Polymorphisms in VDR, CYP27B1, CYP2R1, GC and CYP24A1 Genes as Biomarkers of Survival in Non-Small Cell Lung Cancer: A Systematic Review

 ,
,  ,
,  ,
,  , , ,
, , ,
Abstract
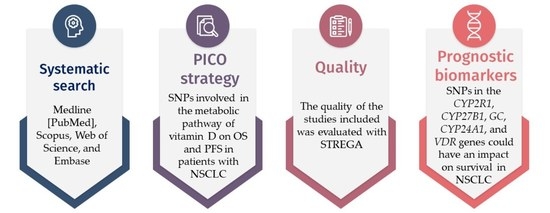
:
1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Eligibility Criteria
2.2.1. Inclusion Criteria
2.2.2. Exclusion Criteria
2.3. Data Collection and Analysis
2.3.1. Study Selection
2.3.2. Data Extraction
2.4. Quality Assessment
3. Results
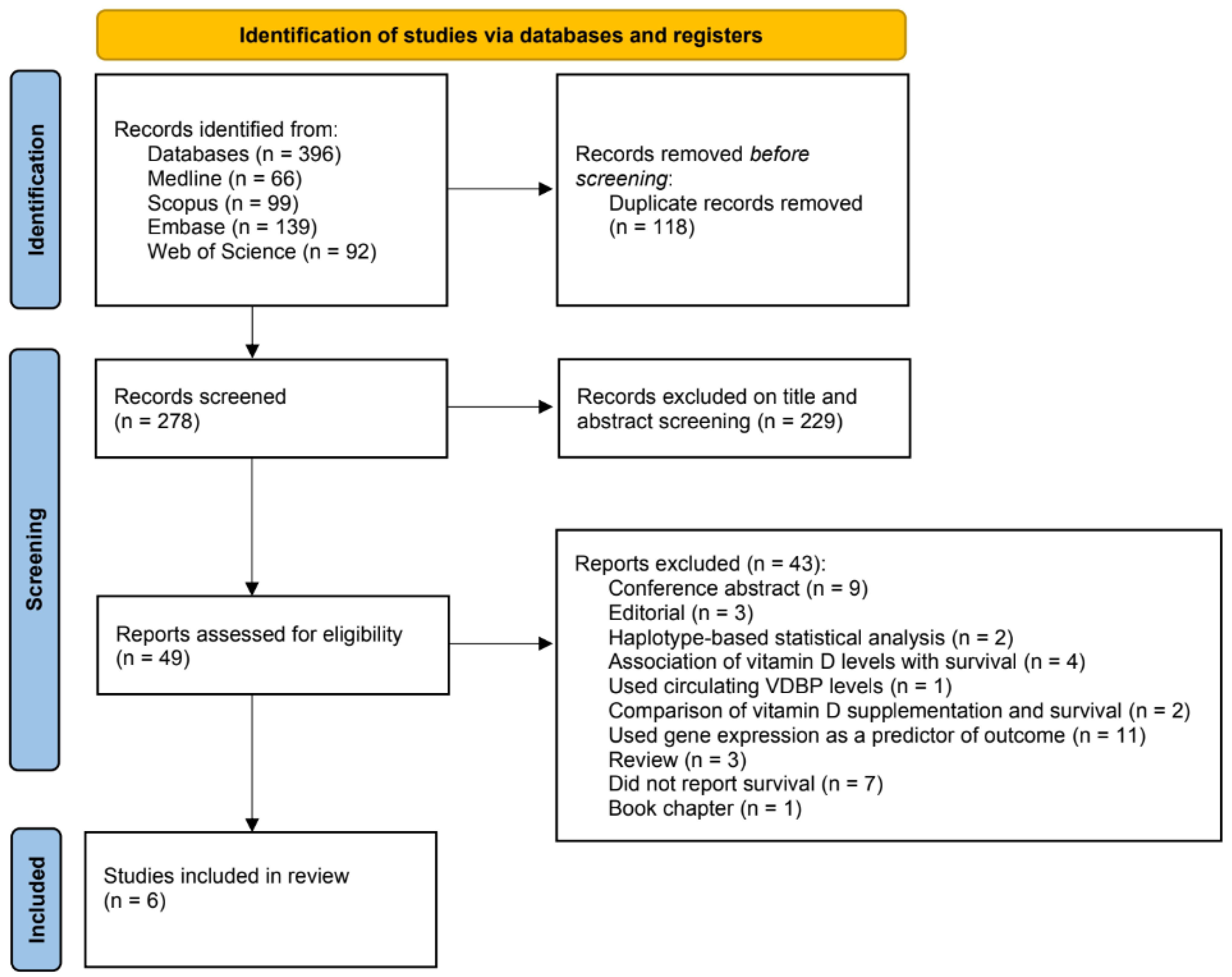
3.1. Search Results
3.2. Study Characteristics
3.3. Influence of Genetic Polymorphisms on Survival in NSCLC
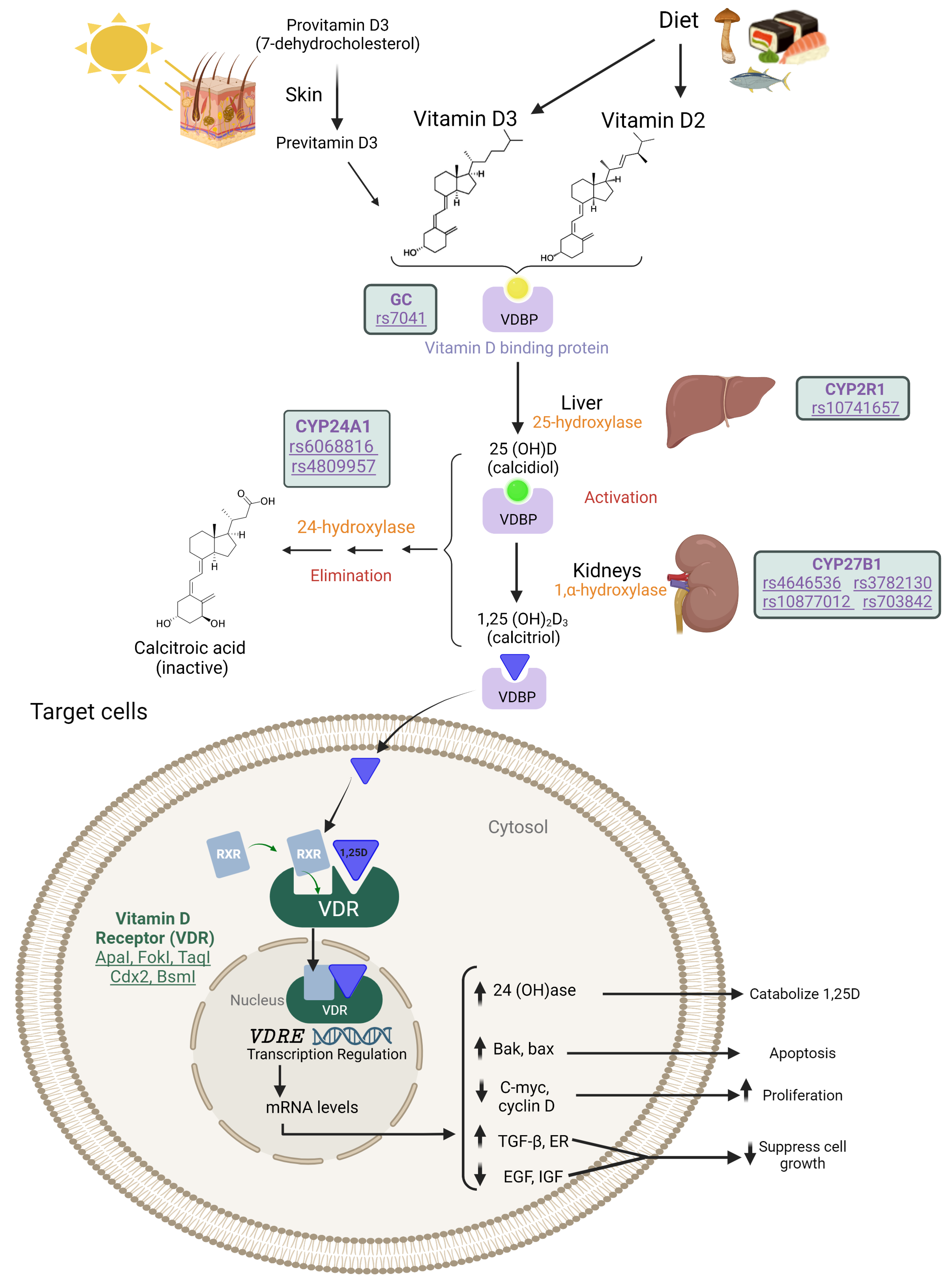
3.3.1. VDR: Vitamin D Receptor
3.3.2. CYP27B1: Cytochrome P450 Family 27 Subfamily B Member 1
3.3.3. CYP24A1: Cytochrome P450 Family 24 Subfamily A Member 1
3.3.4. GC: Vitamin D Binding Protein (Group-Specific Component)
3.3.5. CYP2R1: Cytochrome P450 Family 2 Subfamily R Member 1
3.4. Quality Assessment
4. Discussion
5. Guidelines for Future Research
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- OMS. Cancer. 2022. Available online: https://www.who.int/es/news-room/fact-sheets/detail/cancer (accessed on 15 November 2022).
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Patterson, A.D.; González, F.J.; Perdew, G.H.; Peters, J.M. Molecular Regulation of Carcinogenesis: Friend and Foe. Toxicol. Sci. 2018, 165, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Society of Clinical Oncology (ASCO). Lung Cancer-Non-Small Cell. 2023. Available online: https://www.cancer.net/cancer-t%C3%BDpes/lung-cancer-non-small-cell (accessed on 12 March 2023).
- American Society of Clinical Oncology (ASCO). Lung Cancer-Small Cell. 2023. Available online: https://www.cancer.net/cancer-types/lung-cancer-small-cell (accessed on 12 March 2023).
- Alexander, M.; Kim, S.Y.; Cheng, H. Update 2020: Management of Non-Small Cell Lung Cancer. Lung 2020, 198, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Araghi, M.; Fidler-Benaoudia, M.; Arnold, M.; Rutherford, M.; Bardot, A.; Ferlay, J.; Bucher, O.; De, P.; Engholm, G.; Gavin, A.; et al. International differences in lung cancer survival by sex, histological type and stage at diagnosis: An ICBP SURVMARK-2 Study. Thorax 2022, 77, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Önal, O.; Koçer, M.; Eroğlu, N.H.; Yilmaz, S.D.; Eroğlu, I.; Karadoğan, D. Survival analysis and factors affecting survival in patients who presented to the medical oncology unit with non-small cell lung cancer. Turk. J. Med Sci. 2020, 50, 1838–1850. [Google Scholar] [CrossRef]
- Pineda Lancheros, L.E.; Pérez Ramírez, C.; Sánchez Martín, A.; Gálvez Navas, J.M.; Martínez Martínez, F.; Ramírez Tortosa, M.D.C.; Jiménez Morales, A. Impact of Genetic Polymorphisms on the Metabolic Pathway of Vitamin D and Survival in Non-Small Cell Lung Cancer. Nutrients 2021, 13, 3783. [Google Scholar] [CrossRef]
- El-Sharkawy, A.; Malki, A. Vitamin D Signaling in Inflammation and Cancer: Molecular Mechanisms and Therapeutic Implica-tions. Molecules 2020, 25, 3219. [Google Scholar] [CrossRef]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef] [Green Version]
- Picotto, G.; Liaudat, A.C.; Bohl, L.; Tolosa de Talamoni, N. Molecular aspects of vitamin D anticancer activity. Cancer Investig. 2012, 30, 604–614. [Google Scholar] [CrossRef]
- Pineda Lancheros, L.E.; Rojo Tolosa, S.; Galvez Navas, J.M.; Martinez Martinez, F.; Sanchez Martin, A.; Jimenez Morales, A.; Perez Ramirez, C. Effect of Single Nucleotide Polymorphisms in the Vitamin D Metabolic Pathway on Susceptibility to Non-Small-Cell Lung Cancer. Nutrients 2022, 14, 4668. [Google Scholar] [CrossRef]
- Krasniqi, E.; Boshnjaku, A.; Wagner, K.H.; Wessner, B. Association between Polymorphisms in Vitamin D Pathway-Related Genes, Vitamin D Status, Muscle Mass and Function: A Systematic Review. Nutrients 2021, 13, 3109. [Google Scholar] [CrossRef]
- Karcıoğlu Batur, L.; Özaydın, A.; Maviş, M.E.; Gürsu, G.G.; Harbige, L.; Hekim, N. Vitamin-D Binding Protein Gene Polymorphisms and Serum 25-Hydroxyvitamin-D in a Turkish Population. Metabolites 2021, 11, 696. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez Rojo, P.; Perez Ramirez, C.; Galvez Navas, J.M.; Pineda Lancheros, L.E.; Rojo Tolosa, S.; Ramirez Tortosa, M.D.C.; Jimenez Morales, A. Vitamin D-Related Single Nucleotide Polymorphisms as Risk Biomarker of Cardiovascular Disease. Int. J. Mol. Sci. 2022, 23, 8686. [Google Scholar] [CrossRef] [PubMed]
- Márquez-Pete, N.; Pérez-Ramírez, C.; Maldonado Montoro, M.D.M.; Martínez-Martínez, F.; Fernández-Llimos, F.; Sánchez-Pozo, A.; Ramírez-Tortosa, M.C.; Jiménez-Morales, A. Association of vitamin D receptor genes polymorphisms with rheumatoid arthritis. Arch. Med. Sci. 2021. [Google Scholar] [CrossRef]
- Cancela Díez, B.; Pérez-Ramírez, C.; Maldonado-Montoro, M.D.M.; Carrasco-Campos, M.I.; Sánchez Martín, A.; Pineda Lancheros, L.E.; Martínez-Martínez, F.; Calleja-Hernández, M.; Ramírez-Tortosa, M.C.; Jiménez-Morales, A. Association between polymorphisms in the vitamin D receptor and susceptibility to multiple sclerosis. Pharm. Genom. 2021, 31, 40–47. [Google Scholar] [CrossRef]
- Sakaki, T.; Kagawa, N.; Yamamoto, K.; Inouye, K. Metabolism of vitamin D3 by cytochromes P450. Front. Biosci. 2005, 10, 119–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, S.M.; Shin, E.A. Exploring vitamin D metabolism and function in cancer. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Guan, H.; Liu, C.; Chen, Z.; Wang, L.; Li, C.; Zhao, J.; Yu, Y.; Zhang, P.; Chen, W.; Jiang, A. 1,25-Dihydroxyvitamin D3 up-regulates expression of hsa-let-7a-2 through the interaction of VDR/VDRE in human lung cancer A549 cells. Gene 2013, 522, 142–146. [Google Scholar] [CrossRef]
- Linowiecka, K.; Wolnicka-Glubisz, A.; Brozyna, A.A. Vitamin D endocrine system in breast cancer. Acta Biochim. Pol. 2021, 68, 489–497. [Google Scholar] [CrossRef]
- Miki, H.; Okito, A.; Akiyama, M.; Ono, T.; Tachikawa, N.; Nakahama, K.I. Genetic and epigenetic regulation of osteopontin by cyclic adenosine 3’5’-monophosphate in osteoblasts. Gene 2020, 763, 145059. [Google Scholar] [CrossRef] [PubMed]
- Heist, R.S.; Zhou, W.; Wang, Z.; Liu, G.; Neuberg, D.; Su, L.; Asomaning, K.; Hollis, B.W.; Lynch, T.J.; Wain, J.C.; et al. Circulating 25-hydroxyvitamin D, VDR polymorphisms, and survival in advanced non-small-cell lung cancer. J. Clin. Oncol. 2008, 26, 5596–5602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, J.; Chen, X.; Wang, J.; Li, J.; Xu, F.; Gao, S.; Yu, H.; Qian, B. Genetic Polymorphisms in the Vitamin D Pathway and Non-small Cell Lung Cancer Survival. Pathol. Oncol. Res. 2020, 26, 1709–1715. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, W.; Hu, Z.B.; Xu, L.; Shu, Y.Q.; Pan, S.Y.; Dai, J.C.; Jin, G.F.; Ma, H.X.; Shen, H.B. Plasma Vitamin D Levels and Vitamin D Receptor Polymorphisms Are Associated with Survival of Non-small Cell Lung Cancer. Chin. J. Cancer Res. 2011, 23, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.; Cheng, J.; Gao, J.; Wang, J.; Liu, X.; Wang, L. Vitamin D receptor genetic variants are associated with chemotherapy response and prognosis in patients with advanced non-small-cell lung cancer. Clin. Lung Cancer 2013, 14, 433–439. [Google Scholar] [CrossRef]
- Zhou, W.; Heist, R.S.; Liu, G.; Neuberg, D.S.; Asomaning, K.; Su, L.; Wain, J.C.; Lynch, T.J.; Giovannucci, E.; Christiani, D.C. Polymorphisms of vitamin D receptor and survival in early-stage non-small cell lung cancer patients. Cancer Epidemiol. Biomark. Prev. 2006, 15, 2239–2245. [Google Scholar] [CrossRef] [Green Version]
- Ge, N.; Chu, X.M.; Xuan, Y.P.; Ren, D.Q.; Wang, Y.; Ma, K.; Gao, H.J.; Jiao, W.J. Associations between abnormal vitamin D metabolism pathway function and non-small cell lung cancer. Oncol. Lett. 2017, 14, 7538–7544. [Google Scholar] [CrossRef] [PubMed]
- Gromowski, T.; Gapska, P.; Scott, R.J.; Kaklewski, K.; Marciniak, W.; Durda, K.; Lener, M.; Gorski, B.; Cybulski, C.; Sukiennicki, G.; et al. Serum 25(OH)D concentration, common variants of the VDR gene and lung cancer occurrence. Int. J. Cancer 2017, 141, 336–341. [Google Scholar] [CrossRef] [Green Version]
- Maj, E.; Trynda, J.; Maj, B.; Gebura, K.; Bogunia-Kubik, K.; Chodynski, M.; Kutner, A.; Wietrzyk, J. Differential response of lung cancer cell lines to vitamin D derivatives depending on EGFR, KRAS, p53 mutation status and VDR polymorphism. J. Steroid Biochem. Mol. Biol. 2019, 193, 105431. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cheng, J.; Yang, K. Vitamin D-Related Gene Polymorphisms, Plasma 25-Hydroxy-Vitamin D, Cigarette Smoke and Non-Small Cell Lung Cancer (NSCLC) Risk. Int. J. Mol. Sci. 2016, 17, 1597. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couzin, J. Human genome. HapMap launched with pledges of $100 million. Science 2002, 298, 941–942. [Google Scholar] [CrossRef] [PubMed]
- Phan, L.; Jin, Y.; Zhang, H.; Qiang, W.; Shekhtman, E.; Shao, D.; Revoe, D.; Villamarin, R.; Ivanchenko, E.; Kimura, M.; et al. “ALFA: Allele Frequency Aggregator”. National Center for Biotechnology Information, U.S. National Library of Medicine. Available online: www.ncbi.nlm.nih.gov/snp/docs/gsr/alfa/ (accessed on 10 March 2020).
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. PLoS Med. 2009, 6, e1000100. [Google Scholar] [CrossRef] [PubMed]
- Schardt, C.; Adams, M.B.; Owens, T.; Keitz, S.; Fontelo, P. Utilization of the PICO framework to improve searching PubMed for clinical questions. BMC Med. Inform. Decis. Mak. 2007, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Little, J.; Higgins, J.P.; Ioannidis, J.P.; Moher, D.; Gagnon, F.; von Elm, E.; Khoury, M.J.; Cohen, B.; Davey-Smith, G.; Grimshaw, J.; et al. STrengthening the REporting of Genetic Association Studies (STREGA)—An extension of the STROBE statement. Genet. Epidemiol. 2009, 33, 581–598. [Google Scholar] [CrossRef]
- Andric, M.; Jacimovic, J.; Jakovljevic, A.; Nikolic, N.; Milasin, J. Gene polymorphisms in odontogenic keratocysts and ameloblastomas: A systematic review. Oral Dis. 2022, 28, 1421–1430. [Google Scholar] [CrossRef]
- Chair, S.Y.; Chan, J.Y.W.; Law, B.M.H.; Waye, M.M.Y.; Chien, W.T. Genetic susceptibility in pneumoconiosis in China: A systematic review. Int. Arch. Occup. Environ. Health 2022, 96, 45–56. [Google Scholar] [CrossRef]
- Chair, S.Y.; Law, B.M.H.; Chan, J.Y.W.; So, W.K.W.; Waye, M.M.Y. Association of genetic polymorphisms with psychological symptoms in cancer: A systematic review. Asia Pac. J. Oncol. Nurs. 2022, 9, 12–20. [Google Scholar] [CrossRef]
- Vaughan-Shaw, P.G.; O’Sullivan, F.; Farrington, S.M.; Theodoratou, E.; Campbell, H.; Dunlop, M.G.; Zgaga, L. The impact of vitamin D pathway genetic variation and circulating 25-hydroxyvitamin D on cancer outcome: Systematic review and meta-analysis. Br. J. Cancer 2017, 116, 1092–1110. [Google Scholar] [CrossRef]
- Kong, J.; Xu, F.; Qu, J.; Wang, Y.; Gao, M.; Yu, H.; Qian, B. Genetic polymorphisms in the vitamin D pathway in relation to lung cancer risk and survival. Oncotarget 2015, 6, 2573–2582. [Google Scholar] [CrossRef] [Green Version]
- Duan, L.; Xue, Z.; Ji, H.; Zhang, D.; Wang, Y. Effects of CYP2R1 gene variants on vitamin D levels and status: A systematic review and meta-analysis. Gene 2018, 678, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Duan, G.Q.; Zheng, X.; Li, W.K.; Zhang, W.; Li, Z.; Tan, W. The Association Between VDR and GC Polymorphisms and Lung Cancer Risk: A systematic Review and Meta-Analysis. Genet. Test. Mol. Biomark. 2020, 24, 285–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiba, T.; Morikawa, T.; Odaka, M.; Nakada, T.; Kamiya, N.; Yamashita, M.; Yabe, M.; Inagaki, T.; Asano, H.; Mori, S.; et al. Vitamin D Supplementation and Survival of Patients with Non-small Cell Lung Cancer: A Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Cancer Res. 2018, 24, 4089–4097. [Google Scholar] [CrossRef] [Green Version]
- Trummer, O.; Schweighofer, N.; Haudum, C.W.; Trummer, C.; Pilz, S.; Theiler-Schwetz, V.; Keppel, M.H.; Grübler, M.; Pieber, T.R.; Renner, W.; et al. Genetic Components of 25-Hydroxyvitamin D Increase in Three Randomized Controlled Trials. J. Clin. Med. 2020, 9, 570. [Google Scholar] [CrossRef] [Green Version]
- Carlberg, C. Vitamin D in the Context of Evolution. Nutrients 2022, 14, 3018. [Google Scholar] [CrossRef] [PubMed]
- Rochel, N. Vitamin D and Its Receptor from a Structural Perspective. Nutrients 2022, 14, 2847. [Google Scholar] [CrossRef]
- Carlberg, C. Vitamin D and Its Target Genes. Nutrients 2022, 14, 1354. [Google Scholar] [CrossRef]
- Makishima, M. Update on recent progress in vitamin D research. Vitamin D receptor and the nuclear receptor superfamily. Clin. Calcium 2017, 27, 1533–1541. [Google Scholar]
- Afzal, S.; Brøndum-Jacobsen, P.; Bojesen, S.E.; Nordestgaard, B.G. Genetically low vitamin D concentrations and increased mortality: Mendelian randomisation analysis in three large cohorts. BMJ 2014, 349, g6330. [Google Scholar] [CrossRef] [Green Version]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a knowledge-based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef]
- Yokomura, K.; Suda, T.; Sasaki, S.; Inui, N.; Chida, K.; Nakamura, H. Increased expression of the 25-hydroxyvitamin D(3)-1alpha-hydroxylase gene in alveolar macrophages of patients with lung cancer. J. Clin. Endocrinol. Metab. 2003, 88, 5704–5709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramagopalan, S.V.; Heger, A.; Berlanga, A.J.; Maugeri, N.J.; Lincoln, M.R.; Burrell, A.; Handunnetthi, L.; Handel, A.E.; Disanto, G.; Orton, S.M.; et al. A ChIP-seq defined genome-wide map of vitamin D receptor binding: Associations with disease and evolution. Genome Res. 2010, 20, 1352–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holick, M.F. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef]
- Afzal, M.; Kazmi, I.; Al-Abbasi, F.A.; Alshehri, S.; Ghoneim, M.M.; Imam, S.S.; Nadeem, M.S.; Al-Zahrani, M.H.; Alzarea, S.I.; Alquraini, A. Current Overview on Therapeutic Potential of Vitamin D in Inflammatory Lung Diseases. Biomedicines 2021, 9, 1843. [Google Scholar] [CrossRef]
- Kim, S.H.; Chen, G.; King, A.N.; Jeon, C.K.; Christensen, P.J.; Zhao, L.; Simpson, R.U.; Thomas, D.G.; Giordano, T.J.; Brenner, D.E.; et al. Characterization of vitamin D receptor (VDR) in lung adenocarcinoma. Lung Cancer 2012, 77, 265–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, M.; Parwani, A.V.; Hershberger, P.A.; Lenzner, D.E.; Weissfeld, J.L. Nuclear vitamin D receptor expression is associated with improved survival in non-small cell lung cancer. J. Steroid Biochem. Mol. Biol. 2011, 123, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Latacz, M.; Snarska, J.; Kostyra, E.; Fiedorowicz, E.; Savelkoul, H.F.; Grzybowski, R.; Cieślińska, A. Single Nucleotide Polymorphisms in 25-Hydroxyvitamin D3 1-Alpha-Hydroxylase (CYP27B1) Gene: The Risk of Malignant Tumors and Other Chronic Diseases. Nutrients 2020, 12, 801. [Google Scholar] [CrossRef] [Green Version]
- Jones, G. 100 Years of Vitamin D: Historical aspects of vitamin D. Endocr. Connect. 2022, 11, e210594. [Google Scholar] [CrossRef]
- Gong, C.; Long, Z.; Yu, Y.; Zhu, L.; Tian, J.; Li, S.; Li, J.; Yu, H.; Chi, Q.; Piao, D.; et al. Dietary factors and polymorphisms in vitamin D metabolism genes: The risk and prognosis of colorectal cancer in northeast China. Sci. Rep. 2017, 7, 8827. [Google Scholar] [CrossRef] [Green Version]
- Parise, R.A.; Egorin, M.J.; Kanterewicz, B.; Taimi, M.; Petkovich, M.; Lew, A.M.; Chuang, S.S.; Nichols, M.; El-Hefnawy, T.; Hershberger, P.A. CYP24, the enzyme that catabolizes the antiproliferative agent vitamin D, is increased in lung cancer. Int. J. Cancer 2006, 119, 1819–1828. [Google Scholar] [CrossRef]
- Scazzone, C.; Agnello, L.; Ragonese, P.; Lo Sasso, B.; Bellia, C.; Bivona, G.; Schillaci, R.; Salemi, G.; Ciaccio, M. Association of CYP2R1 rs10766197 with MS risk and disease progression. J. Neurosci. Res. 2018, 96, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Ramnath, N.; Nadal, E.; Jeon, C.K.; Sandoval, J.; Colacino, J.; Rozek, L.S.; Christensen, P.J.; Esteller, M.; Beer, D.G.; Kim, S.H. Epigenetic regulation of vitamin D metabolism in human lung adenocarcinoma. J. Thorac. Oncol. 2014, 9, 473–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Kim, S.H.; King, A.N.; Zhao, L.; Simpson, R.U.; Christensen, P.J.; Wang, Z.; Thomas, D.G.; Giordano, T.J.; Lin, L.; et al. CYP24A1 is an independent prognostic marker of survival in patients with lung adenocarcinoma. Clin. Cancer Res. 2011, 17, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Zeng, R.; Li, H.; Jia, L.; Lee, S.H.; Jiang, R.; Zhang, Y.; Hu, X.; Ye, T.; Wang, X.; Yan, X.; et al. Association of CYP24A1 with survival and drug resistance in clinical cancer patients: A meta-analysis. BMC Cancer 2022, 22, 1317. [Google Scholar] [CrossRef]
- Cargill, M.; Altshuler, D.; Ireland, J.; Sklar, P.; Ardlie, K.; Patil, N.; Shaw, N.; Lane, C.R.; Lim, E.P.; Kalyanaraman, N.; et al. Characterization of single-nucleotide polymorphisms in coding regions of human genes. Nat. Genet. 1999, 22, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Rozmus, D.; Ciesielska, A.; Płomiński, J.; Grzybowski, R.; Fiedorowicz, E.; Kordulewska, N.; Savelkoul, H.; Kostyra, E.; Cieślińska, A. Vitamin D Binding Protein (VDBP) and Its Gene Polymorphisms-The Risk of Malignant Tumors and Other Diseases. Int. J. Mol. Sci. 2020, 21, 7822. [Google Scholar] [CrossRef]
- Chun, R.F. New perspectives on the vitamin D binding protein. Cell Biochem. Funct. 2012, 30, 445–456. [Google Scholar] [CrossRef]
- Speeckaert, M.; Huang, G.; Delanghe, J.R.; Taes, Y.E. Biological and clinical aspects of the vitamin D binding protein (Gc-globulin) and its polymorphism. Clin. Chim. Acta 2006, 372, 33–42. [Google Scholar] [CrossRef]
- Turner, A.M.; McGowan, L.; Millen, A.; Rajesh, P.; Webster, C.; Langman, G.; Rock, G.; Tachibana, I.; Tomlinson, M.G.; Berditchevski, F.; et al. Circulating DBP level and prognosis in operated lung cancer: An exploration of pathophysiology. Eur. Respir. J. 2013, 41, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Lafi, Z.M.; Irshaid, Y.M.; El-Khateeb, M.; Ajlouni, K.M.; Hyassat, D. Association of rs7041 and rs4588 Polymorphisms of the Vitamin D Binding Protein and the rs10741657 Polymorphism of CYP2R1 with Vitamin D Status Among Jordanian Patients. Genet. Test. Mol. Biomark. 2015, 19, 629–636. [Google Scholar] [CrossRef]
- Weinstein, S.J.; Mondul, A.M.; Layne, T.M.; Yu, K.; Huang, J.; Stolzenberg-Solomon, R.Z.; Ziegler, R.G.; Purdue, M.P.; Huang, W.Y.; Abnet, C.C.; et al. Prediagnostic Serum Vitamin D, Vitamin D Binding Protein Isoforms, and Cancer Survival. JNCI Cancer Spectr. 2022, 6, pkac019. [Google Scholar] [CrossRef]
- Dong, A.N.; Tan, B.H.; Pan, Y.; Ong, C.E. The CYP2R1 Enzyme: Structure, Function, Enzymatic Properties and Genetic Polymorphism. J. Pharm. Pharm. Sci. 2021, 24, 94–112. [Google Scholar] [CrossRef] [PubMed]
- Nissen, J.; Rasmussen, L.B.; Ravn-Haren, G.; Andersen, E.W.; Hansen, B.; Andersen, R.; Mejborn, H.; Madsen, K.H.; Vogel, U. Common variants in CYP2R1 and GC genes predict vitamin D concentrations in healthy Danish children and adults. PLoS ONE 2014, 9, e89907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slater, N.A.; Rager, M.L.; Havrda, D.E.; Harralson, A.F. Genetic Variation in CYP2R1 and GC Genes Associated with Vitamin D Deficiency Status. J. Pharm. Pract. 2017, 30, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lopez, E.; Brück, P.; Jansen, T.; Herwig, J.; Badenhoop, K. CYP2R1 (vitamin D 25-hydroxylase) gene is associated with susceptibility to type 1 diabetes and vitamin D levels in Germans. Diabetes Metab. Res. Rev. 2007, 23, 631–636. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Non-small-cell lung cancer OR lung cancer OR NSCLC |
| AND |
| vitamin D receptor OR VDR OR rs1544410 OR BsmI OR rs2228570 OR FokI OR rs7975232 OR ApaI OR rs11568820 OR Cdx-2 OR rs731236 OR TaqI OR CYP27B1 OR 1-a-hydroxylase OR rs10877012 OR rs4646536 OR rs3782130 OR rs703842 OR CYP2R1 OR 25-hydroxylase OR rs10741657 OR CYP24A1 OR 24-hydroxylase OR rs6068816 OR rs4809957 OR GC OR VDBP OR rs7041 OR vitamin D binding protein. |
| AND |
| Survival OR progression-free survival OR prognosis OR mortality OR death. |
| First Author (year) | Ethnicity | Study Design | Sample Size (Deaths/ Total) | NSCLC Stage | Median Follow-Up | VDR | CYP27B1 | CYP24A1 | GC | CYP2R1 | Outcome | PMID | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BmsI | Cdx-2 | FokI | ApaI | TaqI | rs10877012 | rs4646536 | rs3782130 | rs703842 | rs6068816 | rs4809957 | rs7041 | rs10741657 | OS | PFS | |||||||
| Zhou et al. (2006) [29] | Caucasians (USA) | Cohort | 186/373 | IA-IIB | 71 months | X | X | X | X | X | 17119052 | ||||||||||
| Heist et al. (2008) [25] | Caucasians (USA) | Cohort | 233/294 | III-IV | 42 months | X | X | X | X | 18936471 | |||||||||||
| Liu et al. (2011) [27] | Asiatic (China) | Cohort | 311/568 | I-IV | 19 months | X | X | X | X | X | 23467735 | ||||||||||
| Xiong et al. (2013) [28] | Asiatic (China) | Cohort | NA/755 | III-IV | NA | X | X | X | X | X | X | 23522953 | |||||||||
| Kong et al. (2020) [26] | Asiatic (China) | Cohort | 278/542 | I-IV | 80 months a | X | X | X | X | X | X | X | X | X | 31625015 | ||||||
| Pineda et al. (2021) [10] | Caucasians (Spain) | Cohort | 154/194 | I-IV | 204 months a | X | X | X | X | X | X | X | X | X | X | X | X | X | X | 34836039 | |
| dbSNP ID | SNP Position | Frequency (ALFA) | Sample Size | Overall Survival (OS) | Progression-Free Survival (PFS) | PMID | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| log Rank p | Ref. | HR (95% CI) | Comments | log Rank p | Ref. | HR (95% CI) | Comments | |||||
| Gene VDR (12q13.11) | ||||||||||||
| rs1544410 (BsmI) | Intron 8, C > T | T = 0.388066 (85636/220674) | 373 early-stage NSCLC | 0.31 | CC | 0.83 (0.59–1.16) CT 1.35 (0.90–2.03) TT | Multivariate Cox regression | >0.05 | 17119052 [29] | |||
| 180 early-stage Adenocarcinoma | 0.30 | CC | 0.88 (0.52–1.48) CT 1.52 (0.81–2.83) TT | Multivariate Cox regression | >0.05 | 17119052 [29] | ||||||
| 108 early-stage Squamous | 0.83 | CC | 0.59 (0.33–1.05) CT 1.18 (0.62–2.23) TT | Multivariate Cox regression | >0.05 | 17119052 [29] | ||||||
| 294 Advanced NSCLC | 0.61 | CC | 0.89 (0.66–1.19) CT 0.93 (0.64–1.35) TT | Multivariate Cox regression | 18936471 [25] | |||||||
| 562 NSCLC | 0.008 0.004 | CC CC | 1.55 (1.09–2.21) CT 4.33 (1.34–14.0) TT 1.64 (1.16–2.31) T | Multivariate Cox regression | 23467735 [27] | |||||||
| 755 Advanced NSCLC | >0.05 | >0.05 | 23522953 [28] | |||||||||
| 194 NSCLC | 0.500 | 0.900 | 34836039 [10] | |||||||||
| 48 resected NSCLC | 0.600 | 0.700 | 34836039 [10] | |||||||||
| 146 non-resected NSCLC | 0.0073 | C | 2.08 (1.22–3.56) TT | Univariate Cox Model | 0.500 | 34836039 [10] | ||||||
| rs11568820 (Cdx-2) | Intron 1, G > A | A = 0.28140 (14569/51774) | 373 early-stage NSCLC | 0.37 | GG | 0.84 (0.62–1.14) AG 0.92 (0.50–1.68) AA | Multivariate Cox regression | >0.05 | 17119052 [29] | |||
| 180 early-stage Adenocarcinoma | 0.33 | GG | 1.02 (0.64–1.62) AG 1.71 (0.81–3.60) AA | Multivariate Cox regression | >0.05 | 17119052 [29] | ||||||
| 108 early-stage Squamous | 0.05 0.04 | GG GG | 0.55 (0.32–0.95) AG 0.69 (0.16–2.96) AA 0.56 (0.33–0.95) A | Multivariate Cox regression | 0.03 | GG | 0.57 (0.34–0.94) A | Multivariate Cox regression | 17119052 [29] | |||
| 294 Advanced NSCLC | 0.59 | GG | 0.85 (0.63–1.15) GA 1.02 (0.64–1.63) AA | Multivariate Cox regression | 18936471 [25] | |||||||
| 586 NSCLC | 0.773 0.957 0.470 | Additive Dominant Recessive | Log-rank P | 23467735 [27] | ||||||||
| 194 NSCLC | 0.400 | 0.600 | 34836039 [10] | |||||||||
| 48 resected NSCLC | 0.0129 | G | 7.43 (1.53–36.15) AA | Multivariate Cox regression | 0.055 | G | 4.34 (0.97–19.5) AA | Univariate Cox Model | 34836039 [10] | |||
| 146 non-resected NSCLC | 0.700 | 0.400 | 34836039 [10] | |||||||||
| rs2228570 (FokI) | Exon 2, C > T | T = 0.388743 (91849/236272) | 373 early-stage NSCLC | 0.93 | CC | 0.84 (0.61–1.16) CT 1.13 (0.74–1.74) TT | Multivariate Cox regression | >0.05 | 17119052 [29] | |||
| 180 early-stage Adenocarcinoma | 0.40 | CC | 1.13 (0.67–1.88) CT 1.31 (0.70–2.46) TT | Multivariate Cox regression | >0.05 | 17119052 [29] | ||||||
| 108 early-stage Squamous | 0.64 | CC | 0.75 (0.44–1.28) CT 0.98 (0.47–2.03) TT | Multivariate Cox regression | >0.05 | 17119052 [29] | ||||||
| 294 Advanced NSCLC | 0.04 | CC | 1.32 (0.98–1.77) CT 1.41 (0.96–2.07) TT | Multivariate Cox regression | 18936471 [25] | |||||||
| 755 Advanced NSCLC | >0.05 | >0.05 | 23522953 [28] | |||||||||
| 194 NSCLC | 0.600 | 0.600 | 34836039 [10] | |||||||||
| 48 resected NSCLC | 1.000 | 0.400 | 34836039 [10] | |||||||||
| 146 non-resected NSCLC | 0.700 | 0.400 | 34836039 [10] | |||||||||
| rs7975232 (ApaI) | Intron 8, C > A | C = 0.44552 (17435/39134) | 586 NSCLC | NR | Removed (ApaI was not in HWE) | 23467735 [27] | ||||||
| 755 Advanced NSCLC | <0.001 | CC | 2.84 (2.63–3.94) AA | Multivariate Cox regression | 0.053 | CC | 1.43 (0.99–2.78) AA | Multivariate Cox regression | 23522953 [28] | |||
| 194 NSCLC | 0.400 | 0.600 | 34836039 [10] | |||||||||
| 48 resected NSCLC | 1.000 | 1.000 | 34836039 [10] | |||||||||
| 146 non-resected NSCLC | 0.0068 | C | 1.73 (1.16–2.58) AA | Univariate Cox Model | 0.0002 | C | 3.08 (1.71–5.54) AA | Multivariate Cox regression | 34836039 [10] | |||
| rs731236 (TaqI) | Exon 9, A > G | G = 0.387180 (74890/193424) | 586 NSCLC | 0.027 0.016 | AA AA | 1.41 (1.00–1.99) AG 4.26 (1.32–13.8) GG 1.49 (1.07–2.08) G | Multivariate Cox regression | 23467735 [27] | ||||
| 755 Advanced NSCLC | >0.05 | >0.05 | 23522953 [28] | |||||||||
| 194 NSCLC | 0.200 | 0.900 | 34836039 [10] | |||||||||
| 48 resected NSCLC | 0.700 | 0.500 | 34836039 [10] | |||||||||
| 146 non-resected NSCLC | 0.0005 | A | 2.71 (1.55–4.75) GG | Multivariate Cox regression | 0.0463 | A | 1.74 (1.01–2.99) GG | Multivariate Cox regression | 34836039 [10] | |||
| CYP27B1 (12q14.1) | ||||||||||||
| rs10877012 | 5′UTR, G > T | T = 0.292364 (46918/160478) | 542 NSCLC | 0.695 | TT | 1.28 (0.69–1.97) TG 1.39 (0.38–1.85) GG | Multivariate Cox regression | 31625015 [26] | ||||
| 194 NSCLC | 0.098 | TT | 1.826 (0.89–3.73) G | Univariate Cox Model | 0.400 | 34836039 [10] | ||||||
| 48 resected NSCLC | 0.0827 | T | 2.42 (0.89–6.58) GG | Univariate Cox Model | 0.400 | 34836039 [10] | ||||||
| 146 non-resected NSCLC | 0.200 | 0.044 | T | 2.05 (1.02–4.14) GG | Univariate Cox Model | 34836039 [10] | ||||||
| rs4646536 | Intron 6, A > G | G = 0.32704 (27483/84036) | 542 NSCLC | 0.625 | GG | 1.42 (0.73–2.74) GA 1.43 (0.68–3.04) AA | Multivariate Cox regression | 31625015 [26] | ||||
| 194 NSCLC | 0.056 | GG | 2.01 (0.98–4.14) A | Multivariate Cox regression | 0.023 | GG | 2.11 (1.11–4.04) A | Multivariate Cox regression | 34836039 [10] | |||
| 48 resected NSCLC | 0.0676 | G | 2.54 (0.93–6.89) AA | Univariate Cox Model | 0.300 | 34836039 [10] | ||||||
| 146 non-resected NSCLC | 0.200 | 0.004 | G | 8.77 (1.94–39.7) AA | Multivariate Cox regression | 34836039 [10] | ||||||
| rs3782130 | Promotor 5′, G > C | C = 0.18262 (2560/14018) | 542 NSCLC | 0.263 | CC | 0.63 (0.22–1.77) CG 1.16 (0.33–4.18) GG | Multivariate Cox regression | 31625015 [26] | ||||
| 194 NSCLC | 0.200 | 0.400 | 34836039 [10] | |||||||||
| 48 resected NSCLC | 0.0827 | C | 2.42 (0.89–6.59) GG | Univariate Cox Model | 0.400 | 34836039 [10] | ||||||
| 146 non-resected NSCLC | 0.200 | 0.045 | C | 2.05 (1.01–4.13) GG | Univariate Cox Model | 34836039 [10] | ||||||
| rs703842 | 3′UTR, T > C | C = 0.326395 (80258/245892) | 542 NSCLC | 0.627 | CC | 1.27 (0.67–3.25) CT 1.16 (0.45–2.78) TT | Multivariate Cox regression | 31625015 [26] | ||||
| CYP24A1 (20q13.2) | ||||||||||||
| rs6068816 | Exon 6, C > T | T = 0.108153 (33297/307870) | 542 NSCLC | 0.072 | CC | 1.13 (0.86–1.49) CT 0.76 (0.49–1.19) TT | Multivariate Cox regression | 31625015 [26] | ||||
| 194 NSCLC | 0.900 | 1.000 | 34836039 [10] | |||||||||
| 48 resected NSCLC | 0.117 | T | 4.99 (0.67–37.2) CC | Univariate Cox Model | 0.0359 | T | 8.49 (1.15–62.67) CC | Univariate Cox Model a | 34836039 [10] | |||
| 146 non-resected NSCLC | 0.0089 | C | 3.47 (1.37–8.79) TT | Multivariate Cox regression | 0.0048 | C | 8.77 (1.94–39.7) TT | Multivariate Cox regression | 34836039 [10] | |||
| rs4809957 | 3′UTR, A > G | G = 0.232972 (62226/267096) | 542 NSCLC | 0.790 | GG | 0.97 (0.74–1.26) GA 0.92 (0.58–1.45) AA | Multivariate Cox regression | 31625015 [26] | ||||
| 194 NSCLC | 0.300 | 0.089 | G | 2.03 (0.89–4.59) | Univariate Cox Model | 34836039 [10] | ||||||
| 48 resected NSCLC | 0.700 | 0.700 | 34836039 [10] | |||||||||
| 146 non-resected NSCLC | 0.700 | 0.900 | 34836039 [10] | |||||||||
| GC (4q13.3) | ||||||||||||
| rs7041 | Exon 11, T > G | T = 0.457674 (154076 /336650) | 542 NSCLC | 0.693 | TT | 0.82 (0.64–1.07) TG 1.13 (0.67–1.92) GG | Multivariate Cox regression | 31625015 [26] | ||||
| 194 NSCLC | 0.300 | 0.300 | 34836039 [10] | |||||||||
| 48 resected NSCLC | 0.0242 | T | 2.72 (1.14–6.47) GG | Univariate Cox Model | 0.044 | T | 2.26 (1.02–5.02) GG | Multivariate Cox regression | 34836039 [10] | |||
| 146 non-resected NSCLC | 0.700 | 0.400 | 34836039 [10] | |||||||||
| CYP2R1 (11p15.2) | ||||||||||||
| rs10741657 | 5′UTR, A > G | A = 0.379194 (73776/194560) | 542 NSCLC | 0.033 | GG | 0.79 (0.61–1.03) GA 0.69 (0.46–0.97) AA | Multivariate Cox regression | 31625015 [26] | ||||
| 270 NSCLC (Age group >60) | 0.014 | GG | 0.71 (0.51–0.99) A | Multivariate Cox regression | 31625015 [26] | |||||||
| 246 NSCLC (Chemotherapy: No) | 0.002 | GG | 0.65 (0.45–0.95) A | Multivariate Cox regression | 31625015 [26] | |||||||
| 194 NSCLC | 0.0525 | G | 1.58 (0.99–2.52) AA | Univariate Cox Model | 0.300 | 34836039 [10] | ||||||
| 48 resected NSCLC | 0.800 | 0.300 | 34836039 [10] | |||||||||
| 146 non-resected NSCLC | 0.700 | 1.000 | 34836039 [10] | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pineda-Lancheros, L.E.; Gálvez-Navas, J.M.; Rojo-Tolosa, S.; Membrive-Jiménez, C.; Valverde-Merino, M.I.; Martínez-Martínez, F.; Sánchez-Martín, A.; Ramírez-Tortosa, M.; Pérez-Ramírez, C.; Jiménez-Morales, A. Polymorphisms in VDR, CYP27B1, CYP2R1, GC and CYP24A1 Genes as Biomarkers of Survival in Non-Small Cell Lung Cancer: A Systematic Review. Nutrients 2023, 15, 1525. https://doi.org/10.3390/nu15061525
Pineda-Lancheros LE, Gálvez-Navas JM, Rojo-Tolosa S, Membrive-Jiménez C, Valverde-Merino MI, Martínez-Martínez F, Sánchez-Martín A, Ramírez-Tortosa M, Pérez-Ramírez C, Jiménez-Morales A. Polymorphisms in VDR, CYP27B1, CYP2R1, GC and CYP24A1 Genes as Biomarkers of Survival in Non-Small Cell Lung Cancer: A Systematic Review. Nutrients. 2023; 15(6):1525. https://doi.org/10.3390/nu15061525
Chicago/Turabian StylePineda-Lancheros, Laura Elena, José María Gálvez-Navas, Susana Rojo-Tolosa, Cristina Membrive-Jiménez, María Isabel Valverde-Merino, Fernando Martínez-Martínez, Almudena Sánchez-Martín, MCarmen Ramírez-Tortosa, Cristina Pérez-Ramírez, and Alberto Jiménez-Morales. 2023. "Polymorphisms in VDR, CYP27B1, CYP2R1, GC and CYP24A1 Genes as Biomarkers of Survival in Non-Small Cell Lung Cancer: A Systematic Review" Nutrients 15, no. 6: 1525. https://doi.org/10.3390/nu15061525