
Inflammatory Response: A Crucial Way for Gut Microbes to Regulate Cardiovascular Diseases
 , , and
, , and 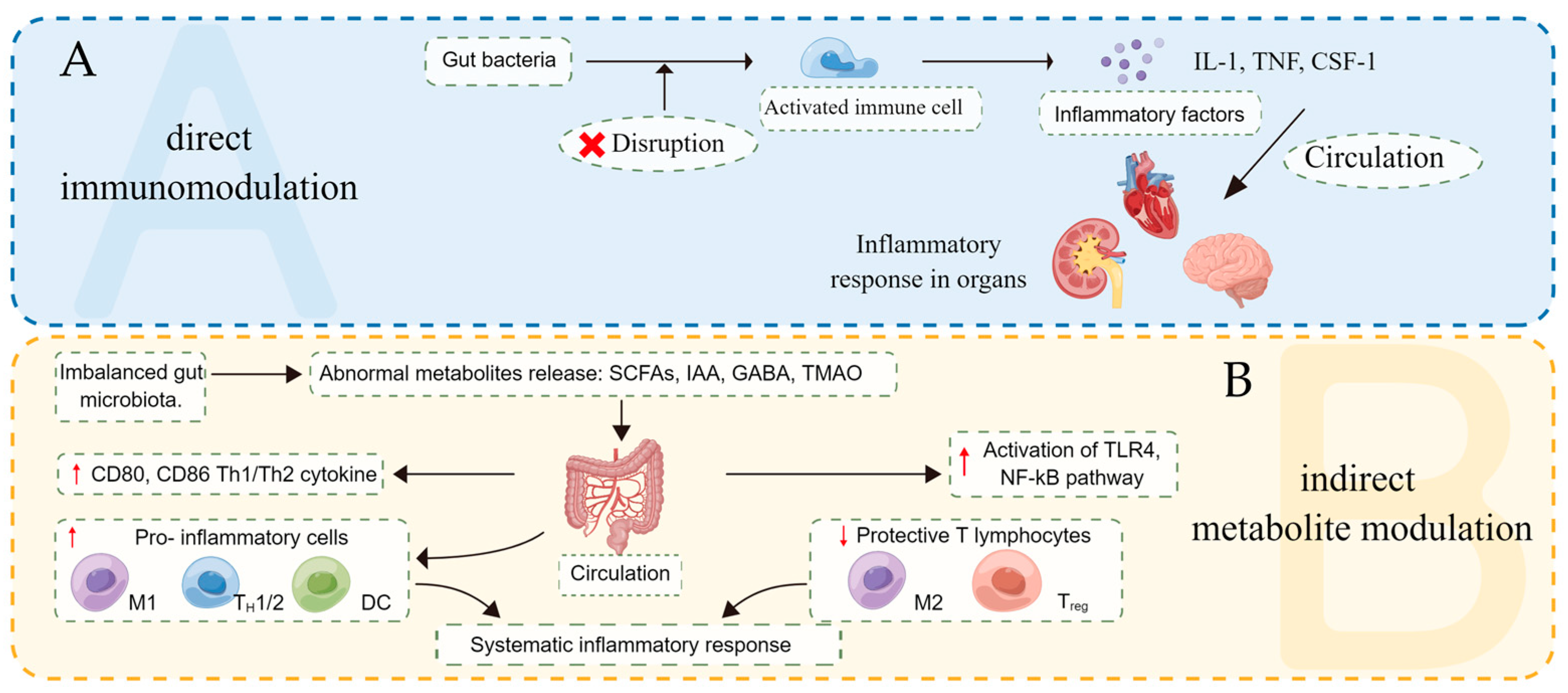{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Enterotype and Inflammatory Responses
2.1. Enterotype and Intestinal Microenvironment
2.2. Two Ways of Regulating Inflammatory Response
2.3. Disturbance of Gut Microbes Induces Inflammatory Response
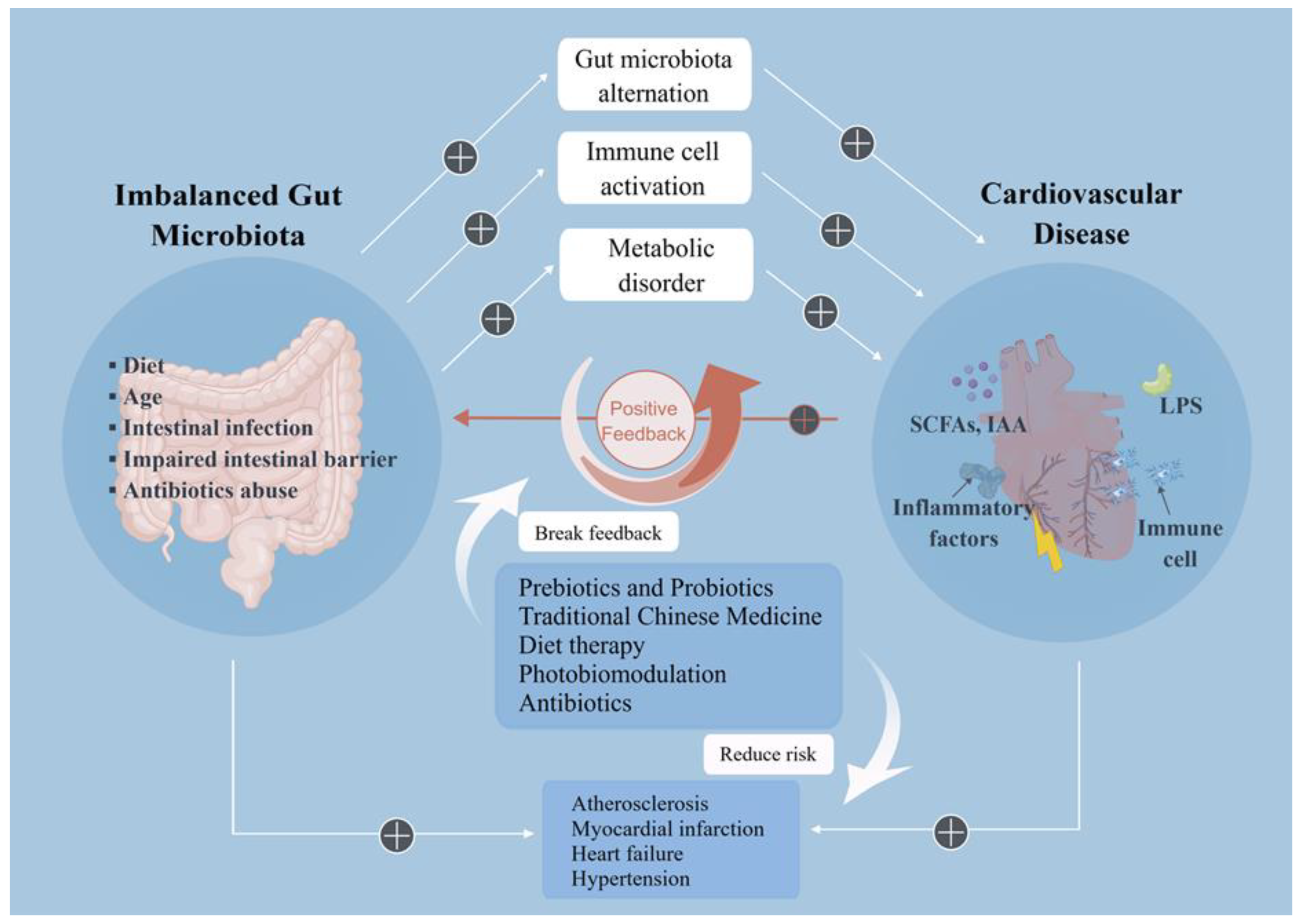
3. Imbalance of Gut Microbes and CVD
3.1. Atherosclerosis
3.2. Myocardial Infarction
3.3. Heart Failure
3.4. Hypertension
4. Therapeutic Strategies Target the Intestinal Microenvironment
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ridker, P.M.; Everett, B.M.; Thuren, T. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, F.K.; Dube, P.; Mohamed, A. Cardiotonic Steroids and the Sodium Trade Balance: New Insights into Trade-Off Mechanisms Mediated by the Na⁺/K⁺-ATPase. Int. J. Mol. Sci. 2018, 19, 2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díez-Sainz, E.; Milagro, F.I.; Riezu-Boj, J.I. Effects of gut microbiota-derived extracellular vesicles on obesity and diabetes and their potential modulation through diet. J. Physiol. Biochem. 2022, 78, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Riazi-Rad, F.; Behrouzi, A.; Mazaheri, H. Impact of gut microbiota on immune system. Acta Microbiol. Immunol. Hung. 2021, 68, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Jiang, L.; Shum, T. Lactobacillus rhamnosusElucidation of Anti-Hypertensive Mechanism by a Novel AC1 Fermented Soymilk in the Deoxycorticosterone Acetate-Salt Hypertensive Rats. Nutrients 2022, 14, 3174. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, Y.; Kong, Y. Microbiota-derived metabolite Indoles induced aryl hydrocarbon receptor activation and inhibited neuroinflammation in APP/PS1 mice. Brain Behav. Immun. 2022, 106, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Shen, M.; Chen, T. Mesona chinensis Benth Polysaccharides Alleviate DSS-Induced Ulcerative Colitis via Inhibiting of TLR4/MAPK/NF-κB Signaling Pathways and Modulating Intestinal Microbiota. Mol. Nutr. Food Res. 2022, 66, e2200047. [Google Scholar] [CrossRef]
- Chi, M.; Ma, K.; Wang, J. The Immunomodulatory Effect of the Gut Microbiota in Kidney Disease. J. Immunol. Res. 2021, 2021, 5516035. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cai, Z.; Ferrari, M.W. The Correlation between Gut Microbiota and Serum Metabolomic in Elderly Patients with Chronic Heart Failure. Mediators Inflamm. 2021, 2021, 5587428. [Google Scholar] [CrossRef]
- Alkushi, A.G.; Elazab, S.T.; Abdelfattah-Hassan, A. Multi-Strain-Probiotic-Loaded Nanoparticles Reduced Colon Inflammation and Orchestrated the Expressions of Tight Junction, NLRP3 Inflammasome and Caspase-1 Genes in DSS-Induced Colitis Model. Pharmaceutics 2022, 14, 1183. [Google Scholar] [CrossRef]
- Zhang, W.Q.; Quan, K.Y.; Feng, C.J. The Lactobacillus gasseri G098 Strain Mitigates Symptoms of DSS-Induced Inflammatory Bowel Disease in Mice. Nutrients 2022, 14, 3745. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Huang, H.; Zhou, M. Paeonol Attenuated Vascular Fibrosis Through Regulating Treg/Th17 Balance in a Gut Microbiota-Dependent Manner. Front. Pharmacol. 2021, 12, 765482. [Google Scholar] [CrossRef]
- Puspitasari, Y.; Ministrini, S.; Schwarz, L. Modern Concepts in Cardiovascular Disease: Inflamm-Aging. Front. Cell. Dev. Biol. 2022, 10, 882211. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Du, X.; Zuo, B. Qige Huxin Formula Attenuates Isoprenaline-Induced Cardiac Fibrosis in Mice via Modulating Gut Microbiota and Protecting Intestinal Integrity. Evid. Based Complement Alternat. Med. 2022, 2022, 2894659. [Google Scholar] [CrossRef]
- Zhao, H.; Lyu, Y.; Zhai, R. Metformin Mitigates Sepsis-Related Neuroinflammation Modulating Gut Microbiota and Metabolites. Front. Immunol. 2022, 13, 797312. [Google Scholar] [CrossRef]
- Wang, H.; He, C.; Liu, Y. Soluble dietary fiber protects intestinal mucosal barrier by improving intestinal flora in a murine model of sepsis. Biomed. Pharmacother. 2020, 129, 110343. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Chen, J.; Hoffmann, C. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Arumugam, M.; Raes, J.; Pelletier, E. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laursen, M. Gut Microbiota Development: Influence of Diet from Infancy to Toddlerhood. Ann. Nutr. Metab. 2021, 77, 21–34. [Google Scholar] [CrossRef]
- Zafar, H.; Saier, M. Bacteroides Gut species in health and disease. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Gorvitovskaia, A.; Holmes, S.; Huse, S.J.M. Interpreting Prevotella and Bacteroides as biomarkers of diet and lifestyle. Microbiome 2016, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Yuan, Y.; Zhang, S. Intestinal Flora and Disease Mutually Shape the Regional Immune System in the Intestinal Tract. Front. Immunol. 2020, 11, 575. [Google Scholar] [CrossRef] [Green Version]
- Kim, C. Control of lymphocyte functions by gut microbiota-derived short-chain fatty acids. Cell. Mol. Immunol. 2021, 18, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, D.; Evans, D.; Coyte, K. Understanding the development and function of the gut microbiota in health and inflammation. Frontline Gastroenterol. 2022, 13, e13–e21. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, E.C.; Murphy, C.; O’Neill, L.A. The role of TLRs, NLRs, and RLRs in mucosal innate immunity and homeostasis. Mucosal Immunol. 2010, 3, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steimle, A.; Frick, J.S. Molecular Mechanisms of Induction of Tolerant and Tolerogenic Intestinal Dendritic Cells in Mice. J. Immunol. Res. 2016, 2016, 1958650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, S.C.; Kamm, M.A.; Stagg, A.J. Intestinal dendritic cells: Their role in bacterial recognition, lymphocyte homing, and intestinal inflammation. Inflamm. Bowel Dis. 2010, 16, 1787–1807. [Google Scholar] [CrossRef]
- Wang, R.; Yao, L.; Meng, T. Rhodomyrtus tomentosa (Ait.) Hassk fruit phenolic-rich extract mitigates intestinal barrier dysfunction and inflammation in mice. Food Chem. 2022, 393, 133438. [Google Scholar] [CrossRef]
- Mao, J.; Li, Y.; Bian, Q. The Bufei Jianpi Formula Improves Mucosal Immune Function by Remodeling Gut Microbiota Through the SCFAs/GPR43/NLRP3 Pathway in Chronic Obstructive Pulmonary Disease Rats. Int. J. Chron. Obstruct. Pulmon. Dis. 2022, 17, 1285–1298. [Google Scholar] [CrossRef]
- Ilyas, A.; Wijayasinghe, Y.S.; Khan, I. Implications of trimethylamine N-oxide (TMAO) and Betaine in Human Health: Beyond Being Osmoprotective Compounds. Front. Mol. Biosci. 2022, 9, 964624. [Google Scholar] [CrossRef]
- Su, X.; Gao, Y.; Yang, R. Gut Microbiota-Derived Tryptophan Metabolites Maintain Gut and Systemic Homeostasis. Cells 2022, 11, 2296. [Google Scholar] [CrossRef]
- Fukuda, S.; Toh, H.; Hase, K. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef]
- Fukuda, S.; Toh, H.; Taylor, T.D. Acetate-producing bifidobacteria protect the host from enteropathogenic infection via carbohydrate transporters. Gut Microbes 2012, 3, 449–454. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, J.; He, T. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Sun, M.; Chen, F. Microbiota metabolite short-chain fatty acid acetate promotes intestinal IgA response to microbiota which is mediated by GPR43. Mucosal Immunol. 2017, 10, 946–956. [Google Scholar] [CrossRef] [Green Version]
- Arpaia, N.; Campbell, C.; Fan, X. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [Green Version]
- Li, H.B.; Xu, M.L.; Xu, X.D. Faecalibacterium prausnitzii Attenuates CKD via Butyrate-Renal GPR43 Axis. Circ. Res. 2022, 131, e120–e134. [Google Scholar] [CrossRef]
- Ren, Y.; Zhang, X.; He, F. Butyrate increases the monocytic myeloid-derived suppressor cells and promotes the secretion of anti-inflammatory cytokines in mice with alcoholic liver disease. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2022, 38, 865–871. [Google Scholar]
- Boyle, J.J. Macrophage activation in atherosclerosis: Pathogenesis and pharmacology of plaque rupture. Curr. Vasc. Pharmacol. 2005, 3, 63–68. [Google Scholar] [CrossRef]
- Kinne, R.W.; Bräuer, R.; Stuhlmüller, B. Macrophages in rheumatoid arthritis. Arthritis Res. 2000, 2, 189–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, M.A.; Jackson, J.; Stanton, M. Short-chain fatty acids act as antiinflammatory mediators by regulating prostaglandin E(2) and cytokines. World J. Gastroenterol. 2009, 15, 5549–5557. [Google Scholar] [CrossRef]
- Tedelind, S.; Westberg, F.; Kjerrulf, M. Anti-inflammatory properties of the short-chain fatty acids acetate and propionate: A study with relevance to inflammatory bowel disease. World J. Gastroenterol. 2007, 13, 2826–2832. [Google Scholar] [CrossRef]
- Sikalidis, A.K.; Maykish, A. The Gut Microbiome and Type 2 Diabetes Mellitus: Discussing a Complex Relationship. Biomedicines 2020, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Yin, J.; Zhang, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef] [PubMed]
- Graham, C.; Mullen, A.; Whelan, K. Obesity and the gastrointestinal microbiota: A review of associations and mechanisms. Nutr. Rev. 2015, 73, 376–385. [Google Scholar] [CrossRef]
- Gury-BenAri, M.; Thaiss, C.A.; Serafini, N. The Spectrum and Regulatory Landscape of Intestinal Innate Lymphoid Cells Are Shaped by the Microbiome. Cell 2016, 166, 1231–1246.e13. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, H.S.; Liu, Y.; Menkiti, O.R. The microbiota regulates neutrophil homeostasis and host resistance to Escherichia coli K1 sepsis in neonatal mice. Nat. Med. 2014, 20, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jiang, W.; Mellins, E.D. TCR-like antibodies targeting autoantigen-mhc complexes: A mini-review. Front. Immunol. 2022, 13, 968432. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, A.; Vermi, W.; Lee, J.S. Intraepithelial type 1 innate lymphoid cells are a unique subset of IL-12- and IL-15-responsive IFN-γ-producing cells. Immunity 2013, 38, 769–781. [Google Scholar] [CrossRef] [Green Version]
- Semin, I.; Ninnemann, J.; Bondareva, M. Interplay Between Microbiota, Toll-Like Receptors and Cytokines for the Maintenance of Epithelial Barrier Integrity. Front. Med. 2021, 8, 644333. [Google Scholar] [CrossRef]
- Kedmi, R.; Najar, T.A.; Mesa, K.R. A RORγt+ cell instructs gut microbiota-specific Treg cell differentiation. Nature 2022, 610, 737–743. [Google Scholar] [CrossRef]
- Ochoa-Repáraz, J.; Mielcarz, D.; Wang, Y. A polysaccharide from the human commensal Bacteroides fragilis protects against CNS demyelinating disease. Mucosal Immunol. 2010, 3, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Rocha, E.; Smith, C.J. Ferritin-like family proteins in the anaerobe Bacteroides fragilis: When an oxygen storm is coming, take your iron to the shelter. Biometals 2013, 26, 577–591. [Google Scholar] [CrossRef] [Green Version]
- Montesi, A.; García-Albiach, R.; Pozuelo, M. Molecular and microbiological analysis of caecal microbiota in rats fed with diets supplemented either with prebiotics or probiotics. Int. J. Food Microbiol. 2005, 98, 281–289. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, D.; Wang, J. Assessment of probiotic adhesion and inhibitory effect on Escherichia coli and Salmonella adhesion. Arch. Microbiol. 2021, 203, 6267–6274. [Google Scholar] [CrossRef]
- Chowdhury, R.; Pavinski Bitar, P.; Keresztes, I. A diffusible signal factor of the intestine dictates Salmonella invasion through its direct control of the virulence activator HilD. PLoS Pathog. 2021, 17, e1009357. [Google Scholar] [CrossRef]
- Lezutekong, J.; Nikhanj, A.; Oudit, G. Imbalance of gut microbiome and intestinal epithelial barrier dysfunction in cardiovascular disease. Clin. Sci. 2018, 132, 901–904. [Google Scholar] [CrossRef]
- Zhang, X.; Gérard, P. Diet-gut microbiota interactions on cardiovascular disease. Comput. Struct. Biotechnol. J. 2022, 20, 1528–1540. [Google Scholar] [CrossRef]
- Hu, Y.; Jin, X.; Gao, F. Selenium-enriched Bifidobacterium longum DD98 effectively ameliorates dextran sulfate sodium-induced ulcerative colitis in mice. Front. Microbiol. 2022, 13, 955112. [Google Scholar] [CrossRef]
- Jena, P.; Setayesh, T.; Sheng, L. Intestinal Microbiota Remodeling Protects Mice from Western Diet-Induced Brain Inflammation and Cognitive Decline. Cells 2022, 11, 504. [Google Scholar] [CrossRef]
- Mima, A.; Nagahara, D.; Tansho, K. Methotrexate induced pneumatosis intestinalis under hemodialysis patient. Hemodial Int. 2017, 21, E9–E12. [Google Scholar] [CrossRef]
- Ikeda, Y.; Ishii, S.; Yazaki, M. Portal congestion and intestinal edema in hospitalized patients with heart failure. Heart Vessels 2018, 33, 740–751. [Google Scholar] [CrossRef]
- Rossi, M.; Johnson, D.; Campbell, K.L. The Kidney-Gut Axis: Implications for Nutrition Care. J. Ren. Nutr. 2015, 25, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Chen, J.; Huang, Y. Yishen Qingli Heluo Granule Ameliorates Renal Dysfunction in 5/6 Nephrectomized Rats by Targeting Gut Microbiota and Intestinal Barrier Integrity. Front. Pharmacol. 2022, 13, 858881. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, D.; Deng, F. The Role of Vitamin D in Immune System and Inflammatory Bowel Disease. J. Inflamm. Res. 2022, 15, 3167–3185. [Google Scholar] [CrossRef]
- You, Y.; Song, H.; Wang, L. Supplement of Caulerpa lentillifera polysaccharide by pre-prandial gavage and free feeding demonstrates differences to prevent obesity and gut microbiota disturbance in mice. J. Sci. Food Agric. 2022. [Google Scholar] [CrossRef]
- Sun, Q.; Cheng, L.; Zeng, X. The modulatory effect of plant polysaccharides on gut flora and the implication for neurodegenerative diseases from the perspective of the microbiota-gut-brain axis. Int. J. Biol. Macromol. 2020, 164, 1484–1492. [Google Scholar] [CrossRef]
- Yan, N.; Wang, L.; Li, Y. Metformin intervention ameliorates AS in ApoE-/- mice through restoring gut dysbiosis and anti-inflammation. PLoS ONE 2021, 16, e0254321. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, Y.; Nabavi, S. Mechanisms of Oxidized LDL-Mediated Endothelial Dysfunction and Its Consequences for the Development of Atherosclerosis. Front. Cardiovasc. Med. 2022, 9, 925923. [Google Scholar] [CrossRef]
- Shao, B.; Xu, H.; Zhao, Y. NLRP3 Inflammasome in Atherosclerosis: Putting Out the Fire of Inflammation. Inflammation 2022. [Google Scholar] [CrossRef]
- Zhu, S.; Xu, K.; Jiang, Y. The gut microbiome in subclinical atherosclerosis: A population-based multiphenotype analysis. Rheumatology 2021, 61, 258–269. [Google Scholar] [CrossRef]
- Shi, G.; Lin, Y.; Wu, Y. Bacteroides fragilis Supplementation Deteriorated Metabolic Dysfunction, Inflammation, and Aorta Atherosclerosis by Inducing Gut Microbiota Dysbiosis in Animal Model. Nutrients 2022, 14, 2199. [Google Scholar] [CrossRef]
- Sun, Y.; Wu, D.; Zeng, W. The Role of Intestinal Dysbacteriosis Induced Arachidonic Acid Metabolism Disorder in Inflammaging in Atherosclerosis. Front. Cell. Infect. Microbiol. 2021, 11, 618265. [Google Scholar] [CrossRef]
- Deng, B.; Tao, L.; Wang, Y. Natural products against inflammation and atherosclerosis: Targeting on gut microbiota. Front. Microbiol. 2022, 13, 997056. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, R.; Mu, H. Oral Administration of Branched-Chain Amino Acids Attenuates Atherosclerosis by Inhibiting the Inflammatory Response and Regulating the Gut Microbiota in ApoE-Deficient Mice. Nutrients 2022, 14, 5065. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, L.; Chen, C. The gut microbiota-artery axis: A bridge between dietary lipids and atherosclerosis? Prog. Lipid Res. 2022, 89, 101209. [Google Scholar] [CrossRef]
- Xu, M.; Lv, C.; Wang, H. Peanut skin extract ameliorates high-fat diet-induced atherosclerosis by regulating lipid metabolism, inflammation reaction and gut microbiota in ApoE-/- mice. Food Res. Int. 2022, 154, 111014. [Google Scholar] [CrossRef]
- Qi, Y.; Liu, W.; Yan, X. Tongxinluo May Alleviate Inflammation and Improve the Stability of Atherosclerotic Plaques by Changing the Intestinal Flora. Front. Pharmacol. 2022, 13, 805266. [Google Scholar] [CrossRef]
- Dong, C.; Yang, Y.; Wang, Y. Gut microbiota combined with metabolites reveals unique features of acute myocardial infarction patients different from stable coronary artery disease. J. Adv. Res. 2022. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, H.; Xie, R. Gut microbiota aggravate cardiac ischemia-reperfusion injury via regulating the formation of neutrophils extracellular traps. Life Sci. 2022, 303, 120670. [Google Scholar] [CrossRef]
- Tang, T.; Chen, H.; Chen, C. Loss of Gut Microbiota Alters Immune System Composition and Cripples Postinfarction Cardiac Repair. Circulation 2019, 139, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Li, S.; Chen, H. The changes of gut microbiota after acute myocardial infarction in rats. PLoS ONE 2017, 12, e0180717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Ye, L.; Li, J. Metagenomic and metabolomic analyses unveil dysbiosis of gut microbiota in chronic heart failure patients. Sci. Rep. 2018, 8, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosheva, I.; Zheng, D.; Levy, M. High-Throughput Screen Identifies Host and Microbiota Regulators of Intestinal Barrier Function. Gastroenterology 2020, 159, 1807–1823. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.; von der Weid, P.Y. Lipopolysaccharides modulate intestinal epithelial permeability and inflammation in a species-specific manner. Gut Microbes 2020, 11, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Krüger, S.; Kunz, D.; Graf, J. Endotoxin hypersensitivity in chronic heart failure. Int. J. Cardiol. 2007, 115, 159–163. [Google Scholar] [CrossRef]
- Carrillo-Salinas, F.; Anastasiou, M.; Ngwenyama, N. Gut dysbiosis induced by cardiac pressure overload enhances adverse cardiac remodeling in a T cell-dependent manner. Gut Microbes 2020, 12, 1823801. [Google Scholar] [CrossRef]
- McMillan, A.; Hazen, S. Gut Microbiota Involvement in Ventricular Remodeling Post-Myocardial Infarction. Circulation 2019, 139, 660–662. [Google Scholar] [CrossRef]
- Organ, C.; Li, Z.; Sharp, T. Nonlethal Inhibition of Gut Microbial Trimethylamine N-oxide Production Improves Cardiac Function and Remodeling in a Murine Model of Heart Failure. J. Am. Heart Assoc. 2020, 9, e016223. [Google Scholar] [CrossRef]
- Lu, Y.; Xiang, M.; Xin, L. Qiliqiangxin Modulates the Gut Microbiota and NLRP3 Inflammasome to Protect Against Ventricular Remodeling in Heart Failure. Front. Pharmacol. 2022, 13, 905424. [Google Scholar] [CrossRef] [PubMed]
- Pullen, A.; Jadapalli, J.; Rhourri-Frih, B. Re-evaluating the causes and consequences of non-resolving inflammation in chronic cardiovascular disease. Heart Fail. Rev. 2020, 25, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Sarhene, M.; Wang, Y.; Wei, J. Biomarkers in heart failure: The past, current and future. Heart Fail. Rev. 2019, 24, 867–903. [Google Scholar] [CrossRef]
- Sandek, A.; Bauditz, J.; Swidsinski, A. Altered intestinal function in patients with chronic heart failure. J. Am. Coll. Cardiol. 2007, 50, 1561–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandek, A.; Anker, S.; von Haehling, S. The gut and intestinal bacteria in chronic heart failure. Curr. Drug Metab. 2009, 10, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Sandek, A.; Bjarnason, I.; Volk, H. Studies on bacterial endotoxin and intestinal absorption function in patients with chronic heart failure. Int. J. Cardiol. 2012, 157, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Feng, Z. The Role of Toll-Like Receptor Signaling in the Progression of Heart Failure. Mediators Inflamm. 2018, 2018, 9874109. [Google Scholar] [CrossRef] [Green Version]
- Jaworska, K.; Koper, M.; Ufnal, M. Gut microbiota and renin-angiotensin system: A complex interplay at local and systemic levels. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 321, G355–G366. [Google Scholar] [CrossRef]
- Saha, P.; Mell, B.; Golonka, R.M. Selective IgA Deficiency in Spontaneously Hypertensive Rats with Gut Dysbiosis. Hypertension 2022, 79, 2239–2249. [Google Scholar] [CrossRef]
- Hsu, C.N.; Yu, H.R.; Lin, I.C. Sodium butyrate modulates blood pressure and gut microbiota in maternal tryptophan-free diet-induced hypertension rat offspring. J. Nutr. Biochem. 2022, 108, 109090. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Nelson, J.W.; Phillips, S. Alterations of the gut microbial community structure and function with aging in the spontaneously hypertensive stroke prone rat. Sci. Rep. 2022, 12, 8534. [Google Scholar] [CrossRef] [PubMed]
- Drapkina, O.M.; Yafarova, A.A.; Kaburova, A.N. Targeting Gut Microbiota as a Novel Strategy for Prevention and Treatment of Hypertension, Atrial Fibrillation and Heart Failure: Current Knowledge and Future Perspectives. Biomedicines 2022, 10, 2019. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Wang, Y.; Fang, Z. Bifidobacterium longum CCFM752 prevented hypertension and aortic lesion, improved antioxidative ability, and regulated the gut microbiome in spontaneously hypertensive rats. Food Funct. 2022, 13, 6373–6386. [Google Scholar] [CrossRef]
- Toral, M.; Robles-Vera, I.; de la Visitación, N. Critical Role of the Interaction Gut Microbiota—Sympathetic Nervous System in the Regulation of Blood Pressure. Front. Physiol. 2019, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.K.; Yang, T.; Oliveira, A.C. Microglial Cells Impact Gut Microbiota and Gut Pathology in Angiotensin II-Induced Hypertension. Circ. Res. 2019, 124, 727–736. [Google Scholar] [CrossRef]
- Wang, T.; Gao, L.; Yang, Z. Restraint Stress in Hypertensive Rats Activates the Intestinal Macrophages and Reduces Intestinal Barrier Accompanied by Intestinal Flora Dysbiosis. J. Inflamm. Res. 2021, 14, 1085–1110. [Google Scholar] [CrossRef]
- Santisteban, M.; Qi, Y.; Zubcevic, J. Hypertension-Linked Pathophysiological Alterations in the Gut. Circ. Res. 2017, 120, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Marques, F.Z. How Dietary Fibre, Acting via the Gut Microbiome, Lowers Blood Pressure. Curr. Hypertens. Rep. 2022, 24, 509–521. [Google Scholar] [CrossRef]
- He, F.J.; Tan, M.; Ma, Y. Salt Reduction to Prevent Hypertension and Cardiovascular Disease: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2020, 75, 632–647. [Google Scholar] [CrossRef]
- Li, J.; Raizada, M.; Richards, E. Gut-brain-bone marrow axis in hypertension. Curr. Opin. Nephrol. Hypertens. 2021, 30, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Lange, T.; Luebber, F.; Grasshoff, H. The contribution of sleep to the neuroendocrine regulation of rhythms in human leukocyte traffic. Semin. Immunopathol. 2022, 44, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R. Catecholamine-Directed Epithelial Cell Interactions with Bacteria in the Intestinal Mucosa. Adv. Exp. Med. Biol. 2016, 874, 79–99. [Google Scholar] [PubMed]
- Foye, O.; Huang, I.; Chiou, C. Early administration of probiotic Lactobacillus acidophilus and/or prebiotic inulin attenuates pathogen-mediated intestinal inflammation and Smad 7 cell signaling. FEMS Immunol. Med. Microbiol. 2012, 65, 467–480. [Google Scholar] [CrossRef] [Green Version]
- Marques, F.; Nelson, E.; Chu, P. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef]
- Hsu, C.; Hou, C.; Hsu, W. Cardiovascular Diseases of Developmental Origins: Preventive Aspects of Gut Microbiota-Targeted Therapy. Nutrients 2021, 13, 2290. [Google Scholar] [CrossRef]
- Karlsson, C.; Ahrné, S.; Molin, G. Probiotic therapy to men with incipient arteriosclerosis initiates increased bacterial diversity in colon: A randomized controlled trial. Atherosclerosis 2010, 208, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, F.C.; Ferraresi, C. Photobiomodulation Therapy on the Treatment of Insulin Resistance: A Narrative Review. Photobiomodul. Photomed. Laser. Surg. 2022, 40, 597–603. [Google Scholar] [CrossRef]
- Cournos, F. The impact of environmental factors on outcome in residential programs. Hosp. Community. Psychiatry 1987, 38, 848–852. [Google Scholar] [CrossRef]
- Zhou, P.; Zhao, X.N.; Ma, Y.Y. Virtual screening analysis of natural flavonoids as trimethylamine (TMA)-lyase inhibitors for coronary heart disease. J. Food Biochem. 2022, 46, e14376. [Google Scholar] [CrossRef]
- Rahman, M.; Islam, F.; Or-Rashid, M.; Mamun, A.A.; Rahaman, M.; Islam, M.; Meem, A.F.K.; Sutradhar, P.R.; Mitra, S.; Mimi, A.A.; et al. The Gut Microbiota (Microbiome) in Cardiovascular Disease and Its Therapeutic Regulation. Front. Cell. Infect. Microbiol. 2022, 12, 903570. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Song, R.J.; Jiang, H. Oat fiber supplementation alleviates intestinal inflammation and ameliorates intestinal mucosal barrier via acting on gut microbiota-derived metabolites in LDLR-/- mice. Nutrition 2022, 95, 111558. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, R.; Yamasaki, Y.; Takeo, J. Long-chain monounsaturated fatty acids improve endothelial function with altering microbial flora. Transl. Res. 2021, 237, 16–30. [Google Scholar] [CrossRef]
- Sikalidis, A.K. Amino acids and immune response: A role for cysteine, glutamine, phenylalanine, tryptophan and arginine in T-cell function and cancer? Pathol. Oncol. Res. 2015, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Zhu, L.-J.; Leng, Y.-Q.; Wang, Y.-W.; Shi, T.; Wang, W.-Z.; Sun, J.-C. Inflammatory Response: A Crucial Way for Gut Microbes to Regulate Cardiovascular Diseases. Nutrients 2023, 15, 607. https://doi.org/10.3390/nu15030607
Wang W, Zhu L-J, Leng Y-Q, Wang Y-W, Shi T, Wang W-Z, Sun J-C. Inflammatory Response: A Crucial Way for Gut Microbes to Regulate Cardiovascular Diseases. Nutrients. 2023; 15(3):607. https://doi.org/10.3390/nu15030607
Chicago/Turabian StyleWang, Wen, Luo-Jiang Zhu, Yue-Qi Leng, Yu-Wan Wang, Te Shi, Wei-Zhong Wang, and Jia-Cen Sun. 2023. "Inflammatory Response: A Crucial Way for Gut Microbes to Regulate Cardiovascular Diseases" Nutrients 15, no. 3: 607. https://doi.org/10.3390/nu15030607