
Effects of Early Nutrition Factors on Baseline Neurodevelopment during the First 6 Months of Life: An EEG Study
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Anthropometrics and Behavioral Assessments
2.3. EEG Recordings and Signal Processing
2.4. Statistical Analyses
3. Results
3.1. Anthropometrics and Behavioral Assessments
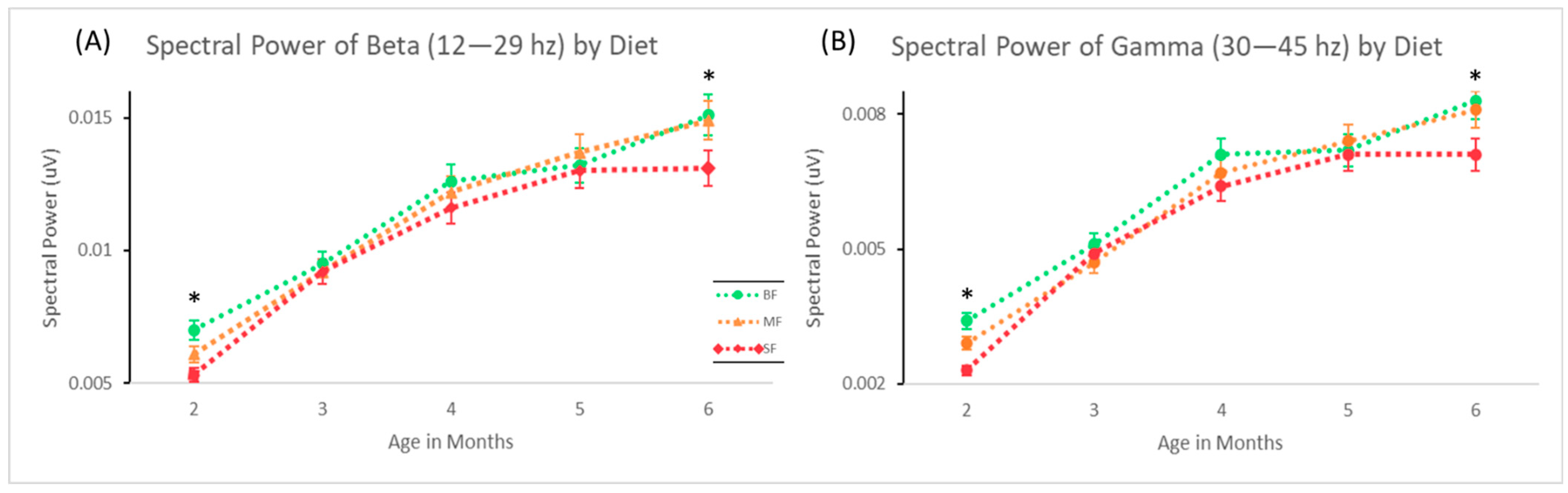
3.2. Spectral Power in Sensor Space
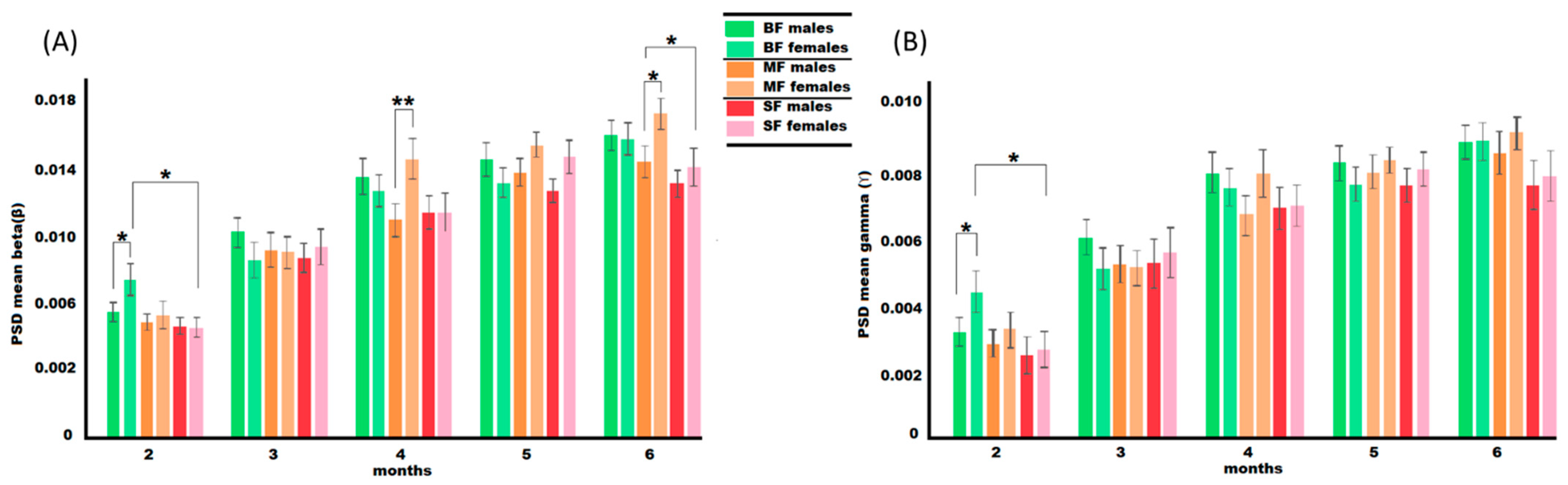
3.3. Spectral Power Covariate Analysis in Sensor Space
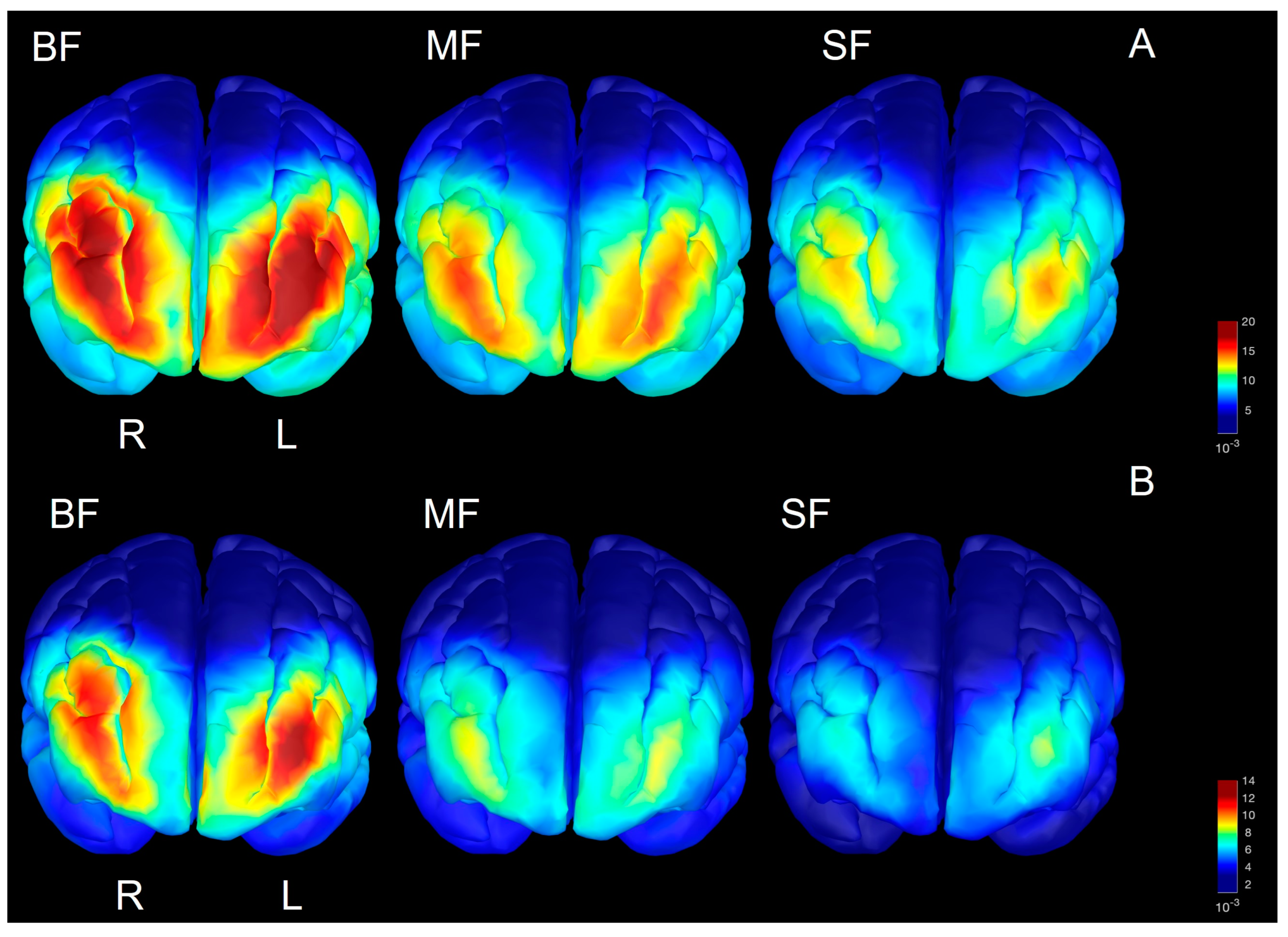
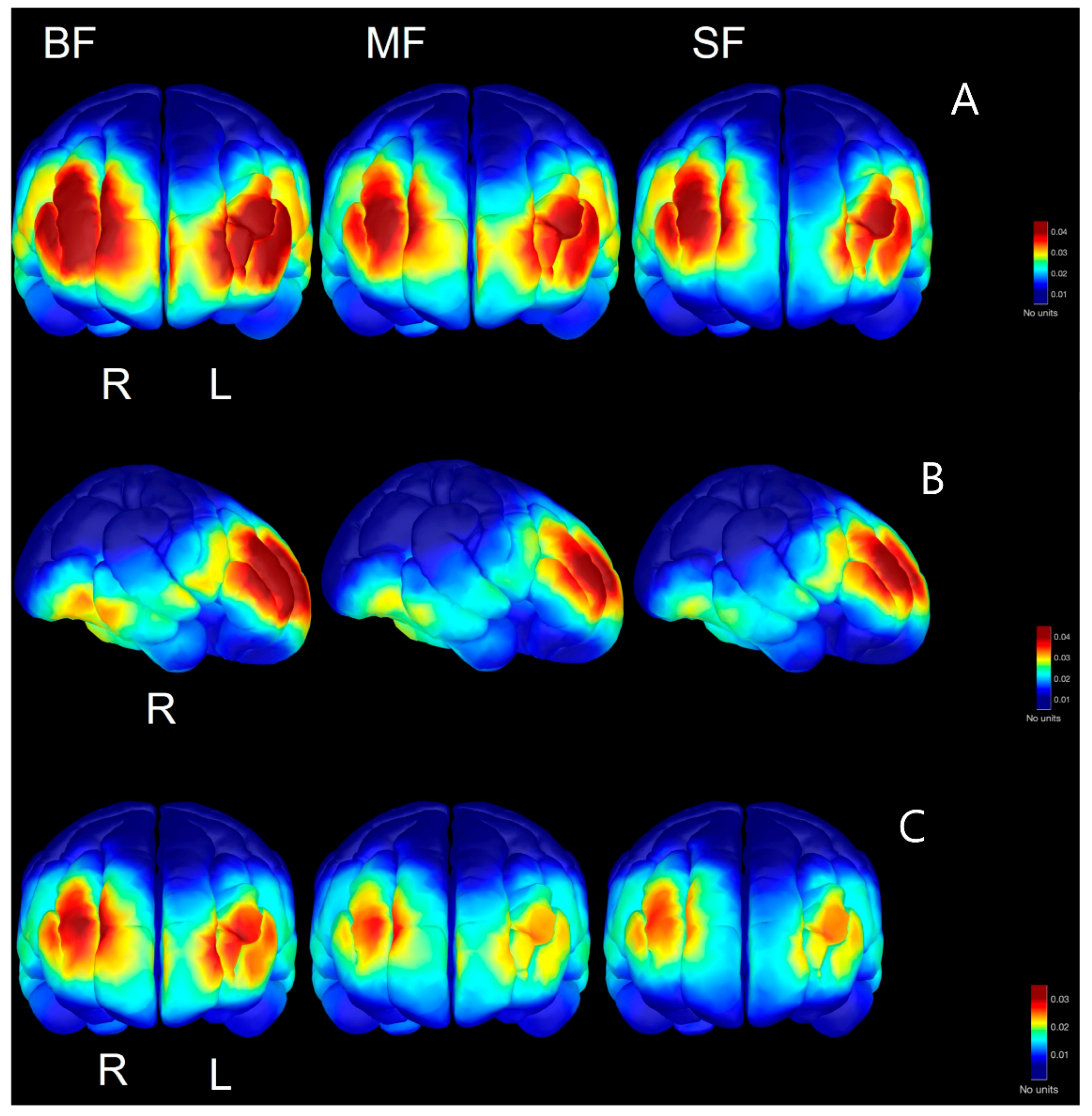
3.4. Spectral Power in Source Space
4. Discussion
4.1. Age-Related Development of Higher Frequencies
4.2. Regional Development of Beta/Gamma Defined in Source Space
4.3. Dietary Effects on Neurocognitive Testing
4.4. Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cusick, S.E.; Georgieff, M.K. The Role of Nutrition in Brain Development: The Golden Opportunity of the “First 1000 Days” Brain Development in Late Fetal and Early Postnatal Life. J. Pediatr. 2016, 175, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Lockyer, F.; McCann, S.; Moore, S.E. Breast Milk Micronutrients and Infant Neurodevelopmental Outcomes: A Systematic Review. Nutrients 2021, 13, 3848. [Google Scholar] [CrossRef]
- Innis, S.M. Dietary Omega 3 Fatty Acids and the Developing Brain. Brain Res. 2008, 1237, 35–43. [Google Scholar] [CrossRef]
- Deoni, S.; Dean, D.; Joelson, S.; O’Regan, J.; Schneider, N. Early Nutrition Influences Developmental Myelination and Cognition in Infants and Young Children. Neuroimage 2018, 178, 649–659. [Google Scholar] [CrossRef]
- Ottolini, K.M.; Andescavage, N.; Keller, S.; Limperopoulos, C. Nutrition and the Developing Brain: The Road to Optimizing Early Neurodevelopment: A Systematic Review. Pediatr. Res. 2020, 87, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.; Morley, R.; Isaacs, E. Nutrition and Mental Development. Nutr. Rev. 2001, 59, S24–S33. [Google Scholar] [CrossRef]
- Kafouri, S.; Kramer, M.; Leonard, G.; Perron, M.; Pike, B.; Richer, L.; Toro, R.; Veillette, S.; Pausova, Z.; Paus, T. Breastfeeding and Brain Structure in Adolescence. Int. J. Epidemiol. 2013, 42, 150–159. [Google Scholar] [CrossRef]
- Lozoff, B. Early Iron Deficiency Has Brain and Behavior Effects Consistent with Dopaminergic Dysfunction1–3. J. Nutr. 2011, 141, 740S–746S. [Google Scholar] [CrossRef] [Green Version]
- Georgieff, M.K.; Innis, S.M. Preterm Neurodevelopment : Essential Fatty Acids and Iron. Pediatr. Res. 2005, 57, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, J.; Greer, F. Use of Soy Protein-Based Formulas in Infant Feeding. Pediatrics 2008, 121, 1062–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meek, J.Y.; Noble, L. Policy Statement: Breastfeeding and the Use of Human Milk. Pediatrics 2022, 150, e2022057988. [Google Scholar] [CrossRef]
- Prado, E.L.; Dewey, K.G. Nutrition and Brain Development in Early Life. Nutr. Rev. 2014, 72, 267–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andres, A.; Cleves, M.A.; Bellando, J.B.; Pivik, R.T.; Casey, P.H.; Badger, T.M. Developmental Status of 1-Year-Old Infants Fed Breast Milk, Cow’s Milk Formula, or Soy Formula. Pediatrics 2012, 129, 1134–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deoni, S.C.L.; Dean, D.C., 3rd; Piryatinsky, I.; O’Muircheartaigh, J.; Waskiewicz, N.; Lehman, K.; Han, M.; Dirks, H. Breastfeeding and Early White Matter Development: A Cross-Sectional Study. NeuroImage 2013, 82, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Kramer, M.S.; Aboud, F.; Mironova, E.; Vanilovich, I.; Platt, R.W.; Matush, L.; Igumnov, S.; Fombonne, E.; Bogdanovich, N.; Ducruet, T.; et al. Breastfeeding and Child Cognitive Development. Am. Med. Assoc. 2008, 65, 578. [Google Scholar] [CrossRef] [Green Version]
- Victora, C.G.; Barros, F.C.; Horta, B.L.; Lima, R.C. Breastfeeding and School Achievement in Brazilian Adolescents. Acta Paediatr. Int. J. Paediatr. 2005, 94, 1656–1660. [Google Scholar] [CrossRef]
- Brion, M.J.A.; Lawlor, D.A.; Matijasevich, A.; Horta, B.; Anselmi, L.; Araújo, C.L.; Menezes, A.M.B.; Victora, C.G.; Smith, G.D. What Are the Causal Effects of Breastfeeding on IQ, Obesity and Blood Pressure? Evidence from Comparing High-Income with Middle-Income Cohorts. Int. J. Epidemiol. 2011, 40, 670–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luby, J.L.; Belden, A.C.; Whalen, D.; Harms, M.P.; Barch, D.M. Breastfeeding and Childhood IQ: The Mediating Role of Gray Matter Volume. J. Am. Acad. Child Adolesc. Psychiatry 2016, 55, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Ou, X.; Andres, A.; Pivik, R.T.; Cleves, M.A.; Snow, J.H.; Ding, Z.; Badger, T.M. Voxel-Based Morphometry and FMRI Revealed Differences in Brain Gray Matter in Breastfed and Milk Formula-Fed Children. Am. J. Neuroradiol. 2016, 37, 713–719. [Google Scholar] [CrossRef] [Green Version]
- Buzsáki, G. Rhythms of the Brain; Oxford University Press: Oxford, UK, 2006. [Google Scholar] [CrossRef] [Green Version]
- Begus, K.; Bonawitz, E. The Rhythm of Learning: Theta Oscillations as an Index of Active Learning in Infancy. Dev. Cogn. Neurosci. 2020, 45, 100810. [Google Scholar] [CrossRef]
- Köster, M.; Langeloh, M.; Hoehl, S. Visually Entrained Theta Oscillations Increase for Unexpected Events in the Infant Brain. Psychol. Sci. 2019, 30, 1656–1663. [Google Scholar] [CrossRef]
- Pivik, R.T.; Andres, A.; Badger, T.M. Effects of Diet on Early Stage Cortical Perception and Discrimination of Syllables Differing in Voice-Onset Time : A Longitudinal ERP Study in 3 and 6 Month Old Infants. Brain Lang. 2012, 120, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Norton, E.S.; MacNeill, L.A.; Harriott, E.M.; Allen, N.; Krogh-Jespersen, S.; Smyser, C.D.; Rogers, C.E.; Smyser, T.A.; Luby, J.; Wakschlag, L. EEG/ERP as a Pragmatic Method to Expand the Reach of Infant-Toddler Neuroimaging in HBCD: Promises and Challenges. Dev. Cogn. Neurosci. 2021, 51, 100988. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods Find. Exp. Clin. Pharmacol. 2002, 24 (Suppl. D), 5–12. [Google Scholar] [PubMed]
- Khazipov, R.; Milh, M. Early Patterns of Activity in the Developing Cortex: Focus on the Sensorimotor System. Semin. Cell Dev. Biol. 2018, 76, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Egorov, A.V.; Draguhn, A. Development of Coherent Neuronal Activity Patterns in Mammalian Cortical Networks: Common Principles and Local Hetereogeneity. Mech. Dev. 2013, 130, 412–423. [Google Scholar] [CrossRef]
- Gasser, T.; Verleger, R.; Bächer, P.; Sroka, L. Development of the EEG of School-Age Children and Adolescents. I. Analysis of Band Power. Electroencephalogr. Clin. Neurophysiol. 1988, 69, 91–99. [Google Scholar] [CrossRef]
- Saby, J.N.; Marshall, P.J. The Utility of EEG Band Power Analysis in the Study of Infancy and Early Childhood. Dev. Neuropsychol. 2012, 37, 253–273. [Google Scholar] [CrossRef] [Green Version]
- Stroganova, T.A.; Orekhova, E.V.; Posikera, I.N. EEG Alpha Rhythm in Infants. Clin. Neurophysiol. 1999, 110, 997–1012. [Google Scholar] [CrossRef]
- Csibra, G.; Davis, G.; Spratling, M.W.; Johnson, M.H. Gamma Oscillations and Object Processing in the Infant Brain. Science 2000, 290, 1582–1585. [Google Scholar] [CrossRef] [Green Version]
- Gilley, P.M.; Uhler, K.; Watson, K.; Yoshinaga-Itano, C. Spectral-Temporal EEG Dynamics of Speech Discrimination Processing in Infants during Sleep. BMC Neurosci. 2017, 18, 34. [Google Scholar] [CrossRef] [Green Version]
- Jing, H.; Gilchrist, J.M.; Badger, T.M.; Pivik, R.T. A Longitudinal Study of Differences in Electroencephalographic Activity among Breastfed, Milk Formula-Fed, and Soy Formula-Fed Infants during the First Year of Life. Early Hum. Dev. 2010, 86, 119–125. [Google Scholar] [CrossRef]
- Andres, A.; Casey, P.H.; Cleves, M.A.; Badger, T.M. Body Fat and Bone Mineral Content of Infants Fed Breast Milk, Cow’s Milk Formula, or Soy Formula during the First Year of Life. J. Pediatr. 2013, 163, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Bayley, N. Bayley Scales of Infant and Toddler Development, 2nd ed.; Administration Manual; Harcourt Assessment: San Antonio, TX, USA, 1993. [Google Scholar]
- Wechsler, D. Wechsler Abbreviated Scale of Intelligence; The Psychological Corporation: San Antonio, TX, USA, 1999. [Google Scholar]
- Gabard-Durnam, L.J.; Leal, A.S.M.; Wilkinson, C.L.; Levin, A.R. The Harvard Automated Processing Pipeline for Electroencephalography (HAPPE): Standardized Processing Software for Developmental and High-Artifact Data. Front. Neurosci. 2018, 12, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, D. A Method to Standardize a Reference of Scalp EEG Recordings to a Point at Infinity. Physiol. Meas. 2001, 22, 693–711. [Google Scholar] [CrossRef]
- Tadel, F.; Baillet, S.; Mosher, J.C.; Pantazis, D.; Leahy, R.M. Brainstorm: A User-Friendly Application for MEG/EEG Analysis. Comput. Intell. Neurosci. 2011, 2011, 879716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, C.; Larson, E.; Richards, J.E.; Elsabbagh, M. Structural Templates for Imaging EEG Cortical Sources in Infants. Neuroimage 2021, 227, 117682. [Google Scholar] [CrossRef]
- Pivik, R.T.; Andres, A.; Tennal, K.B.; Gu, Y.; Downs, H.; Bellando, B.J.; Jarratt, K.; Cleves, M.A.; Badger, T.M. Resting Gamma Power during the Postnatal Critical Period for GABAergic System Development Is Modulated by Infant Diet and Sex. Int. J. Psychophysiol. 2019, 135, 73–94. [Google Scholar] [CrossRef]
- Best, J.R.; Miller, P.H. A Developmental Perspective on Executive Function. Child Dev. 2010, 81, 1641–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, D.; Chang, L.; Ernst, T.M.; Curran, M.; Buchthal, S.D.; Alicata, D.; Skranes, J.; Johansen, H.; Hernandez, A.; Yamakawa, R.; et al. Structural Growth Trajectories and Rates of Change in the First 3 Months of Infant Brain Development. JAMA Neurol. 2014, 71, 1266–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Infantes, D.; Cereijo, R.; Sebastiani, G.; Pérez-Cruz, M.; Villarroya, F.; Ibáñez, L. Nerve Growth Factor Levels in Term Human Infants: Relationship to Prenatal Growth and Early Postnatal Feeding. Int. J. Endocrinol. 2018, 2018, 7562702. [Google Scholar] [CrossRef] [PubMed]
- Blüml, S.; Wisnowski, J.L.; Nelson, M.D., Jr.; Paquette, L.; Gilles, F.H.; Kinney, H.C.; Panigrahy, A. Metabolic Maturation of the Human Brain from Birth through Adolescence: Insights from in Vivo Magnetic Resonance Spectroscopy. Cereb. Cortex 2013, 23, 2944–2955. [Google Scholar] [CrossRef] [Green Version]
- Lenroot, R.K.; Giedd, J.N. Brain Development in Children and Adolescents: Insights from Anatomical Magnetic Resonance Imaging. Neurosci. Biobehav. Rev. 2006, 30, 718–729. [Google Scholar] [CrossRef]
- Georgieff, M.K. Nutrition and the developing brain: Nutrient priorities and measurement. Am. J. Clin. Nutr. 2007, 85, 614S–620S. [Google Scholar] [CrossRef] [PubMed]
- Başar, E. A Review of Gamma Oscillations in Healthy Subjects and in Cognitive Impairment. Int. J. Psychophysiol. 2013, 90, 99–117. [Google Scholar] [CrossRef]
- Wang, X.-J. Neurophysiological and Computational Principles of Cortical Rhythms in Cognition. Physiol. Rev. 2010, 90, 1195–1268. [Google Scholar] [CrossRef]
- Uhlhaas, P.J.; Roux, F.; Singer, W.; Haenschel, C.; Sireteanu, R.; Rodriguez, E. The Development of Neural Synchrony Reflects Late Maturation and Restructuring of Functional Networks in Humans. Proc. Natl. Acad. Sci. USA 2009, 106, 9866–9871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golbs, A.; Nimmervoll, B.; Sun, J.-J.; Sava, I.E.; Luhmann, H.J. Control of Programmed Cell Death by Distinct Electrical Activity Patterns. Cereb. Cortex 2011, 21, 1192–1202. [Google Scholar] [CrossRef] [Green Version]
- Lessmann, V.; Gottmann, K.; Malcangio, M. Neurotrophin Secretion: Current Facts and Future Prospects. Prog. Neurobiol. 2003, 69, 341–374. [Google Scholar] [CrossRef]
- Hong, E.J.; McCord, A.E.; Greenberg, M.E. A Biological Function for the Neuronal Activity-Dependent Component of Bdnf Transcription in the Development of Cortical Inhibition. Neuron 2008, 60, 610–624. [Google Scholar] [CrossRef] [Green Version]
- Gottmann, K.; Mittmann, T.; Lessmann, V. BDNF Signaling in the Formation, Maturation and Plasticity of Glutamatergic and GABAergic Synapses. Exp. Brain Res. 2009, 199, 203–234. [Google Scholar] [CrossRef]
- Nassar, M.F.; Younis, N.T.; El-Arab, S.E.; Fawzi, F.A. Neuro-Developmental Outcome and Brain-Derived Neurotrophic Factor Level in Relation to Feeding Practice in Early Infancy. Matern. Child Nutr. 2011, 7, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Abdel Salam, S.; Menazie, E.; Elhady, M.; El Attar, S. The Effect of Brain-Derived Neurotrophic Factor on Motor Development and Its Relation to the Pattern of Feeding in Early Infancy. Sci. J. Al-Azhar Med. Fac. Girls 2021, 5, 223–228. [Google Scholar] [CrossRef]
- Porcher, C.; Medina, I.; Gaiarsa, J.-L. Mechanism of BDNF Modulation in GABAergic Synaptic Transmission in Healthy and Disease Brains. Front. Cell. Neurosci. 2018, 12, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredes, M.F.; James, D.; Gil-Perotin, S.; Kim, H.; Cotter, J.A.; Ng, C.; Sandoval, K.; Rowitch, D.H.; Xu, D.; Mcquillen, P.S.; et al. Extensive Migration of Young Neurons into the Infant Human Frontal Lobe. Science 2016, 354, aaf7073. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ari, Y. The GABA Excitatory/Inhibitory Developmental Sequence : A Personal Journey. Neuroscience 2014, 279, 187–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyzio, R.; Represa, A.; Jorquera, I.; Ben-ari, Y.; Gozlan, H.; Aniksztejn, L. The Establishment of GABAergic and Glutamatergic Synapses on CA1 Pyramidal Neurons Is Sequential and Correlates with the Development of the Apical Dendrite. J. Neurosci. 1999, 19, 10372–10382. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Broadbelt, K.G.; Haynes, R.L.; Rebecca, D.; Volpe, J.J.; Kinney, H.C. Late Development of the GABAergic System in the Human Cerebral Cortex and White Matter. J. Neuropathol. Exp. Neurol. 2012, 70, 841–858. [Google Scholar] [CrossRef] [Green Version]
- Galanopoulou, A.S. Sexually Dimorphic Expression of KCC2 and GABA Function. Epilepsy Res. 2008, 80, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Niemarkt, H.J.; Jennekens, W.; Pasman, J.W.; Katgert, T.; van Pul, C.; Gavilanes, A.W.D.; Kramer, B.W.; Zimmermann, L.U.C.J.; Oetomo, S.B.; Intensive, N.; et al. Maturational Changes in Automated EEG Spectral Power Analysis in Preterm Infants. Pediatr. Res. 2011, 70, 529–534. [Google Scholar] [CrossRef] [Green Version]
- Van Elk, M.; Van Schie, H.T.; Hunnius, S.; Vesper, C.; Bekkering, H. NeuroImage You’ll Never Crawl Alone : Neurophysiological Evidence for Experience-Dependent Motor Resonance in Infancy. Neuroimage 2008, 43, 808–814. [Google Scholar] [CrossRef]
- Perone, S.; Gartstein, M.A. Mapping Cortical Rhythms to Infant Behavioral Tendencies via Baseline EEG and Parent-Report. Dev. Psychobiol. 2019, 61, 815–823. [Google Scholar] [CrossRef]
- Williams, I.A.; Tarullo, A.R.; Grieve, P.G.; Wilpers, A.; Vignola, E.F.; Myers, M.M.; Fifer, W.P. Fetal Cerebrovascular Resistance and Neonatal EEG Predict 18-Month Neurodevelopmental Outcome in Infants with Congenital Heart Disease. Ultrasound Obstet. Gynecol. 2012, 40, 304–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, A.R.; Barry, R.J.; McCarthy, R.; Selikowitz, M. Age and Sex Effects in the EEG: Differences in Two Subtypes of Attention-Deficit/Hyperactivity Disorder. Clin. Neurophysiol. 2001, 112, 815–826. [Google Scholar] [CrossRef]
- Alzu’bi, A.; Lindsay, S.; Kerwin, J.; Looi, S.J.; Khalil, F.; Clowry, G.J. Distinct Cortical and Sub-Cortical Neurogenic Domains for GABAergic Interneuron Precursor Transcription Factors NKX2.1, OLIG2 and COUP-TFII in Early Fetal Human Telencephalon. Brain Struct. Funct. 2017, 222, 2309–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jabbi, M.; Kohn, P.D.; Nash, T.; Ianni, A.; Coutlee, C.; Holroyd, T.; Carver, F.W.; Chen, Q.; Cropp, B.; Kippenhan, J.S.; et al. Convergent BOLD and Beta-Band Activity in Superior Temporal Sulcus and Frontolimbic Circuitry Underpins Human Emotion Cognition. Cereb. Cortex 2015, 25, 1878–1888. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, J.; Cheung, R.T.H.; Chan, R.H.M.; Chen, C.-Y. Right Temporal Oscillations of Infants in Relation to Contingent Learning. In Proceedings of the 2020 42nd Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC), Montreal, QC, Canada, 20–24 July 2020; Volume 2020, pp. 3273–3276. [Google Scholar]
- Daniels, M.C.; Adair, L.S. Breast-Feeding Influences Cognitive Development in Filipino Children. J. Nutr. 2005, 135, 2589–2595. [Google Scholar] [CrossRef] [Green Version]
- Bartels, M.; Van Beijsterveldt, C.E.M.; Boomsma, D.I. Breastfeeding, Maternal Education and Cognitive Function: A Prospective Study in Twins. Behav. Genet. 2009, 39, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Jensen, C.L.; Voigt, R.G.; Prager, T.C.; Zou, Y.L.; Fraley, J.K.; Rozelle, J.C.; Turcich, M.R.; Llorente, A.M.; Anderson, R.E.; Heird, W.C. Effects of Maternal Docosahexaenoic Acid Intake on Visual Function and Neurodevelopment in Breastfed Term Infants. Am. J. Clin. Nutr. 2005, 82, 125–132. [Google Scholar] [CrossRef]
- Ferguson, M.; Molfese, P.J. Breast-Fed Infants Process Speech Differently from Bottle-Fed Infants: Evidence from Neuroelectrophysiology. Dev. Neuropsychol. 2007, 31, 337–347. [Google Scholar] [CrossRef]
- Wieland, L.S.; Santesso, N. A Summary of a Cochrane Review: Supplementation with Long Chain Polyunsaturated Fatty Acids (LCPUFA) to Breastfeeding Mothers for Improving Child Growth and Development. Eur. J. Integr. Med. 2016, 8, 113–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.-J.; Yeung, C.-Y. Recent Advance in Infant Nutrition: Human Milk Oligosaccharides. Pediatr. Neonatol. 2021, 62, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Hauser, J.; Pisa, E.; Arias Vásquez, A.; Tomasi, F.; Traversa, A.; Chiodi, V.; Martin, F.P.; Sprenger, N.; Lukjancenko, O.; Zollinger, A.; et al. Sialylated Human Milk Oligosaccharides Program Cognitive Development through a Non-Genomic Transmission Mode. Mol. Psychiatry 2021, 26, 2854–2871. [Google Scholar] [CrossRef] [PubMed]
- Berger, P.K.; Plows, J.F.; Jones, R.B.; Alderete, T.L.; Yonemitsu, C.; Poulsen, M.; Ryoo, J.H.; Peterson, B.S.; Bode, L.; Goran, M.I. Human Milk Oligosaccharide 2’-Fucosyllactose Links Feedings at 1 Month to Cognitive Development at 24 Months in Infants of Normal and Overweight Mothers. PLoS ONE 2020, 15, e0228323. [Google Scholar] [CrossRef] [Green Version]
- Borre, Y.E.; O’Keeffe, G.W.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota and Neurodevelopmental Windows: Implications for Brain Disorders. Trends Mol. Med. 2014, 20, 509–518. [Google Scholar] [CrossRef]
- van den Elsen, L.W.J.; Garssen, J.; Burcelin, R.; Verhasselt, V. Shaping the Gut Microbiota by Breastfeeding: The Gateway to Allergy Prevention? Front. Pediatr. 2019, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Notarbartolo, V.; Giuffrè, M.; Montante, C.; Corsello, G.; Carta, M. Composition of Human Breast Milk Microbiota and Its Role in Children’s Health. Pediatr. Gastroenterol. Hepatol. Nutr. 2022, 25, 194–210. [Google Scholar] [CrossRef]
- Anderson, A.J.; Perone, S. Developmental Change in the Resting State Electroencephalogram : Insights into Cognition and the Brain. Brain Cogn. 2018, 126, 40–52. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Total | Dietary Group | Maternal WASI |
|---|---|---|---|
| 2 m | 313 | BF: 108 | 108.31 (10.76) |
| MF: 100 | 105.54 (9.65) | ||
| SF: 105 | 102.16 (12.07) | ||
| 3 m | 348 | BF: 114 | 109.34 (10.74) |
| MF: 116 | 105.04 (8.68) | ||
| SF: 119 | 103.19 (11.49) | ||
| 4 m | 342 | BF: 111 | 109.92 (10.39) |
| MF: 110 | 105.82 (8.90) | ||
| SF: 121 | 103.66 (10.96) | ||
| 5 m | 417 | BF: 137 | 109.80 (10.05) |
| MF: 136 | 105.16 (9.31) | ||
| SF: 144 | 103.29 (11.18) | ||
| 6 m | 419 | BF: 135 | 109.78 (10.78) |
| MF: 143 | 105.25 (9.36) | ||
| SF: 141 | 104.07 (10.96) |
| Variables | Age | Dietary Group Means (SD) | Main Effect of Group | |||
|---|---|---|---|---|---|---|
| BF | MF | SF | F | p | ||
| Gestation (Weeks) | 2 m | 39.54 (1.02) | 39.19 (0.93) | 39.30 (1.03) | 4.709 | 0.1 |
| 3 m | 39.65 (1.05) | 39.11 (0.96) | 39.08 (1.01) | 11.626 | <0.001 * | |
| 4 m | 39.61 (1.05) | 39.17 (0.89) | 39.31 (1.05) | 7.446 | <0.001 * | |
| 5 m | 39.56 (1.01) | 39.14 (0.90) | 39.09 (1.06) | 8.992 | <0.001 * | |
| 6 m | 39.60 (1.05) | 39.13 (0.90) | 39.29 (1.01) | 9.807 | <0.001 ** | |
| Birth weight (kgs) | 2 m | 3.53 (.032) | 3.51 (0.38) | 3.42 (0.37) | 3.032 | 0.5 |
| 3 m | 3.55 (0.33) | 3.50 (0.39) | 3.45 (0.36) | 2.385 | 0.094 | |
| 4 m | 3.57 (0.33) | 3.50 (0.36) | 3.50 (0.36) | 2.914 | 0.056 | |
| 5 m | 3.55 (0.34) | 3.47 (0.37) | 3.43 (0.38) | 4.144 | 0.017 * | |
| 6 m | 3.55 (0.34) | 3.49 (0.37) | 3.45 (0.39) | 2.756 | 0.065 | |
| Birth length (cm) | 2 m | 51.27 (2.14) | 51.38 (2.67) | 51.11 (2.16) | 0.346 | 0.708 |
| 3 m | 51.42 (1.96) | 51.08 (2.54) | 51.36 (2.10) | 0.763 | 0.467 | |
| 4 m | 51.58 (2.29) | 51.31 (2.50) | 51.28 (2.11) | 0.572 | 0.565 | |
| 5 m | 51.54 (2.18) | 51.12 (2.30) | 51.22 (2.02) | 1.423 | 0.242 | |
| 6 m | 51.38 (2.21) | 51.24 (2.47) | 51.18 (2.14) | 0.288 | 0.75 | |
| Height (cm) | 2 m | 57.39 (1.77) | 57.15 (2.07) | 56.97 (1.72) | 1.351 | 0.261 |
| 3 m | 60.09 (2.01) | 60.01 (2.18) | 59.68(1.65) | 1.708 | 0.183 | |
| 4 m | 62.72 (2.01) | 62.37 (2.26) | 62.68 (2.04) | 0.959 | 0.384 | |
| 5 m | 64.33 (2.01) | 64.52 (2.17) | 64.65 (1.92) | 0.791 | 0.454 | |
| 6 m | 65.86 (2.32) | 66.44 (2.38) | 66.63 (2.05) | 4.289 | 0.014 * | |
| Weight (kg) | 2 m | 5.45 (0.57) | 5.32 (0.46) | 5.17 (0.49) | 7.912 | <0.001 ** |
| 3 m | 6.12 (0.71) | 6.13 (0.61) | 5.95 (0.52) | 3.338 | 0.037 * | |
| 4 m | 6.75 (0.77) | 6.77 (0.75) | 6.71 (0.64) | 0.201 | 0.818 | |
| 5 m | 7.18 (0.82) | 7.45 (0.86) | 7.33 (0.73) | 3.967 | 0.02 * | |
| 6 m | 7.63 (0.86) | 7.91 (0.83) | 7.89 (0.80) | 4.835 | 0.008 ** | |
| Head circ. (cm) | 2 m | 39.38 (1.06) | 39.38 (1.08) | 39.08 (1.00) | 2.73 | 0.067 |
| 3 m | 40.57 (1.11) | 40.71 (1.06) | 40.60 (1.09) | 0.541 | 0.582 | |
| 4 m | 41.78 (1.24) | 41.77 (1.10) | 41.90 (1.14) | 0.436 | 0.647 | |
| 5 m | 42.63 (1.19) | 42.82 (1.10) | 42.83 (1.26) | 1.187 | 0.206 | |
| 6 m | 43.38 (1.27) | 43.56 (1.17) | 43.78 (1.29) | 3.415 | 0.034 * | |
| 2 m | 108.31 (10.76) | 105.54 (9.65) | 103.16 (12.07) | 8.325 | <0.001 ** | |
| 3 m | 109.34 (10.74) | 105.04 (8.68) | 103.19 (11.49) | 10.569 | <0.001 ** | |
| Maternal WASI | 4 m | 109.92 (10.39) | 105.82 (8.90) | 103.66 (10.96) | 11.23 | <0.001 ** |
| 5 m | 109.80 (10.05) | 105.16 (9.32) | 103.29 (11.18) | 14.89 | <0.001 ** | |
| 6 m | 109.78 (10.78) | 105.25 (9.36) | 104.07 (10.99) | 11.5 | <0.001 ** | |
| M/F | x^2 | p | ||||
| Sex | 2 m | 50/58 | 49/51 | 56/49 | 1.071 | 0.585 |
| 3 m | 55/59 | 63/53 | 64/54 | 3.057 | 0.548 | |
| 4 m | 55/56 | 59/51 | 67/54 | 0.821 | 0.663 | |
| 5 m | 63/74 | 71/65 | 80/64 | 2.637 | 0.267 | |
| 6 m | 62/73 | 74/69 | 80/61 | 3.231 | 0.199 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilbreath, D.; Hagood, D.; Alatorre-Cruz, G.C.; Andres, A.; Downs, H.; Larson-Prior, L.J. Effects of Early Nutrition Factors on Baseline Neurodevelopment during the First 6 Months of Life: An EEG Study. Nutrients 2023, 15, 1535. https://doi.org/10.3390/nu15061535
Gilbreath D, Hagood D, Alatorre-Cruz GC, Andres A, Downs H, Larson-Prior LJ. Effects of Early Nutrition Factors on Baseline Neurodevelopment during the First 6 Months of Life: An EEG Study. Nutrients. 2023; 15(6):1535. https://doi.org/10.3390/nu15061535
Chicago/Turabian StyleGilbreath, Dylan, Darcy Hagood, Graciela Catalina Alatorre-Cruz, Aline Andres, Heather Downs, and Linda J. Larson-Prior. 2023. "Effects of Early Nutrition Factors on Baseline Neurodevelopment during the First 6 Months of Life: An EEG Study" Nutrients 15, no. 6: 1535. https://doi.org/10.3390/nu15061535