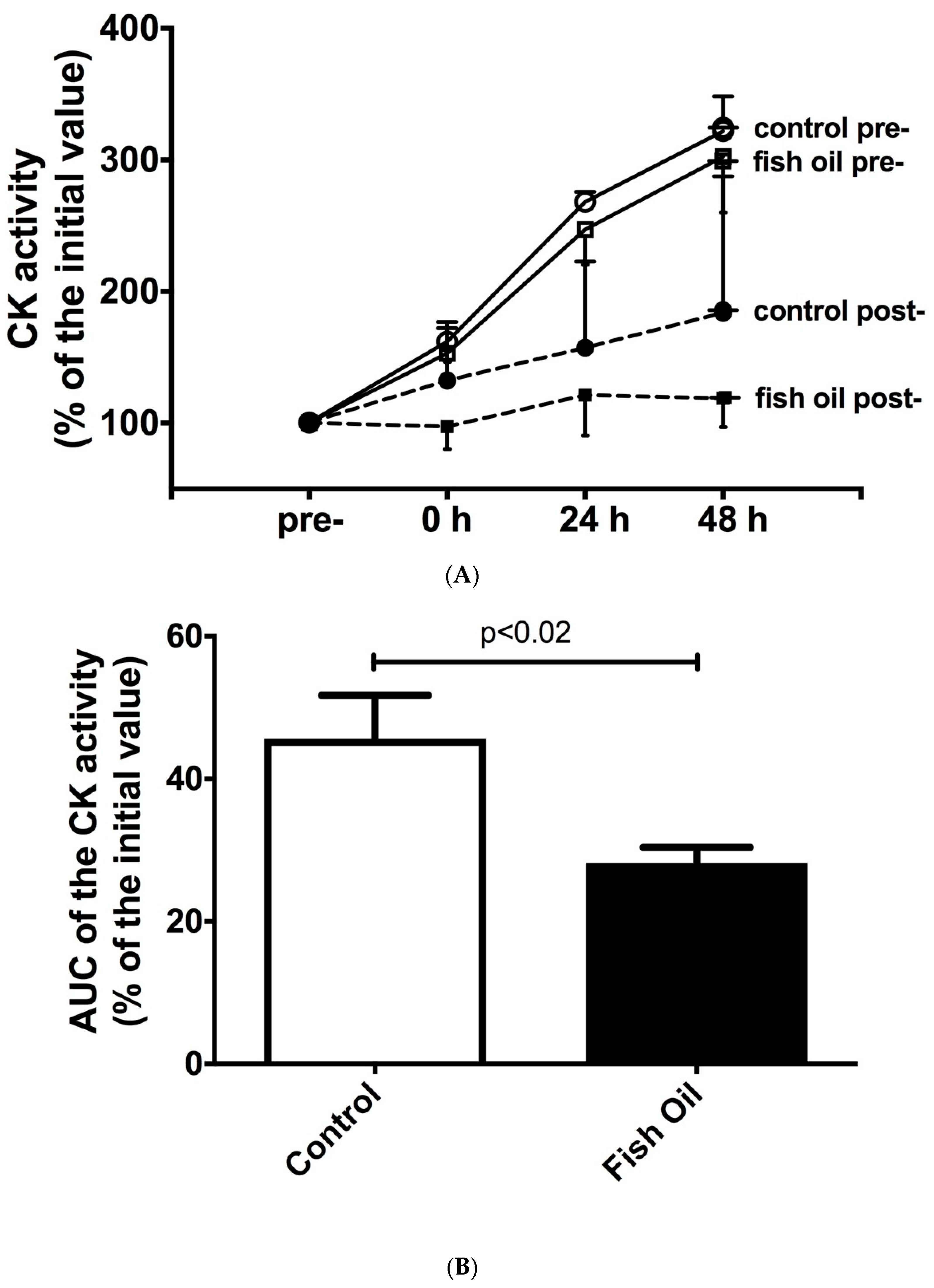
3.1. Plasma Activity of Creatine Kinase and Lactate Dehydrogenase
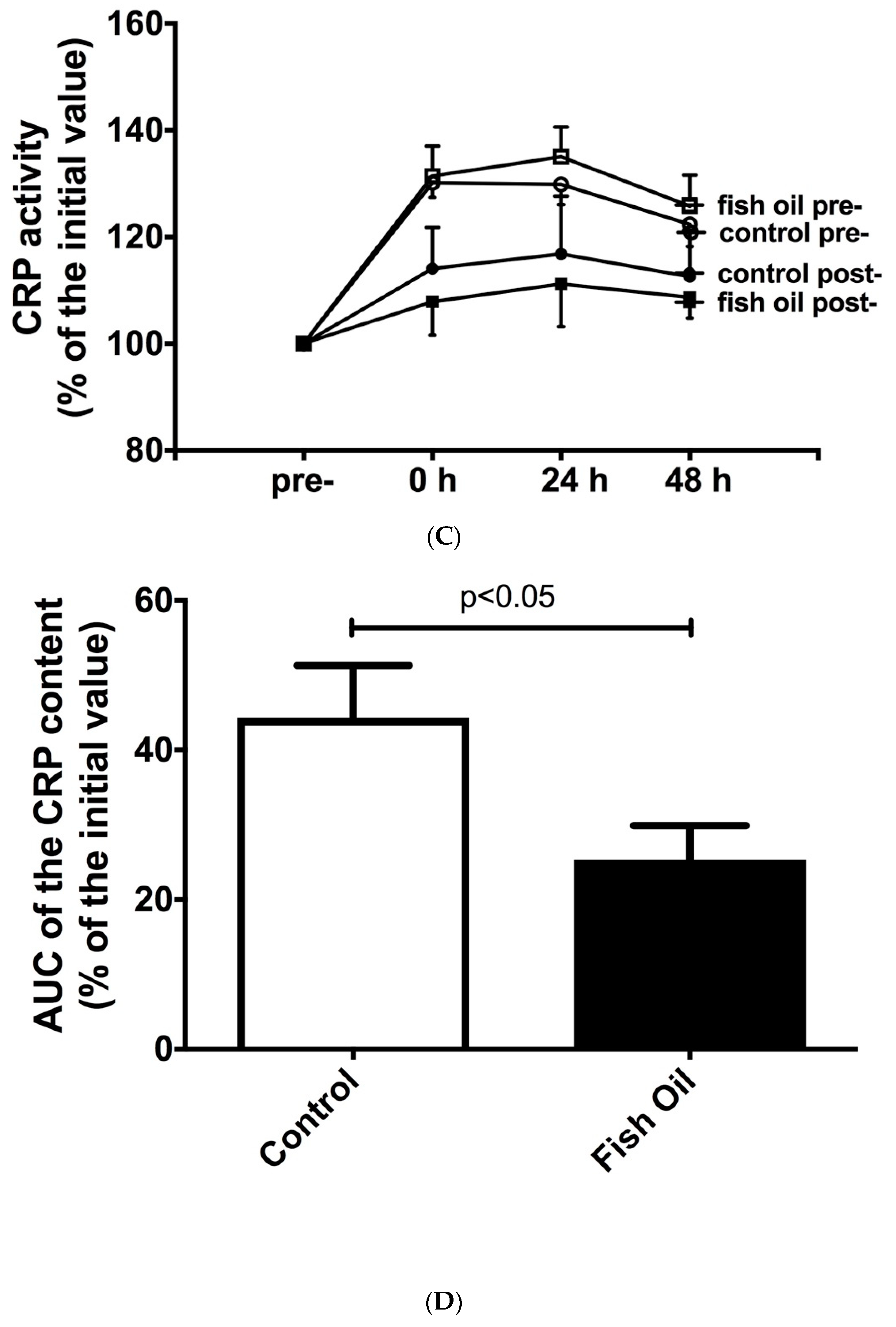
3.2. Determination of Inflammation Markers
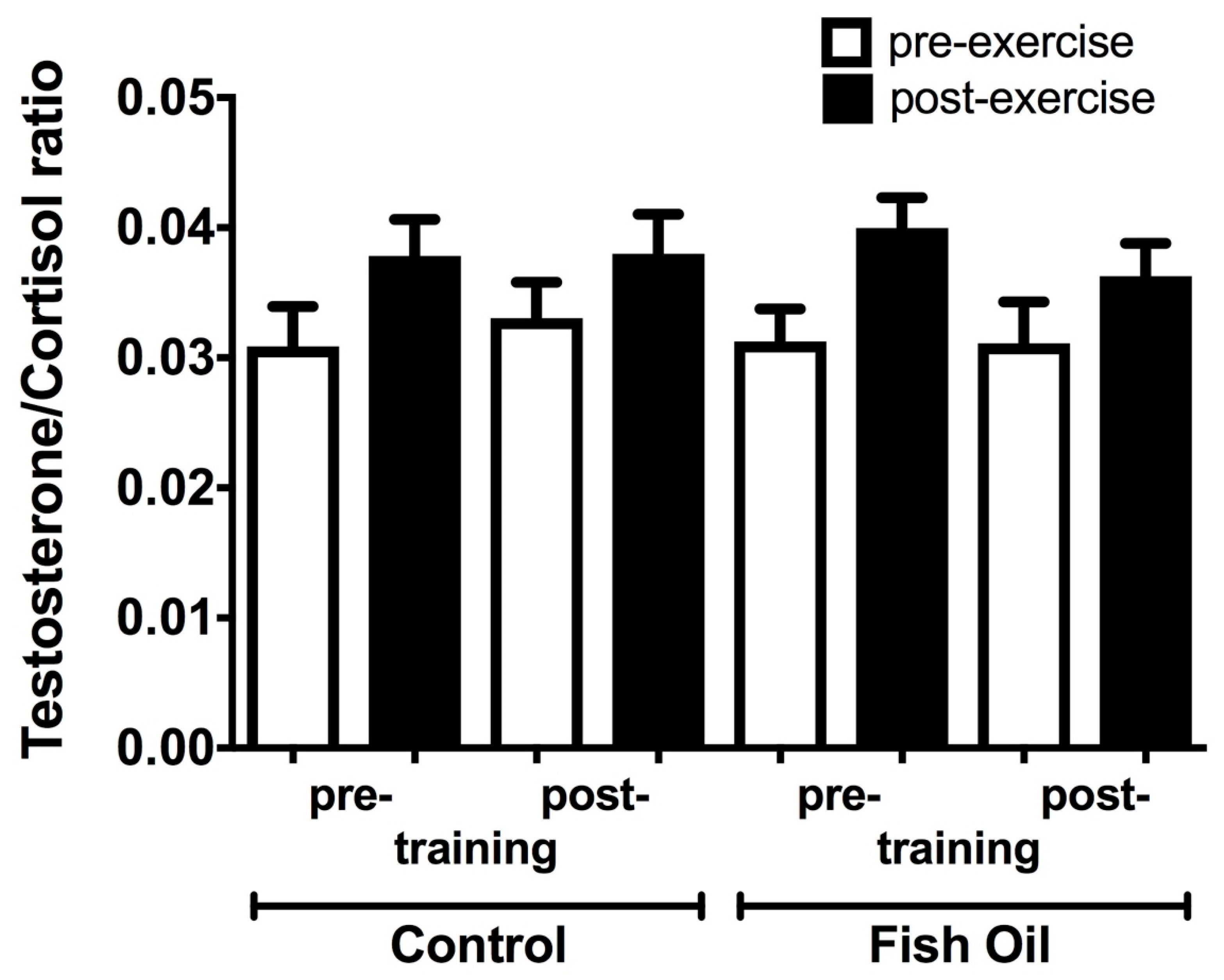
3.3. Plasma Testosterone: Cortisol Ratio
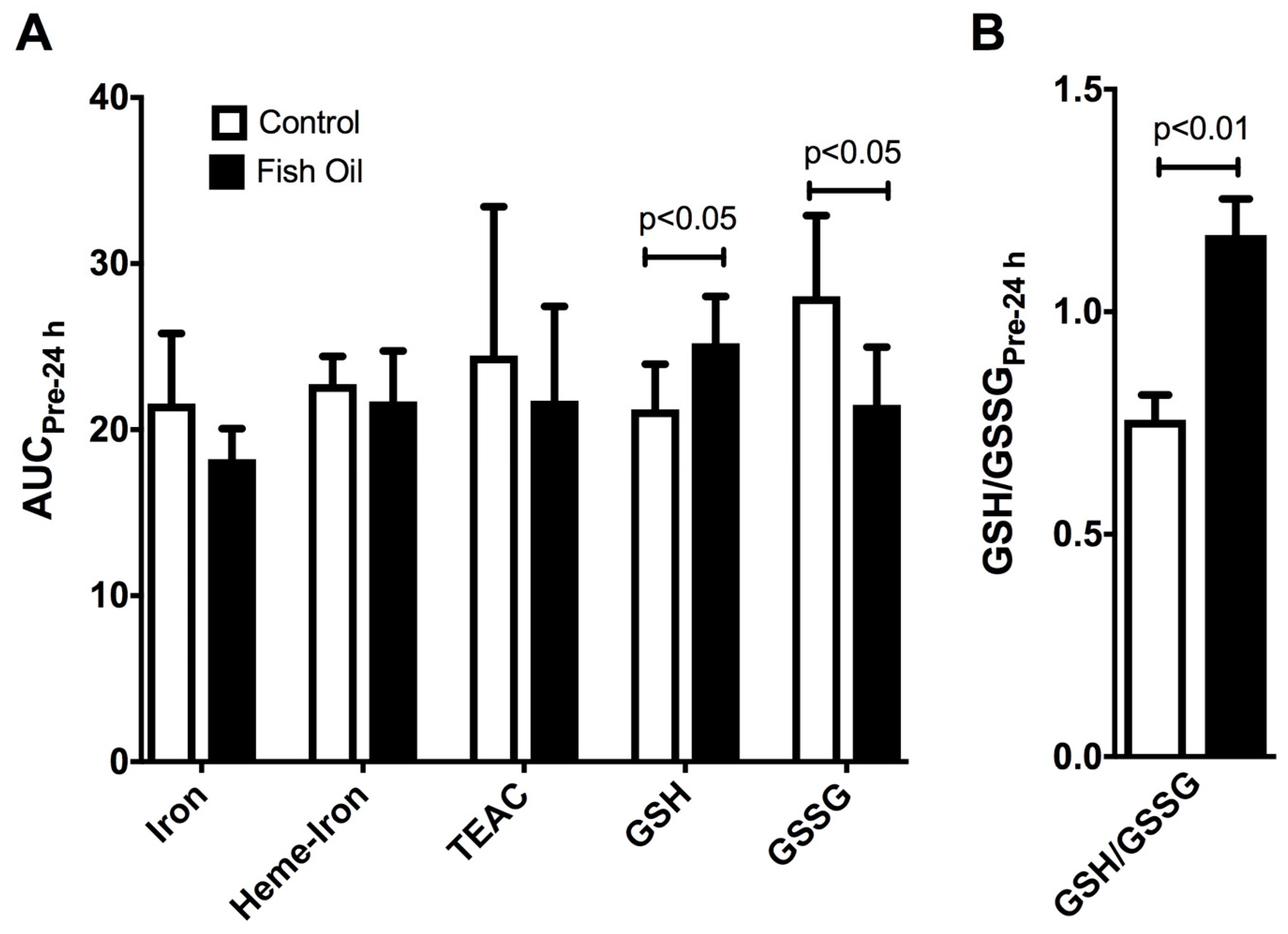
3.4. Measurement of Plasma Redox Parameters
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peake, J.M.; Neubauer, O.; Gatta, P.A.D.; Nosaka, K. Muscle damage and inflammation during recovery from exercise. J. Appl. Physiol. 2017, 122, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.B. Initial events in exercise-induced muscular injury. Med. Sci. Sport. Exerc. 1990, 22, 429–435. [Google Scholar]
- Margonis, K.; Fatouros, I.G.; Jamurtas, A.Z.; Nikolaidis, M.G.; Douroudos, I.; Chatzinikolaou, A.; Mitrakou, A.; Mastorakos, G.; Papassotiriou, I.; Taxildaris, K.; et al. Oxidative stress biomarkers responses to physical overtraining: Implications for diagnosis. Free Radic. Biol. Med. 2007, 43, 901–910. [Google Scholar] [CrossRef] [PubMed]
- González-Hernández, J.M.; Jiménez-Reyes, P.; Cerón, J.J.; Tvarijonaviciute, A.; Llorente-Canterano, F.J.; Martínez-Aranda, L.M.; García-Ramos, A. Response of Muscle Damage Markers to an Accentuated Eccentric Training Protocol: Do Serum and Saliva Measurements Agree? J. Strength Cond. Res. 2022, 36, 2132–2138. [Google Scholar] [CrossRef]
- Uchida, M.C.; Nosaka, K.; Ugrinowitsch, C.; Yamashita, A.; Martins, E., Jr.; Moriscot, A.S.; Aoki, M.S. Effect of bench press exercise intensity on muscle soreness and inflammatory mediators. J. Sport. Sci. 2009, 27, 499–507. [Google Scholar] [CrossRef]
- Chen, T.C.; Yang, T.J.; Huang, M.J.; Wang, H.S.; Tseng, K.W.; Chen, H.L.; Nosaka, K. Damage and the repeated bout effect of arm, leg, and trunk muscles induced by eccentric resistance exercises. Scand. J. Med. Sci. Sport. 2019, 29, 725–735. [Google Scholar] [CrossRef]
- Chen, T.C.; Chen, H.L.; Cheng, L.F.; Chou, T.Y.; Nosaka, K. Effect of Leg Eccentric Exercise on Muscle Damage of the Elbow Flexors after Maximal Eccentric Exercise. Med. Sci. Sport. Exerc. 2021, 53, 1473–1481. [Google Scholar] [CrossRef]
- Paulsen, G.; Mikkelsen, U.R.; Raastad, T.; Peake, J.M. Leucocytes, cytokines and satellite cells: What role do they play in muscle damage and regeneration following eccentric exercise? Exerc. Immunol. Rev. 2012, 18, 42–97. [Google Scholar]
- Kanda, K.; Sugama, K.; Hayashida, H.; Sakuma, J.; Kawakami, Y.; Miura, S.; Yoshioka, H.; Mori, Y.; Suzuki, K. Eccentric exercise-induced delayed-onset muscle soreness and changes in markers of muscle damage and inflammation. Exerc. Immunol. Rev. 2013, 19, 72–85. [Google Scholar]
- Philippou, A.; Tryfonos, A.; Theos, A.; Nezos, A.; Halapas, A.; Maridaki, M.; Koutsilieris, M. Expression of tissue remodelling, inflammation- and angiogenesis-related factors after eccentric exercise in humans. Mol. Biol. Rep. 2021, 48, 4047–4054. [Google Scholar] [CrossRef]
- Bruunsgaard, H.; Galbo, H.; Halkjaer-Kristensen, J.; Johansen, T.L.; MacLean, D.A.; Pedersen, B.K. Exercise-induced increase in serum interleukin-6 in humans is related to muscle damage. J. Physiol. 1997, 499 Pt 3, 833–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, J.G.; Fielding, R.A.; Fiatarone, M.A.; Orencole, S.F.; Dinarello, C.A.; Evans, W.J. Increased interleukin 1 beta in human skeletal muscle after exercise. Am. J. Physiol. 1989, 257, R451–R455. [Google Scholar] [CrossRef]
- Isaacs, A.W.; Macaluso, F.; Smith, C.; Myburgh, K.H. C-Reactive Protein Is Elevated Only in High Creatine Kinase Responders to Muscle Damaging Exercise. Front. Physiol. 2019, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Barquilha, G.; Uchida, M.C.; Santos, V.C.; Moura, N.R.; Lambertucci, R.H.; Hatanaka, E.; Cury-Boaventura, M.F.; Pithon-Curi, T.C.; Gorjão, R.; Hirabara, S.M. Characterization of the Effects of One Maximal Repetition Test on Muscle Injury and Inflammation Markers. WebmedCentral 2011, 2, 1–8. [Google Scholar]
- Bernat-Adell, M.D.; Collado-Boira, E.J.; Moles-Julio, P.; Panizo-González, N.; Martínez-Navarro, I.; Hernando-Fuster, B.; Hernando-Domingo, C. Recovery of Inflammation, Cardiac, and Muscle Damage Biomarkers after Running a Marathon. J. Strength Cond. Res. 2021, 35, 626–632. [Google Scholar] [CrossRef]
- Ebbeling, C.B.; Clarkson, P.M. Exercise-induced muscle damage and adaptation. Sport. Med. 1989, 7, 207–234. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, W.J.; Fleck, S.J.; Evans, W.J. Strength and power training: Physiological mechanisms of adaptation. Exerc. Sport. Sci. Rev. 1996, 24, 363–397. [Google Scholar] [CrossRef]
- Hyldahl, R.D.; Chen, T.C.; Nosaka, K. Mechanisms and Mediators of the Skeletal Muscle Repeated Bout Effect. Exerc. Sport. Sci Rev. 2017, 45, 24–33. [Google Scholar] [CrossRef] [PubMed]
- McHugh, M.P.; Connolly, D.A.; Eston, R.G.; Gleim, G.W. Exercise-induced muscle damage and potential mechanisms for the repeated bout effect. Sport. Med. 1999, 27, 157–170. [Google Scholar] [CrossRef]
- Ferreira-Junior, J.B.; Bottaro, M.; Vieira, A.; Siqueira, A.F.; Vieira, C.A.; Durigan, J.L.Q.; Cadore, E.L.; Coelho, L.G.M.; Simões, H.G.; Bemben, M.G. One session of partial-body cryotherapy (−110 °C) improves muscle damage recovery. Scand. J. Med. Sci. Sport. 2015, 25, e524–e530. [Google Scholar] [CrossRef]
- Kwiecien, S.Y.; McHugh, M.P. The cold truth: The role of cryotherapy in the treatment of injury and recovery from exercise. Eur. J. Appl. Physiol. 2021, 121, 2125–2142. [Google Scholar] [CrossRef] [PubMed]
- Haq, A.; Ribbans, W.J.; Hohenauer, E.; Baross, A.W. The Comparative Effect of Different Timings of Whole Body Cryotherapy Treatment with Cold Water Immersion for Post-Exercise Recovery. Front. Sport. Act. Living 2022, 4, 940516. [Google Scholar] [CrossRef] [PubMed]
- Malaguti, M.; Angeloni, C.; Hrelia, S. Polyphenols in exercise performance and prevention of exercise-induced muscle damage. Oxid. Med. Cell. Longev. 2013, 2013, 825928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calella, P.; Cerullo, G.; Di Dio, M.; Liguori, F.; Di Onofrio, V.; Gallè, F.; Liguori, G. Antioxidant, anti-inflammatory and immunomodulatory effects of spirulina in exercise and sport: A systematic review. Front. Nutr. 2022, 9, 1048258. [Google Scholar] [CrossRef]
- Holdsworth, C.T.; Copp, S.W.; Hirai, D.M.; Ferguson, S.K.; Sims, G.E.; Hageman, K.S.; Stebbins, C.L.; Poole, D.C.; Musch, T.I. The effects of dietary fish oil on exercising skeletal muscle vascular and metabolic control in chronic heart failure rats. Appl. Physiol. Nutr. Metab. 2014, 39, 299–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, C.G.; Santos, V.C.; Levada-Pires, A.C.; Jacintho, T.M.; Gorjão, R.; Pithon-Curi, T.C.; Cury-Boaventura, M.F. Effects of DHA-rich fish oil supplementation on the lipid profile, markers of muscle damage, and neutrophil function in wheelchair basketball athletes before and after acute exercise. Appl. Physiol. Nutr. Metab. 2015, 40, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Visconti, L.M.; Cotter, J.A.; Schick, E.E.; Daniels, N.; Viray, F.E.; Purcell, C.A.; Brotman, C.B.R.; Ruhman, K.E.; Escobar, K.A. Impact of varying doses of omega-3 supplementation on muscle damage and recovery after eccentric resistance exercise. Metabol. Open 2021, 12, 100133. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; He, Q.; Shi, L.; Wu, Y. Impact of Antarctic krill oil supplementation on skeletal muscle injury recovery after resistance exercise. Eur. J. Nutr. 2022, 62, 1345–1356. [Google Scholar] [CrossRef]
- Yosefy, C.; Viskoper, J.R.; Laszt, A.; Priluk, R.; Guita, E.; Varon, D.; Illan, Z.; Berry, E.M.; Savion, N.; Adan, Y.; et al. The effect of fish oil on hypertension, plasma lipids and hemostasis in hypertensive, obese, dyslipidemic patients with and without diabetes mellitus. Prostaglandins Leukot. Essent. Fat. Acids 1999, 61, 83–87. [Google Scholar] [CrossRef]
- Shibabaw, T. Omega-3 polyunsaturated fatty acids: Anti-inflammatory and anti-hypertriglyceridemia mechanisms in cardiovascular disease. Mol. Cell. Biochem. 2021, 476, 993–1003. [Google Scholar] [CrossRef]
- Giordano, E.; Visioli, F. Long-chain omega 3 fatty acids: Molecular bases of potential antioxidant actions. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Feng, X.; Zhang, J.; Zhang, Y.; Xia, F.; Liu, L.; Jin, Z.; Lu, C.; Xia, Y.; Papadimos, T.J.; et al. ω-3 Fish oil fat emulsion preconditioning mitigates myocardial oxidative damage in rats through aldehydes stress. Biomed. Pharmacother. 2019, 118, 109198. [Google Scholar] [CrossRef]
- Tsitouras, P.D.; Gucciardo, F.; Salbe, A.D.; Heward, C.; Harman, S.M. High omega-3 fat intake improves insulin sensitivity and reduces CRP and IL6, but does not affect other endocrine axes in healthy older adults. Horm. Metab. Res. 2008, 40, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Martins, A.R.; Crisma, A.R.; Masi, L.N.; Amaral, C.L.; Marzuca-Nassr, G.N.; Bomfim, L.H.M.; Teodoro, B.G.; Queiroz, A.L.; Serdan, T.D.A.; Torres, R.P.; et al. Attenuation of obesity and insulin resistance by fish oil supplementation is associated with improved skeletal muscle mitochondrial function in mice fed a high-fat diet. J. Nutr. Biochem. 2018, 55, 76–88. [Google Scholar] [CrossRef]
- Djuricic, I.; Calder, P.C. Beneficial Outcomes of Omega-6 and Omega-3 Polyunsaturated Fatty Acids on Human Health: An Update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.M.; Worthley, C.; Murphy, K.J.; Buckley, J.D.; Ferrante, A.; Howe, P.R. n-3 Fatty acid supplementation and regular moderate exercise: Differential effects of a combined intervention on neutrophil function. Br. J. Nutr. 2007, 98, 300–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monnard, C.R.; Dulloo, A.G. Polyunsaturated fatty acids as modulators of fat mass and lean mass in human body composition regulation and cardiometabolic health. Obes. Rev. 2021, 22 (Suppl. 2), e13197. [Google Scholar] [CrossRef]
- Warner, J.G., Jr.; Ullrich, I.H.; Albrink, M.J.; Yeater, R.A. Combined effects of aerobic exercise and omega-3 fatty acids in hyperlipidemic persons. Med. Sci. Sport. Exerc. 1989, 21, 498–505. [Google Scholar] [CrossRef]
- Ma, T.; He, L.; Luo, Y.; Zhang, G.; Cheng, X.; Bai, Y. Use of fish oil and mortality of patients with cardiometabolic multimo rbidity: A prospective study of UK biobank. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 2751–2759. [Google Scholar] [CrossRef]
- Jost, Z.; Tomczyk, M.; Chroboczek, M.; Calder, P.C.; Laskowski, R. Improved Oxygen Uptake Efficiency Parameters Are Not Correlated with VO2peak or Running Economy and Are Not Affected by Omega-3 Fatty Acid Supplementation in Endurance Runners. Int. J. Environ. Res. Public Health 2022, 19, 14043. [Google Scholar] [CrossRef]
- Oostenbrug, G.S.; Mensink, R.P.; Hardeman, M.R.; De Vries, T.; Brouns, F.; Hornstra, G. Exercise performance, red blood cell deformability, and lipid peroxidation: Effects of fish oil and vitamin E. J. Appl. Physiol. 1997, 83, 746–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philpott, J.D.; Witard, O.C.; Galloway, S.D.R. Applications of omega-3 polyunsaturated fatty acid supplementation for sport performance. Res. Sport. Med. 2019, 27, 219–237. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Campo, D.J.; Ávila-Gandía, V.; López-Román, F.J.; Miñarro, J.; Contreras, C.; Soto-Méndez, F.; Domingo Pedrol, J.C.; Luque-Rubia, A.J. Supplementation of Re-Esterified Docosahexaenoic and Eicosapentaenoic Acids Reduce Inflammatory and Muscle Damage Markers after Exercise in Endurance Athletes: A Randomized, Controlled Crossover Trial. Nutrients 2020, 12, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodacki, C.L.; Rodacki, A.L.; Pereira, G.; Naliwaiko, K.; Coelho, I.; Pequito, D.; Fernandes, L.C. Fish-oil supplementation enhances the effects of strength training in elderly women. Am. J. Clin. Nutr. 2012, 95, 428–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornish, S.M.; Cordingley, D.M.; Shaw, K.A.; Forbes, S.C.; Leonhardt, T.; Bristol, A.; Candow, D.G.; Chilibeck, P.D. Effects of Omega-3 Supplementation Alone and Combined with Resistance Exercise on Skeletal Muscle in Older Adults: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 2221. [Google Scholar] [CrossRef]
- da Cruz Alves, N.M.; Pfrimer, K.; Santos, P.C.; de Freitas, E.C.; Neves, T.; Pessini, R.A.; Junqueira-Franco, M.V.M.; Nogueira-Barbosa, M.H.; Greig, C.A.; Ferriolli, E. Randomised Controlled Trial of Fish Oil Supplementation on Responsiveness to Resistance Exercise Training in Sarcopenic Older Women. Nutrients 2022, 14, 2844. [Google Scholar] [CrossRef]
- Leoncini, S.; De Felice, C.; Signorini, C.; Zollo, G.; Cortelazzo, A.; Durand, T.; Galano, J.M.; Guerranti, R.; Rossi, M.; Ciccoli, L.; et al. Cytokine Dysregulation in MECP2- and CDKL5-Related Rett Syndrome: Relationships with Aberrant Redox Homeostasis, Inflammation, and omega-3 PUFAs. Oxid. Med. Cell. Longev. 2015, 2015, 421624. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Ramirez, V.; Macias-Islas, M.A.; Ortiz, G.G.; Pacheco-Moises, F.; Torres-Sanchez, E.D.; Sorto-Gomez, T.E.; Cruz-Ramos, J.A.; Orozco-Aviña, G.; Celis De La Rosa, A.J. Efficacy of fish oil on serum of TNF alpha, IL-1 beta, and IL-6 oxidative stress markers in multiple sclerosis treated with interferon beta-1b. Oxid. Med. Cell. Longev. 2013, 2013, 709493. [Google Scholar] [CrossRef] [Green Version]
- Grosso, G.; Galvano, F.; Marventano, S.; Malaguarnera, M.; Bucolo, C.; Drago, F.; Caraci, F. Omega-3 fatty acids and depression: Scientific evidence and biological mechanisms. Oxid. Med. Cell. Longev. 2014, 2014, 313570. [Google Scholar] [CrossRef] [Green Version]
- Gray, P.; Chappell, A.; Jenkinson, A.M.; Thies, F.; Gray, S.R. Fish oil supplementation reduces markers of oxidative stress but not muscle soreness after eccentric exercise. Int. J. Sport. Nutr. Exerc. Metab. 2014, 24, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Drobnic, F.; Storsve, A.B.; Burri, L.; Ding, Y.; Banquells, M.; Riera, J.; Björk, P.; Ferrer-Roca, V.; Domingo, J.C. Krill-Oil-Dependent Increases in HS-Omega-3 Index, Plasma Choline and Antioxidant Capacity in Well-Conditioned Power Training Athletes. Nutrients 2021, 13, 4237. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Yanagimoto, K.; Nakazato, K.; Hayamizu, K.; Ochi, E. Eicosapentaenoic and docosahexaenoic acids-rich fish oil supplementation attenuates strength loss and limited joint range of motion after eccentric contr actions: A randomized, double-blind, placebo-controlled, parallel-group trial. Eur. J. Appl. Physiol. 2016, 116, 1179–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, Y.; Ueda, H.; Yanagimoto, K.; Kato, A.; Ochi, E. 4-week eicosapentaenoic acid-rich fish oil supplementation partially protects muscular damage following eccentric contractions. J. Int. Soc. Sport. Nutr. 2021, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- Kyriakidou, Y.; Wood, C.; Ferrier, C.; Dolci, A.; Elliott, B. The effect of Omega-3 polyunsaturated fatty acid supplementation on exercise-induced muscle damage. J. Int. Soc. Sport. Nutr. 2021, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- VanDusseldorp, T.A.; Escobar, K.A.; Johnson, K.E.; Stratton, M.T.; Moriarty, T.; Kerksick, C.M.; Mangine, G.T.; Holmes, A.J.; Lee, M.; Endito, M.R.; et al. Impact of Varying Dosages of Fish Oil on Recovery and Soreness Following Eccentric Exercise. Nutrients 2020, 12, 2246. [Google Scholar] [CrossRef]
- Funaki, A.; Gam, H.; Matsuda, T.; Ishikawa, A.; Yamada, M.; Ikegami, N.; Nishikawa, Y.; Sakamaki-Sunaga, M. Influence of Menstrual Cycle on Leukocyte Response Following Exercise-Induced Muscle Damage. Int. J. Environ. Res. Public Health 2022, 19, 9201. [Google Scholar] [CrossRef]
- Uchida, M.C.; Crewther, B.T.; Ugrinowitsch, C.; Bacurau, R.F.; Moriscot, A.S.; Aoki, M.S. Hormonal responses to different resistance exercise schemes of similar total volume. J. Strength Cond. Res. 2009, 23, 2003–2008. [Google Scholar] [CrossRef]
- Helge, J.W.; Ayre, K.J.; Hulbert, A.J.; Kiens, B.; Storlien, L.H. Regular exercise modulates muscle membrane phospholipid profile in rats. J. Nutr. 1999, 129, 1636–1642. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, J.F.; Murphy, B. The colorimetric determination of iron in biological material with reference to its measurement during chelation therapy. Clin. Chem. 1966, 12, 58–69. [Google Scholar] [CrossRef]
- Van den Berg, R.; Haenen, G.; Van den Berg, H.; Bast, A. Applicability of an improved Trolox equivalent antioxidant capacity (TEAC) assay for evaluation of antioxidant capacity measurements of mixtures. Food Chem. 1999, 66, 511–517. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat Protoc 2006, 1, 3159–3165. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge Academic: New York, NY, USA, 1988. [Google Scholar]
- Lakens, D. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Front. Psychol. 2013, 4, 863. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, D.Z.; Cubrilo, D.G.; Puzovic, V.S.; Vuletic, M.S.; Zivkovic, V.I.; Barudzic, N.S.; Radovanovic, D.S.; Djuric, D.M.; Jakovljevic, V.L. Changes in athlete’s redox state induced by habitual and unaccustomed exercise. Oxid. Med. Cell. Longev. 2012, 2012, 805850. [Google Scholar] [CrossRef] [Green Version]
- El Assar, M.; Álvarez-Bustos, A.; Sosa, P.; Angulo, J.; Rodríguez-Mañas, L. Effect of Physical Activity/Exercise on Oxidative Stress and Inflammation in Muscle and Vascular Aging. Int. J. Mol. Sci. 2022, 23, 8713. [Google Scholar] [CrossRef] [PubMed]
- Steensberg, A.; Fischer, C.P.; Keller, C.; Moller, K.; Pedersen, B.K. IL-6 enhances plasma IL-1ra, IL-10, and cortisol in humans. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E433–E437. [Google Scholar] [CrossRef] [PubMed]
- McHugh, M.P. Recent advances in the understanding of the repeated bout effect: The protective effect against muscle damage from a single bout of eccentric exercise. Scand. J. Med. Sci. Sport. 2003, 13, 88–97. [Google Scholar] [CrossRef]
- Urhausen, A.; Gabriel, H.; Kindermann, W. Blood hormones as markers of training stress and overtraining. Sport. Med. 1995, 20, 251–276. [Google Scholar] [CrossRef]
- Tait, J.L.; Bulmer, S.M.; Drake, J.M.; Drain, J.R.; Main, L.C. Impact of 12 weeks of basic military training on testosterone and cortisol responses. BMJ Mil. Health 2022, e002179. [Google Scholar] [CrossRef]
- Crewther, B.; Cronin, J.; Keogh, J.; Cook, C. The salivary testosterone and cortisol response to three loading schemes. J. Strength Cond. Res. 2008, 22, 250–255. [Google Scholar] [CrossRef]
- Gomes, E.C.; Silva os de Oliveira, M.R. Oxidants, antioxidants, and the beneficial roles of exercise-induced production of reactive species. Oxid. Med. Cell. Longev. 2012, 2012, 756132. [Google Scholar] [CrossRef]
- Kanzaki, K.; Watanabe, D.; Shi, J.; Wada, M. Mechanisms of eccentric contraction-induced muscle damage and nutritional supplementations for mitigating it. J. Muscle Res. Cell Motil. 2022, 43, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Kurozumi, R.; Kojima, S. Increase of intracellular glutathione by low-level NO mediated by transcription factor NF-kappaB in RAW 264.7 cells. Biochim. Et Biophys. Acta 2005, 1744, 58–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H.; Moss, K.M. A role of mitochondrial glutathione peroxidase in modulating mitochondrial oxidations in liver. Eur. J. Biochem. 1978, 84, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Garrido, M.; Terron, M.P.; Rodriguez, A.B. Chrononutrition against oxidative stress in aging. Oxid. Med. Cell. Longev. 2013, 2013, 729804. [Google Scholar] [CrossRef]
- Barbosa, A.M.; Francisco, P.D.C.; Motta, K.; Chagas, T.R.; dos Santos, C.; Rafacho, A.; Nunes, E.A. Fish oil supplementation attenuates changes in plasma lipids caused by dexamethasone treatment in rats. Appl. Physiol. Nutr. Metab. 2016, 41, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Gorjao, R.; Dos Santos, C.M.M.; Serdaos, D.A.; Diniz, V.L.S.; Alba-Loureiro, T.C.; Cury-Boaventura, M.F.; Hatanaka, E.; Levada-Pires, A.C.; Sato, F.T.; Pithon-Curi, T.C.; et al. New insights on the regulation of cancer cachexia by N-3 polyunsaturated fatty acids. Pharmacol. Ther. 2019, 196, 117–134. [Google Scholar] [CrossRef]
- Liu, J.; Meng, Q.; Zheng, L.; Yu, P.; Hu, H.; Zhuang, R.; Ge, X.; Liu, Z.; Liang, X.; Zhou, X. Effect of omega-3 polyunsaturated fatty acids on left ventricular remodeling in chronic heart failure: A systematic review and meta-analysis. Br. J. Nutr. 2022, 1–35. [Google Scholar] [CrossRef]
- Siroma, T.K.; Machate, D.J.; Zorgetto-Pinheiro, V.A.; Figueiosdo, P.S.; Marcelino, G.; Hiane, P.A.; Bogo, D.; Pott, A.; Cury, E.R.J.; Guimarães, R.C.A.; et al. Polyphenols and ω-3 PUFAs: Beneficial Outcomes to Obesity and Its Related Metabolic Diseases. Front. Nutr. 2022, 8, 781622. [Google Scholar] [CrossRef]
- Videla, L.A.; Hernandez-Rodas, M.C.; Metherel, A.H.; Valenzuela, R. Influence of the nutritional status and oxidative stress in the desaturation and elongation of n-3 and n-6 polyunsaturated fatty acids: Impact on non-alcoholic fatty liver disease. Prostaglandins Leukot. Essent. Fat. Acids 2022, 181, 102441. [Google Scholar] [CrossRef]
- Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar]
- Ferreira, I.; Falcato, F.; Bandarra, N.; Rauter, A.P. Resolvins, Protectins, and Maresins: DHA-Derived Specialized Pro-Resolving Mediators, Biosynthetic Pathways, Synthetic Approaches, and Their Role in Inflammation. Molecules 2022, 27, 1677. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barquilha, G.; Dos Santos, C.M.M.; Caçula, K.G.; Santos, V.C.; Polotow, T.G.; Vasconcellos, C.V.; Gomes-Santos, J.A.F.; Rodrigues, L.E.; Lambertucci, R.H.; Serdan, T.D.A.; et al. Fish Oil Supplementation Improves the Repeated-Bout Effect and Redox Balance in 20–30-Year-Old Men Submitted to Strength Training. Nutrients 2023, 15, 1708. https://doi.org/10.3390/nu15071708
Barquilha G, Dos Santos CMM, Caçula KG, Santos VC, Polotow TG, Vasconcellos CV, Gomes-Santos JAF, Rodrigues LE, Lambertucci RH, Serdan TDA, et al. Fish Oil Supplementation Improves the Repeated-Bout Effect and Redox Balance in 20–30-Year-Old Men Submitted to Strength Training. Nutrients. 2023; 15(7):1708. https://doi.org/10.3390/nu15071708
Chicago/Turabian StyleBarquilha, Gustavo, Cesar Miguel Momesso Dos Santos, Kim Guimaraes Caçula, Vinícius Coneglian Santos, Tatiana Geraldo Polotow, Cristina Vardaris Vasconcellos, José Alberto Fernandes Gomes-Santos, Luiz Eduardo Rodrigues, Rafael Herling Lambertucci, Tamires Duarte Afonso Serdan, and et al. 2023. "Fish Oil Supplementation Improves the Repeated-Bout Effect and Redox Balance in 20–30-Year-Old Men Submitted to Strength Training" Nutrients 15, no. 7: 1708. https://doi.org/10.3390/nu15071708