
Associations of Homocysteine, Folate, and Vitamin B12 with Osteoarthritis: A Mendelian Randomization Study

Abstract
:1. Introduction
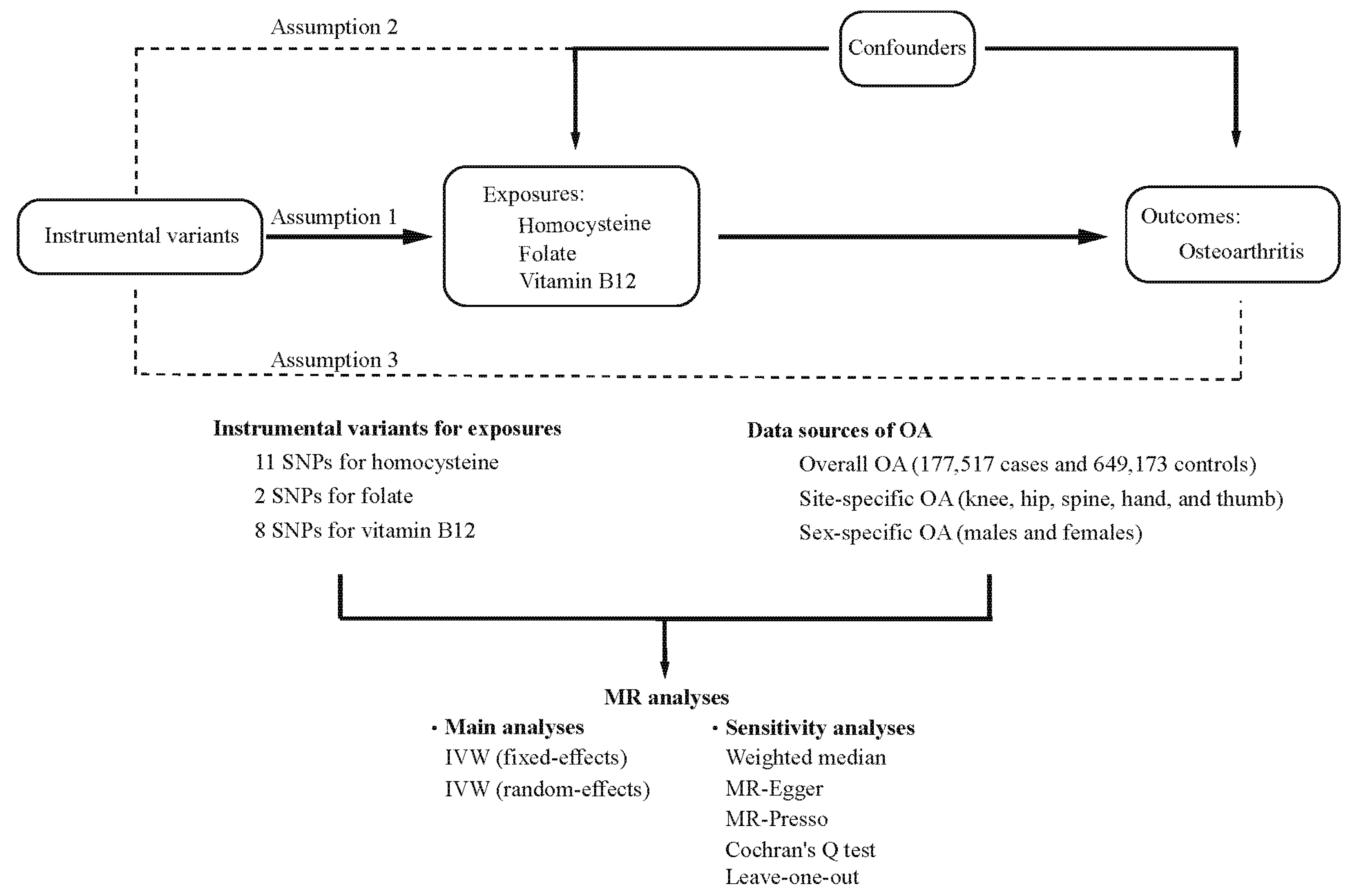
2. Materials and Methods
2.1. Study Design
2.2. Data Sources of Outcomes
2.3. Selection of Instrumental Variables
2.4. MR Analyses
3. Results
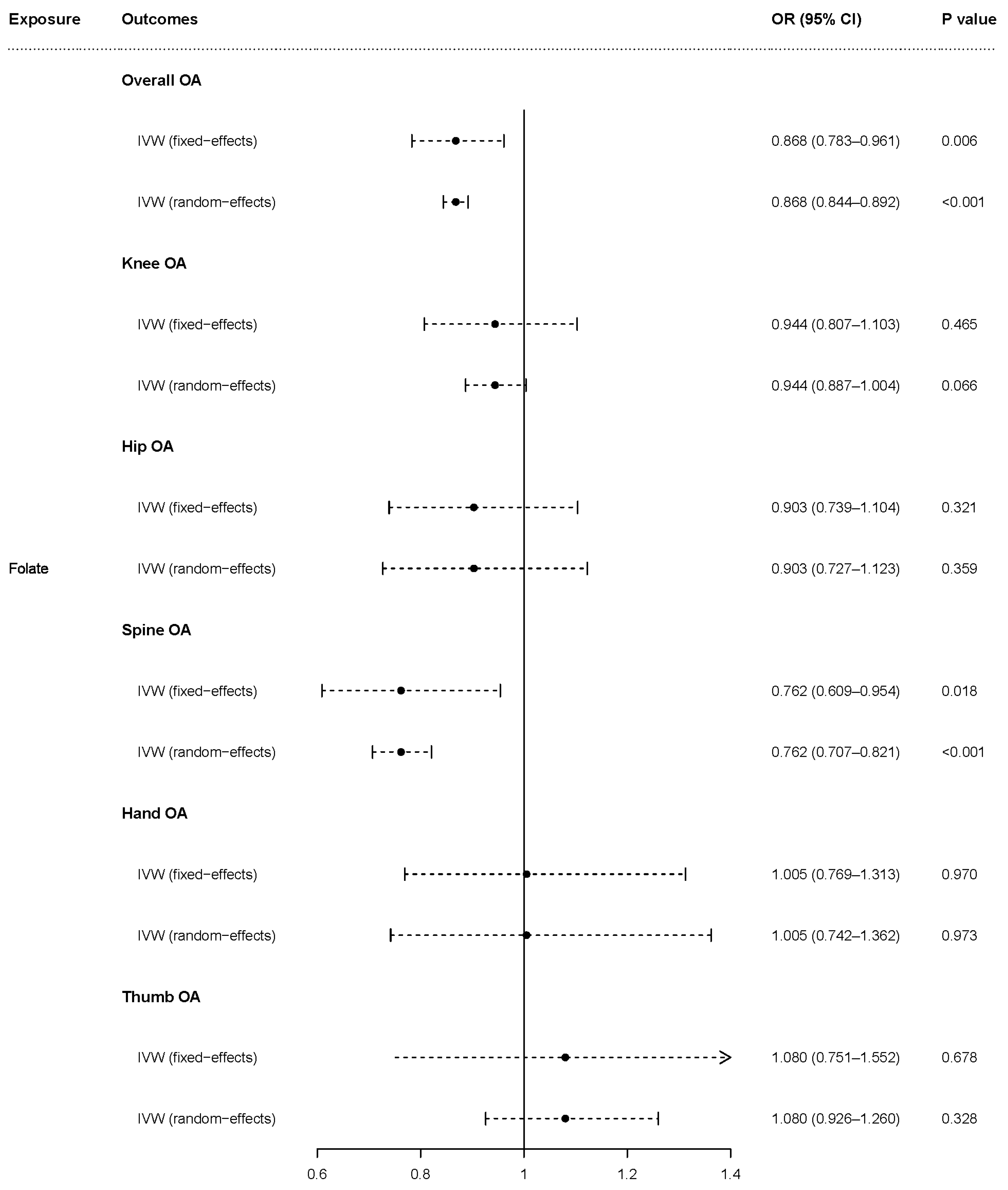
3.1. Causal Effects of Homocysteine, Folate, and Vitamin B12 on OA
3.2. Causal Effects of Homocysteine, Folate, and Vitamin B12 on OA in Females
3.3. Causal Effects of Homocysteine, Folate, and Vitamin B12 on OA in Males
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Alhambra, D.; Judge, A.; Javaid, M.K.; Cooper, C.; Diez-Perez, A.; Arden, N.K. Incidence and risk factors for clinically diagnosed knee, hip and hand osteoarthritis: Influences of age, gender and osteoarthritis affecting other joints. Ann. Rheum. Dis. 2014, 73, 1659–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.P. Osteoarthritis. Nat. Rev. Dis. Prim. 2016, 2, 16072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Fan, S.; Zhi, X.; Wang, Y.; Wang, Y.; Zheng, Q.; Sun, G. Prevalence of Hyperhomocysteinemia in China: A Systematic Review and Meta-Analysis. Nutrients 2015, 7, 74–90. [Google Scholar] [CrossRef] [Green Version]
- McLean, R.R.; Jacques, P.F.; Selhub, J.; Tucker, K.L.; Samelson, E.J.; Broe, K.E.; Hannan, M.T.; Cupples, L.A.; Kiel, D.P. Homocysteine as a Predictive Factor for Hip Fracture in Older Persons. N. Engl. J. Med. 2004, 350, 2042–2049. [Google Scholar] [CrossRef] [Green Version]
- Boushey, C.J.; Beresford, S.A.A.; Omenn, G.S.; Motulsky, A.G. A Quantitative Assessment of Plasma Homocysteine as a Risk Factor for Vascular Disease. Probable benefits of increasing folic acid intakes. JAMA 1995, 274, 1049–1057. [Google Scholar] [CrossRef]
- Koh, J.-M.; Lee, Y.-S.; Kim, Y.S.; Kim, D.J.; Kim, H.-H.; Park, J.-Y.; Lee, K.-U.; Kim, G.S. Homocysteine Enhances Bone Resorption by Stimulation of Osteoclast Formation and Activity through Increased Intracellular ROS Generation. J. Bone Miner. Res. 2006, 21, 1003–1011. [Google Scholar] [CrossRef]
- Ma, C.-H.; Chiua, Y.C.; Wu, C.-H.; Jou, I.-M.; Tu, Y.-K.; Hung, C.-H.; Hsieh, P.-L.; Tsai, K.-L. Homocysteine causes dysfunction of chondrocytes and oxidative stress through repression of SIRT1/AMPK pathway: A possible link between hyperhomocysteinemia and osteoarthritis. Redox Biol. 2018, 15, 504–512. [Google Scholar] [CrossRef]
- Nakano, M.; Nakamura, Y.; Urano, T.; Miyazaki, A.; Suzuki, T.; Watanabe, K.; Takahashi, J.; Shiraki, M. Associations of Homocysteine Metabolism with the Risk of Spinal Osteoarthritis Progression in Postmenopausal Women. J. Clin. Endocrinol. Metab. 2021, 106, 3428–3438. [Google Scholar] [CrossRef]
- Fayfman, M.; Niu, J.; Zhang, Y.; Felson, D.; Sack, B.; Aliabadi, P.; Selhub, J.; Hunter, D. The relation of plasma homocysteine to radiographic knee osteoarthritis. Osteoarthr. Cartil. 2009, 17, 766–771. [Google Scholar] [CrossRef] [Green Version]
- Shu, J.; Sun, X.; Li, J.; Li, F.; Tang, J.; Shi, L. Serum homocysteine levels and their association with clinical characteristics of inflammatory arthritis. Clin. Rheumatol. 2020, 39, 3295–3302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, H.; Zhang, Z.; Yang, F.; Chen, J. Serum Metabolites as Potential Biomarkers for Diagnosis of Knee Osteoarthritis. Dis. Markers 2015, 2015, 684794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.D.; Hemani, G. Mendelian randomization: Genetic anchors for causal inference in epidemiological studies. Hum. Mol. Genet. 2014, 23, R89–R98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.D.; Ebrahim, S. “Mendelian randomization”: Can genetic epidemiology contribute to understanding environmental determinants of disease? Int. J. Epidemiol. 2003, 32, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boer, C.G.; Hatzikotoulas, K.; Southam, L.; Stefánsdóttir, L.; Zhang, Y.; de Almeida, R.C.; Wu, T.T.; Zheng, J.; Hartley, A.; Teder-Laving, M.; et al. Deciphering osteoarthritis genetics across 826,690 individuals from 9 populations. Cell 2021, 184, 4784–4818.e17. [Google Scholar] [CrossRef]
- Van Meurs, J.B.; Pare, G.; Schwartz, S.M.; Hazra, A.; Tanaka, T.; Vermeulen, S.H.; Cotlarciuc, I.; Yuan, X.; Mälarstig, A.; Bandinelli, S.; et al. Common genetic loci influencing plasma homocysteine concentrations and their effect on risk of coronary artery disease. Am. J. Clin. Nutr. 2013, 98, 668–676. [Google Scholar] [CrossRef] [Green Version]
- Grarup, N.; Sulem, P.; Sandholt, C.H.; Thorleifsson, G.; Ahluwalia, T.S.; Steinthorsdottir, V.; Bjarnason, H.; Gudbjartsson, D.; Magnusson, O.T.; Sparsø, T.; et al. Genetic Architecture of Vitamin B12 and Folate Levels Uncovered Applying Deeply Sequenced Large Datasets. PLoS Genet. 2013, 9, e1003530. [Google Scholar] [CrossRef] [Green Version]
- Genomes Project Consortium. A map of human genome variation from population-scale sequencing. Nature 2010, 467, 1061–1073. [Google Scholar] [CrossRef] [Green Version]
- Kamat, M.A.; Blackshaw, J.A.; Young, R.; Surendran, P.; Burgess, S.; Danesh, J.; Butterworth, A.S.; Staley, J.R. PhenoScanner V2: An expanded tool for searching human genotype-phenotype associations. Bioinform. Oxf. Engl. 2019, 35, 4851–4853. [Google Scholar] [CrossRef] [Green Version]
- Hemani, G.; Tilling, K.; Davey Smith, G. Orienting the causal relationship between imprecisely measured traits using GWAS summary data. PLoS Genet. 2017, 13, e1007081. [Google Scholar]
- Hemani, G.; Zheng, J.; Elsworth, B.; Wade, K.H.; Haberland, V.; Baird, D.; Laurin, C.; Burgess, S.; Bowden, J.; Langdon, R.; et al. The MR-Base platform supports systematic causal inference across the human phenome. eLife 2018, 7, e34408. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Foley, C.N.; Zuber, V. Inferring Causal Relationships between Risk Factors and Outcomes from Genome-Wide Association Study Data. Annu. Rev. Genom. Hum. Genet. 2018, 19, 303–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartwig, F.P.; Davies, N.M.; Hemani, G.; Smith, G.D. Two-sample Mendelian randomization: Avoiding the downsides of a powerful, widely applicable but potentially fallible technique. Int. J. Epidemiol. 2016, 45, 1717–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef] [Green Version]
- Bowden, J.; Smith, G.D.; Haycock, P.C.; Burgess, S. Consistent Estimation in Mendelian Randomization with Some Invalid Instruments Using a Weighted Median Estimator. Genet. Epidemiol. 2016, 40, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.P.T.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbanck, M.; Chen, C.-Y.; Neale, B.; Do, R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 2018, 50, 693–698. [Google Scholar] [CrossRef]
- Herrmann, M.; Tami, A.; Wildemann, B.; Wolny, M.; Wagner, A.; Schorr, H.; Taban-Shomal, O.; Umanskaya, N.; Ross, S.; Garcia, P.; et al. Hyperhomocysteinemia induces a tissue specific accumulation of homocysteine in bone by collagen binding and adversely affects bone. Bone 2009, 44, 467–475. [Google Scholar] [CrossRef]
- Zhao, Z.; Tan, Q.; Jiang, A.; Sun, S.; Liu, Z.; Li, W.; Song, C.; Leng, H. Evidence of subchondral bone’s effects on articular cartilage damage in OVX-OA rat. Eng. Fract. Mech. 2020, 233, 107081. [Google Scholar] [CrossRef]
- Jiang, A.; Xu, P.; Yang, Z.; Zhao, Z.; Tan, Q.; Li, W.; Song, C.; Dai, H.; Leng, H. Increased Sparc release from subchondral osteoblasts promotes articular chondrocyte degeneration under estrogen withdrawal. Osteoarthr. Cartil. 2022, 31, 26–38. [Google Scholar] [CrossRef]
- Hosnijeh, F.S.; Kavousi, M.; Boer, C.; Uitterlinden, A.; Hofman, A.; Reijman, M.; Oei, E.; Bierma-Zeinstra, S.; van Meurs, J. Development of a prediction model for future risk of radiographic hip osteoarthritis. Osteoarthr. Cartil. 2018, 26, 540–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacGregor, A.J.; Li, Q.; Spector, T.D.; Williams, F. The genetic influence on radiographic osteoarthritis is site specific at the hand, hip and knee. Rheumatology 2009, 48, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Bijsterbosch, J.; Meulenbelt, I.; Watt, I.; Rosendaal, F.R.; Huizinga, T.W.J.; Kloppenburg, M. Clustering of hand osteoarthritis progression and its relationship to progression of osteoarthritis at the knee. Ann. Rheum. Dis. 2014, 73, 567–572. [Google Scholar] [CrossRef]
- Stürmer, T.; Günther, K.-P.; Brenner, H. Obesity, overweight and patterns of osteoarthritis: The Ulm Osteoarthritis Study. J. Clin. Epidemiol. 2000, 53, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Mijatovic, V.; Kenemans, P.; Jakobs, C.; van Baal, W.M.; Peters-Muller, E.R.; van der Mooren, M.J. A randomized controlled study of the effects of 17beta-estradiol-dydrogesterone on plasma homocysteine in postmenopausal women. Obstet. Gynecol. 1998, 91, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, N.W. Mass spectrometry evidence for formation of estrogen–homocysteine conjugates: Estrogens can regulate homocysteine levels. Free. Radic. Biol. Med. 2013, 65, 1447–1454. [Google Scholar] [CrossRef]
- Castañeda, S.; Vicente-Rabaneda, E.F. Disentangling the molecular interplays between subchondral bone and articular cartilage in estrogen deficiency-induced osteoarthritis. Osteoarthr. Cartil. 2023, 31, 6–8. [Google Scholar] [CrossRef]
- Xu, H.; Shin, M.-H.; Kang, J.-H.; Choi, S.-E.; Park, D.-J.; Kweon, S.-S.; Lee, Y.-H.; Kim, H.-Y.; Lee, J.-K.; Lee, S.-S. Folate deficiency is associated with increased radiographic severity of osteoarthritis in knee joints but not in hand joints. Clin. Exp. Rheumatol. 2022; ahead of print. [Google Scholar] [CrossRef]
- Amirkhizi, F.; Ghoreishy, S.M.; Hamedi-Shahraki, S.; Asghari, S. Higher dietary phytochemical index is associated with lower odds of knee osteoarthritis. Sci. Rep. 2022, 12, 9059. [Google Scholar] [CrossRef]
- Flynn, M.A.; Irvin, W.; Krause, G. The effect of folate and cobalamin on osteoarthritic hands. J. Am. Coll. Nutr. 1994, 13, 351–356. [Google Scholar] [CrossRef]
- Fu, L.; Wang, Y.; Hu, Y.-Q. Causal effects of B vitamins and homocysteine on obesity and musculoskeletal diseases: A Mendelian randomization study. Front. Nutr. 2022, 9, 2741. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | SNP | Chromosome:Position | Gene | EA | OA | EAF | Beta | SE | p |
|---|---|---|---|---|---|---|---|---|---|
| Homocysteine | rs957140 | 11:89201627 | NOX4 | A | G | 0.45 | −0.045 | 0.008 | 2.43 × 10−10 |
| rs9369898 | 6:49382193 | MUT | A | G | 0.62 | 0.0449 | 0.007 | 2.17 × 10−10 | |
| rs7130284 | 11:89148372 | NOX4 | T | C | 0.07 | −0.1242 | 0.013 | 1.88 × 10−20 | |
| rs4660306 | 1:45978675 | MMACHC | T | C | 0.33 | 0.0435 | 0.007 | 2.33 × 10−9 | |
| rs42648 | 7:89977760 | GTPBP10 | A | G | 0.40 | −0.0395 | 0.007 | 1.97 × 10−8 | |
| rs2851391 | 21:44487404 | CBS | T | C | 0.47 | 0.056 | 0.008 | 1.70 × 10−12 | |
| rs234709 | 21:44486964 | CBS | T | C | 0.45 | −0.0718 | 0.007 | 3.90 × 10−24 | |
| rs2275565 | 1:237048676 | MTR | T | G | 0.21 | −0.0542 | 0.009 | 1.96 × 10−10 | |
| rs1801222 | 10:17156151 | CUBN | A | G | 0.34 | 0.0453 | 0.007 | 8.43 × 10−10 | |
| rs12780845 | 10:17223244 | CUBN | A | G | 0.65 | 0.0529 | 0.009 | 7.80 × 10−10 | |
| rs12134663 | 1:11838646 | MTHFR | A | C | 0.80 | −0.101 | 0.011 | 2.54 × 10−21 | |
| Folate | rs17421511 | 1:11857788 | MTHFR | G | A | 0.827 | 0.098 | 0.012 | 1.80 × 10−15 |
| rs652197 | 11:71849741 | FOLR3 | C | T | 0.179 | 0.069 | 0.011 | 2.50 × 10−10 | |
| Vitamin B12 | rs5753231 | 22:31003069 | TCN2 | C | T | 0.790 | 0.064 | 0.010 | 7.50 × 10−10 |
| rs56077122 | 10:17207015 | TRDMT1 | A | C | 0.335 | 0.087 | 0.009 | 4.80 × 10−21 | |
| rs41281112 | 13:100518634 | CLYBL | C | T | 0.948 | 0.17 | 0.016 | 9.60 × 10−27 | |
| rs34528912 | 11:59631535 | TCN1 | T | C | 0.036 | 0.17 | 0.021 | 2.10 × 10−15 | |
| rs2336573 | 19:8367709 | CD320 | T | C | 0.031 | 0.32 | 0.021 | 1.10 × 10−51 | |
| rs1801222 | 10:17156151 | CUBN | G | A | 0.593 | 0.11 | 0.007 | 1.10 × 10−52 | |
| rs117456053 | 11:59616831 | TCN1 | G | A | 0.976 | 0.16 | 0.027 | 1.90 × 10−9 | |
| rs1141321 | 6:49412433 | MUT | C | T | 0.627 | 0.061 | 0.007 | 1.40 × 10−16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, H.; Chen, L.; Zhong, Y.; Yang, Z.; Li, W.; Song, C.; Leng, H. Associations of Homocysteine, Folate, and Vitamin B12 with Osteoarthritis: A Mendelian Randomization Study. Nutrients 2023, 15, 1636. https://doi.org/10.3390/nu15071636
Hong H, Chen L, Zhong Y, Yang Z, Li W, Song C, Leng H. Associations of Homocysteine, Folate, and Vitamin B12 with Osteoarthritis: A Mendelian Randomization Study. Nutrients. 2023; 15(7):1636. https://doi.org/10.3390/nu15071636
Chicago/Turabian StyleHong, Haofeng, Longting Chen, Yiming Zhong, Zihuan Yang, Weishi Li, Chunli Song, and Huijie Leng. 2023. "Associations of Homocysteine, Folate, and Vitamin B12 with Osteoarthritis: A Mendelian Randomization Study" Nutrients 15, no. 7: 1636. https://doi.org/10.3390/nu15071636