

Naringin Alleviates Glucose-Induced Aging by Reducing Fat Accumulation and Promoting Autophagy in Caenorhabditis elegans
 ,
,
Abstract
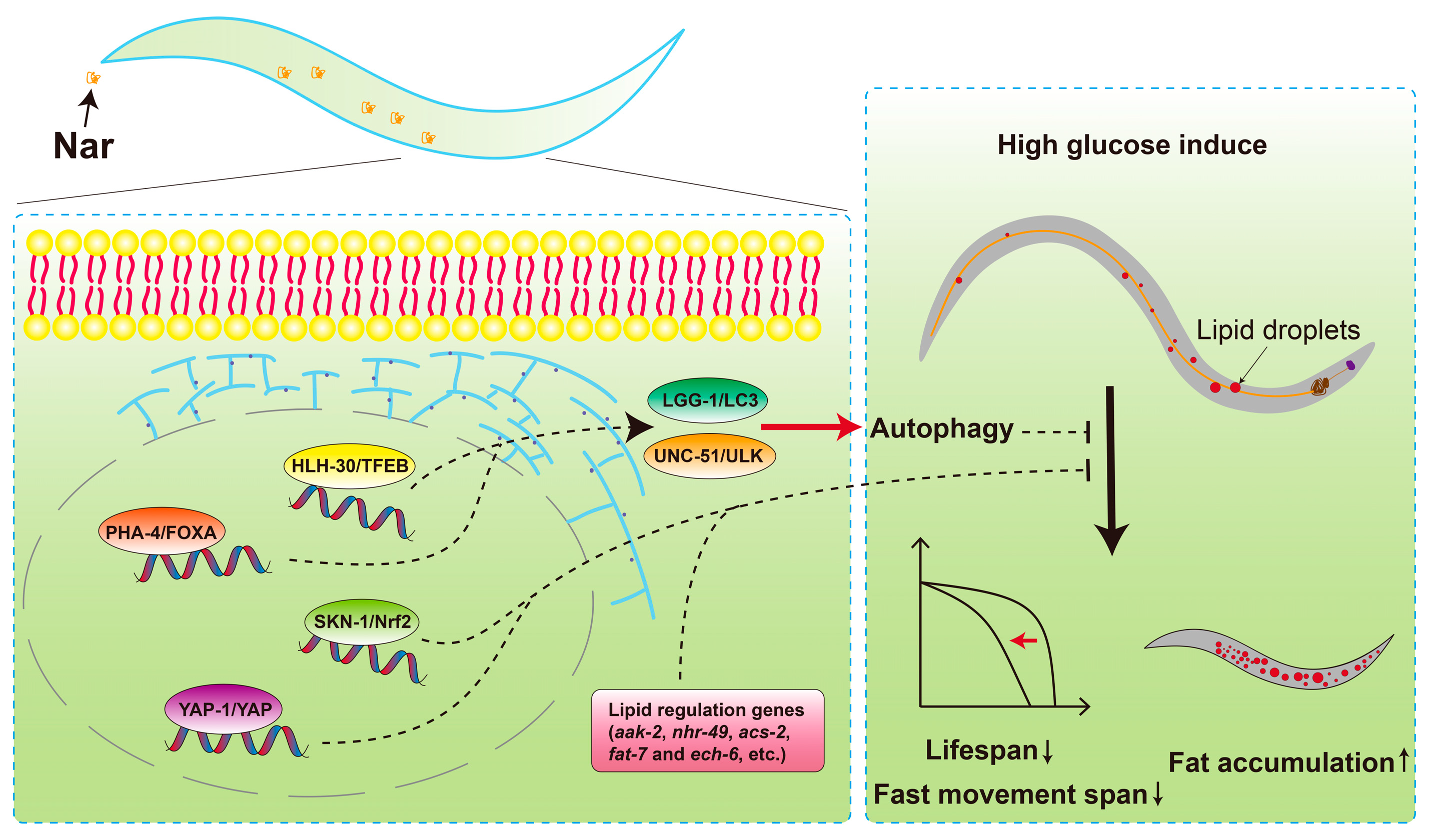
:1. Introduction
2. Materials and Methods
2.1. C. elegans Maintenance
2.2. Lifespan and Fast Movement Span Analysis
2.3. RNA Interference (RNAi)
2.4. Analysis of Auto-Fluorescence of C. elegans
2.5. Fat Accumulation Testing with Oil Red O Staining
2.6. Physiological Phenotype Analysis of C. elegans
2.7. Rhodamine 6G Staining
2.8. Observing Autophagy by dFP::LGG
2.9. Whole Transcriptome Sequencing Analysis
2.10. qPCR Analysis
2.11. Statistical Analysis
3. Results
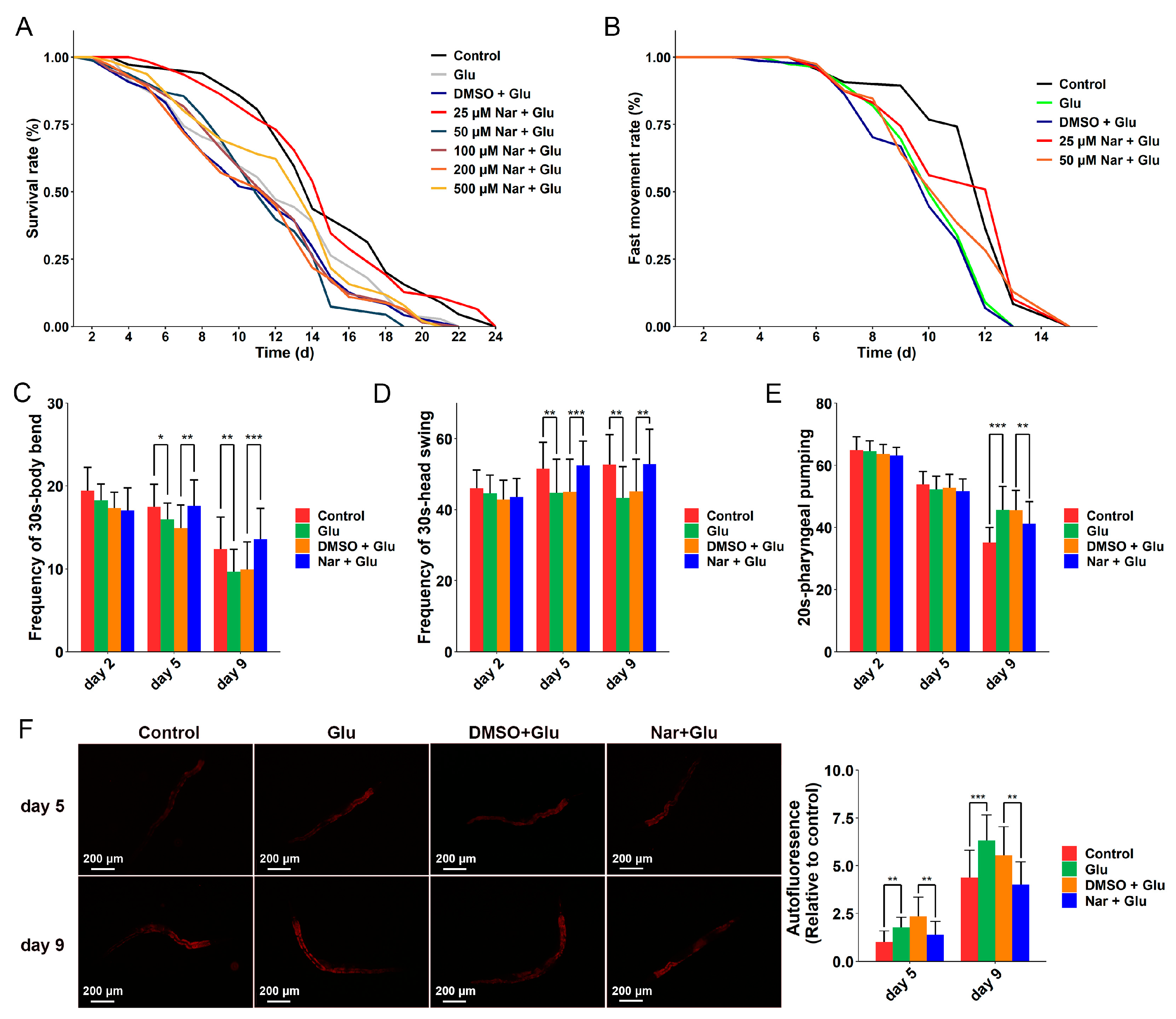
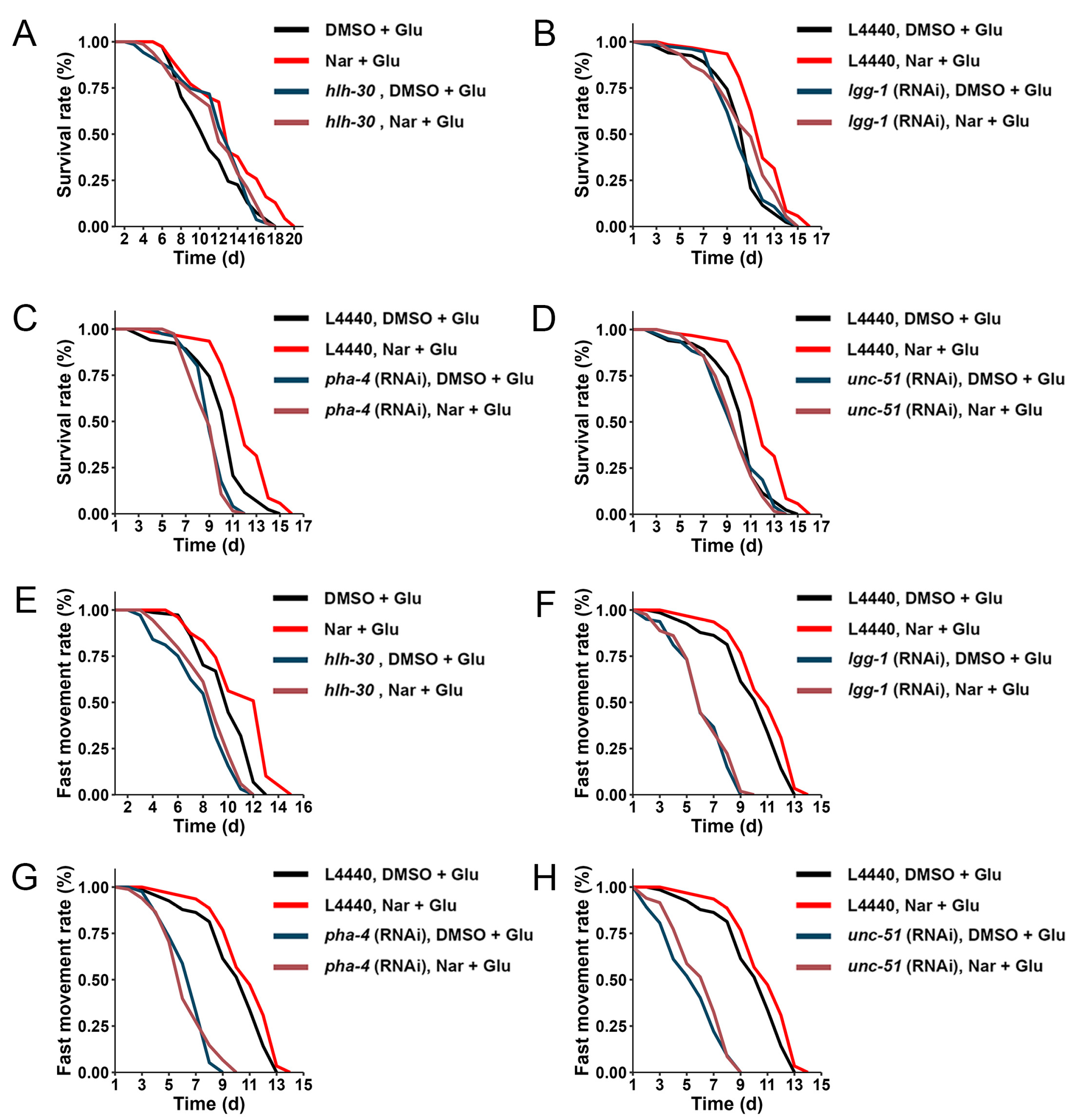
3.1. Anti-Aging Effect of Nar on HGI Worms
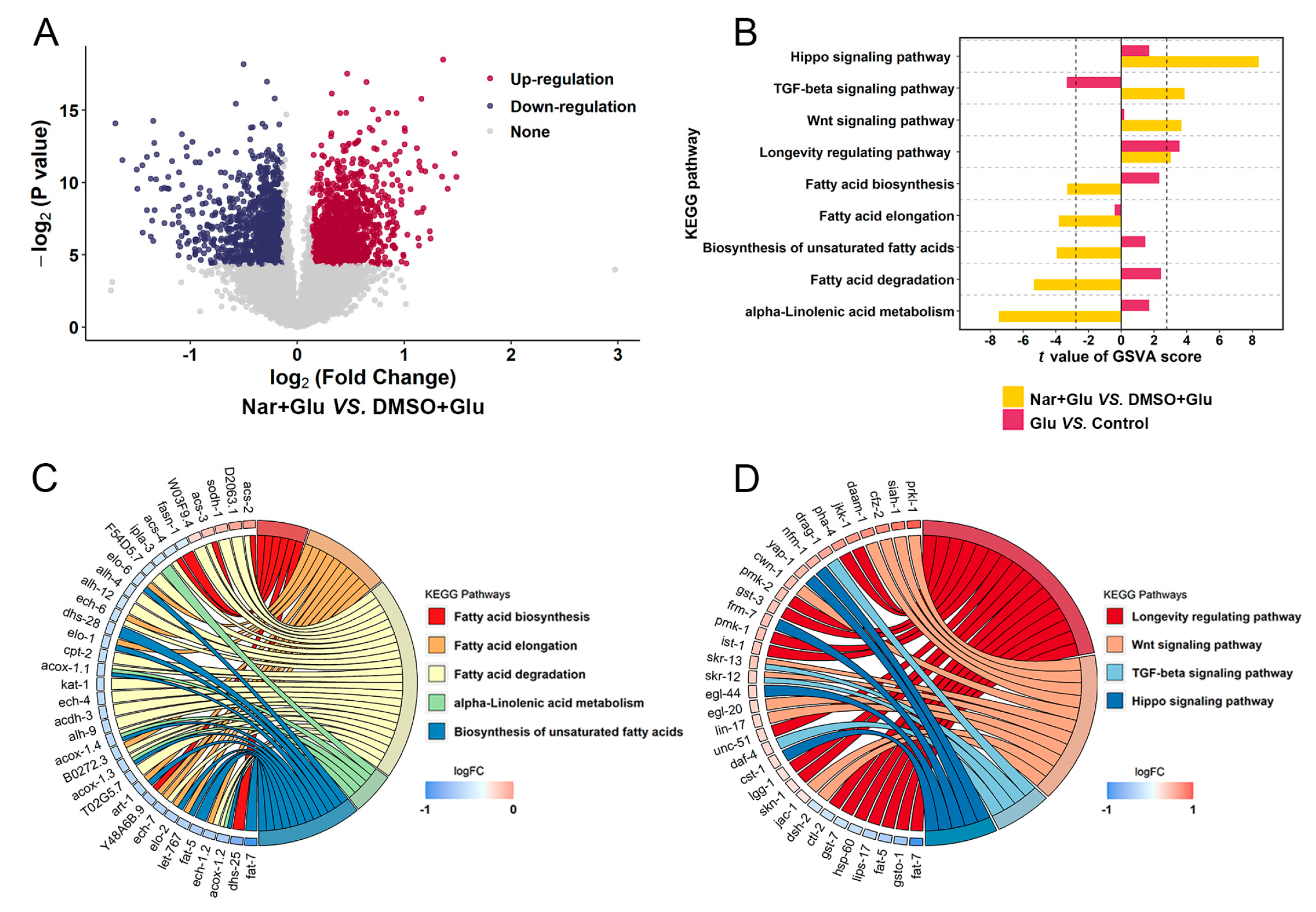
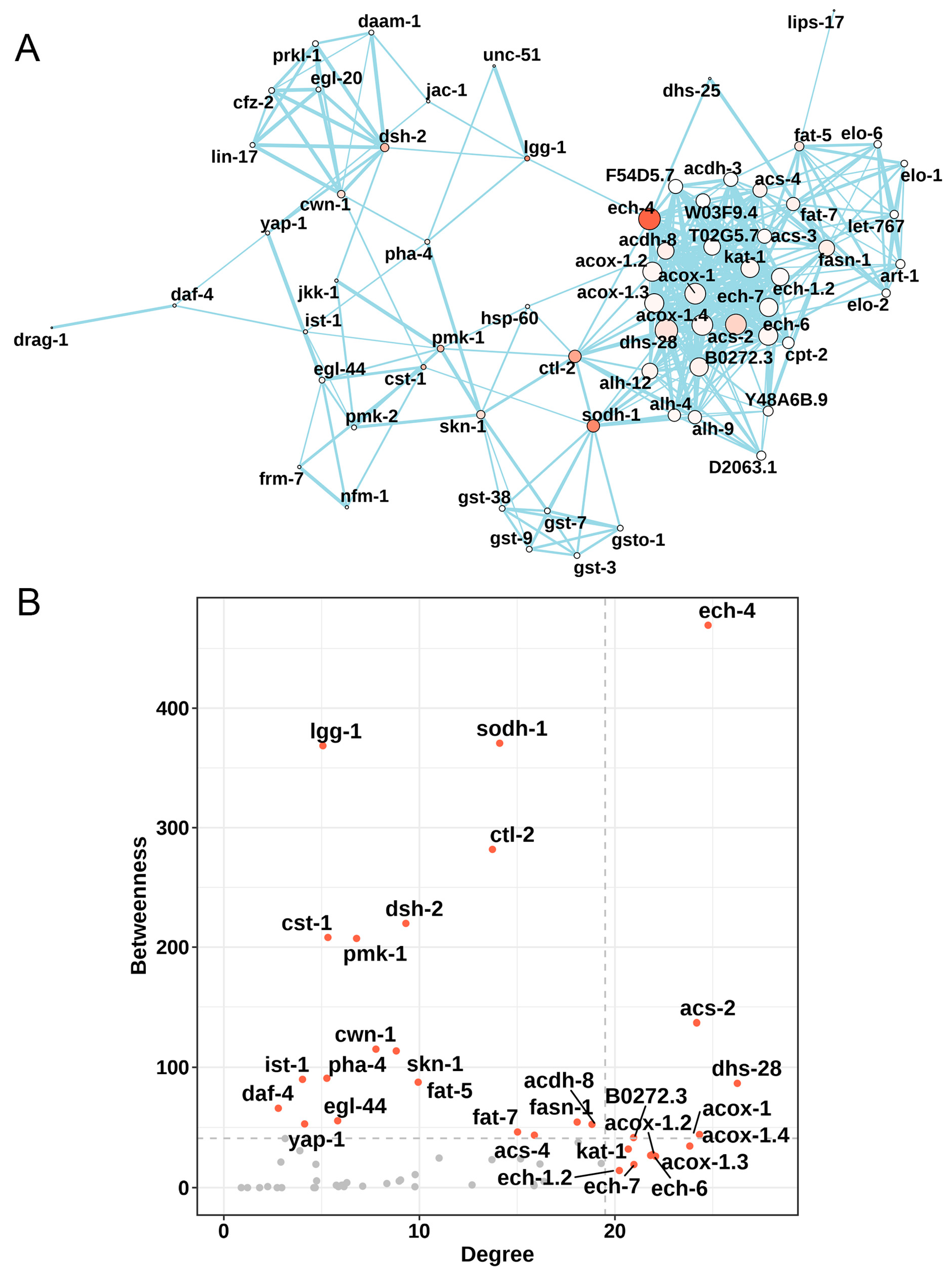
3.2. Analysis of Transcript Sequencing
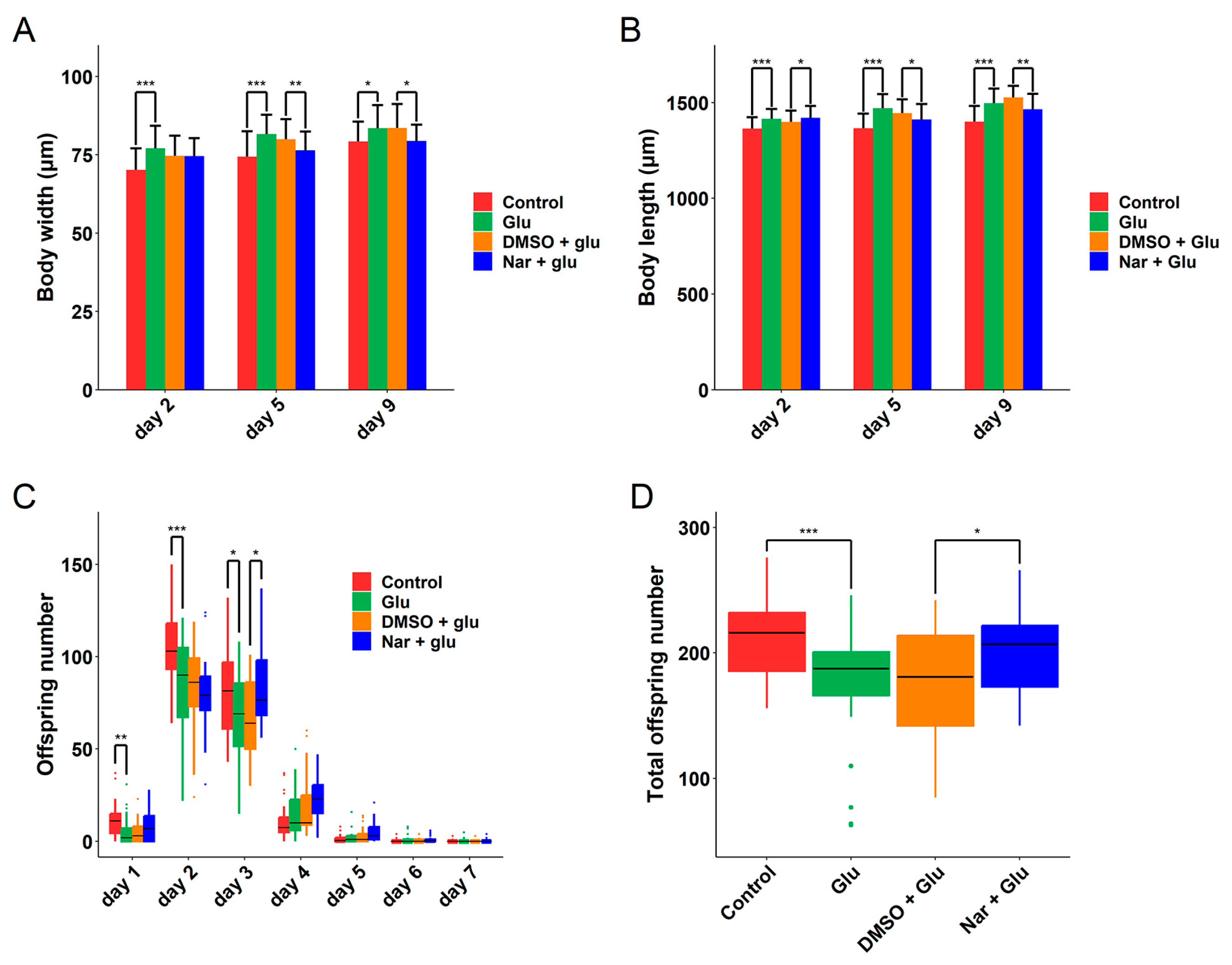
3.3. Improvement of Reproduction and Development in HGI C. elegans
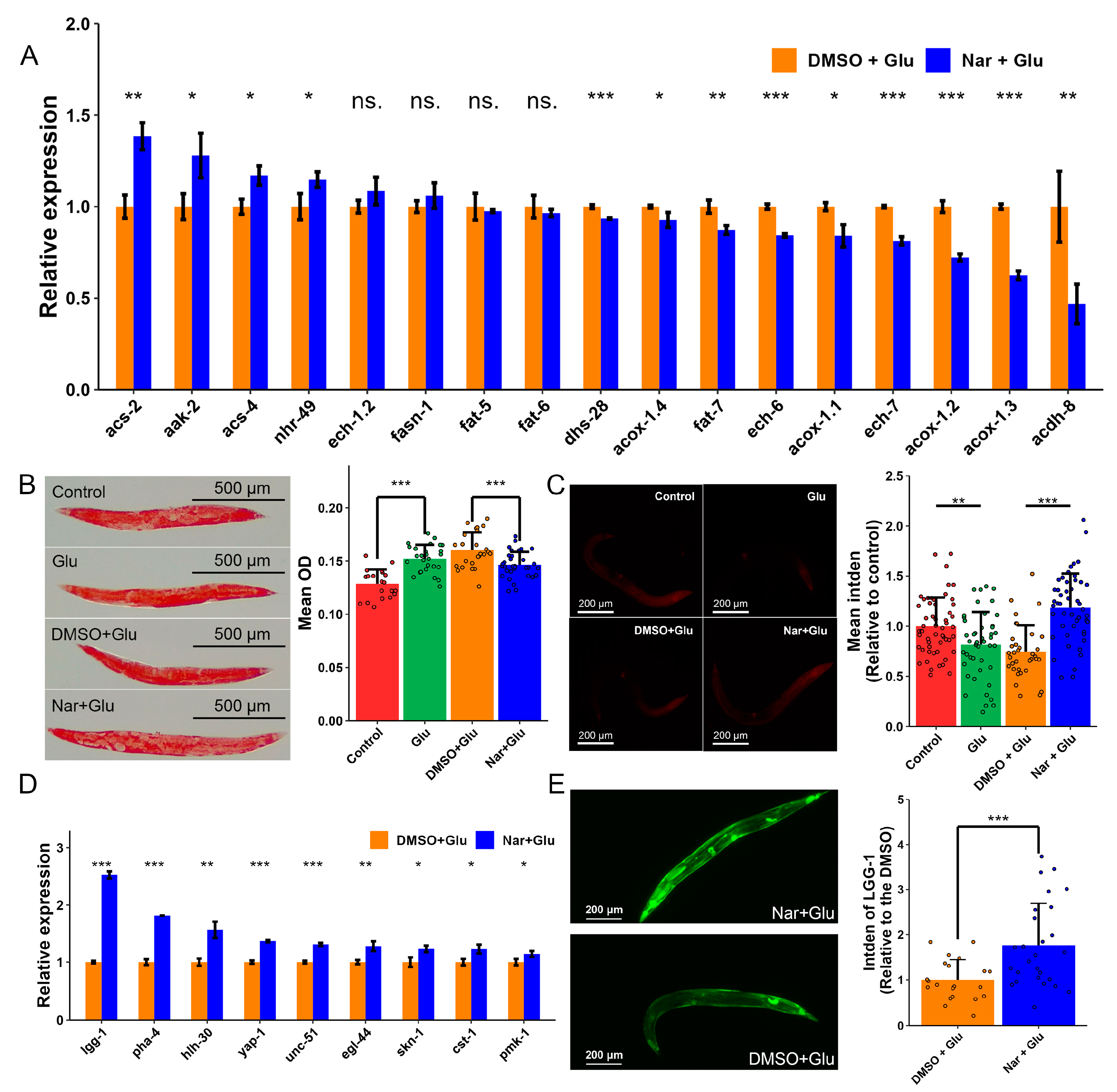
3.4. Reduction in Fat Accumulation in HGI Worms
3.5. Induced Autophagy against High Concentration of Glucose
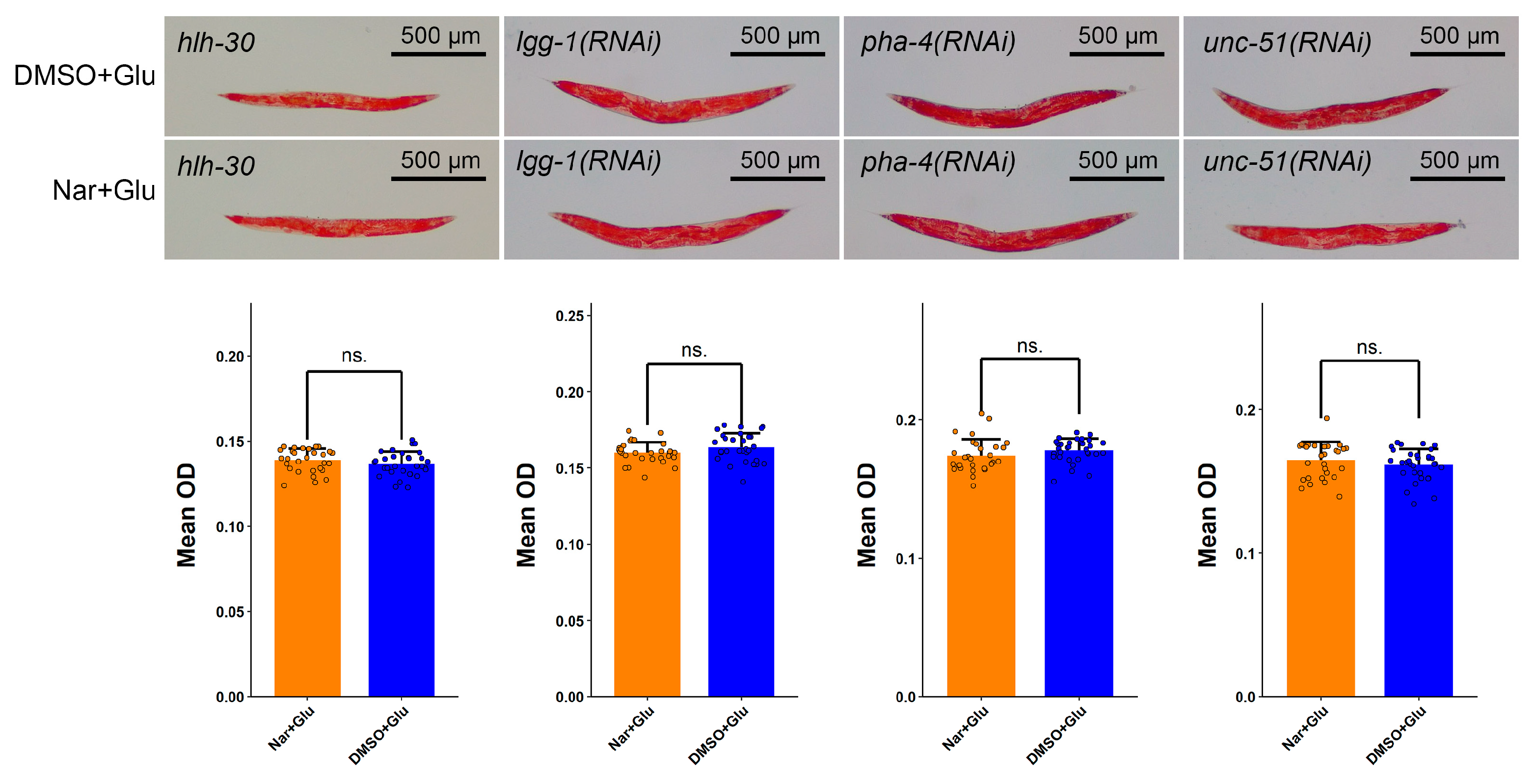
3.6. Reduced Fat Accumulation by Nar Might Be through Autophagy Pathway
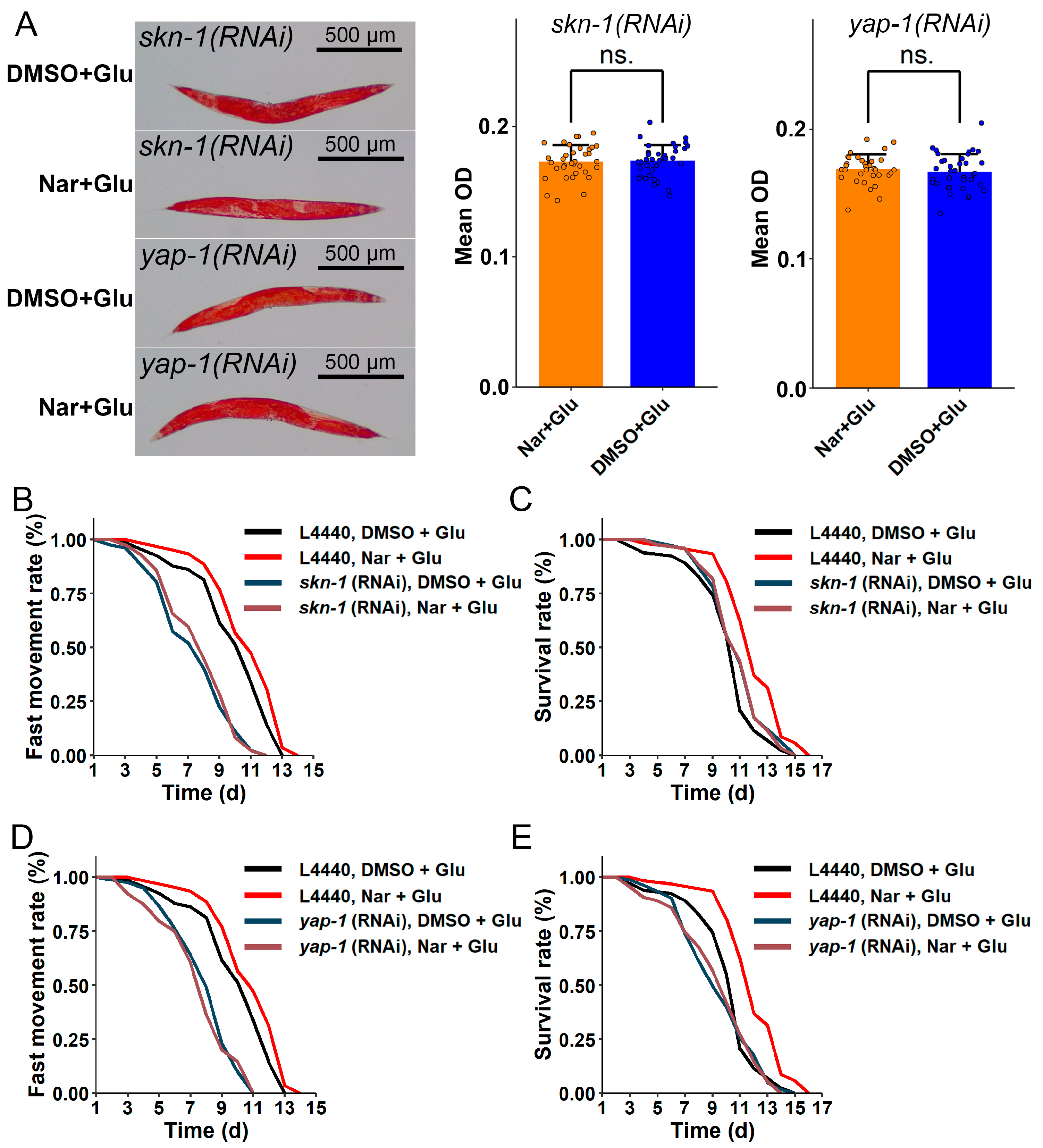
3.7. skn-1 and yap-1 Genes Mediating Fat Reduction Might Be Involved in Delaying Glucose-Induced Aging
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calcinotto, A.; Kohli, J.; Zagato, E.; Pellegrini, L.; Demaria, M.; Alimonti, A. Cellular Senescence: Aging, Cancer, and Injury. Physiol. Rev. 2019, 99, 1047–1078. [Google Scholar] [CrossRef]
- Costantino, S.; Paneni, F.; Cosentino, F. Ageing, metabolism and cardiovascular disease. J. Physiol. 2016, 594, 2061–2073. [Google Scholar] [CrossRef]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Bentley, R.A.; Ross, C.N.; O’Brien, M.J. Obesity, Metabolism, and Aging: A Multiscalar Approach. Prog. Mol. Biol. Transl. Sci. 2018, 155, 25–42. [Google Scholar] [PubMed]
- Ronan, L.; Alexander-Bloch, A.F.; Wagstyl, K.; Farooqi, S.; Brayne, C.; Tyler, L.K.; Cam, C.A.N.; Fletcher, P.C. Obesity associated with increased brain age from midlife. Neurobiol. Aging 2016, 47, 63–70. [Google Scholar] [CrossRef]
- Meccariello, R.; D’Angelo, S. Impact of Polyphenolic-Food on Longevity: An Elixir of Life. An Overview. Antioxidants 2021, 10, 507. [Google Scholar] [CrossRef] [PubMed]
- Csekes, E.; Rackova, L. Skin Aging, Cellular Senescence and Natural Polyphenols. Int. J. Mol. Sci. 2021, 22, 12641. [Google Scholar] [CrossRef]
- Hong, M.; Yu, J.; Wang, X.; Liu, Y.; Zhan, S.; Wu, Z.; Zhang, X. Tea Polyphenols as Prospective Natural Attenuators of Brain Aging. Nutrients 2022, 14, 3012. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, M.; Liu, H.; Suo, R.; Lu, C. Naringin protects endothelial cells from apoptosis and inflammation by regulating the Hippo-YAP Pathway. Biosci. Rep. 2020, 40, BSR20193431. [Google Scholar] [CrossRef]
- Barajas-Vega, J.L.; Raffoul-Orozco, A.K.; Hernandez-Molina, D.; Avila-Gonzalez, A.E.; Garcia-Cobian, T.A.; Rubio-Arellano, E.D.; Ramirez-Lizardo, E.J. Naringin reduces body weight, plasma lipids and increases adiponectin levels in patients with dyslipidemia. Int. J. Vitam. Nutr. Res. 2022, 92, 292–298. [Google Scholar] [CrossRef]
- Zheng, J.; Greenway, F.L. Caenorhabditis elegans as a model for obesity research. Int. J. Obes. 2012, 36, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Schlotterer, A.; Kukudov, G.; Bozorgmehr, F.; Hutter, H.; Du, X.; Oikonomou, D.; Ibrahim, Y.; Pfisterer, F.; Rabbani, N.; Thornalley, P.; et al. C. elegans as model for the study of high glucose- mediated life span reduction. Diabetes 2009, 58, 2450–2456. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.J.; Huang, C.; Hughes, S.; Kornfeld, K. The measurement and analysis of age-related changes in Caenorhabditis elegans. WormBook 2008, 1–21. [Google Scholar] [CrossRef]
- Yue, Y.; Hao, G.; Cho, J.; Park, Y. Curcumin reduced fat accumulation in Caenorhabditis elegans. Curr. Res. Food Sci. 2021, 4, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Li, J.; Pan, R.; Zhu, Y.; Xiao, X.; Li, Y.; Li, C. Polysaccharides from Volvariella volvacea inhibit fat accumulation in C. elegans dependent on the aak-2/nhr-49-mediated pathway. J. Food Biochem. 2021, 45, e13912. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Li, J.; Zou, X.; Greggain, J.; Rodkaer, S.V.; Faergeman, N.J.; Liang, B.; Watts, J.L. Regulation of lipid droplet size and phospholipid composition by stearoyl-CoA desaturase. J. Lipid Res. 2013, 54, 2504–2514. [Google Scholar] [CrossRef]
- Palmisano, N.J.; Melendez, A. Autophagy in C. elegans development. Dev. Biol. 2019, 447, 103–125. [Google Scholar] [CrossRef]
- Fu, M.; Zhang, X.; Zhang, X.; Yang, L.; Luo, S.; Liu, H. Autophagy Plays a Role in the Prolongation of the Life Span of Caenorhabditis elegans by Astaxanthin. Rejuvenation Res. 2021, 24, 198–205. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, P.; Chen, X.; Xie, Y.; Weston-Green, K.; Solowij, N.; Chew, Y.L.; Huang, X.F. Cannabidiol induces autophagy and improves neuronal health associated with SIRT1 mediated longevity. Geroscience 2022, 44, 1505–1524. [Google Scholar] [CrossRef]
- Lapierre, L.R.; Melendez, A.; Hansen, M. Autophagy links lipid metabolism to longevity in C. elegans. Autophagy 2012, 8, 144–146. [Google Scholar] [CrossRef]
- Mailler, E.; Guardia, C.M.; Bai, X.; Jarnik, M.; Williamson, C.D.; Li, Y.; Maio, N.; Golden, A.; Bonifacino, J.S. The autophagy protein ATG9A enables lipid mobilization from lipid droplets. Nat. Commun. 2021, 12, 6750. [Google Scholar] [CrossRef]
- Bai, J.; Zhu, Y.; He, L.; Zhang, J.; Li, J.; Pan, R.; Zhang, J.; Zhao, Y.; Cui, L.; Lu, H.; et al. Saponins from bitter melon reduce lipid accumulation via induction of autophagy in C. elegans and HepG2 cell line. Curr. Res. Food Sci. 2022, 5, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Qu, Y.; Zhou, X.G.; Chen, J.N.; Luo, H.R.; Wu, G.S. A Dihydroflavonoid Naringin Extends the Lifespan of C. elegans and Delays the Progression of Aging-Related Diseases in PD/AD Models via DAF-16. Oxidative Med. Cell. Longev. 2020, 2020, 6069354. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Hibshman, J.D.; Webster, A.K.; Baugh, L.R. Liquid-culture protocols for synchronous starvation, growth, dauer formation, and dietary restriction of Caenorhabditis elegans. STAR Protoc. 2021, 2, 100276. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Murphy, C.T.; Kenyon, C. Glucose shortens the life span of C. elegans by downregulating DAF-16/FOXO activity and aquaporin gene expression. Cell Metab. 2009, 10, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Xiong, C.; Kornfeld, K. Measurements of age-related changes of physiological processes that predict lifespan of Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2004, 101, 8084–8089. [Google Scholar] [CrossRef] [PubMed]
- Walhout, A.J.; Temple, G.F.; Brasch, M.A.; Hartley, J.L.; Lorson, M.A.; van den Heuvel, S.; Vidal, M. GATEWAY recombinational cloning: Application to the cloning of large numbers of open reading frames or ORFeomes. Methods Enzymol. 2000, 328, 575–592. [Google Scholar]
- Escorcia, W.; Ruter, D.L.; Nhan, J.; Curran, S.P. Quantification of Lipid Abundance and Evaluation of Lipid Distribution in Caenorhabditis elegans by Nile Red and Oil Red O Staining. J. Vis. Exp. 2018, 133, e57352. [Google Scholar]
- Raizen, D.; Song, B.M.; Trojanowski, N.; You, Y.J. Methods for measuring pharyngeal behaviors. WormBook 2012, 1–13. [Google Scholar] [CrossRef]
- Vernochet, C.; Kahn, C.R. Mitochondria, obesity and aging. Aging 2012, 4, 859–860. [Google Scholar] [CrossRef]
- Dilberger, B.; Baumanns, S.; Schmitt, F.; Schmiedl, T.; Hardt, M.; Wenzel, U.; Eckert, G.P. Mitochondrial Oxidative Stress Impairs Energy Metabolism and Reduces Stress Resistance and Longevity of C. elegans. Oxidative Med. Cell Longev. 2019, 2019, 6840540. [Google Scholar] [CrossRef]
- Mottram, L.F.; Forbes, S.; Ackley, B.D.; Peterson, B.R. Hydrophobic analogues of rhodamine B and rhodamine 101: Potent fluorescent probes of mitochondria in living C. elegans. Beilstein J. Org. Chem. 2012, 8, 2156–2165. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Frazee, A.C.; Pertea, G.; Jaffe, A.E.; Langmead, B.; Salzberg, S.L.; Leek, J.T. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat. Biotechnol. 2015, 33, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.O.; Dong, R.; Zhang, Y.; Zhang, J.L.; Luo, Z.; Zhang, J.; Chen, L.L.; Yang, L. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res. 2016, 26, 1277–1287. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Analysis and Normalization of Real-Time Polymerase Chain Reaction (PCR) Experimental Data. Cold Spring Harb. Protoc. 2018, 10, pdb.top095000. [Google Scholar] [CrossRef]
- Fang, E.F.; Waltz, T.B.; Kassahun, H.; Lu, Q.; Kerr, J.S.; Morevati, M.; Fivenson, E.M.; Wollman, B.N.; Marosi, K.; Wilson, M.A.; et al. Tomatidine enhances lifespan and healthspan in C. elegans through mitophagy induction via the SKN-1/Nrf2 pathway. Sci. Rep. 2017, 7, 46208. [Google Scholar] [CrossRef] [PubMed]
- Smita, S.S.; Trivedi, S.; Pandey, T.; Trivedi, M.; Pandey, R. A Bioactive compound Shatavarin IV-mediated longevity as revealed by dietary restriction-induced autophagy in Caenorhabditis elegans. Biogerontology 2020, 21, 827–844. [Google Scholar] [CrossRef] [PubMed]
- Pattarachotanant, N.; Sornkaew, N.; Warayanon, W.; Rangsinth, P.; Sillapachaiyaporn, C.; Vongthip, W.; Chuchawankul, S.; Prasansuklab, A.; Tencomnao, T. Aquilaria crassna Leaf Extract Ameliorates Glucose-Induced Neurotoxicity In Vitro and Improves Lifespan in Caenorhabditis elegans. Nutrients 2022, 14, 3668. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, L.R.; De Magalhaes Filho, C.D.; McQuary, P.R.; Chu, C.C.; Visvikis, O.; Chang, J.T.; Gelino, S.; Ong, B.; Davis, A.E.; Irazoqui, J.E.; et al. The TFEB orthologue HLH-30 regulates autophagy and modulates longevity in Caenorhabditis elegans. Nat. Commun. 2013, 4, 2267. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, L.R.; Silvestrini, M.J.; Nunez, L.; Ames, K.; Wong, S.; Le, T.T.; Hansen, M.; Melendez, A. Autophagy genes are required for normal lipid levels in C. elegans. Autophagy 2013, 9, 278–286. [Google Scholar] [CrossRef]
- Okoro, N.O.; Odiba, A.S.; Osadebe, P.O.; Omeje, E.O.; Liao, G.; Fang, W.; Jin, C.; Wang, B. Bioactive Phytochemicals with Anti-Aging and Lifespan Extending Potentials in Caenorhabditis elegans. Molecules 2021, 26, 7323. [Google Scholar] [CrossRef]
- Morselli, E.; Maiuri, M.C.; Markaki, M.; Megalou, E.; Pasparaki, A.; Palikaras, K.; Criollo, A.; Galluzzi, L.; Malik, S.A.; Vitale, I.; et al. Caloric restriction and resveratrol promote longevity through the Sirtuin-1-dependent induction of autophagy. Cell Death Dis. 2010, 1, e10. [Google Scholar] [CrossRef]
- Aranaz, P.; Navarro-Herrera, D.; Zabala, M.; Romo-Hualde, A.; Lopez-Yoldi, M.; Vizmanos, J.L.; Milagro, F.I.; Gonzalez-Navarro, C.J. Phenolic Compounds Reduce the Fat Content in Caenorhabditis elegans by Affecting Lipogenesis, Lipolysis, and Different Stress Responses. Pharmaceuticals 2020, 13, 355. [Google Scholar] [CrossRef]
- Steinbaugh, M.J.; Narasimhan, S.D.; Robida-Stubbs, S.; Moronetti Mazzeo, L.E.; Dreyfuss, J.M.; Hourihan, J.M.; Raghavan, P.; Operana, T.N.; Esmaillie, R.; Blackwell, T.K. Lipid-mediated regulation of SKN-1/Nrf in response to germ cell absence. eLife 2015, 4, e07836. [Google Scholar] [CrossRef]
- Pang, S.; Lynn, D.A.; Lo, J.Y.; Paek, J.; Curran, S.P. SKN-1 and Nrf2 couples proline catabolism with lipid metabolism during nutrient deprivation. Nat. Commun. 2014, 5, 5048. [Google Scholar] [CrossRef]
- Zhang, X.; Li, W.; Tang, Y.; Lin, C.; Cao, Y.; Chen, Y. Mechanism of Pentagalloyl Glucose in Alleviating Fat Accumulation in Caenorhabditis elegans. J. Agric. Food Chem. 2019, 67, 14110–14120. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kang, J.; Ahn, S.; Lee, J. The Hippo Pathway Is Essential for Maintenance of Apicobasal Polarity in the Growing Intestine of Caenorhabditis elegans. Genetics 2019, 213, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Saul, N.; Dhondt, I.; Kuokkanen, M.; Perola, M.; Verschuuren, C.; Wouters, B.; von Chrzanowski, H.; De Vos, W.H.; Temmerman, L.; Luyten, W.; et al. Identification of healthspan-promoting genes in Caenorhabditis elegans based on a human GWAS study. Biogerontology 2022, 23, 431–452. [Google Scholar] [CrossRef] [PubMed]
- Van Gilst, M.R.; Hadjivassiliou, H.; Jolly, A.; Yamamoto, K.R. Nuclear hormone receptor NHR-49 controls fat consumption and fatty acid composition in C. elegans. PLoS Biol. 2005, 3, e53. [Google Scholar] [CrossRef]
- Moreno-Arriola, E.; El Hafidi, M.; Ortega-Cuellar, D.; Carvajal, K. AMP-Activated Protein Kinase Regulates Oxidative Metabolism in Caenorhabditis elegans through the NHR-49 and MDT-15 Transcriptional Regulators. PLoS ONE 2016, 11, e0148089. [Google Scholar] [CrossRef]
- Watts, J.L.; Ristow, M. Lipid and Carbohydrate Metabolism in Caenorhabditis elegans. Genetics 2017, 207, 413–446. [Google Scholar]
- Yue, Y.; Shen, P.; Chang, A.L.; Qi, W.; Kim, K.H.; Kim, D.; Park, Y. trans-Trismethoxy resveratrol decreased fat accumulation dependent on fat-6 and fat-7 in Caenorhabditis elegans. Food Funct. 2019, 10, 4966–4974. [Google Scholar] [CrossRef]
- Liu, Y.J.; Gao, A.W.; Smith, R.L.; Janssens, G.E.; Panneman, D.M.; Jongejan, A.; van Weeghel, M.; Vaz, F.M.; Silvestrini, M.J.; Lapierre, L.R.; et al. Reduced ech-6 expression attenuates fat-induced lifespan shortening in C. elegans. Sci. Rep. 2022, 12, 3350. [Google Scholar] [CrossRef]
- Gumienny, T.L.; Savage-Dunn, C. TGF-beta signaling in C. elegans. WormBook 2013, 1–34. [Google Scholar] [CrossRef]
- Pari, L.; Chandramohan, R. Modulatory effects of naringin on hepatic key enzymes of carbohydrate metabolism in high-fat diet/low-dose streptozotocin-induced diabetes in rats. Gen. Physiol. Biophys. 2017, 36, 343–352. [Google Scholar] [CrossRef]
- Murillo, K.; Samigullin, A.; Humpert, P.M.; Fleming, T.; Ozer, K.; Schlotterer, A.; Hammes, H.P.; Morcos, M. Protective Effects of Transient Glucose Exposure in Adult C. elegans. Antioxidants 2022, 11, 160. [Google Scholar] [CrossRef]
- Wang, C.; Chan, J.S.; Ren, L.; Yan, J.H. Obesity Reduces Cognitive and Motor Functions across the Lifespan. Neural Plast. 2016, 2016, 2473081. [Google Scholar] [CrossRef] [PubMed]
- Landecho, M.F.; Marin-Oto, M.; Recalde-Zamacona, B.; Bilbao, I.; Fruhbeck, G. Obesity as an adipose tissue dysfunction disease and a risk factor for infections—COVID-19 as a case study. Eur. J. Intern. Med. 2021, 91, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Seravalle, G.; Grassi, G. Obesity and hypertension. Pharmacol. Res. 2017, 122, 1–7. [Google Scholar] [CrossRef]
- Singh, R.K.; Kumar, P.; Mahalingam, K. Molecular genetics of human obesity: A comprehensive review. Comptes Rendus Biol. 2017, 340, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Jura, M.; Kozak, L.P. Obesity and related consequences to ageing. Age 2016, 38, 23. [Google Scholar] [CrossRef]
- Kitada, M.; Koya, D. Autophagy in metabolic disease and ageing. Nat. Rev. Endocrinol. 2021, 17, 647–661. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N | Events | Mean ± SE | Median | p-Value |
|---|---|---|---|---|---|
| Control | 70 | 53 | 14.95 ± 0.57 | 14 | - |
| Glu | 81 | 74 | 12.09 ± 0.57 | 12 | 0.005 |
| DMSO + Glu | 92 | 74 | 11.29 ± 0.54 | 12 | - |
| 25 μM Nar + Glu | 64 | 51 | 14.89 ± 0.60 | 15 | <0.001 |
| 50 μM Nar + Glu | 81 | 69 | 11.42 ± 0.45 | 11 | 0.632 |
| 100 μM Nar + Glu | 83 | 69 | 11.87 ± 0.51 | 12 | 0.650 |
| 200 μM Nar + Glu | 65 | 52 | 11.38 ± 0.60 | 12 | 0.921 |
| 500 μM Nar + Glu | 68 | 54 | 12.73 ± 0.58 | 14 | 0.186 |
| Group | N | Events | Mean ± SE | Median | p-Value |
|---|---|---|---|---|---|
| Control | 90 | 72 | 11.67 ± 0.22 | 12 | - |
| Glu | 81 | 62 | 10.28 ± 0.23 | 10 | <0.001 |
| DMSO + Glu | 74 | 54 | 10.01 ± 0.25 | 10 | - |
| 25 μM + Glu | 77 | 44 | 11.25 ± 0.32 | 13 | 0.001 |
| 50 μM + Glu | 78 | 55 | 10.72 ± 0.30 | 11 | 0.060 |
| Strain | Group | N | Events | Mean ± SE | Median | p-Value |
|---|---|---|---|---|---|---|
| hlh-30 | DMSO + Glu | 70 | 59 | 12.05 ± 0.46 | 13 | |
| Nar + Glu | 73 | 56 | 11.95 ± 0.47 | 12 | 0.940 | |
| lgg-1 (RNAi) | DMSO + Glu | 80 | 62 | 10.21 ± 0.29 | 10 | |
| Nar + Glu | 82 | 59 | 10.61 ± 0.36 | 11 | 0.146 | |
| L4440 | DMSO + Glu | 67 | 56 | 10.25 ± 0.31 | 11 | |
| Nar + Glu | 62 | 47 | 12.07 ± 0.29 | 12 | <0.001 | |
| pha-4 (RNAi) | DMSO + Glu | 81 | 72 | 9.36 ± 0.15 | 9 | |
| Nar + Glu | 82 | 70 | 9.17 ± 0.14 | 9 | 0.328 | |
| skn-1 (RNAi) | DMSO + Glu | 82 | 65 | 10.97 ± 0.26 | 11 | |
| Nar + Glu | 86 | 63 | 10.93 ± 0.25 | 11 | 0.703 | |
| unc-51 (RNAi) | DMSO + Glu | 83 | 68 | 9.68 ± 0.29 | 10 | |
| Nar + Glu | 84 | 64 | 9.73 ± 0.26 | 10 | 0.640 | |
| yap-1 (RNAi) | DMSO + Glu | 80 | 64 | 9.53 ± 0.33 | 9 | |
| Nar + Glu | 64 | 57 | 9.49 ± 0.37 | 10 | 0.979 |
| Strain | Group | N | Events | Mean ± SE | Median | p-Value |
|---|---|---|---|---|---|---|
| hlh-30 | DMSO + Glu | 70 | 65 | 8.05 ± 0.30 | 9 | |
| Nar + Glu | 73 | 59 | 8.61 ± 0.27 | 9 | 0.214 | |
| lgg-1 (RNAi) | DMSO + Glu | 80 | 67 | 6.39 ± 0.22 | 6 | |
| Nar + Glu | 82 | 64 | 6.48 ± 0.23 | 6 | 0.599 | |
| L4440 | DMSO + Glu | 67 | 56 | 10.03 ± 0.31 | 11 | |
| Nar + Glu | 62 | 47 | 10.89 ± 0.28 | 11 | 0.034 | |
| pha-4 (RNAi) | DMSO + Glu | 81 | 77 | 6.53 ± 0.17 | 7 | |
| Nar + Glu | 82 | 74 | 6.51 ± 0.21 | 6 | 0.353 | |
| skn-1 (RNAi) | DMSO + Glu | 82 | 68 | 7.48 ± 0.28 | 8 | |
| Nar + Glu | 86 | 64 | 7.85 ± 0.26 | 8 | 0.522 | |
| unc-51 (RNAi) | DMSO + Glu | 83 | 75 | 5.54 ± 0.24 | 6 | |
| Nar + Glu | 83 | 65 | 6.14 ± 0.23 | 7 | 0.141 | |
| yap-1 (RNAi) | DMSO + Glu | 80 | 64 | 7.99 ± 0.24 | 8 | |
| Nar + Glu | 64 | 58 | 7.65 ± 0.29 | 8 | 0.653 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, P.; Wang, P.; Liu, L.; Wang, P.; Lin, G.; Qu, Z.; Yu, Z.; Liu, N. Naringin Alleviates Glucose-Induced Aging by Reducing Fat Accumulation and Promoting Autophagy in Caenorhabditis elegans. Nutrients 2023, 15, 907. https://doi.org/10.3390/nu15040907
Guo P, Wang P, Liu L, Wang P, Lin G, Qu Z, Yu Z, Liu N. Naringin Alleviates Glucose-Induced Aging by Reducing Fat Accumulation and Promoting Autophagy in Caenorhabditis elegans. Nutrients. 2023; 15(4):907. https://doi.org/10.3390/nu15040907
Chicago/Turabian StyleGuo, Peisen, Panpan Wang, Limin Liu, Peixi Wang, Guimiao Lin, Zhi Qu, Zengli Yu, and Nan Liu. 2023. "Naringin Alleviates Glucose-Induced Aging by Reducing Fat Accumulation and Promoting Autophagy in Caenorhabditis elegans" Nutrients 15, no. 4: 907. https://doi.org/10.3390/nu15040907