
Purification of Extracellular Protease from Staphylococcus simulans QB7and Its Ability in Generating Antioxidant and Anti-inflammatory Peptides from Meat Proteins


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Optimization of Culture Conditions for Protease Production
2.3. Protease Activity Assay
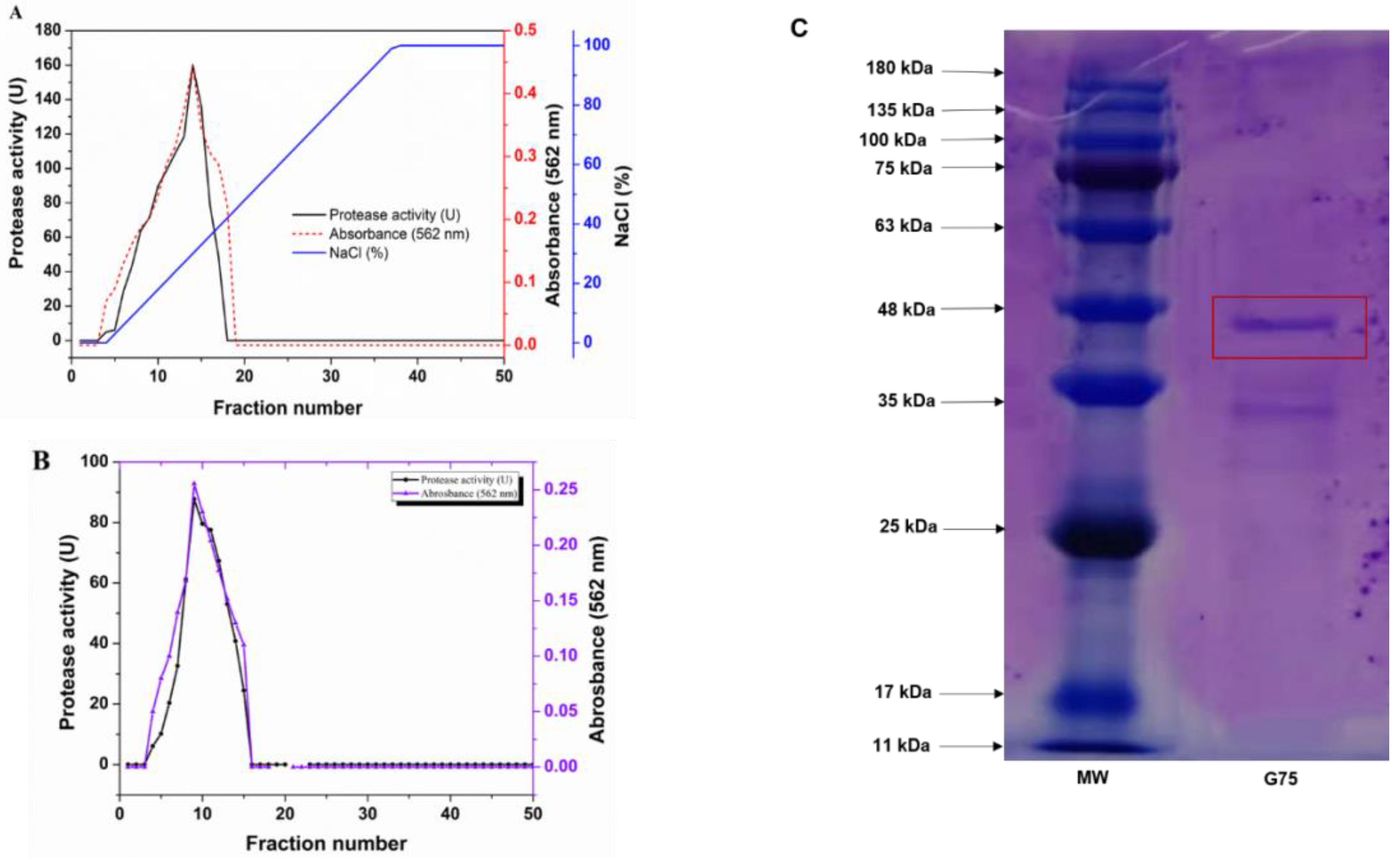
2.4. Purification of S. simulans QB7 Extracellular Protease
2.5. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.6. Enzymatic Properties
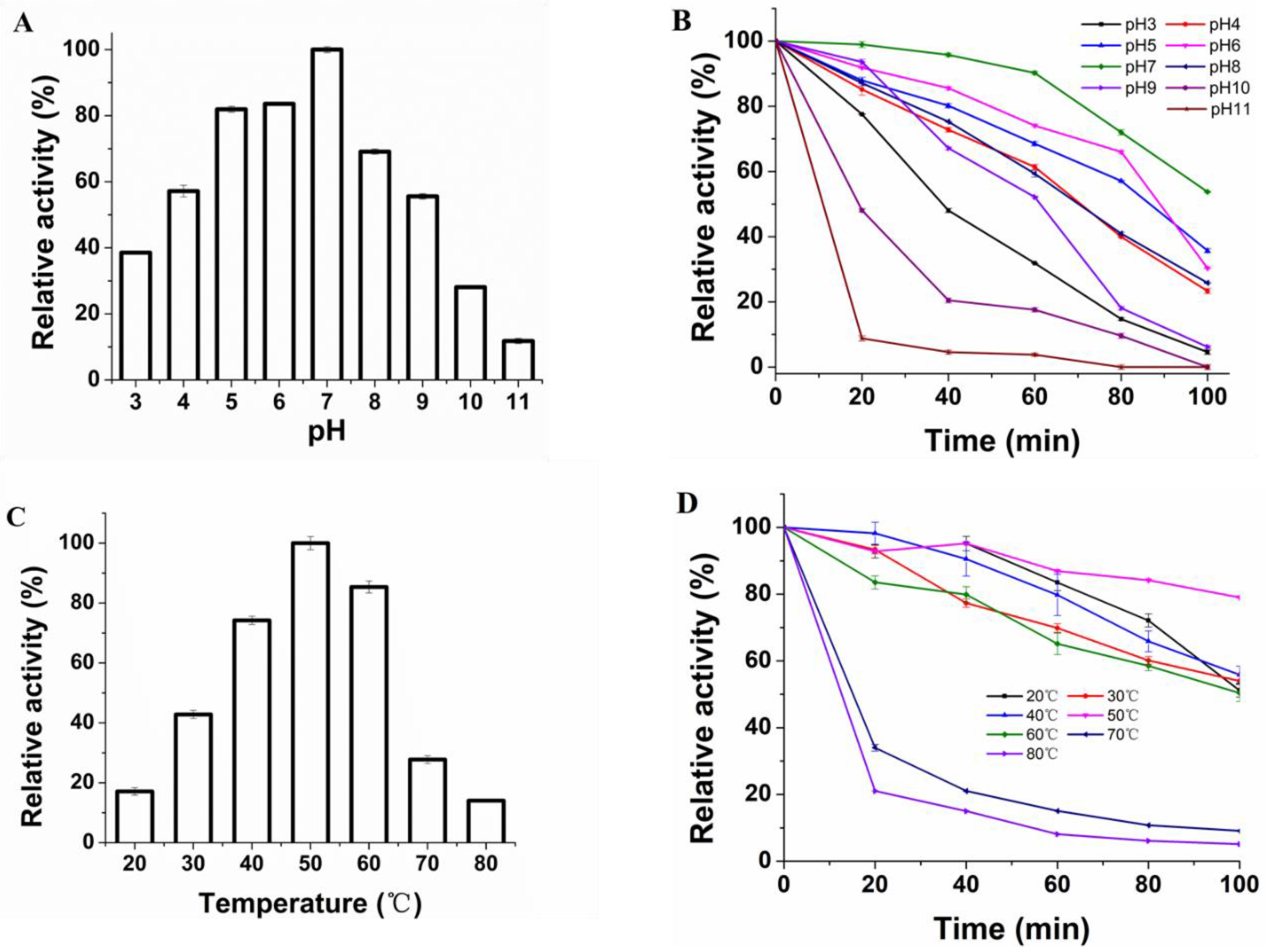
2.6.1. pH and Temperature
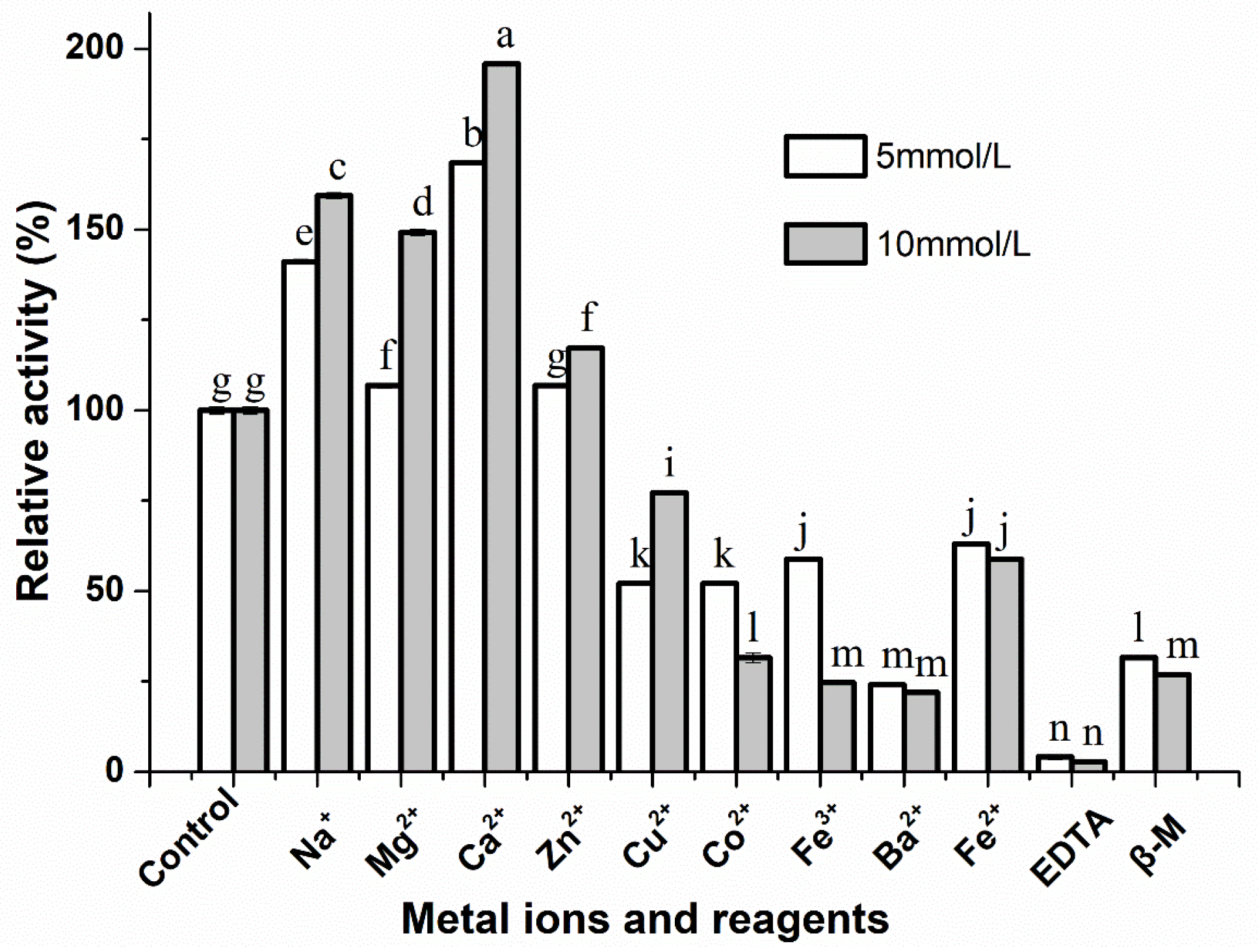
2.6.2. Metal Ions and Inhibitors
2.7. Degradation of Meat Proteins by S. simulans QB7 Extracellular Protease
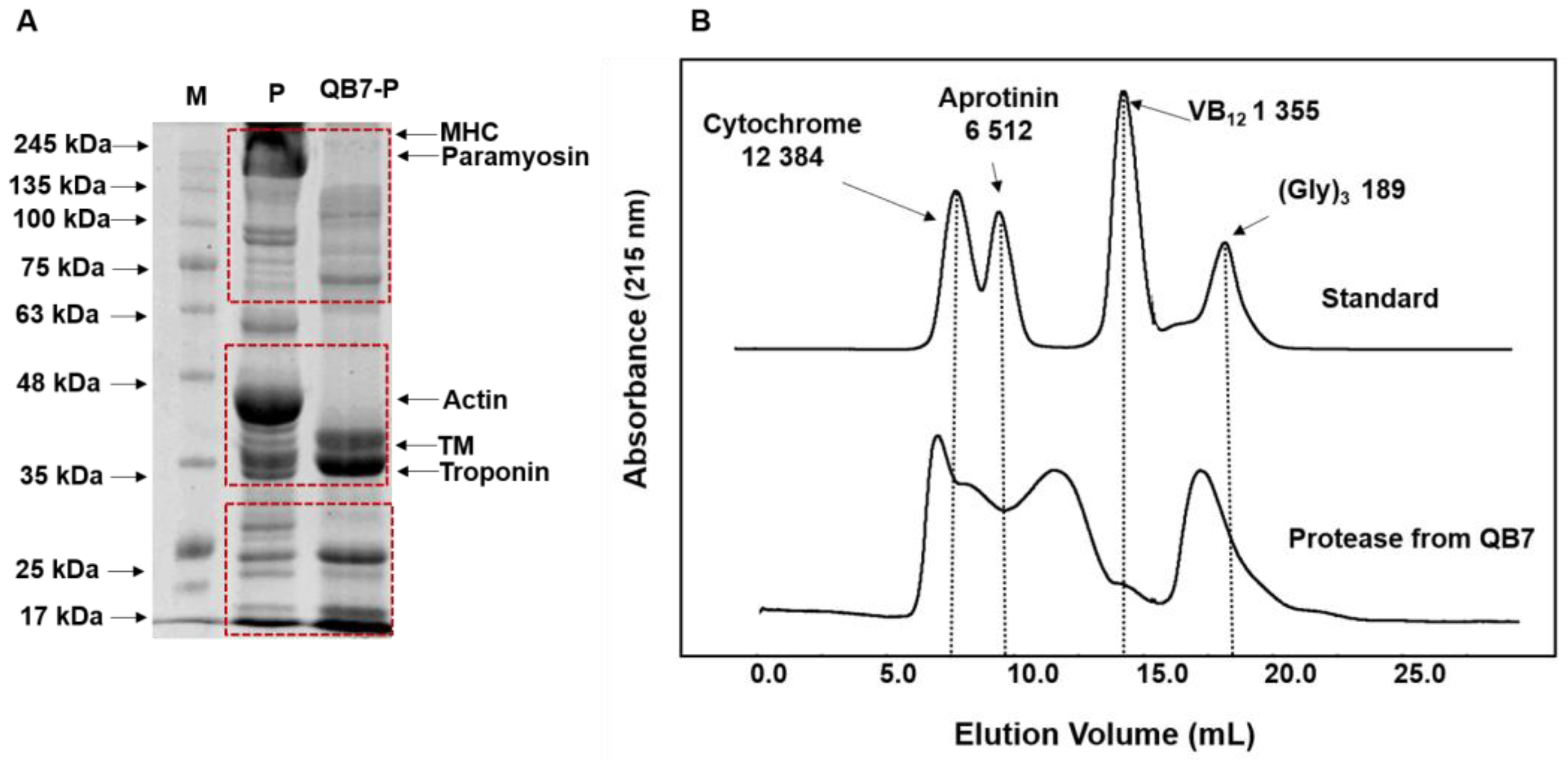
2.8. Size Exclusion Chromatography
2.9. Cytotoxicity, Oxidant Stress, and Anti-Inflammatory Evaluation through EA. Hy926 Cells
2.9.1. Cell Culture of EA.hy926 Cells
2.9.2. Desalting Protocol, Cytotoxicity, Lipid Peroxidation, and Superoxide Detection
2.9.3. Anti-Inflammatory Assay in EA.hy926 Cells
2.10. Statistical Analysis
3. Results and Discussion
3.1. Effect of Fermentation Factors on Crude Protease Production by S. simulans QB7
3.2. Protease Purification
3.3. Effect of pH and Temperature on Protease Activity and Stability
3.4. Effect of Metal Ions and Inhibitors on the Activity of S. simulans QB7 Protease
3.5. The Effect of S. simulans QB7 Protease on Protein Degradation and Generating Bioactive Peptides from Meat Proteins
3.5.1. Molecular Weight Distribution
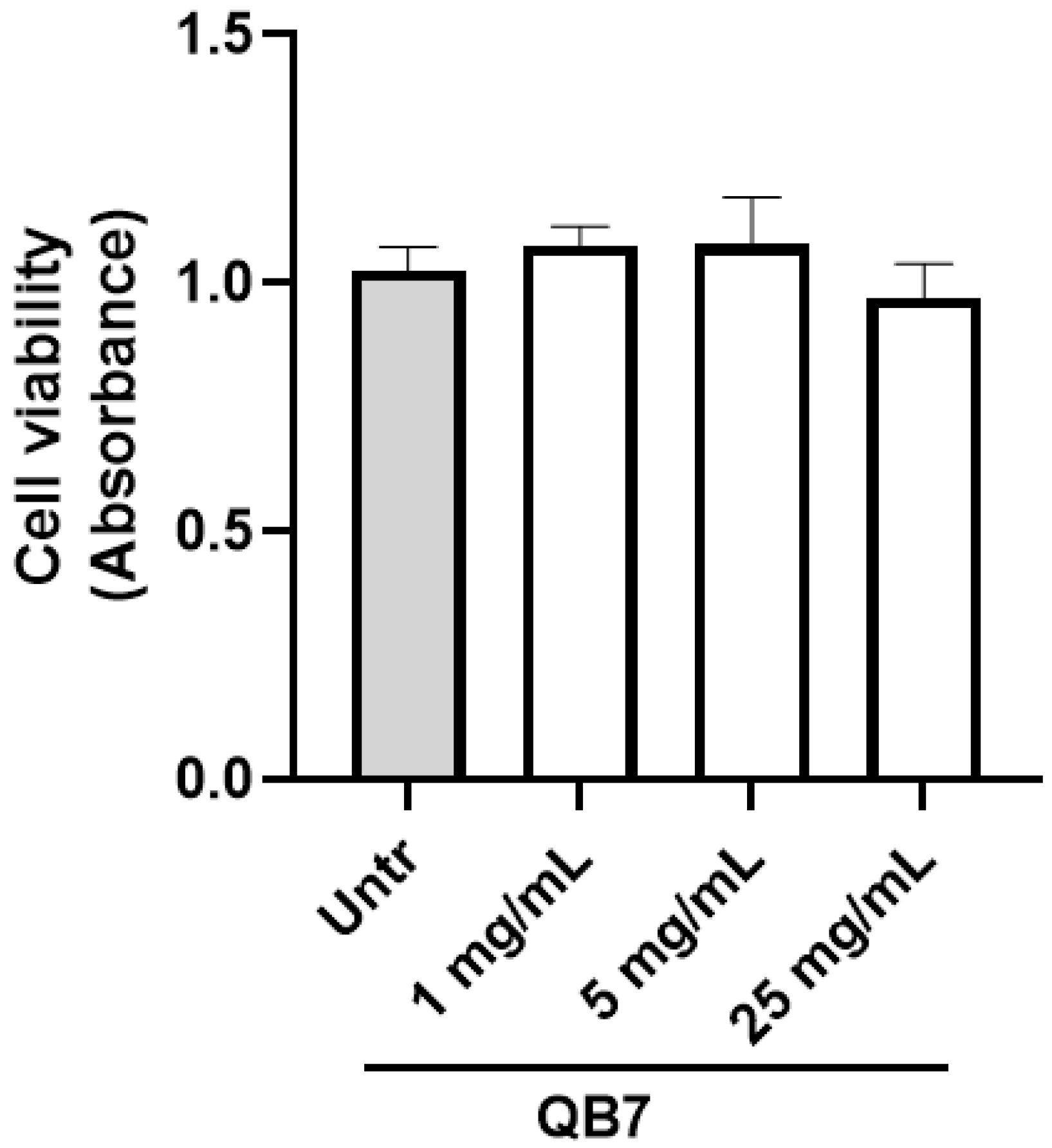
3.5.2. Cell Toxicity Assay
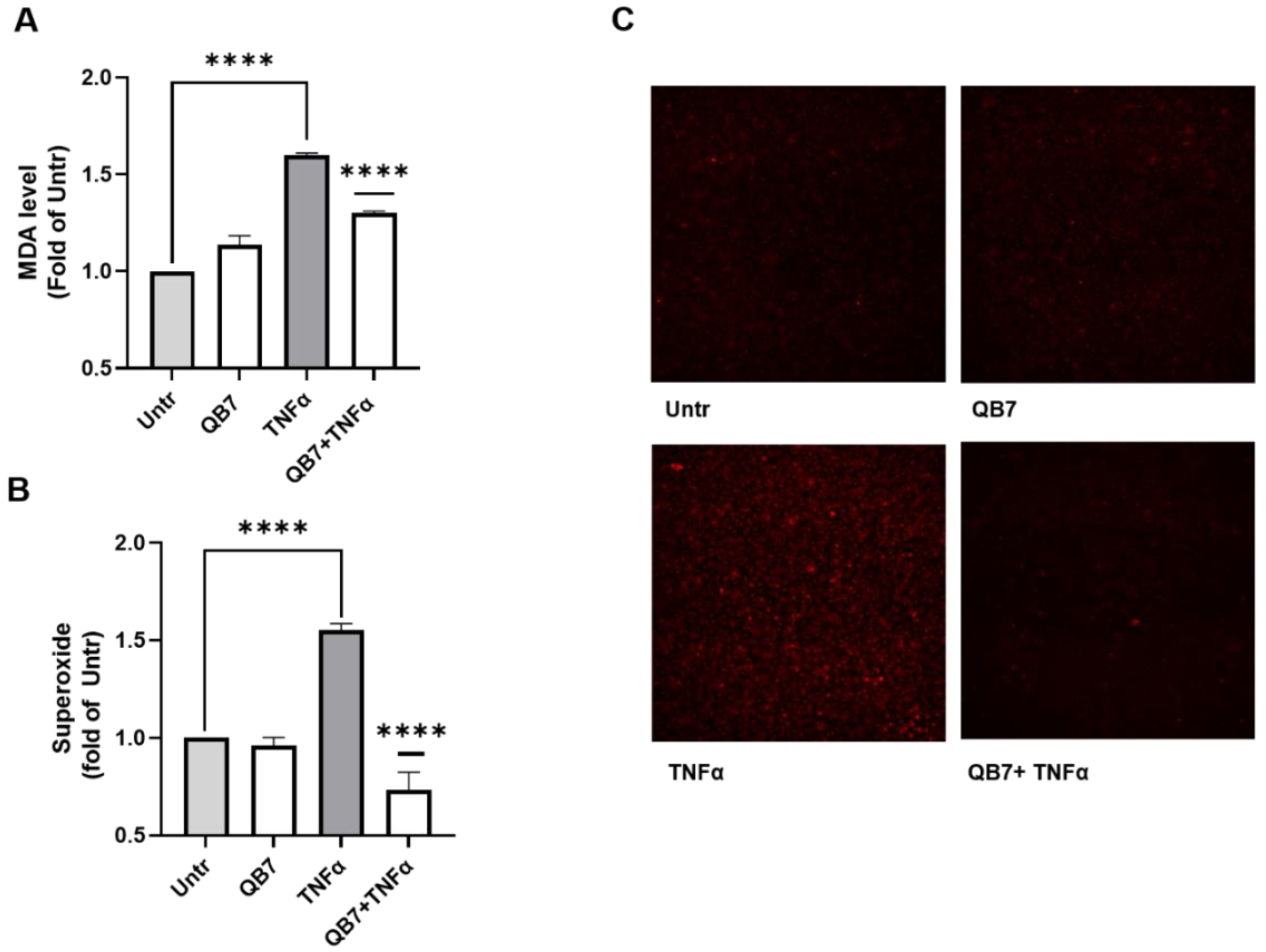
3.5.3. Antioxidant Activity
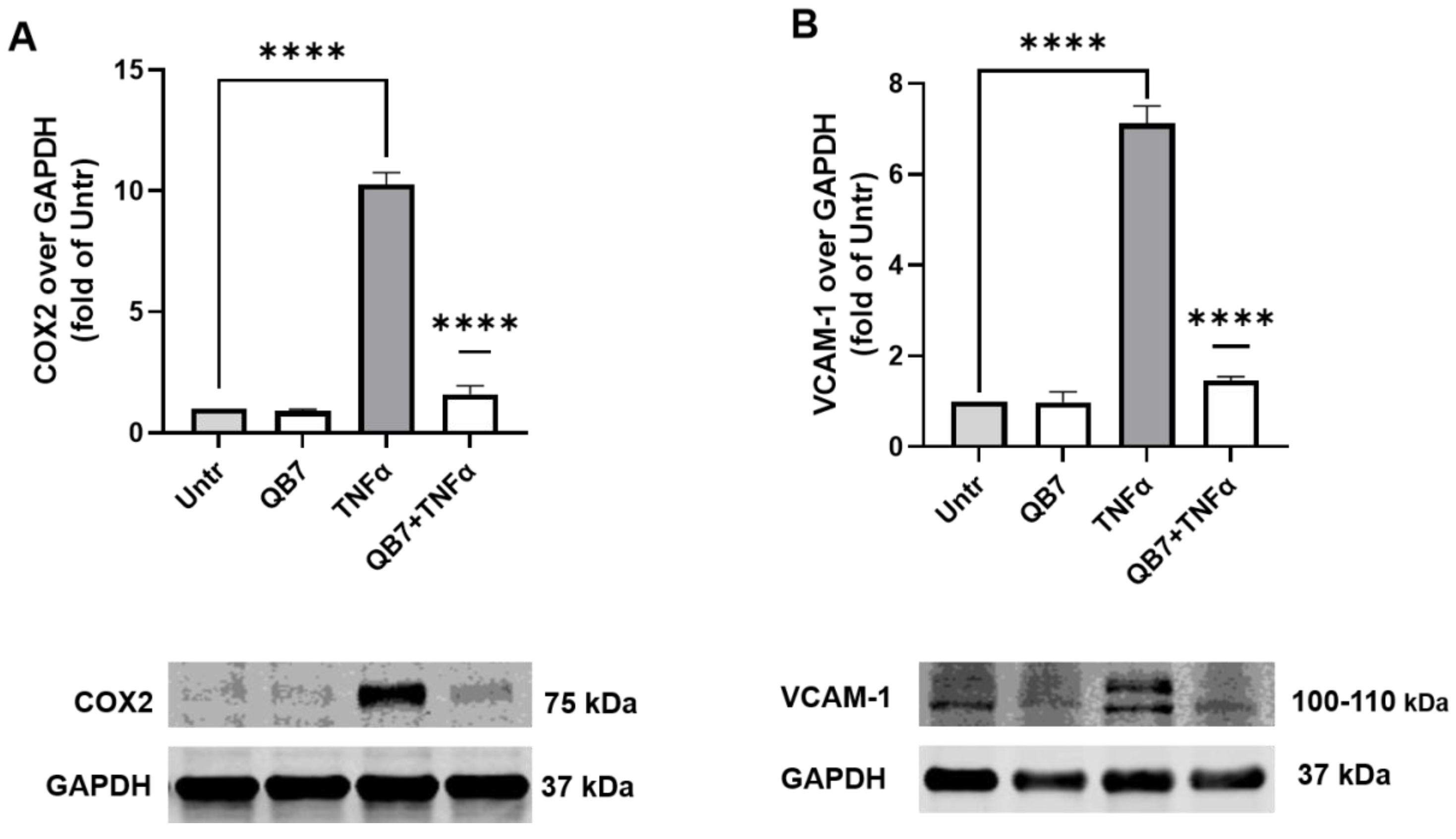
3.5.4. Anti-Inflammatory Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tavano, L.; Berenguer-Murcia, A.; Secundo, F. Biotechnological applications of proteases in food technology. Compr. Rev. Food Sci. Food Saf. 2018, 17, 412–436. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos Aguilar, H.; Sato, G. Microbial proteases: Production and application in obtaining protein hydrolysates. Food Res. Int. 2018, 103, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Huang, X.; Ren, Q.; Wang, X. Purification and characterization of a H2O2-tolerant alkaline protease from Bacillus sp. ZJ1502, a newly isolated strain from fermented bean curd. Food Chem. 2019, 274, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Nadeem, F.; Mehmood, T.; Muhammad Bilal, M.; Anwar, Z.; Fazeeha Amjad, F. Protease—A Versatile and Ecofriendly Biocatalyst with Multi-Industrial Applications: An Updated Review. Catal. Lett. 2021, 151, 307–323. [Google Scholar] [CrossRef]
- Ahangari, H.; Yazdani, P.; Ebrahimi, V.; Soofiyani, R.; Azargun, R.; Tarhriz, V.; Eyvazi, S. An Updated review on production of food derived bioactive peptides; focus on the psychrotrophic bacterial proteases. Biocatal. Agric. Biotechnol. 2021, 35, 102051. [Google Scholar] [CrossRef]
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial proteases applications. Front. Bioeng. Biotechnol. 2019, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammad Badrud Duza, M. Microbial enzymes and their applications-a review. Indo Am. J. Pharm. Res. 2013, 3, 6208–6219. [Google Scholar]
- Ullah, N.; Rehman, M.U.; Sarwar, A.; Nadeem, M.; Nelofer, R.; Shakir, H.A.; Irfan, M.; Idrees, M.; Naz, S.; Nabi, G.; et al. Purification, Characterization, and Application of Alkaline Protease Enzyme from a Locally Isolated Bacillus cereus Strain. Fermentation 2022, 8, 628. [Google Scholar] [CrossRef]
- Salihi, A.; Asoodeh, A.; Aliabadian, M. Production and biochemical characterization of an alkaline protease from Aspergillus oryzae CH93. Int. J. Biol. Macromol. 2017, 94, 827–835. [Google Scholar] [CrossRef]
- Toldrá, F.; Gallego, M.; Reig, M.; Aristoy, M.; Mora, L. Recent Progress in Enzymatic Release of Peptides in Foods of Animal Origin and Assessment of Bioactivity. J. Agric. Food Chem. 2020, 68, 12842–12855. [Google Scholar] [CrossRef]
- Candogan, K.; Wardlaw, B.; Acton, C. Effect of starter culture on proteolytic changes during processing of fermented beef sausages. Food Chem. 2009, 116, 731–737. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Q.; Xia, X.; Sun, F.; Kong, B. Biochemical properties of extracellular protease from Staphylococcus carnosus RT6 isolated from Harbin dry sausages, and its hydrolysis of meat proteins. J. Food Sci. 2021, 86, 1642–1655. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, J.; Kong, B.; Liu, Q.; Xia, X.; Sun, F. Purification and Characterization of the Protease from Staphylococcus xylosus A2 Isolated from Harbin Dry Sausages. Foods 2022, 11, 1094. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Feng, M.; Sun, J. Effects of Lactobacillus plantarum CD101 and Staphylococcus simulans NJ201 on proteolytic changes and bioactivities (antioxidant and antihypertensive activities) in fermented pork sausage. LWT-Food Sci. Technol. 2020, 133, 109985. [Google Scholar] [CrossRef]
- Yu, D.; Feng, M.; Sun, J.; Xu, X.; Zhou, G. Protein degradation and peptide formation with antioxidant activity in pork protein extracts inoculated with Lactobacillus plantarum and Staphylococcus simulans. Meat Sci. 2020, 160, 107958. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Feng, M.; Sun, J. Influence of mixed starters on the degradation of proteins and the formation of peptides with antioxidant activities in dry fermented sausages. Food Control 2021, 123, 107743. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Q.; Chen, X.; Zhou, J.; Wu, J. Isolation and characterization of coagulase negative staphylococci with high proteolytic activity from dry fermented sausages as a potential starter culture. Food Res. Int. 2022, 162, 111957. [Google Scholar] [CrossRef]
- Farhadian, S.; Asoodeh, A.; Lagzian, M. Purification, biochemical characterization, and structural modeling of a potential htrA-like serine protease from Bacillus subtilis DR8806. J. Mol. Catal. B: Enzym. 2015, 115, 51–58. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, Z.; Zhang, J. An acidic protease from the grass carp intestine (Ctenopharyngodon idellus). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 149, 83–90. [Google Scholar] [CrossRef]
- Afsharnezhad, M.; Shahangian, S.; Sariri, R. A novel milk-clotting cysteine protease from Ficus johannis: Purification and characterization. Int. J. Biol. Macromol. 2019, 121, 173–182. [Google Scholar] [CrossRef]
- Laemmli, K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, J.; Chen, Q.; Kong, B.; Sun, F. Biochemical properties of extracellular protease from Staphylococcus epidermidis isolated from Harbin dry sausages and its hydrolysis of meat protein. Food Biosci. 2021, 42, 101130. [Google Scholar] [CrossRef]
- Wang, C.; Wu, J. Preparation and characterization of adhesive from spent hen proteins. Int. J. Adhes. Adhes. 2012, 36, 8–14. [Google Scholar] [CrossRef]
- Fan, H.; Yu, W.; Liao, W.; Wu, J. Spent Hen Protein Hydrolysate with Good Gastrointestinal Stability and Permeability in Caco-2 Cells Shows Antihypertensive Activity in SHR. Foods 2020, 9, 1384. [Google Scholar] [CrossRef]
- Fan, H.; Bhullar, S.; Wang, Z.; Wu, J. Chicken Muscle Protein-Derived Peptide VVHPKESF Reduces TNFα-Induced Inflammation and Oxidative Stress by Suppressing TNFR1 Signaling in Human Vascular Endothelial Cells. Mol. Nutr. Food Res. 2022, 66, 2200184. [Google Scholar] [CrossRef]
- Fan, H.; Wang, J.; Liao, W.; Jiang, X.; Wu, J. Identification and Characterization of Gastrointestinal-Resistant Angiotensin-Converting Enzyme Inhibitory Peptides from Egg White Proteins. J. Agric. Food Chem. 2019, 67, 7147–7156. [Google Scholar] [CrossRef]
- Fan, H.; Bhullar, K.; Wu, J. Spent Hen Muscle Protein-Derived RAS Regulating Peptides Show Antioxidant Activity in Vascular Cells. Antioxidants 2021, 10, 290. [Google Scholar] [CrossRef]
- Singh, S.; Tripathi, V.; Jain, R.; Vikram, S.; Garg, S. An antibiotic, heavy metal resistant and halotolerant Bacillus cereus SIU1 and its thermoalkaline protease. Microb. Cell Factories 2010, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Sun, Q.; Zhang, H.; Kong, B.; Xia, X. Purification and biochemical characteristics of the microbial extracellular protease from Lactobacillus curvatus isolated from Harbin dry sausages. Int. J. Biol. Macromol. 2019, 133, 987–997. [Google Scholar] [CrossRef]
- Sharma, K.; Kumar, R.; Kumar, A. Microbial alkaline proteases: Optimization of production parameters and their properties. J. Genet. Eng. Biotechnol. 2017, 15, 115–126. [Google Scholar] [CrossRef]
- Gupta, A.; Khare, S. Enhanced production and characterization of a solvent stable protease from solvent tolerant Pseudomonas aeruginosa PseA. Enzym. Microb. Technol. 2007, 42, 11–16. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, H.; Mats, L.; Liu, R.; Deng, Z.; Mine, Y.; Tsao, R. Anti-inflammatory Effect and Cellular Uptake Mechanism of Peptides from Common Bean (Phaseolus vulga L.) Milk and Yogurts in Caco-2 Mono- and Caco-2/EA.hy926 Co-culture Models. J. Agric. Food Chem. 2019, 67, 8370–8381. [Google Scholar] [CrossRef] [PubMed]
- Worsztynowicz, P.; Bialas, W.; Grajek, W. Integrated approach for obtaining bioactive peptides from whey proteins hydrolysed using a new proteolytic lactic acid bacteria. Food Chem. 2019, 312, 126035. [Google Scholar] [CrossRef] [PubMed]
- Bisswanger, H. Enzyme assays. Perspect. Sci. 2020, 1, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Lücke, F. Fermented Meat Products: An Overview. Fermented Meat Prod. Health Asp. 2016, 2, 1–15. [Google Scholar]
- Abdel-Naby, M.; Ahmed, S.; Wehaidy, H.; El-Mandy, S. Catalytic, kinetic and thermodynamic properties of stabilized Bacillus stearothermophilus alkaline protease. Int. J. Biol. Macromol. 2017, 96, 265–271. [Google Scholar] [CrossRef]
- Jaouadi, N.; Rekik, H.; Ben Elhoul, M.; Rahem, F.; Hila, C.; Ben Aicha, C.; Badis, A.; Toumi, A.; Bejar, S.; Jaouadi, B. A novel keratinase from Bacillus tequilensis strain Q7 with promising potential for the leather bating process. Int. J. Biol. Macromol. 2015, 79, 952–964. [Google Scholar] [CrossRef]
- Broncano, J.; Timon, M.; Parra, V.; Andres, A.; Petron, M. Use of proteases to improve oxidative stability of fermented sausages by increasing low molecular weight compounds with antioxidant activity. Food Res. Int. 2011, 44, 2655–2659. [Google Scholar] [CrossRef]
- Fan, H.; Liao, W.; Wu, J. Chicken muscle hydrolysate reduces blood pressure in spontaneously hypertensive rats, upregulates ACE2, and ameliorates vascular inflammation, fibrosis, and oxidative stress. J. Food Sci. 2022, 87, 1292–1305. [Google Scholar] [CrossRef]
- Liang, Q.; Chalamaiah, M.; Ren, X.; Ma, H.; Wu, J. Identification of New Anti-inflammatory Peptides from Zein Hydrolysate after Simulated Gastrointestinal Digestion and Transport in Caco-2 Cells. J. Agric. Food Chem. 2018, 66, 1114–1120. [Google Scholar] [CrossRef]
- Baranska, P.; Jerczynska, H.; Pawlowska, Z.; Koziolkiewicz, W.; Cierniewski, S. Expression of Integrins and Adhesive Properties of Human Endothelial Cell Line EA.hy 926. Cancer Genom. Proteom. 2005, 2, 265–269. [Google Scholar]
- Touyz, M.; Schiffrin, L. Signal transduction mechanisms mediating the physiological and pathophysiological actions of angiotensin II in vascular smooth muscle cells. Pharmacol. Rev. 2000, 52, 639–672. [Google Scholar] [PubMed]
- Touyz, M.; Schiffrin, L. Reactive oxygen species in vascular biology: Implications in hypertension. Histochem. Cell Biol. 2004, 122, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Galle, J.; Quaschning, T.; Seibold, S.; Wanner, C. Endothelial dysfunction and inflammation: What is the link? Kidney Int. 2003, 63, S45–S49. [Google Scholar] [CrossRef] [Green Version]
- Odegaard, O.; Jacobs, R.; Sanchez, A.; Goff, C.; Reiner, P.; Gross, D. Oxidative stress, inflammation, endothelial dysfunction and incidence of type 2 diabetes. Cardiovasc. Diabetol. 2016, 15, 51. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Touyz, M.; Park, B.; Schiffrin, L. Antioxidant Effects of Vitamins C and E Are Associated with Altered Activation of Vascular NADPH Oxidase and Superoxide Dismutase in Stroke-Prone SHR. Hypertension (Dallas, Tex. 1979) 2001, 38, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Fleenor, S.; Seals, R.; Zigler, L.; Sindler, L. Superoxide-lowering therapy with TEMPOL reverses arterial dysfunction with aging in mice. Aging Cell 2012, 11, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Gallego, M.; Mora, M.; Hayes, M.; Reig, M.; Toldra, F. Peptides with Potential Cardioprotective Effects Derived from Dry-Cured Ham Byproducts. J. Agric. Food Chem. 2019, 67, 1115–1126. [Google Scholar] [CrossRef]
- Jahandideh, F.; Chakrabarti, S.; Davidge, S.; Wu, J. Antioxidant Peptides Identified from Ovotransferrin by the ORAC Method Did Not Show Anti-Inflammatory and Antioxidant Activities in Endothelial Cells. J. Agric. Food Chem. 2016, 64, 113–119. [Google Scholar] [CrossRef]
- Yamawaki, H.; Kuramoto, J.; Kameshima, S.; Usui, T.; Okada, M.; Hara, Y. Omentin, a novel adipocytokine inhibits TNF-induced vascular inflammation in human endothelial cells. Biochem. Biophys. Res. 2011, 408, 339–343. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Steps | Total Activity (U) | Total Protein (mg) | Enzymatic Activity (U/mg Protein) | Purificatio Fold | Yield (%) |
|---|---|---|---|---|---|
| Culture medium supernatant | 107.90 ± 1.22 | 49.38 ± 0.021 | 2.19 ± 0.025 | 1 | 100 |
| Ammonium sulfate precipitation | 58.60 ± 1.23 | 15.10 ± 0.011 | 3.88 ± 0.049 | 1.77 | 54.31 |
| DEAE-Sepharose FF | 34.78 ± 1.61 | 1.44 ± 0.012 | 24.15 ± 0.41 | 15.88 | 32.23 |
| Sephadex-G75 chromatography | 15.92 ± 1.32 | 0.24 ± 0.077 | 66.33 ± 1.63 | 30.29 | 14.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Fan, H.; Lu, K.; Zhu, Q.; Wu, J. Purification of Extracellular Protease from Staphylococcus simulans QB7and Its Ability in Generating Antioxidant and Anti-inflammatory Peptides from Meat Proteins. Nutrients 2023, 15, 65. https://doi.org/10.3390/nu15010065
Li H, Fan H, Lu K, Zhu Q, Wu J. Purification of Extracellular Protease from Staphylococcus simulans QB7and Its Ability in Generating Antioxidant and Anti-inflammatory Peptides from Meat Proteins. Nutrients. 2023; 15(1):65. https://doi.org/10.3390/nu15010065
Chicago/Turabian StyleLi, Hongying, Hongbing Fan, Kuan Lu, Qiujin Zhu, and Jianping Wu. 2023. "Purification of Extracellular Protease from Staphylococcus simulans QB7and Its Ability in Generating Antioxidant and Anti-inflammatory Peptides from Meat Proteins" Nutrients 15, no. 1: 65. https://doi.org/10.3390/nu15010065