

Artepillin C Time−Dependently Alleviates Metabolic Syndrome in Obese Mice by Regulating CREB/CRTC2−BMAL1 Signaling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Cell Culture
2.3. Luciferase Activity Assays
2.4. Isolation of Total RNA Isolation and Quantitative RT−PCR Analysis
2.5. Protein Extraction and Western Blotting
2.6. Plasmid
2.7. Adenoviruses
2.8. Immunofluorescence Staining
2.9. Haematoxylin and Eosin Staining
2.10. Pyruvate Tolerance Tests (PTT) and Insulin Tolerance Tests (ITT)
2.11. Plasma Lipid Assay
2.12. Lipid Production Assay
2.13. Statistical Analyses
3. Results
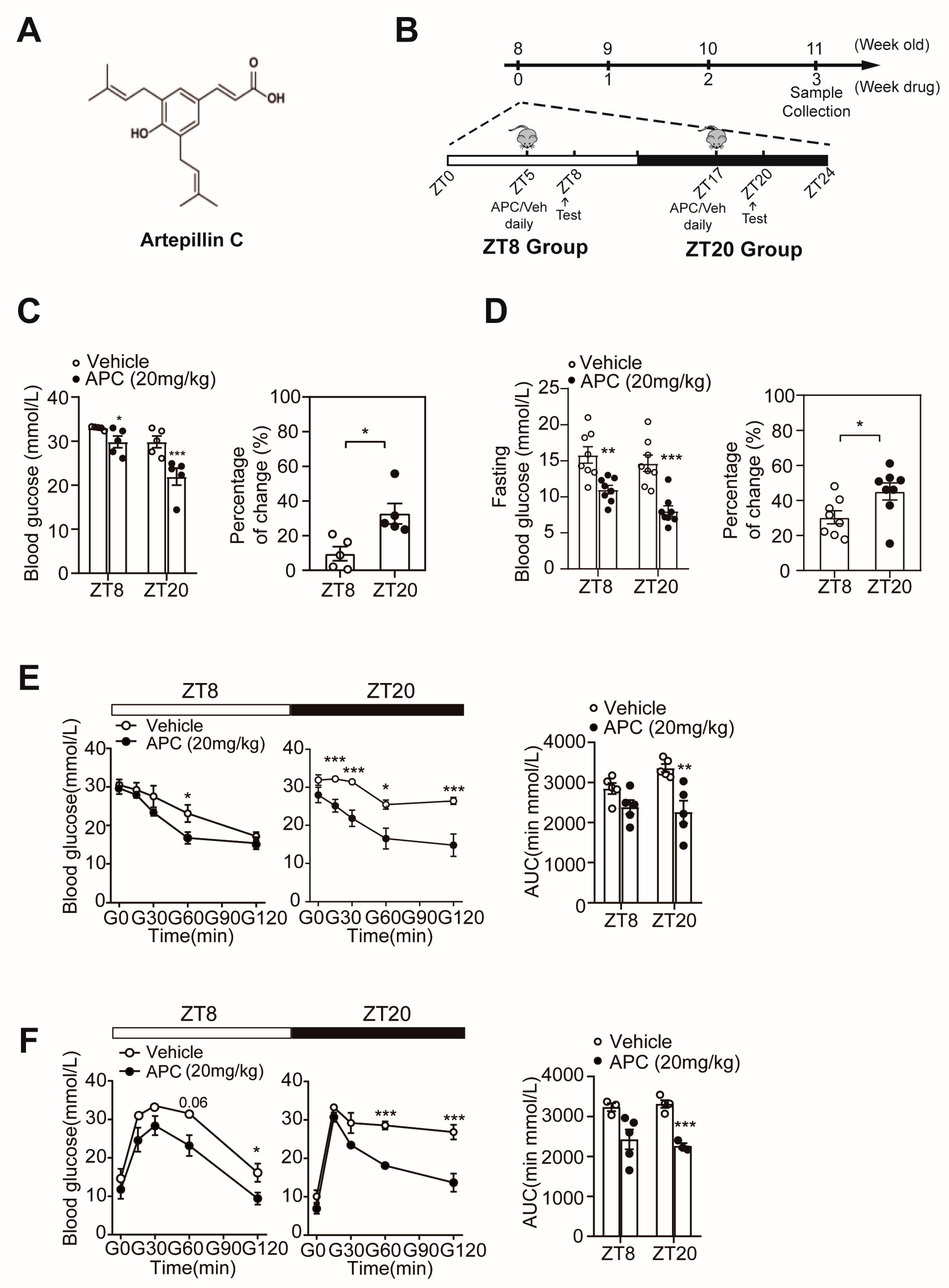
3.1. APC Could Enhance Glucose Homeostasis in a Circadian Rhythmic Manner
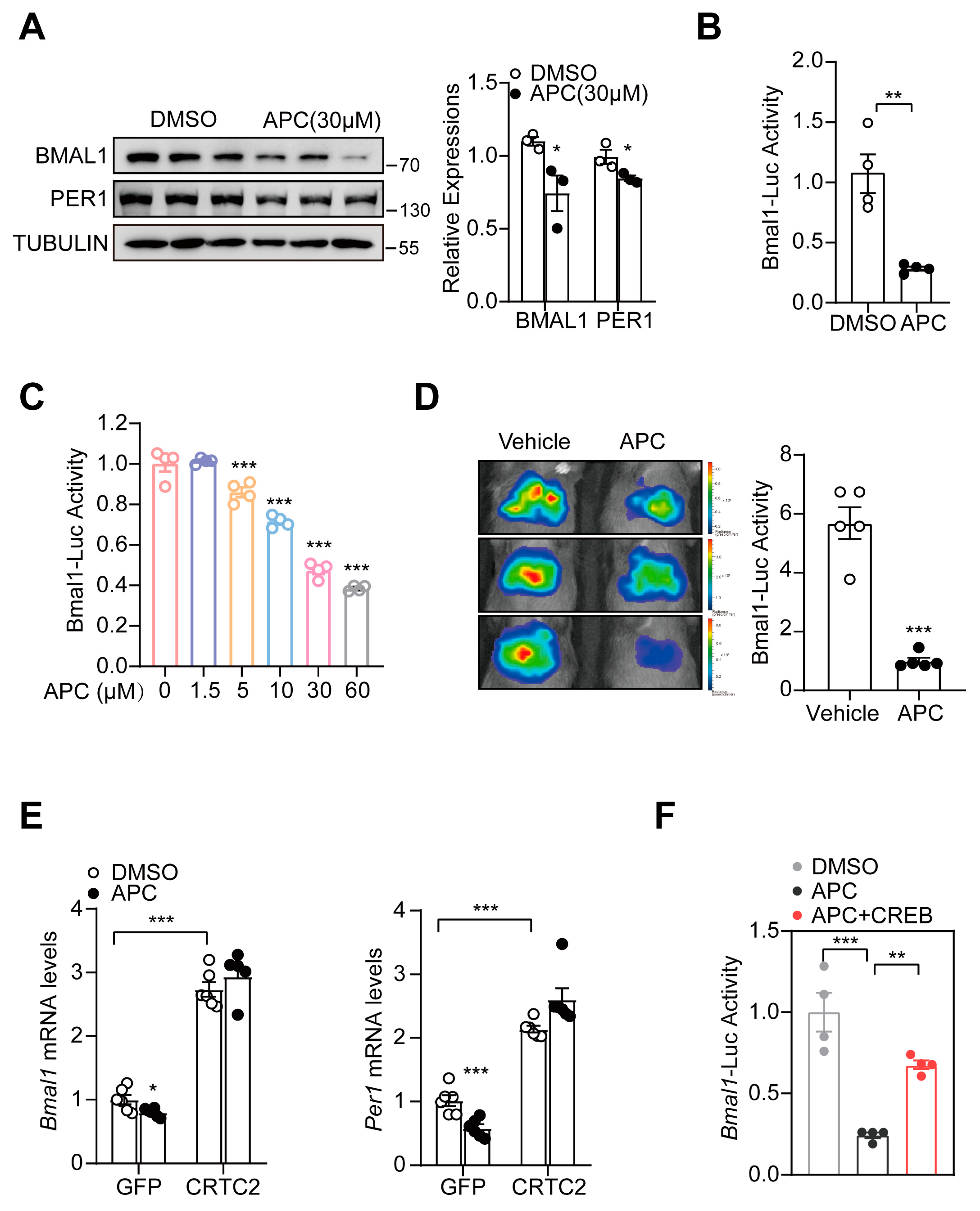
3.2. APC Decreased Hepatic BMAL1 Expression by Blocking CREB/CRTC2 Complex
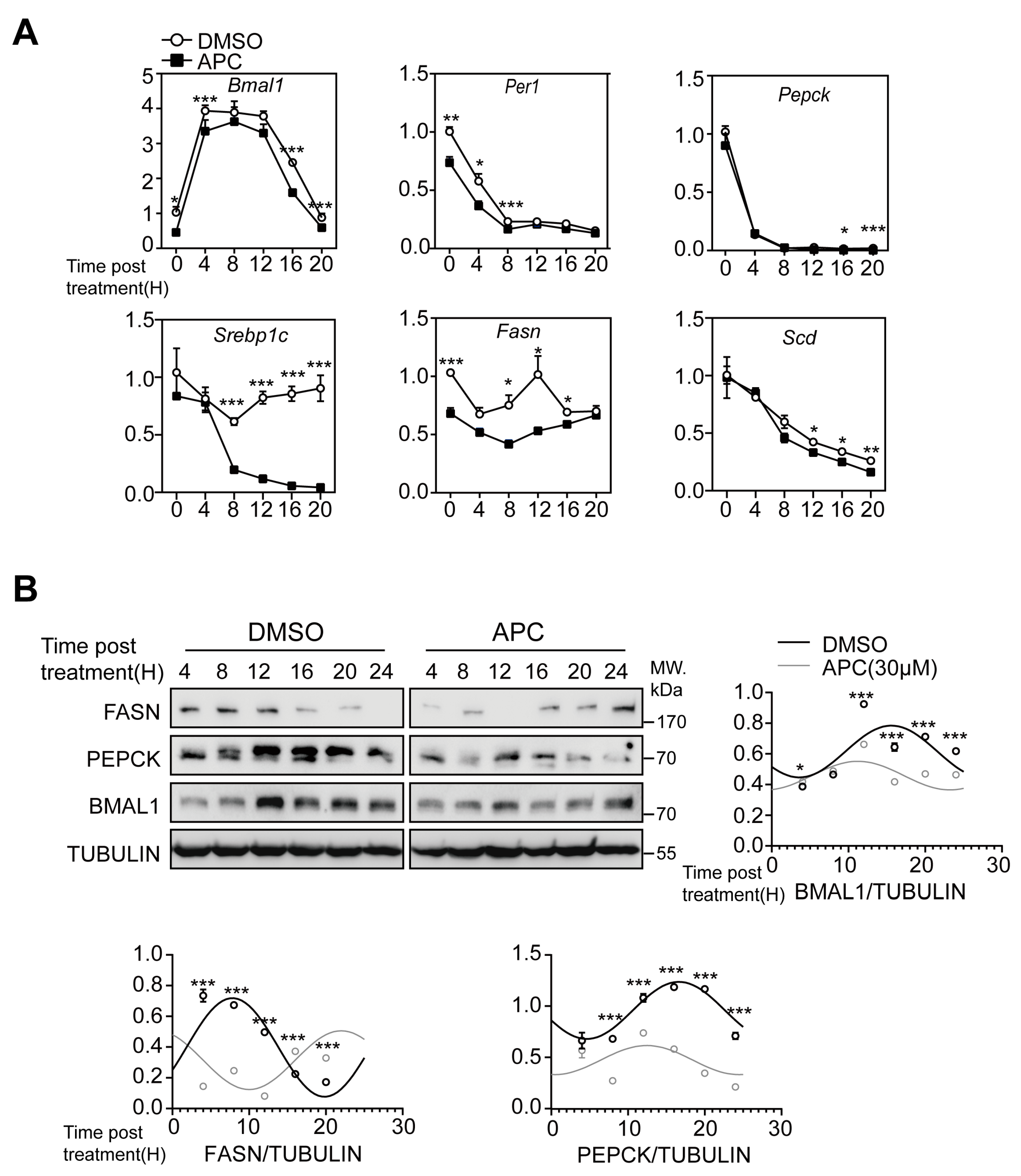
3.3. APC Alters the Circadian Oscillation of BMAL1 Expression in Primary Hepatocytes
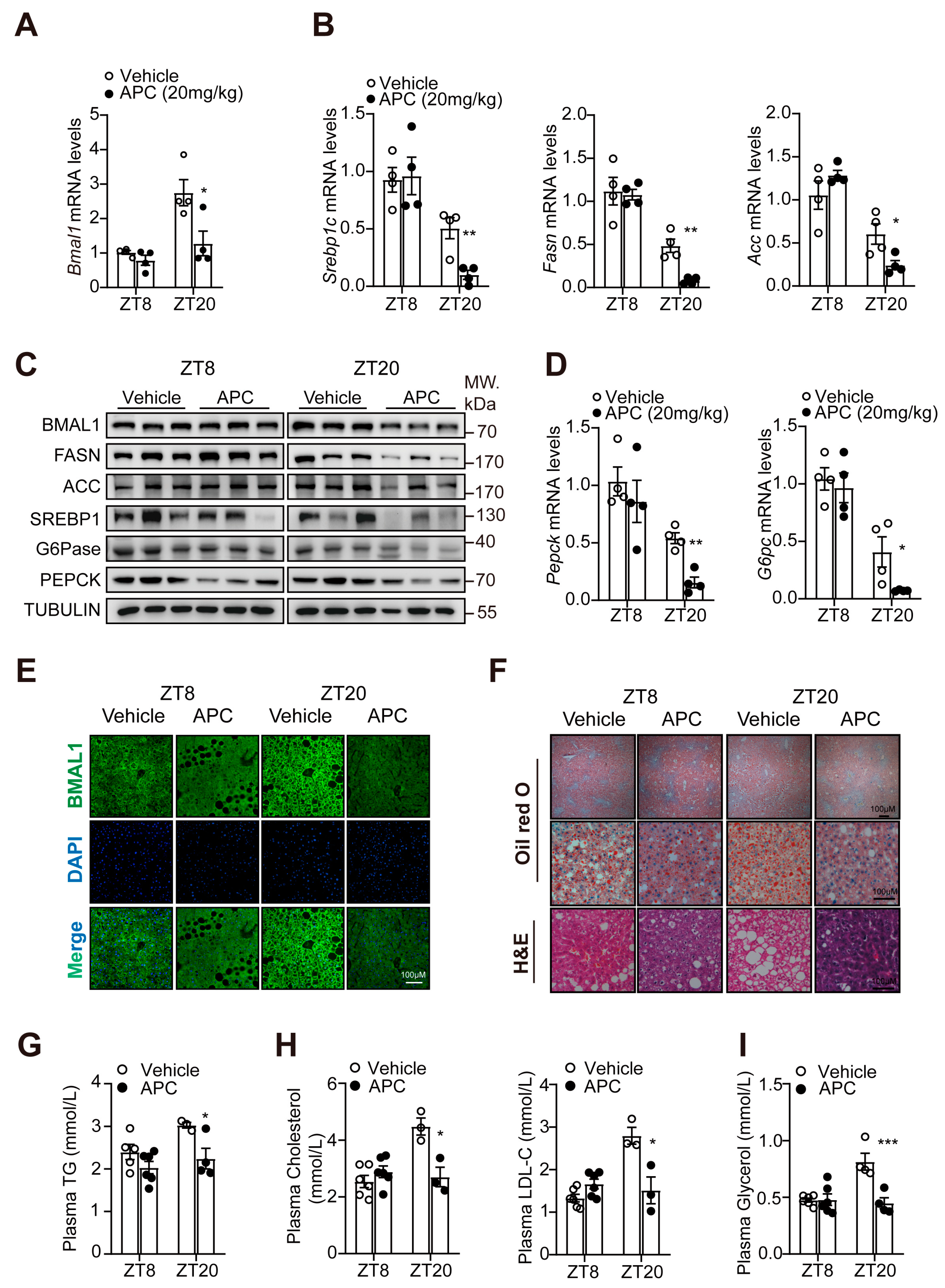
3.4. APC Rhythmically Modulated Hepatic Glucose and Lipid Metabolism in db/db Mice
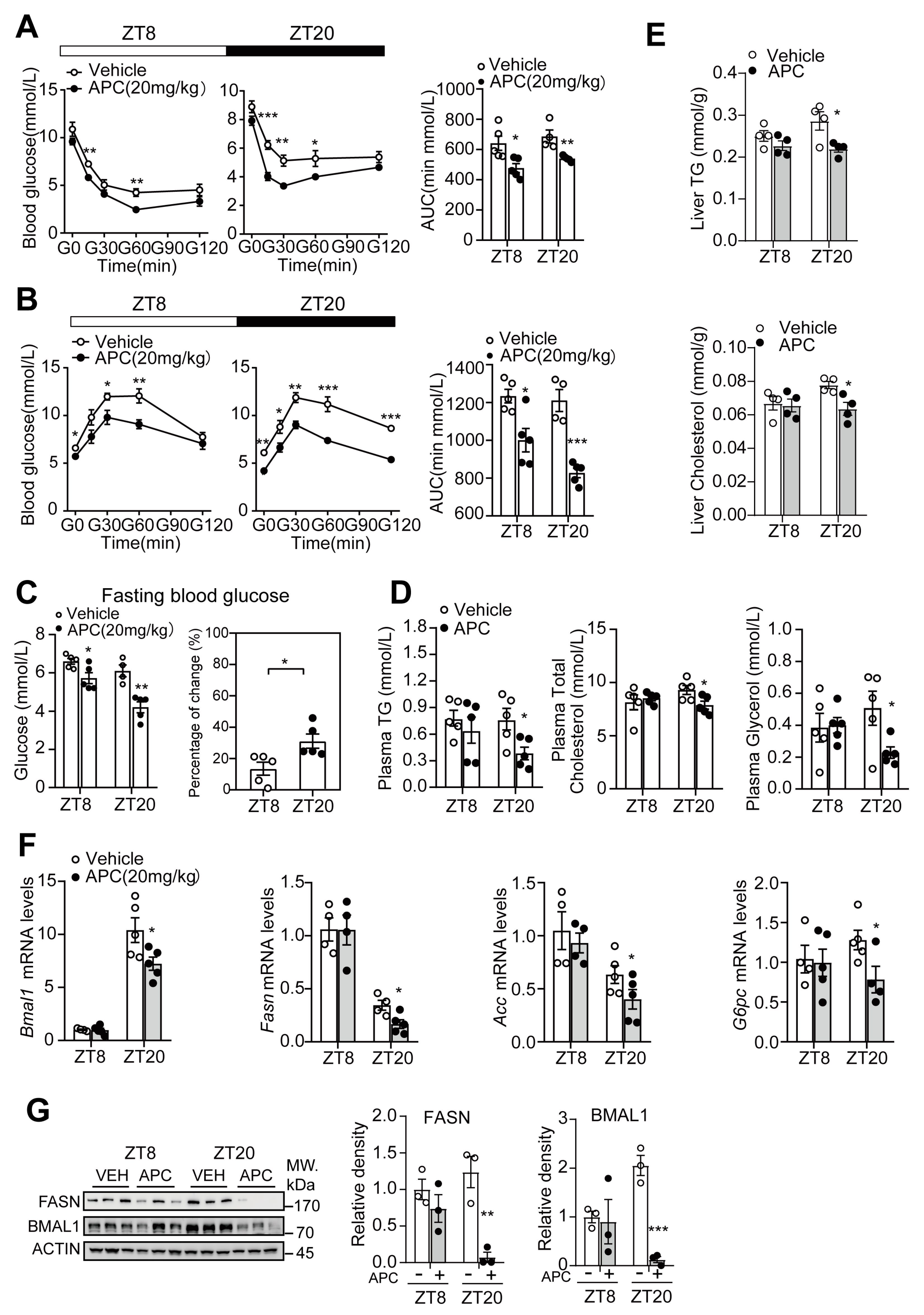
3.5. Time−Dependent Metabolic Protection of APC in DIO Mice
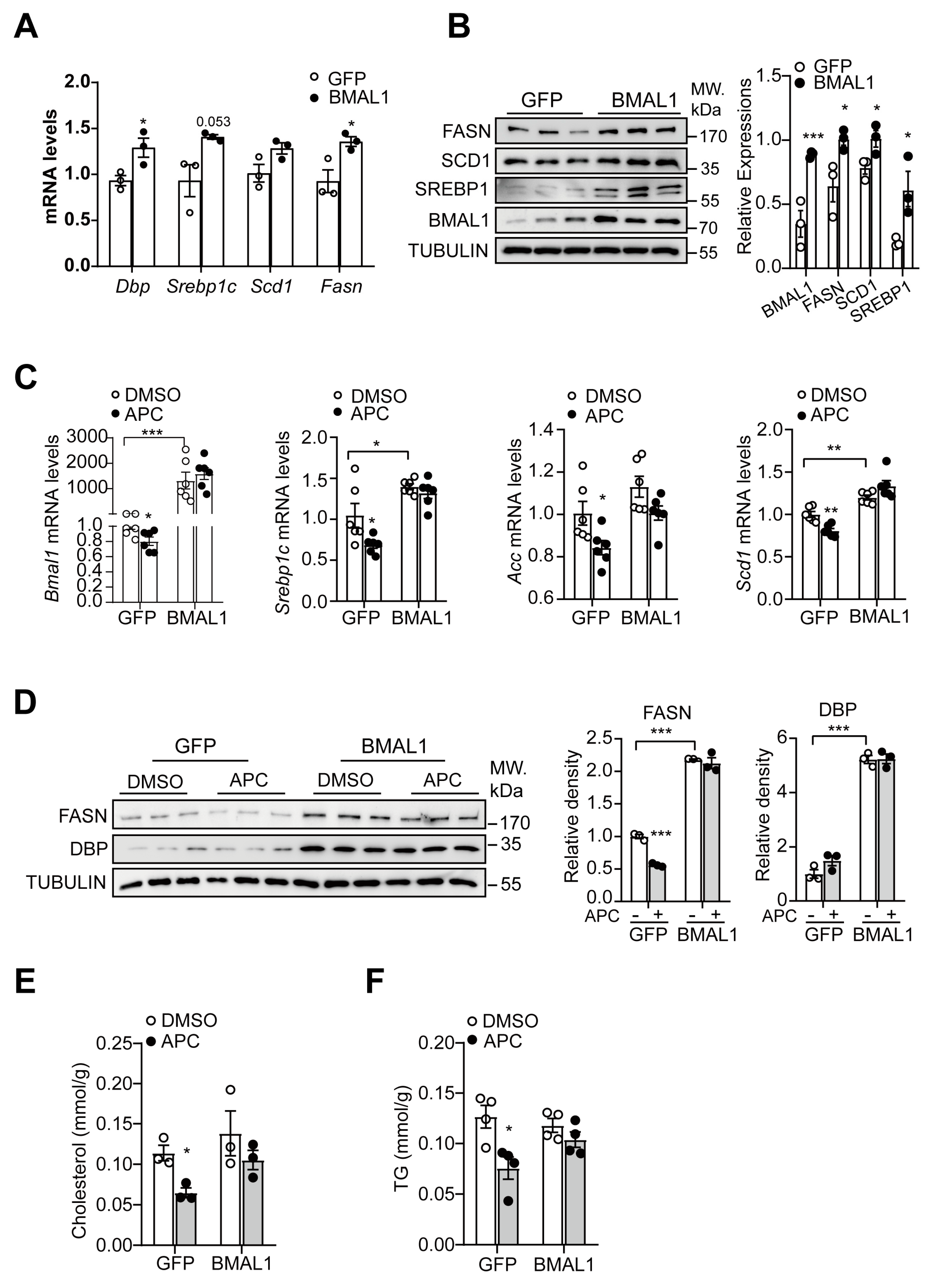
3.6. Involvement of BMAL1 in APC−Mediated Regulation of Hepatic Lipid Synthesis
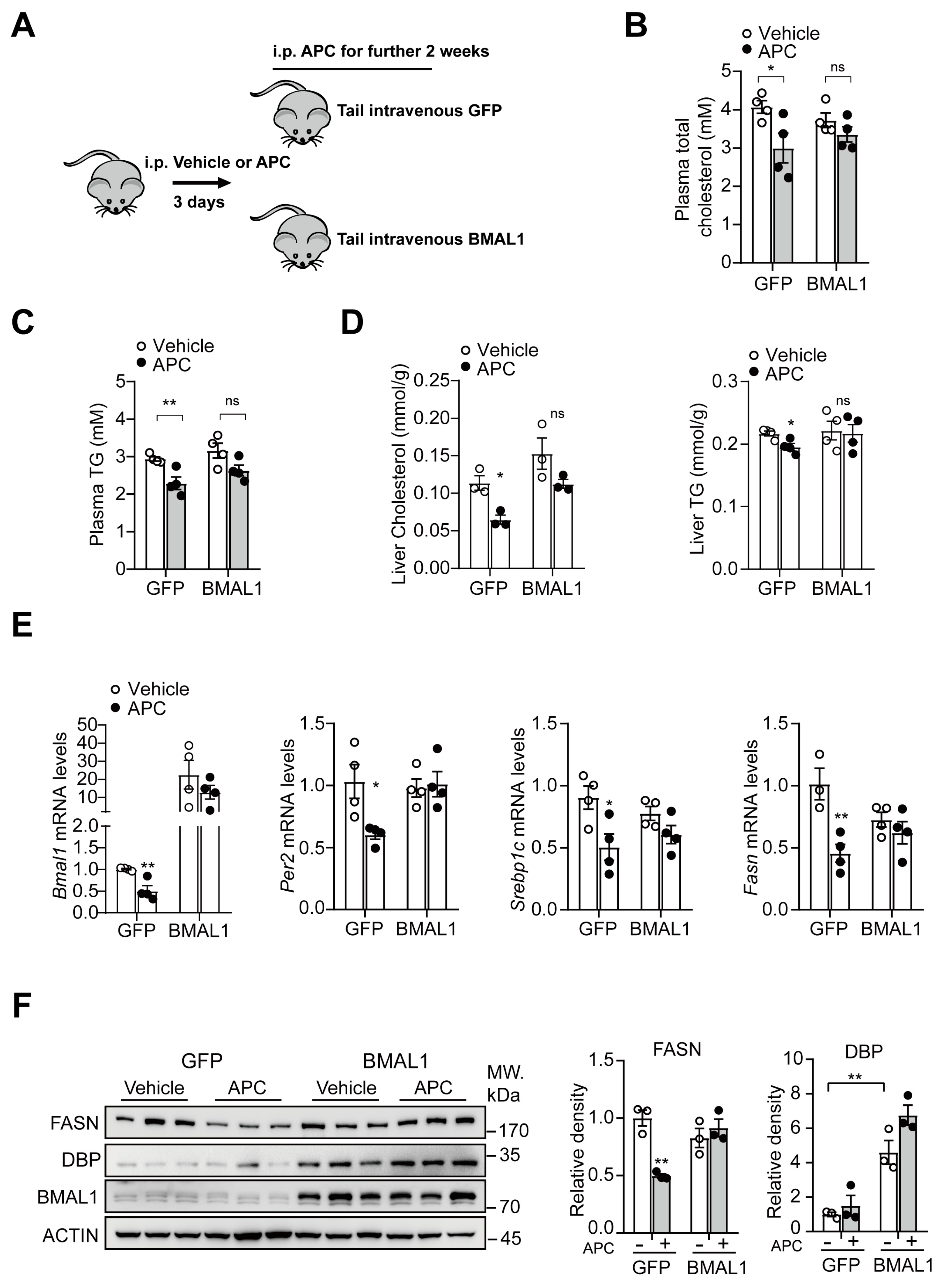
3.7. BMAL1−Overexpression in the Liver Antagonizes APC−Mediated Inhibition in Hepatic Lipid Synthesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Veiga, R.S.; De Mendonça, S.; Mendes, P.B.; Paulino, N.; Mimica, M.J.; Lagareiro Netto, A.A.; Lira, I.; López, B.-C.; Negrão, V.; Marcucci, M. Artepillin C and Phenolic Compounds Responsible for Antimicrobial and Antioxidant Activity of Green Propolis and Baccharis Dracunculifolia Dc. J. Appl. Microbiol. 2017, 122, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Hirata, Y.; Motoyama, M.; Kimura, S.; Takashima, M.; Ikawa, T.; Oh-Hashi, K.; Kamatari, Y.O. Artepillin C, a Major Component of Brazilian Green Propolis, Inhibits Endoplasmic Reticulum Stress and Protein Aggregation. Eur. J. Pharmacol. 2021, 912, 174572. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Mertas, A.; Czuba, Z.P.; Król, W. Inhibition of Inflammatory Response by Artepillin C in Activated Raw264.7 Macrophages. Evid.-Based Complement. Altern. Med. 2013, 2013, 735176. [Google Scholar] [CrossRef] [Green Version]
- Martins, N.S.; Fraga-Silva, T.F.D.C.; Correa, G.F.; Boko, M.M.M.; Ramalho, L.N.Z.; Rodrigues, D.M.; Hori, J.I.; Costa, D.L.; Bastos, J.K.; Bonato, V.L.D. Artepillin C Reduces Allergic Airway Inflammation by Induction of Monocytic Myeloid−Derived Suppressor Cells. Pharmaceutics 2021, 13, 1763. [Google Scholar] [CrossRef] [PubMed]
- Franchi, G.C., Jr.; Moraes, C.S.; Toreti, V.C.; Daugsch, A.; Nowill, A.E.; Park, Y.K. Comparison of Effects of the Ethanolic Extracts of Brazilian Propolis on Human Leukemic Cells as Assessed with the Mtt Assay. Evid.-Based Complement. Altern. Med. 2012, 2012, 918956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahinozzaman, M.; Basak, B.; Emran, R.; Rozario, P.; Obanda, D.N. Artepillin C: A Comprehensive Review of Its Chemistry, Bioavailability, and Pharmacological Properties. Fitoterapia 2020, 147, 104775. [Google Scholar] [CrossRef]
- Cheung, K.-W.; Sze, D.M.-Y.; Chan, W.K.; Deng, R.-X.; Tu, W.; Chan, G.C.F. Brazilian Green Propolis and Its Constituent, Artepillin C Inhibits Allogeneic Activated Human Cd4 T Cells Expansion and Activation. J. Ethnopharmacol. 2011, 138, 463–471. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Wang, Y.; Wang, P.; Zhou, Z.; Wu, R.; Xu, Q.; You, H.; Liu, Y.; Wang, L.; et al. A Propolis−Derived Small Molecule Ameliorates Metabolic Syndrome in Obese Mice by Targeting the Creb/Crtc2 Transcriptional Complex. Nat. Commun. 2022, 13, 246. [Google Scholar] [CrossRef]
- Gachon, F.; Firsov, D. The Role of Circadian Timing System on Drug Metabolism and Detoxification. Expert Opin. Drug Metab. Toxicol. 2011, 7, 147–158. [Google Scholar] [CrossRef]
- Chen, M.; Zhou, C.; Xu, H.; Zhang, T.; Wu, B. Chronopharmacological Targeting of Rev-Erbalpha by Puerarin Alleviates Hyperhomocysteinemia in Mice. Biomed. Pharmacother. 2020, 125, 109936. [Google Scholar] [CrossRef]
- Dunlap, J.C. Molecular Bases for Circadian Clocks. Cell 1999, 96, 271–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, D.K.; Takahashi, J.S.; Kay, S.A. Suprachiasmatic Nucleus: Cell Autonomy and Network Properties. Annu. Rev. Physiol. 2010, 72, 551–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masri, S.; Sassone-Corsi, P. The Emerging Link between Cancer, Metabolism, and Circadian Rhythms. Nat. Med. 2018, 24, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Mure, L.S.; Le, H.D.; Benegiamo, G.; Chang, M.W.; Rios, L.; Jillani, N.; Ngotho, M.; Kariuki, T.; Dkhissi-Benyahya, O.; Cooper, H.M.; et al. Diurnal Transcriptome Atlas of a Primate across Major Neural and Peripheral Tissues. Science 2018, 359, eaao0318. [Google Scholar] [CrossRef] [Green Version]
- Panda, S. Circadian Physiology of Metabolism. Science 2016, 354, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Dyar, K.A.; Treebak, J.T.; Jepsen, S.L.; Ehrlich, A.M.; Ashcroft, S.P.; Trost, K.; Kunzke, T.; Prade, V.M.; Small, L.; et al. Atlas of Exercise Metabolism Reveals Time-Dependent Signatures of Metabolic Homeostasis. Cell Metab. 2022, 34, 329–345.e8. [Google Scholar] [CrossRef]
- Roenneberg, T.; Allebrandt, K.V.; Merrow, M.; Vetter, C. Social Jetlag and Obesity. Curr. Biol. 2012, 22, 939–943. [Google Scholar] [CrossRef] [Green Version]
- Crosby, P.; Hamnett, R.; Putker, M.; Hoyle, N.P.; Reed, M.; Karam, C.J.; Maywood, E.S.; Stangherlin, A.; Chesham, J.E.; Hayter, E.A.; et al. Insulin/Igf-1 Drives Period Synthesis to Entrain Circadian Rhythms with Feeding Time. Cell 2019, 177, 896–909.e20. [Google Scholar] [CrossRef] [Green Version]
- Xing, C.; Zhou, Y.; Xu, H.; Ding, M.; Zhang, Y.; Zhang, M.; Hu, M.; Huang, X.; Song, L. Sleep Disturbance Induces Depressive Behaviors and Neuroinflammation by Altering the Circadian Oscillations of Clock Genes in Rats. Neurosci. Res. 2021, 171, 124–132. [Google Scholar] [CrossRef]
- Cox, K.H.; Takahashi, J.S. Circadian Clock Genes and the Transcriptional Architecture of the Clock Mechanism. J. Mol. Endocrinol. 2019, 63, R93–R102. [Google Scholar] [CrossRef] [Green Version]
- Dibner, C.; Schibler, U.; Albrecht, U. The Mammalian Circadian Timing System: Organization and Coordination of Central and Peripheral Clocks. Annu. Rev. Physiol. 2010, 72, 517–549. [Google Scholar] [CrossRef] [Green Version]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and Peripheral Circadian Clocks in Mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, E.E.; Kay, S.A. Clocks Not Winding Down: Unravelling Circadian Networks. Nat. Rev. Mol. Cell. Biol. 2010, 11, 764–776. [Google Scholar] [CrossRef]
- Marcheva, B.; Ramsey, K.M.; Buhr, E.D.; Kobayashi, Y.; Su, H.; Ko, C.H.; Ivanova, G.; Omura, C.; Mo, S.; Vitaterna, M.H.; et al. Disruption of the Clock Components Clock and Bmal1 Leads to Hypoinsulinaemia and Diabetes. Nature 2010, 466, 627–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobi, D.; Liu, S.; Burkewitz, K.; Kory, N.; Knudsen, N.H.; Alexander, R.K.; Unluturk, U.; Li, X.; Kong, X.; Hyde, A.L.; et al. Hepatic Bmal1 Regulates Rhythmic Mitochondrial Dynamics and Promotes Metabolic Fitness. Cell Metab. 2015, 22, 709–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peek, C.B.; Levine, D.C.; Cedernaes, J.; Taguchi, A.; Kobayashi, Y.; Tsai, S.J.; Bonar, N.A.; McNulty, M.R.; Ramsey, K.M.; Bass, J. Circadian Clock Interaction with Hif1α Mediates Oxygenic Metabolism and Anaerobic Glycolysis in Skeletal Muscle. Cell Metab. 2017, 25, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamia, K.A.; Storch, K.F.; Weitz, C.J. Physiological Significance of a Peripheral Tissue Circadian Clock. Proc. Natl. Acad. Sci. USA 2008, 105, 15172–15177. [Google Scholar] [CrossRef] [Green Version]
- Dang, F.; Sun, X.; Ma, X.; Wu, R.; Zhang, D.; Chen, Y.; Xu, Q.; Wu, Y.; Liu, Y. Insulin Post-Transcriptionally Modulates Bmal1 Protein to Affect the Hepatic Circadian Clock. Nat. Commun. 2016, 7, 12696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Dang, F.; Li, P.; Wang, P.; Xu, Q.; Liu, Z.; Li, Y.; Wu, Y.; Chen, Y.; Liu, Y. The Circadian Protein Period2 Suppresses Mtorc1 Activity Via Recruiting Tsc1 to Mtorc1 Complex. Cell Metab. 2019, 29, 653–667.e6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, E.; Liu, Y.; Dentin, R.; Pongsawakul, P.Y.; Liu, A.C.; Hirota, T.; Nusinow, D.; Sun, X.; Landais, S.; Kodama, Y.; et al. Cryptochrome Mediates Circadian Regulation of Camp Signaling and Hepatic Gluconeogenesis. Nat. Med. 2010, 16, 1152–1156. [Google Scholar] [CrossRef]
- Hirota, T.; Lee, J.W.; Lewis, W.G.; Zhang, E.; Breton, G.; Liu, X.; Garcia, M.; Peters, E.C.; Etchegaray, J.-P.; Traver, D.; et al. High-Throughput Chemical Screen Identifies a Novel Potent Modulator of Cellular Circadian Rhythms and Reveals Ckialpha as a Clock Regulatory Kinase. PLoS Biol. 2010, 8, e1000559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Dentin, R.; Chen, D.; Hedrick, S.; Ravnskjaer, K.; Schenk, S.; Milne, J.; Meyers, D.J.; Cole, P.; Iii, J.Y.; et al. A Fasting Inducible Switch Modulates Gluconeogenesis Via Activator/Coactivator Exchange. Nature 2008, 456, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Bass, J. Circadian Topology of Metabolism. Nature 2012, 491, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Seok, S.; Fu, T.; Choi, S.-E.; Li, Y.; Zhu, R.; Kumar, S.; Sun, X.; Yoon, G.; Kang, Y.; Zhong, W.; et al. Transcriptional Regulation of Autophagy by an Fxr-Creb Axis. Nature 2014, 516, 108–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Vera, L.; Fischer, W.H.; Montminy, M. The Creb Coactivator Crtc2 Links Hepatic Er Stress and Fasting Gluconeogenesis. Nature 2009, 460, 534–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Inoue, H.; Ravnskjaer, K.; Viste, K.; Miller, N.; Liu, Y.; Hedrick, S.; Vera, L.; Montminy, M. Targeted Disruption of the Creb Coactivator Crtc2 Increases Insulin Sensitivity. Proc. Natl. Acad. Sci. USA 2010, 107, 3087–3092. [Google Scholar] [CrossRef] [Green Version]
- Sinturel, F.; Gerber, A.; Mauvoisin, D.; Wang, J.; Gatfield, D.; Stubblefield, J.J.; Green, C.B.; Gachon, F.; Schibler, U. Diurnal Oscillations in Liver Mass and Cell Size Accompany Ribosome Assembly Cycles. Cell 2017, 169, 651–663.e14. [Google Scholar] [CrossRef] [Green Version]
- Hatori, M.; Panda, S. Cry Links the Circadian Clock and Creb-Mediated Gluconeogenesis. Cell Res. 2010, 20, 1285–1288. [Google Scholar] [CrossRef]
- Altarejos, J.Y.; Montminy, M. Creb and the Crtc Co-Activators: Sensors for Hormonal and Metabolic Signals. Nat. Rev. Mol. Cell Biol. 2011, 12, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Li, E.; Chen, L.; Zhang, Y.; Wei, F.; Liu, J.; Deng, H.; Wang, Y. The Creb Coactivator Crtc2 Controls Hepatic Lipid Metabolism by Regulating Srebp1. Nature 2015, 524, 243–246. [Google Scholar] [CrossRef]
- Sun, X.; Dang, F.; Zhang, D.; Yuan, Y.; Zhang, C.; Wu, Y.; Wang, Y.; Liu, Y.; Sun, X.; Dang, F.; et al. Glucagon-Creb/Crtc2 Signaling Cascade Regulates Hepatic Bmal1 Protein. J. Biol. Chem. 2015, 290, 2189–2197. [Google Scholar] [CrossRef] [Green Version]
- Koyanagi, S.; Kuramoto, Y.; Nakagawa, H.; Aramaki, H.; Ohdo, S.; Soeda, S.; Shimeno, H. A Molecular Mechanism Regulating Circadian Expression of Vascular Endothelial Growth Factor in Tumor Cells. Cancer Res. 2003, 63, 7277–7283. [Google Scholar] [PubMed]
- Alexander, R.K.; Liou, Y.-H.; Knudsen, N.H.; Starost, K.A.; Xu, C.; Hyde, A.L.; Liu, S.; Jacobi, D.; Liao, N.-S.; Lee, C.-H. Bmal1 Integrates Mitochondrial Metabolism and Macrophage Activation. eLife 2020, 9, e54090. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Wang, Z.; Zhang, T.; Chen, X.; Xu, H.; Wang, F.; Guo, L.; Chen, M.; Liu, K.; Wu, B. Deficiency of Intestinal Bmal1 Prevents Obesity Induced by High-Fat Feeding. Nat. Commun. 2021, 12, 5323. [Google Scholar] [CrossRef]
- Zhang, D.; Tong, X.; Arthurs, B.; Guha, A.; Rui, L.; Kamath, A.; Inoki, K.; Yin, L. Liver Clock Protein Bmal1 Promotes De Novo Lipogenesis through Insulin-Mtorc2-Akt Signaling. J. Biol. Chem. 2014, 289, 25925–25935. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhou, L.; Liu, S.; Liu, Y.; Zhao, J.; Chen, Y.; Liu, Y. Artepillin C Time−Dependently Alleviates Metabolic Syndrome in Obese Mice by Regulating CREB/CRTC2−BMAL1 Signaling. Nutrients 2023, 15, 1644. https://doi.org/10.3390/nu15071644
Wang L, Zhou L, Liu S, Liu Y, Zhao J, Chen Y, Liu Y. Artepillin C Time−Dependently Alleviates Metabolic Syndrome in Obese Mice by Regulating CREB/CRTC2−BMAL1 Signaling. Nutrients. 2023; 15(7):1644. https://doi.org/10.3390/nu15071644
Chicago/Turabian StyleWang, Lei, Lingqin Zhou, Shuai Liu, Yaxin Liu, Jia Zhao, Yaqiong Chen, and Yi Liu. 2023. "Artepillin C Time−Dependently Alleviates Metabolic Syndrome in Obese Mice by Regulating CREB/CRTC2−BMAL1 Signaling" Nutrients 15, no. 7: 1644. https://doi.org/10.3390/nu15071644