
The Role of Omega-3 Polyunsaturated Fatty Acids and Their Lipid Mediators on Skeletal Muscle Regeneration: A Narrative Review
 , and
, and 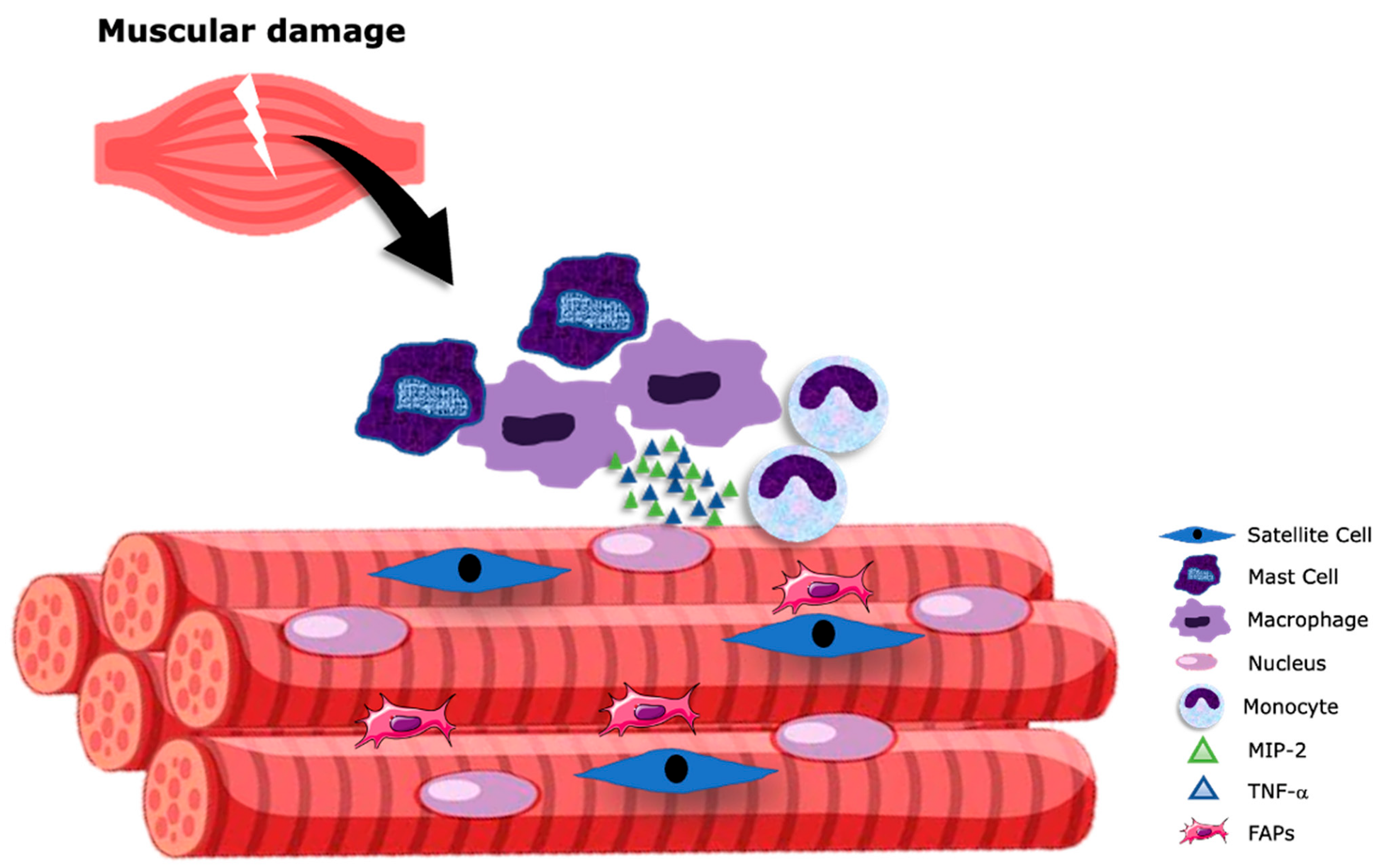{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. An Inflammatory Process Initiates Skeletal Muscle Repair
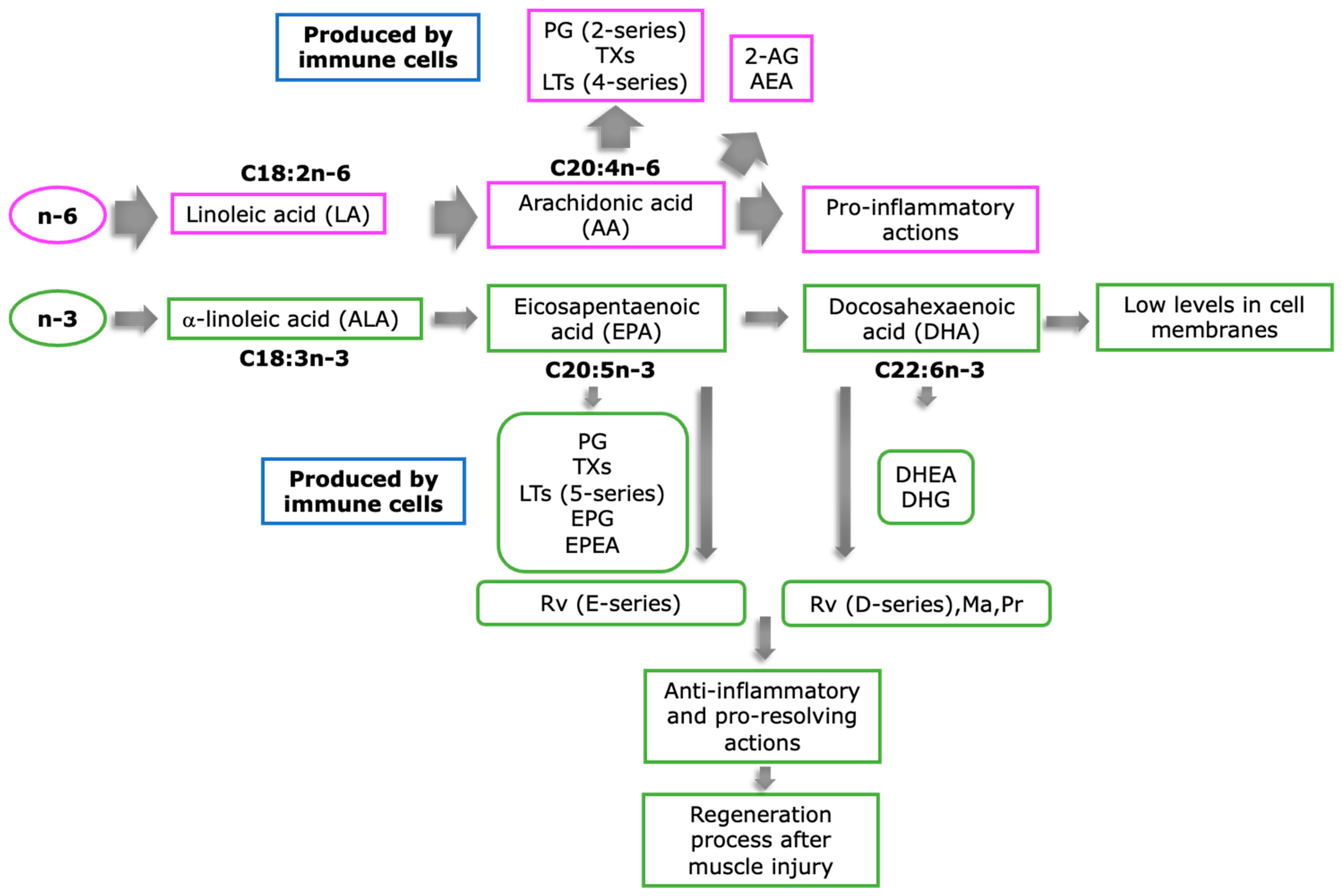
3. Role of PUFAs on Inflammation Resolution
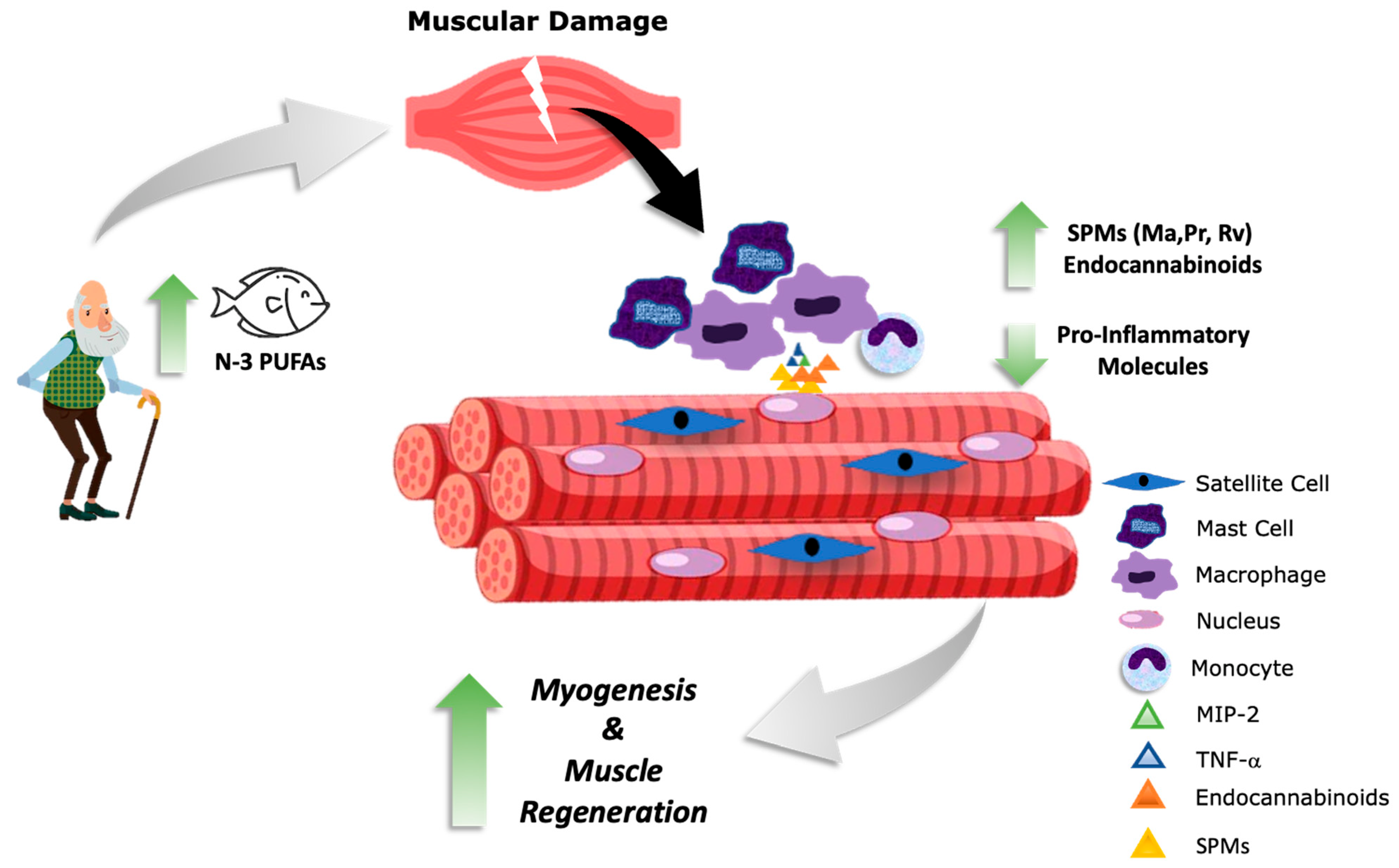
4. Potential Beneficial Effects of n3-PUFA-Derived Metabolites in Muscle Regeneration: Evidence from In Vitro Muscular Cell Lines, Adult Skeletal Fibers, and Animal Models
5. Clinical Interventions Supporting the Consumption of n-3 PUFAs to Aid Muscle Recovery
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Janssen, I.; Heymsfield, S.B.; Wang, Z.M.; Ross, R. Skeletal Muscle Mass and Distribution in 468 Men and Women Aged 18–88 Yr. J. Appl. Physiol. 2000, 89, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Järvinen, T.A.H.; Järvinen, T.L.N.; Kääriäinen, M.; Kalimo, H.; Järvinen, M. Muscle Injuries: Biology and Treatment. Am. J. Sports Med. 2005, 33, 745–764. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite Cells and the Muscle Stem Cell Niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [PubMed]
- Sinha, I.; Sakthivel, D.; Varon, D.E. Systemic Regulators of Skeletal Muscle Regeneration in Obesity. Front. Endocrinol. 2017, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Domingues-Faria, C.; Vasson, M.-P.; Goncalves-Mendes, N.; Boirie, Y.; Walrand, S. Skeletal Muscle Regeneration and Impact of Aging and Nutrition. Ageing Res. Rev. 2016, 26, 22–36. [Google Scholar] [CrossRef]
- Almada, A.E.; Wagers, A.J. Molecular Circuitry of Stem Cell Fate in Skeletal Muscle Regeneration, Ageing and Disease. Nat. Rev. Mol. Cell Biol. 2016, 17, 267–279. [Google Scholar] [CrossRef]
- Tedesco, F.S.; Dellavalle, A.; Diaz-Manera, J.; Messina, G.; Cossu, G. Repairing Skeletal Muscle: Regenerative Potential of Skeletal Muscle Stem Cells. J. Clin. Investig. 2010, 120, 11–19. [Google Scholar] [CrossRef]
- Allbrook, D.B.; Han, M.F.; Hellmuth, A.E. Population of Muscle Satellite Cells in Relation to Age and Mitotic Activity. Pathology 1971, 3, 223–243. [Google Scholar] [CrossRef]
- Mauro, A. Satellite Cell of Skeletal Muscle Fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef]
- Theret, M.; Rossi, F.M.V.; Contreras, O. Evolving Roles of Muscle-Resident Fibro-Adipogenic Progenitors in Health, Regeneration, Neuromuscular Disorders, and Aging. Front. Physiol. 2021, 12, 673404. [Google Scholar] [CrossRef]
- Wosczyna, M.N.; Rando, T.A. A Muscle Stem Cell Support Group: Coordinated Cellular Responses in Muscle Regeneration. Dev. Cell 2018, 46, 135–143. [Google Scholar] [CrossRef]
- Markworth, J.F.; Maddipati, K.R.; Cameron-Smith, D. Emerging Roles of Pro-Resolving Lipid Mediators in Immunological and Adaptive Responses to Exercise-Induced Muscle Injury. Exerc. Immunol. Rev. 2016, 22, 110–134. [Google Scholar]
- Bentzinger, C.F.; Wang, Y.X.; Dumont, N.A.; Rudnicki, M.A. Cellular Dynamics in the Muscle Satellite Cell Niche. EMBO Rep. 2013, 14, 1062–1072. [Google Scholar] [CrossRef]
- Heredia, J.E.; Mukundan, L.; Chen, F.M.; Mueller, A.A.; Deo, R.C.; Locksley, R.M.; Rando, T.A.; Chawla, A. Type 2 Innate Signals Stimulate Fibro/Adipogenic Progenitors to Facilitate Muscle Regeneration. Cell 2013, 153, 376–388. [Google Scholar] [CrossRef]
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in Our Understanding of Oxylipins Derived from Dietary PUFAs. Adv. Nutr. 2015, 6, 513–540. [Google Scholar] [CrossRef]
- Serhan, C.N.; Dalli, J.; Colas, R.A.; Winkler, J.W.; Chiang, N. Protectins and Maresins: New pro-Resolving Families of Mediators in Acute Inflammation and Resolution Bioactive Metabolome. Biochim. Biophys. Acta 2015, 1851, 397–413. [Google Scholar] [CrossRef]
- Tidball, J.G.; Villalta, S.A. Regulatory Interactions between Muscle and the Immune System during Muscle Regeneration. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R1173–R1187. [Google Scholar] [CrossRef]
- McKenna, C.F.; Fry, C.S. Altered Satellite Cell Dynamics Accompany Skeletal Muscle Atrophy during Chronic Illness, Disuse, and Aging. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 447–452. [Google Scholar] [CrossRef]
- Tidball, J.G. Regulation of Muscle Growth and Regeneration by the Immune System. Nat. Rev. Immunol. 2017, 17, 165–178. [Google Scholar] [CrossRef]
- Giannakis, N.; Sansbury, B.E.; Patsalos, A.; Hays, T.T.; Riley, C.O.; Han, X.; Spite, M.; Nagy, L. Dynamic Changes to Lipid Mediators Support Transitions among Macrophage Subtypes during Muscle Regeneration. Nat. Immunol. 2019, 20, 626–636. [Google Scholar] [CrossRef]
- Markworth, J.F.; Brown, L.A.; Lim, E.; Floyd, C.; Larouche, J.; Castor-Macias, J.A.; Sugg, K.B.; Sarver, D.C.; Macpherson, P.C.; Davis, C.; et al. Resolvin D1 Supports Skeletal Myofiber Regeneration via Actions on Myeloid and Muscle Stem Cells. JCI Insight 2020, 5, e137713. [Google Scholar] [CrossRef] [PubMed]
- Phillips, T.; Childs, A.C.; Dreon, D.M.; Phinney, S.; Leeuwenburgh, C. A Dietary Supplement Attenuates IL-6 and CRP after Eccentric Exercise in Untrained Males. Med. Sci. Sports Exerc. 2003, 35, 2032–2037. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Thorlacius, H. Mast Cell-Derived Tumour Necrosis Factor-Alpha Mediates Macrophage Inflammatory Protein-2-Induced Recruitment of Neutrophils in Mice. Br. J. Pharmacol. 2005, 145, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, C.; Micheletti, A.; Cassatella, M.A. Neutrophil-Derived Cytokines: Facts beyond Expression. Front. Immunol. 2014, 5, 508. [Google Scholar] [CrossRef]
- Molina, T.; Fabre, P.; Dumont, N.A. Fibro-Adipogenic Progenitors in Skeletal Muscle Homeostasis, Regeneration and Diseases. Open Biol. 2021, 11, 210110. [Google Scholar] [CrossRef]
- Lemos, D.R.; Babaeijandaghi, F.; Low, M.; Chang, C.-K.; Lee, S.T.; Fiore, D.; Zhang, R.-H.; Natarajan, A.; Nedospasov, S.A.; Rossi, F.M. V Nilotinib Reduces Muscle Fibrosis in Chronic Muscle Injury by Promoting TNF-Mediated Apoptosis of Fibro/Adipogenic Progenitors. Nat. Med. 2015, 21, 786–794. [Google Scholar] [CrossRef]
- Cheng, A.J.; Yamada, T.; Rassier, D.E.; Andersson, D.C.; Westerblad, H.; Lanner, J.T. Reactive Oxygen/Nitrogen Species and Contractile Function in Skeletal Muscle during Fatigue and Recovery. J. Physiol. 2016, 594, 5149–5160. [Google Scholar] [CrossRef]
- Powers, S.K.; Morton, A.B.; Ahn, B.; Smuder, A.J. Redox Control of Skeletal Muscle Atrophy. Free Radic. Biol. Med. 2016, 98, 208–217. [Google Scholar] [CrossRef]
- Vasilaki, A.; Jackson, M.J. Role of Reactive Oxygen Species in the Defective Regeneration Seen in Aging Muscle. Free Radic. Biol. Med. 2013, 65, 317–323. [Google Scholar] [CrossRef]
- Calder, P.C. Fatty Acids and Inflammation: The Cutting Edge between Food and Pharma. Eur. J. Pharmacol. 2011, 668, 50–58. [Google Scholar] [CrossRef]
- Tang, J.; Yang, B.; Yan, Y.; Tong, W.; Zhou, R.; Zhang, J.; Mi, J.; Li, D. Palmitoleic Acid Protects against Hypertension by Inhibiting NF-ΚB-Mediated Inflammation. Mol. Nutr. Food Res. 2021, 65, e2001025. [Google Scholar] [CrossRef]
- Gorzynik-Debicka, M.; Przychodzen, P.; Cappello, F.; Kuban-Jankowska, A.; Marino Gammazza, A.; Knap, N.; Wozniak, M.; Gorska-Ponikowska, M. Potential Health Benefits of Olive Oil and Plant Polyphenols. Int. J. Mol. Sci. 2018, 19, 686. [Google Scholar] [CrossRef]
- González-Mañán, D.; Tapia, G.; Gormaz, J.G.; D’Espessailles, A.; Espinosa, A.; Masson, L.; Varela, P.; Valenzuela, A.; Valenzuela, R. Bioconversion of α-Linolenic Acid to n-3 LCPUFA and Expression of PPAR-Alpha, Acyl Coenzyme A Oxidase 1 and Carnitine Acyl Transferase I Are Incremented after Feeding Rats with α-Linolenic Acid-Rich Oils. Food Funct. 2012, 3, 765–772. [Google Scholar] [CrossRef]
- Rincón-Cervera, M.Á.; Valenzuela, R.; Hernandez-Rodas, M.C.; Barrera, C.; Espinosa, A.; Marambio, M.; Valenzuela, A. Vegetable Oils Rich in Alpha Linolenic Acid Increment Hepatic N-3 LCPUFA, Modulating the Fatty Acid Metabolism and Antioxidant Response in Rats. Prostaglandins. Leukot. Essent. Fatty Acids 2016, 111, 25–35. [Google Scholar] [CrossRef]
- Videla, L.A.; Hernandez-Rodas, M.C.; Metherel, A.H.; Valenzuela, R. Influence of the Nutritional Status and Oxidative Stress in the Desaturation and Elongation of N-3 and n-6 Polyunsaturated Fatty Acids: Impact on Non-Alcoholic Fatty Liver Disease. Prostaglandins. Leukot. Essent. Fatty Acids 2022, 181, 102441. [Google Scholar] [CrossRef]
- Calder, P.C.; Grimble, R.F. Polyunsaturated Fatty Acids, Inflammation and Immunity. Eur. J. Clin. Nutr. 2002, 56, S14–S19. [Google Scholar] [CrossRef]
- Gerling, C.J.; Mukai, K.; Chabowski, A.; Heigenhauser, G.J.F.; Holloway, G.P.; Spriet, L.L.; Jannas-Vela, S. Incorporation of Omega-3 Fatty Acids into Human Skeletal Muscle Sarcolemmal and Mitochondrial Membranes Following 12 Weeks of Fish Oil Supplementation. Front. Physiol. 2019, 10, 348. [Google Scholar] [CrossRef]
- Calder, P.C. Eicosapentaenoic and Docosahexaenoic Acid Derived Specialised Pro-Resolving Mediators: Concentrations in Humans and the Effects of Age, Sex, Disease and Increased Omega-3 Fatty Acid Intake. Biochimie 2020, 178, 105–123. [Google Scholar] [CrossRef]
- Egan, R.W.; Kuehl, F.A. Prostaglandins, Arachidonic Acid, and Inflammation. Science 1980, 210, 978–984. [Google Scholar] [CrossRef]
- Kuda, O. Bioactive Metabolites of Docosahexaenoic Acid. Biochimie 2017, 136, 12–20. [Google Scholar] [CrossRef]
- Serhan, C.N. Novel Omega—3-Derived Local Mediators in Anti-Inflammation and Resolution. Pharmacol. Ther. 2005, 105, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Echeverría, F.; Valenzuela, R.; Espinosa, A.; Bustamante, A.; Álvarez, D.; Gonzalez-Mañan, D.; Ortiz, M.; Soto-Alarcon, S.A.; Videla, L.A. Reduction of High-Fat Diet-Induced Liver Proinflammatory State by Eicosapentaenoic Acid plus Hydroxytyrosol Supplementation: Involvement of Resolvins RvE1/2 and RvD1/2. J. Nutr. Biochem. 2019, 63, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga-Hernández, J.; Sambra, V.; Echeverría, F.; Videla, L.A.; Valenzuela, R. N-3 PUFAs and Their Specialized pro-Resolving Lipid Mediators on Airway Inflammatory Response: Beneficial Effects in the Prevention and Treatment of Respiratory Diseases. Food Funct. 2022, 13, 4260–4272. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Bioactive Lipids in Age-Related Disorders. Adv. Exp. Med. Biol. 2020, 1260, 33–83. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Pro-Resolving Lipid Mediators Are Leads for Resolution Physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef]
- McDougle, D.R.; Watson, J.E.; Abdeen, A.A.; Adili, R.; Caputo, M.P.; Krapf, J.E.; Johnson, R.W.; Kilian, K.A.; Holinstat, M.; Das, A. Anti-Inflammatory ω-3 Endocannabinoid Epoxides. Proc. Natl. Acad. Sci. USA 2017, 114, E6034–E6043. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-6 and Omega-3 Fatty Acids: Endocannabinoids, Genetics and Obesity. OCL 2020, 27, 7. [Google Scholar] [CrossRef]
- Mackie, K. Cannabinoid Receptors: Where They Are and What They Do. J. Neuroendocrinol. 2008, 20, 10–14. [Google Scholar] [CrossRef]
- Carnevale, L.N.; Das, A. Novel Anti-Inflammatory and Vasodilatory ω-3 Endocannabinoid Epoxide Regioisomers. Adv. Exp. Med. Biol. 2019, 1161, 219–232. [Google Scholar] [CrossRef]
- Shahbazi, F.; Grandi, V.; Banerjee, A.; Trant, J.F. Cannabinoids and Cannabinoid Receptors: The Story so Far. iScience 2020, 23, 101301. [Google Scholar] [CrossRef]
- Aguirre, C.A.; Castillo, V.A.; Llanos, M.N. The Endocannabinoid Anandamide during Lactation Increases Body Fat Content and CB1 Receptor Levels in Mice Adipose Tissue. Nutr. Diabetes 2015, 5, e167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, J.E.; Kim, J.S.; Das, A. Emerging Class of Omega-3 Fatty Acid Endocannabinoids & Their Derivatives. Prostaglandins Other Lipid Mediat. 2019, 143, 106337. [Google Scholar] [CrossRef]
- Zelasko, S.; Arnold, W.R.; Das, A. Endocannabinoid Metabolism by Cytochrome P450 Monooxygenases. Prostaglandins Other Lipid Mediat. 2015, 116–117, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Rouzer, C.A.; Marnett, L.J. Endocannabinoid Oxygenation by Cyclooxygenases, Lipoxygenases, and Cytochromes P450: Cross-Talk between the Eicosanoid and Endocannabinoid Signaling Pathways. Chem. Rev. 2011, 111, 5899–5921. [Google Scholar] [CrossRef] [PubMed]
- Risha, M.A.; Siengdee, P.; Dannenberger, D.; Wimmers, K.; Ponsuksili, S. PUFA Treatment Affects C2C12 Myocyte Differentiation, Myogenesis Related Genes and Energy Metabolism. Genes 2021, 12, 192. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, T.-Y.; Baum, J.I.; Huang, Y. Effect of Eicosapentaenoic Acid and Docosahexaenoic Acid on Myogenesis and Mitochondrial Biosynthesis during Murine Skeletal Muscle Cell Differentiation. Front. Nutr. 2018, 5, 15. [Google Scholar] [CrossRef]
- Haghani, K.; Pashaei, S.; Vakili, S.; Taheripak, G.; Bakhtiyari, S. TNF-α Knockdown Alleviates Palmitate-Induced Insulin Resistance in C2C12 Skeletal Muscle Cells. Biochem. Biophys. Res. Commun. 2015, 460, 977–982. [Google Scholar] [CrossRef]
- Baker, L.A.; Martin, N.R.W.; Kimber, M.C.; Pritchard, G.J.; Lindley, M.R.; Lewis, M.P. Resolvin E1 (R(v) E(1)) Attenuates LPS Induced Inflammation and Subsequent Atrophy in C2C12 Myotubes. J. Cell. Biochem. 2018, 119, 6094–6103. [Google Scholar] [CrossRef]
- Machado, R.V.; Mauricio, A.F.; Taniguti, A.P.T.; Ferretti, R.; Neto, H.S.; Marques, M.J. Eicosapentaenoic Acid Decreases TNF-α and Protects Dystrophic Muscles of Mdx Mice from Degeneration. J. Neuroimmunol. 2011, 232, 145–150. [Google Scholar] [CrossRef]
- Yang, B.; Lin, L.; Bazinet, R.P.; Chien, Y.-C.; Chang, J.P.-C.; Satyanarayanan, S.K.; Su, H.; Su, K.-P. Clinical Efficacy and Biological Regulations of ω-3 PUFA-Derived Endocannabinoids in Major Depressive Disorder. Psychother. Psychosom. 2019, 88, 215–224. [Google Scholar] [CrossRef]
- Ramsden, C.E.; Zamora, D.; Makriyannis, A.; Wood, J.T.; Mann, J.D.; Faurot, K.R.; MacIntosh, B.A.; Majchrzak-Hong, S.F.; Gross, J.R.; Courville, A.B.; et al. Diet-Induced Changes in n-3- and n-6-Derived Endocannabinoids and Reductions in Headache Pain and Psychological Distress. J. Pain 2015, 16, 707–716. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.I.; Atherton, P.; Reeds, D.N.; Mohammed, B.S.; Rankin, D.; Rennie, M.J.; Mittendorfer, B. Omega-3 Polyunsaturated Fatty Acids Augment the Muscle Protein Anabolic Response to Hyperinsulinaemia-Hyperaminoacidaemia in Healthy Young and Middle-Aged Men and Women. Clin. Sci. 2011, 121, 267–278. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Yanagimoto, K.; Nakazato, K.; Hayamizu, K.; Ochi, E. Eicosapentaenoic and Docosahexaenoic Acids-Rich Fish Oil Supplementation Attenuates Strength Loss and Limited Joint Range of Motion after Eccentric Contractions: A Randomized, Double-Blind, Placebo-Controlled, Parallel-Group Trial. Eur. J. Appl. Physiol. 2016, 116, 1179–1188. [Google Scholar] [CrossRef]
- Gollnick, P.D.; Sjödin, B.; Karlsson, J.; Jansson, E.; Saltin, B. Human Soleus Muscle: A Comparison of Fiber Composition and Enzyme Activities with Other Leg Muscles. Pflugers Arch. 1974, 348, 247–255. [Google Scholar] [CrossRef]
- Penner, A.L.; Waytt, V.; Winter, T.; Leng, S.; Duhamel, T.A.; Aukema, H.M. Oxylipin Profiles and Levels Vary by Skeletal Muscle Type, Dietary Fat and Sex in Young Rats. Appl. Physiol. Nutr. Metab. Physiol. Appl. Nutr. Metab. 2021, 46, 1378–1388. [Google Scholar] [CrossRef]
- Yamazaki, H.; Nishimura, M.; Uehara, M.; Kuribara-Souta, A.; Yamamoto, M.; Yoshikawa, N.; Morohashi, K.-I.; Tanaka, H. Eicosapentaenoic Acid Changes Muscle Transcriptome and Intervenes in Aging-Related Fiber Type Transition in Male Mice. Am. J. Physiol. Endocrinol. Metab. 2021, 320, E346–E358. [Google Scholar] [CrossRef]
- Markworth, J.F.; Brown, L.A.; Lim, E.; Castor-Macias, J.A.; Larouche, J.; Macpherson, P.C.D.; Davis, C.; Aguilar, C.A.; Maddipati, K.R.; Brooks, S.V. Metabolipidomic Profiling Reveals an Age-Related Deficiency of Skeletal Muscle pro-Resolving Mediators That Contributes to Maladaptive Tissue Remodeling. Aging Cell 2021, 20, e13393. [Google Scholar] [CrossRef]
- Jeromson, S.; Mackenzie, I.; Doherty, M.K.; Whitfield, P.D.; Bell, G.; Dick, J.; Shaw, A.; Rao, F.; Ashcroft, S.; Philp, A.; et al. Lipid Remodelling and an Altered Membrane Proteome May Drive the Effects of EPA and DHA Treatment on Skeletal Muscle Glucose Uptake and Protein Accretion. Am. J. Physiol.-Endocrinol. Metab. 2018, 314, E605–E619. [Google Scholar] [CrossRef]
- Isesele, P.O.; Mazurak, V.C. Regulation of Skeletal Muscle Satellite Cell Differentiation by Omega-3 Polyunsaturated Fatty Acids: A Critical Review. Front. Physiol. 2021, 12, 682091. [Google Scholar] [CrossRef]
- Ghnaimawi, S.; Shelby, S.; Baum, J.; Huang, Y. Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on C2C12 Cell Adipogenesis and Inhibition of Myotube Formation. Anim. Cells Syst. 2019, 23, 355–364. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, X.; Liu, Y.; Zhang, L.; Odle, J.; Lin, X.; Zhu, H.; Wang, X.; Liu, Y. EPA and DHA Inhibit Myogenesis and Downregulate the Expression of Muscle-Related Genes in C2C12 Myoblasts. Genes 2019, 10, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacham-Kaplan, O.; Camera, D.M.; Hawley, J.A. Divergent Regulation of Myotube Formation and Gene Expression by E2 and EPA during In-Vitro Differentiation of C2C12 Myoblasts. Int. J. Mol. Sci. 2020, 21, 745. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, Q.; Zheng, P.; Zhang, J.; Huang, F. DHA Inhibits Protein Degradation More Efficiently than EPA by Regulating the PPARγ/NFκB Pathway in C2C12 Myotubes. Biomed Res. Int. 2013, 2013, 318981. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, A.; Campos, C.; Díaz-Vegas, A.; Galgani, J.E.; Juretic, N.; Osorio-Fuentealba, C.; Bucarey, J.L.; Tapia, G.; Valenzuela, R.; Contreras-Ferrat, A.; et al. Insulin-Dependent H2O2 Production Is Higher in Muscle Fibers of Mice Fed with a High-Fat Diet. Int. J. Mol. Sci. 2013, 14, 15740–15754. [Google Scholar] [CrossRef] [PubMed]
- Roseno, S.L.; Davis, P.R.; Bollinger, L.M.; Powell, J.J.S.; Witczak, C.A.; Brault, J.J. Short-Term, High-Fat Diet Accelerates Disuse Atrophy and Protein Degradation in a Muscle-Specific Manner in Mice. Nutr. Metab. 2015, 12, 39. [Google Scholar] [CrossRef]
- Woodworth-Hobbs, M.E.; Hudson, M.B.; Rahnert, J.A.; Zheng, B.; Franch, H.A.; Price, S.R. Docosahexaenoic Acid Prevents Palmitate-Induced Activation of Proteolytic Systems in C2C12 Myotubes. J. Nutr. Biochem. 2014, 25, 868–874. [Google Scholar] [CrossRef]
- Bryner, R.W.; Woodworth-Hobbs, M.E.; Williamson, D.L.; Alway, S.E. Docosahexaenoic Acid Protects Muscle Cells from Palmitate-Induced Atrophy. ISRN Obes. 2012, 2012, 647348. [Google Scholar] [CrossRef]
- Saini, A.; Sharples, A.P.; Al-Shanti, N.; Stewart, C.E. Omega-3 Fatty Acid EPA Improves Regenerative Capacity of Mouse Skeletal Muscle Cells Exposed to Saturated Fat and Inflammation. Biogerontology 2017, 18, 109–129. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Nishida, Y.; Maeshige, N.; Moriguchi, M.; Uemura, M.; Ma, X.; Miyoshi, M.; Kondo, H.; Fujino, H. Preventive Effect of Docosahexaenoic Acid (DHA) and Eicosapentaenoic Acid (EPA) against Endotoxin-Induced Muscle Atrophy. Clin. Nutr. ESPEN 2021, 45, 503–506. [Google Scholar] [CrossRef]
- Chen, S.-C.; Chen, P.-Y.; Wu, Y.-L.; Chen, C.-W.; Chen, H.-W.; Lii, C.-K.; Sun, H.-L.; Liu, K.-L. Long-Chain Polyunsaturated Fatty Acids Amend Palmitate-Induced Inflammation and Insulin Resistance in Mouse C2C12 Myotubes. Food Funct. 2016, 7, 270–278. [Google Scholar] [CrossRef]
- Wei, H.-K.; Deng, Z.; Jiang, S.-Z.; Song, T.-X.; Zhou, Y.-F.; Peng, J.; Tao, Y.-X. Eicosapentaenoic Acid Abolishes Inhibition of Insulin-Induced MTOR Phosphorylation by LPS via PTP1B Downregulation in Skeletal Muscle. Mol. Cell. Endocrinol. 2017, 439, 116–125. [Google Scholar] [CrossRef]
- Jung, T.W.; Kim, H.-C.; Abd El-Aty, A.M.; Jeong, J.H. Protectin DX Ameliorates Palmitate- or High-Fat Diet-Induced Insulin Resistance and Inflammation through an AMPK-PPARα-Dependent Pathway in Mice. Sci. Rep. 2017, 7, 1397. [Google Scholar] [CrossRef]
- Dort, J.; Orfi, Z.; Fabre, P.; Molina, T.; Conte, T.C.; Greffard, K.; Pellerito, O.; Bilodeau, J.-F.; Dumont, N.A. Resolvin-D2 Targets Myogenic Cells and Improves Muscle Regeneration in Duchenne Muscular Dystrophy. Nat. Commun. 2021, 12, 6264. [Google Scholar] [CrossRef]
- Fogagnolo Mauricio, A.; Minatel, E.; Santo Neto, H.; Marques, M.J. Effects of Fish Oil Containing Eicosapentaenoic Acid and Docosahexaenoic Acid on Dystrophic Mdx Mice. Clin. Nutr. 2013, 32, 636–642. [Google Scholar] [CrossRef]
- Apolinário, L.M.; De Carvalho, S.C.; Santo Neto, H.; Marques, M.J. Long-Term Therapy with Omega-3 Ameliorates Myonecrosis and Benefits Skeletal Muscle Regeneration in Mdx Mice. Anat. Rec. 2015, 298, 1589–1596. [Google Scholar] [CrossRef]
- Carvalho, S.C.D.; Apolinário, L.M.; Matheus, S.M.M.; Santo Neto, H.; Marques, M.J. EPA Protects against Muscle Damage in the Mdx Mouse Model of Duchenne Muscular Dystrophy by Promoting a Shift from the M1 to M2 Macrophage Phenotype. J. Neuroimmunol. 2013, 264, 41–47. [Google Scholar] [CrossRef]
- de Carvalho, S.C.; Hindi, S.M.; Kumar, A.; Marques, M.J. Effects of Omega-3 on Matrix Metalloproteinase-9, Myoblast Transplantation and Satellite Cell Activation in Dystrophin-Deficient Muscle Fibers. Cell Tissue Res. 2017, 369, 591–602. [Google Scholar] [CrossRef]
- Pinel, A.; Rigaudière, J.P.; Jouve, C.; Montaurier, C.; Jousse, C.; LHomme, M.; Morio, B.; Capel, F. Transgenerational Supplementation with Eicosapentaenoic Acid Reduced the Metabolic Consequences on the Whole Body and Skeletal Muscle in Mice Receiving an Obesogenic Diet. Eur. J. Nutr. 2021, 60, 3143–3157. [Google Scholar] [CrossRef]
- Wang, Z.-G.; Zhu, Z.-Q.; He, Z.-Y.; Cheng, P.; Liang, S.; Chen, A.-M.; Yang, Q. Endogenous Conversion of N-6 to n-3 Polyunsaturated Fatty Acids Facilitates the Repair of Cardiotoxin-Induced Skeletal Muscle Injury in Fat-1 Mice. Aging 2021, 13, 8454–8466. [Google Scholar] [CrossRef]
- Chalchat, E.; Gaston, A.-F.; Charlot, K.; Peñailillo, L.; Valdés, O.; Tardo-Dino, P.-E.; Nosaka, K.; Martin, V.; Garcia-Vicencio, S.; Siracusa, J. Appropriateness of Indirect Markers of Muscle Damage Following Lower Limbs Eccentric-Biased Exercises: A Systematic Review with Meta-Analysis. PLoS ONE 2022, 17, e0271233. [Google Scholar] [CrossRef]
- Lieber, R.L.; Fridén, J. Mechanisms of Muscle Injury after Eccentric Contraction. J. Sci. Med. Sport 1999, 2, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, P.M. Exercise-Induced Muscle Damage--Animal and Human Models. Med. Sci. Sports Exerc. 1992, 24, 510–511. [Google Scholar] [CrossRef] [PubMed]
- Ochi, E.; Tsuchiya, Y.; Yanagimoto, K. Effect of Eicosapentaenoic Acids-Rich Fish Oil Supplementation on Motor Nerve Function after Eccentric Contractions. J. Int. Soc. Sports Nutr. 2017, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Lenn, J.; Uhl, T.; Mattacola, C.; Boissonneault, G.; Yates, J.; Ibrahim, W.; Bruckner, G. The Effects of Fish Oil and Isoflavones on Delayed Onset Muscle Soreness. Med. Sci. Sports Exerc. 2002, 34, 1605–1613. [Google Scholar] [CrossRef]
- Gray, P.; Chappell, A.; Jenkinson, A.M.E.; Thies, F.; Gray, S.R. Fish Oil Supplementation Reduces Markers of Oxidative Stress but Not Muscle Soreness after Eccentric Exercise. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 206–214. [Google Scholar] [CrossRef]
- Rajabi, A.; Lotfi, N.; Abdolmaleki, A.; Rashid-Amiri, S. The Effects of Omega-3 Intake on Delayed Onset Muscle Sorness in Non-Athlet Men. Pedagog. Psychol. Med. -Biol. Probl. Phys. Train. Sport. 2013, 17, 91–95. [Google Scholar]
- Tartibian, B.; Maleki, B.H.; Abbasi, A. The Effects of Ingestion of Omega-3 Fatty Acids on Perceived Pain and External Symptoms of Delayed Onset Muscle Soreness in Untrained Men. Clin. J. Sport Med. Off. J. Can. Acad. Sport Med. 2009, 19, 115–119. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Yanagimoto, K.; Ueda, H.; Ochi, E. Supplementation of Eicosapentaenoic Acid-Rich Fish Oil Attenuates Muscle Stiffness after Eccentric Contractions of Human Elbow Flexors. J. Int. Soc. Sports Nutr. 2019, 16, 19. [Google Scholar] [CrossRef]
- Philpott, J.D.; Donnelly, C.; Walshe, I.H.; MacKinley, E.E.; Dick, J.; Galloway, S.D.R.; Tipton, K.D.; Witard, O.C. Adding Fish Oil to Whey Protein, Leucine, and Carbohydrate over a Six-Week Supplementation Period Attenuates Muscle Soreness Following Eccentric Exercise in Competitive Soccer Players. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 26–36. [Google Scholar] [CrossRef]
- McKinley-Barnard, S.K.; Andre, T.L.; Gann, J.J.; Hwang, P.S.; Willoughby, D.S. Effectiveness of Fish Oil Supplementation in Attenuating Exercise-Induced Muscle Damage in Women during Midfollicular and Midluteal Menstrual Phases. J. Strength Cond. Res. 2018, 32, 1601–1612. [Google Scholar] [CrossRef]
- Lembke, P.; Capodice, J.; Hebert, K.; Swenson, T. Influence of Omega-3 (N3) Index on Performance and Wellbeing in Young Adults after Heavy Eccentric Exercise. J. Sports Sci. Med. 2014, 13, 151–156. [Google Scholar]
- Kyriakidou, Y.; Wood, C.; Ferrier, C.; Dolci, A.; Elliott, B. The Effect of Omega-3 Polyunsaturated Fatty Acid Supplementation on Exercise-Induced Muscle Damage. J. Int. Soc. Sports Nutr. 2021, 18, 9. [Google Scholar] [CrossRef]
- Xin, G.; Eshaghi, H. Effect of Omega-3 Fatty Acids Supplementation on Indirect Blood Markers of Exercise-Induced Muscle Damage: Systematic Review and Meta-Analysis of Randomized Controlled Trials. Food Sci. Nutr. 2021, 9, 6429–6442. [Google Scholar] [CrossRef]
- Lv, Z.T.; Zhang, J.M.; Zhu, W.T. Omega-3 Polyunsaturated Fatty Acid Supplementation for Reducing Muscle Soreness after Eccentric Exercise: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Biomed Res. Int. 2020, 2020, 8062017. [Google Scholar] [CrossRef]
- Bloomer, R.J.; Larson, D.E.; Fisher-Wellman, K.H.; Galpin, A.J.; Schilling, B.K. Effect of Eicosapentaenoic and Docosahexaenoic Acid on Resting and Exercise-Induced Inflammatory and Oxidative Stress Biomarkers: A Randomized, Placebo Controlled, Cross-over Study. Lipids Health Dis. 2009, 8, 36. [Google Scholar] [CrossRef]
- Cornish, S.M.; Johnson, S.T. Systemic Cytokine Response to Three Bouts of Eccentric Exercise. Results Immunol. 2014, 4, 23–29. [Google Scholar] [CrossRef]
- Rodríguez-Cruz, M.; Atilano-Miguel, S.; Barbosa-Cortés, L.; Bernabé-García, M.; Almeida-Becerril, T.; Cárdenas-Conejo, A.; Del Rocío Cruz-Guzmán, O.; Maldonado-Hernández, J. Evidence of Muscle Loss Delay and Improvement of Hyperinsulinemia and Insulin Resistance in Duchenne Muscular Dystrophy Supplemented with Omega-3 Fatty Acids: A Randomized Study. Clin. Nutr. 2019, 38, 2087–2097. [Google Scholar] [CrossRef]
- Rodríguez-Cruz, M.; Cruz-Guzmán, O.D.R.; Almeida-Becerril, T.; Solís-Serna, A.D.; Atilano-Miguel, S.; Sánchez-González, J.R.; Barbosa-Cortés, L.; Ruíz-Cruz, E.D.; Huicochea, J.C.; Cárdenas-Conejo, A.; et al. Potential Therapeutic Impact of Omega-3 Long Chain-Polyunsaturated Fatty Acids on Inflammation Markers in Duchenne Muscular Dystrophy: A Double-Blind, Controlled Randomized Trial. Clin. Nutr. 2018, 37, 1840–1851. [Google Scholar] [CrossRef]
- Sitzia, C.; Meregalli, M.; Belicchi, M.; Farini, A.; Arosio, M.; Bestetti, D.; Villa, C.; Valenti, L.; Brambilla, P.; Torrente, Y. Preliminary Evidences of Safety and Efficacy of Flavonoids- and Omega 3-Based Compound for Muscular Dystrophies Treatment: A Randomized Double-Blind Placebo Controlled Pilot Clinical Trial. Front. Neurol. 2019, 10, 755. [Google Scholar] [CrossRef]
- Henderson, G.C.; Evans, N.P.; Grange, R.W.; Tuazon, M.A. Compared with That of MUFA, a High Dietary Intake of n-3 PUFA Does Not Reduce the Degree of Pathology in Mdx Mice. Br. J. Nutr. 2014, 111, 1791–1800. [Google Scholar] [CrossRef] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jannas-Vela, S.; Espinosa, A.; Candia, A.A.; Flores-Opazo, M.; Peñailillo, L.; Valenzuela, R. The Role of Omega-3 Polyunsaturated Fatty Acids and Their Lipid Mediators on Skeletal Muscle Regeneration: A Narrative Review. Nutrients 2023, 15, 871. https://doi.org/10.3390/nu15040871
Jannas-Vela S, Espinosa A, Candia AA, Flores-Opazo M, Peñailillo L, Valenzuela R. The Role of Omega-3 Polyunsaturated Fatty Acids and Their Lipid Mediators on Skeletal Muscle Regeneration: A Narrative Review. Nutrients. 2023; 15(4):871. https://doi.org/10.3390/nu15040871
Chicago/Turabian StyleJannas-Vela, Sebastian, Alejandra Espinosa, Alejandro A. Candia, Marcelo Flores-Opazo, Luis Peñailillo, and Rodrigo Valenzuela. 2023. "The Role of Omega-3 Polyunsaturated Fatty Acids and Their Lipid Mediators on Skeletal Muscle Regeneration: A Narrative Review" Nutrients 15, no. 4: 871. https://doi.org/10.3390/nu15040871