
Ketone Body β-Hydroxy-Butyrate Sustains Progressive Motility in Capacitated Human Spermatozoa: A Possible Role in Natural Fertility
 , ,
, , {kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Semen Samples
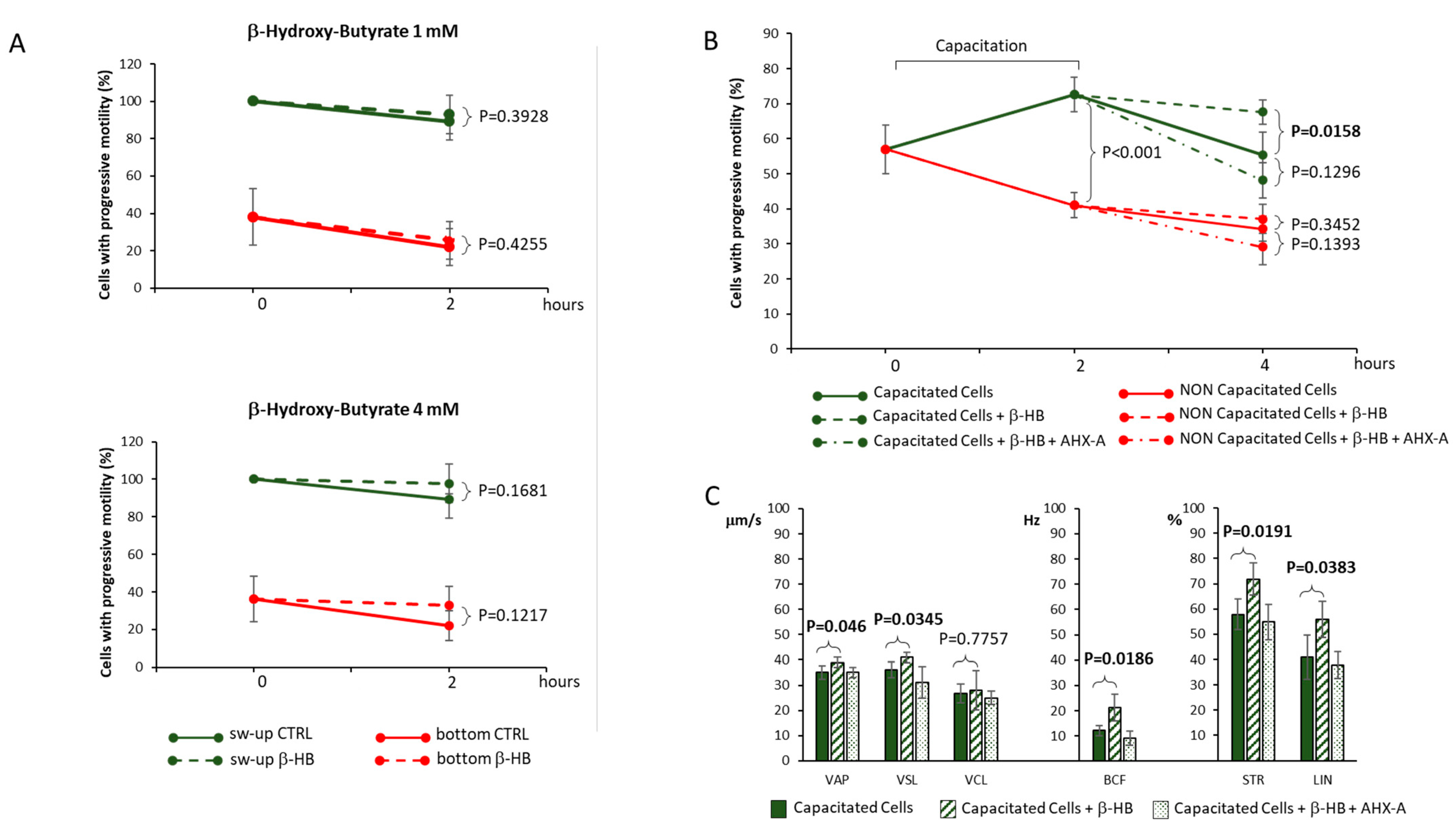
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brostow, D.P.; Hirsch, A.T.; Collins, T.C.; Kurzer, M.S. The Role of Nutrition and Body Composition in Peripheral Arterial Disease. Nat. Rev. Cardiol. 2012, 9, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.M. Variation of Leptin During Menstrual Cycle and Its Relation to the Hypothalamic–Pituitary–Gonadal (HPG) Axis: A Systematic Review. Int. J. Women’s Health 2021, 13, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Pascoal, G.d.F.L.; Geraldi, M.V.; Maróstica, M.R.; Ong, T.P. Effect of Paternal Diet on Spermatogenesis and Offspring Health: Focus on Epigenetics and Interventions with Food Bioactive Compounds. Nutrients 2022, 14, 2150. [Google Scholar] [CrossRef] [PubMed]
- Pini, T.; Raubenheimer, D.; Simpson, S.J.; Crean, A.J. Obesity and Male Reproduction; Placing the Western Diet in Context. Front. Endocrinol. 2021, 12, 622292. [Google Scholar] [CrossRef]
- Wei, W.; Zhang, X.; Zhou, B.; Ge, B.; Tian, J.; Chen, J. Effects of Female Obesity on Conception, Pregnancy and the Health of Offspring. Front. Endocrinol. 2022, 13, 949228. [Google Scholar] [CrossRef]
- Erdem, N.Z.; Ozelgun, D.; Taskin, H.E.; Avsar, F.M. Comparison of a Pre-Bariatric Surgery Very Low-Calorie Ketogenic Diet and the Mediterranean Diet Effects on Weight Loss, Metabolic Parameters, and Liver Size Reduction. Sci. Rep. 2022, 12, 20686. [Google Scholar] [CrossRef]
- Drabińska, N.; Wiczkowski, W.; Piskuła, M.K. Recent Advances in the Application of a Ketogenic Diet for Obesity Management. Trends Food Sci. Technol. 2021, 110, 28–38. [Google Scholar] [CrossRef]
- Laffel, L. Ketone Bodies: A Review of Physiology, Pathophysiology and Application of Monitoring to Diabetes. Diabetes Metab. Res. Rev. 1999, 15, 412–426. [Google Scholar] [CrossRef]
- Shilpa, J.; Mohan, V. Ketogenic Diets: Boon or Bane? Indian J. Med. Res. 2018, 148, 251. [Google Scholar] [CrossRef]
- Lee, P.R.; Kossoff, E.H. Dietary Treatments for Epilepsy: Management Guidelines for the General Practitioner. Epilepsy Behav. 2011, 21, 115–121. [Google Scholar] [CrossRef]
- Furini, C.; Spaggiari, G.; Simoni, M.; Greco, C.; Santi, D. Ketogenic State Improves Testosterone Serum Levels—Results from a Systematic Review and Meta-Analysis. Endocrine 2022, 79, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Corona, G.; Rastrelli, G.; Morelli, A.; Sarchielli, E.; Cipriani, S.; Vignozzi, L.; Maggi, M. Treatment of Functional Hypogonadism Besides Pharmacological Substitution. World J. Men’s Health 2020, 38, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultan, S.; Patel, A.G.; El-Hassani, S.; Whitelaw, B.; Leca, B.M.; Vincent, R.P.; le Roux, C.W.; Rubino, F.; Aywlin, S.J.B.; Dimitriadis, G.K. Male Obesity Associated Gonadal Dysfunction and the Role of Bariatric Surgery. Front. Endocrinol. 2020, 11, 408. [Google Scholar] [CrossRef] [PubMed]
- Mongioì, L.M.; Cimino, L.; Condorelli, R.A.; Magagnini, M.C.; Barbagallo, F.; Cannarella, R.; la Vignera, S.; Calogero, A.E. Effectiveness of a Very Low Calorie Ketogenic Diet on Testicular Function in Overweight/Obese Men. Nutrients 2020, 12, 2967. [Google Scholar] [CrossRef]
- Cignarelli, A.; Santi, D.; Genchi, V.A.; Conte, E.; Giordano, F.; di Leo, S.; Natalicchio, A.; Laviola, L.; Giorgino, F.; Perrini, S. Very Low-calorie Ketogenic Diet Rapidly Augments Testosterone Levels in Non-diabetic Obese Subjects. Andrology 2023, 11, 234–244. [Google Scholar] [CrossRef]
- Huang, K.; Thomas, N.; Gooley, P.R.; Armstrong, C.W. Systematic Review of NMR-Based Metabolomics Practices in Human Disease Research. Metabolites 2022, 12, 963. [Google Scholar] [CrossRef] [PubMed]
- Blaurock, J.; Baumann, S.; Grunewald, S.; Schiller, J.; Engel, K.M. Metabolomics of Human Semen: A Review of Different Analytical Methods to Unravel Biomarkers for Male Fertility Disorders. Int. J. Mol. Sci. 2022, 23, 9031. [Google Scholar] [CrossRef]
- Castiglione Morelli, M.A.; Iuliano, A.; Schettini, S.C.A.; Petruzzi, D.; Ferri, A.; Colucci, P.; Viggiani, L.; Cuviello, F.; Ostuni, A. NMR Metabolic Profiling of Follicular Fluid for Investigating the Different Causes of Female Infertility: A Pilot Study. Metabolomics 2019, 15, 19. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; World Health Organization: Geneva, Switzerland, 2010; ISBN 9789241547789.
- Garolla, A.; de Toni, L.; Menegazzo, M.; Foresta, C. Caution in the Use of Standard Sperm-Washing Procedures for Assisted Reproduction in HPV-Infected Patients. Reprod. Biomed. Online 2020, 41, 967–968. [Google Scholar] [CrossRef]
- Walker, J.M. Part V Sperm Preparation and Selection Techniques. In Methods in Molecular Biology Series; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Signorelli, J.; Diaz, E.S.; Morales, P. Kinases, Phosphatases and Proteases during Sperm Capacitation. Cell Tissue Res. 2012, 349, 765–782. [Google Scholar] [CrossRef]
- Saris, C.G.J.; Timmers, S. Ketogenic Diets and Ketone Suplementation: A Strategy for Therapeutic Intervention. Front. Nutr. 2022, 9, 947567. [Google Scholar] [CrossRef] [PubMed]
- Calcaterra, V.; Verduci, E.; Pascuzzi, M.C.; Magenes, V.C.; Fiore, G.; di Profio, E.; Tenuta, E.; Bosetti, A.; Todisco, C.F.; D’Auria, E.; et al. Metabolic Derangement in Pediatric Patient with Obesity: The Role of Ketogenic Diet as Therapeutic Tool. Nutrients 2021, 13, 2805. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Moehl, K.; Ghena, N.; Schmaedick, M.; Cheng, A. Intermittent Metabolic Switching, Neuroplasticity and Brain Health. Nat. Rev. Neurosci. 2018, 19, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Gomes, C.P.C.; Almeida, J.A.; Franco, O.L.; Petriz, B. Omics and the Molecular Exercise Physiology. Adv. Clin. Chem. 2020, 96, 55–84. [Google Scholar] [CrossRef] [PubMed]
- Sherwin, R.S.; Hendler, R.G.; Felig, P. Effect of Ketone Infusions on Amino Acid and Nitrogen Metabolism in Man. J. Clin. Investig. 1975, 55, 1382–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, K.S.; Welle, S.L.; Halliday, D.; Campbell, R.G. Effect of Beta-Hydroxybutyrate on Whole-Body Leucine Kinetics and Fractional Mixed Skeletal Muscle Protein Synthesis in Humans. J. Clin. Investig. 1988, 82, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.; Cogan, K.E.; Egan, B. Metabolism of Ketone Bodies during Exercise and Training: Physiological Basis for Exogenous Supplementation. J. Physiol. 2017, 595, 2857–2871. [Google Scholar] [CrossRef] [Green Version]
- Yakupova, E.I.; Bocharnikov, A.D.; Plotnikov, E.Y. Effects of Ketogenic Diet on Muscle Metabolism in Health and Disease. Nutrients 2022, 14, 3842. [Google Scholar] [CrossRef]
- Carneiro, L.; Pellerin, L. Nutritional Impact on Metabolic Homeostasis and Brain Health. Front. Neurosci. 2022, 15, 767405. [Google Scholar] [CrossRef]
- Zhao, H.; Zhao, Y.; Li, T.; Li, M.; Li, J.; Li, R.; Liu, P.; Yu, Y.; Qiao, J. Metabolism Alteration in Follicular Niche: The Nexus among Intermediary Metabolism, Mitochondrial Function, and Classic Polycystic Ovary Syndrome. Free Radic. Biol. Med. 2015, 86, 295–307. [Google Scholar] [CrossRef]
- Leroy, J.L.M.R.; Vanholder, T.; Delanghe, J.R.; Opsomer, G.; van Soom, A.; Bols, P.E.J.; Dewulf, J.; de Kruif, A. Metabolic Changes in Follicular Fluid of the Dominant Follicle in High-Yielding Dairy Cows Early Post Partum. Theriogenology 2004, 62, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- Missio, D.; dos Santos Brum, D.; Dalle Laste Dacampo, L.; Weber Santos Cibin, F.; Silveira Mesquita, F.; Germano Ferst, J.; Fiordalisi, G.; Dias Gonçalves, P.B.; Ferreira, R. High Concentrations of Β-hydroxybutyrate Alter the Kinetics of Bovine Spermatozoa. Andrologia 2021, 53, e14148. [Google Scholar] [CrossRef] [PubMed]
- Pocate-Cheriet, K.; Santulli, P.; Kateb, F.; Bourdon, M.; Maignien, C.; Batteux, F.; Chouzenoux, S.; Patrat, C.; Wolf, J.P.; Bertho, G.; et al. The Follicular Fluid Metabolome Differs According to the Endometriosis Phenotype. Reprod. Biomed. Online 2020, 41, 1023–1037. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Iguchi, N.; Miyagawa, Y.; Koga, M.; Kohroki, J.; Nishimune, Y. Differential Expression of Succinyl CoA Transferase (SCOT) Genes in Somatic and Germline Cells of the Mouse Testis. Int. J. Androl. 2003, 26, 52–56. [Google Scholar] [CrossRef]
- Zaferani, M.; Suarez, S.S.; Abbaspourrad, A. Mammalian Sperm Hyperactivation Regulates Navigation via Physical Boundaries and Promotes Pseudo-Chemotaxis. Proc. Natl. Acad. Sci. USA 2021, 118, e2107500118. [Google Scholar] [CrossRef]
- Miki, K.; Clapham, D.E. Rheotaxis Guides Mammalian Sperm. Curr. Biol. 2013, 23, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Ramal-Sanchez, M.; Bernabò, N.; Valbonetti, L.; Cimini, C.; Taraschi, A.; Capacchietti, G.; Machado-Simoes, J.; Barboni, B. Role and Modulation of TRPV1 in Mammalian Spermatozoa: An Updated Review. Int. J. Mol. Sci. 2021, 22, 4306. [Google Scholar] [CrossRef]
- Han, S.; Chu, X.-P.; Goodson, R.; Gamel, P.; Peng, S.; Vance, J.; Wang, S. Cholesterol Inhibits Human Voltage-Gated Proton Channel HHv1. Proc. Natl. Acad. Sci. USA 2022, 119, e2205420119. [Google Scholar] [CrossRef]
- Matamoros-Volante, A.; Treviño, C.L. Capacitation-Associated Alkalization in Human Sperm Is Differentially Controlled at the Subcellular Level. J. Cell Sci. 2020, 133, jcs238816. [Google Scholar] [CrossRef]
- Boguenet, M.; Bouet, P.-E.; Spiers, A.; Reynier, P.; May-Panloup, P. Mitochondria: Their Role in Spermatozoa and in Male Infertility. Hum. Reprod. Update 2021, 27, 697–719. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pappalardo, C.; Finocchi, F.; Pedrucci, F.; Di Nisio, A.; Ferlin, A.; De Toni, L.; Foresta, C. Ketone Body β-Hydroxy-Butyrate Sustains Progressive Motility in Capacitated Human Spermatozoa: A Possible Role in Natural Fertility. Nutrients 2023, 15, 1622. https://doi.org/10.3390/nu15071622
Pappalardo C, Finocchi F, Pedrucci F, Di Nisio A, Ferlin A, De Toni L, Foresta C. Ketone Body β-Hydroxy-Butyrate Sustains Progressive Motility in Capacitated Human Spermatozoa: A Possible Role in Natural Fertility. Nutrients. 2023; 15(7):1622. https://doi.org/10.3390/nu15071622
Chicago/Turabian StylePappalardo, Claudia, Federica Finocchi, Federica Pedrucci, Andrea Di Nisio, Alberto Ferlin, Luca De Toni, and Carlo Foresta. 2023. "Ketone Body β-Hydroxy-Butyrate Sustains Progressive Motility in Capacitated Human Spermatozoa: A Possible Role in Natural Fertility" Nutrients 15, no. 7: 1622. https://doi.org/10.3390/nu15071622