
Faba Bean: An Untapped Source of Quality Plant Proteins and Bioactives


Abstract
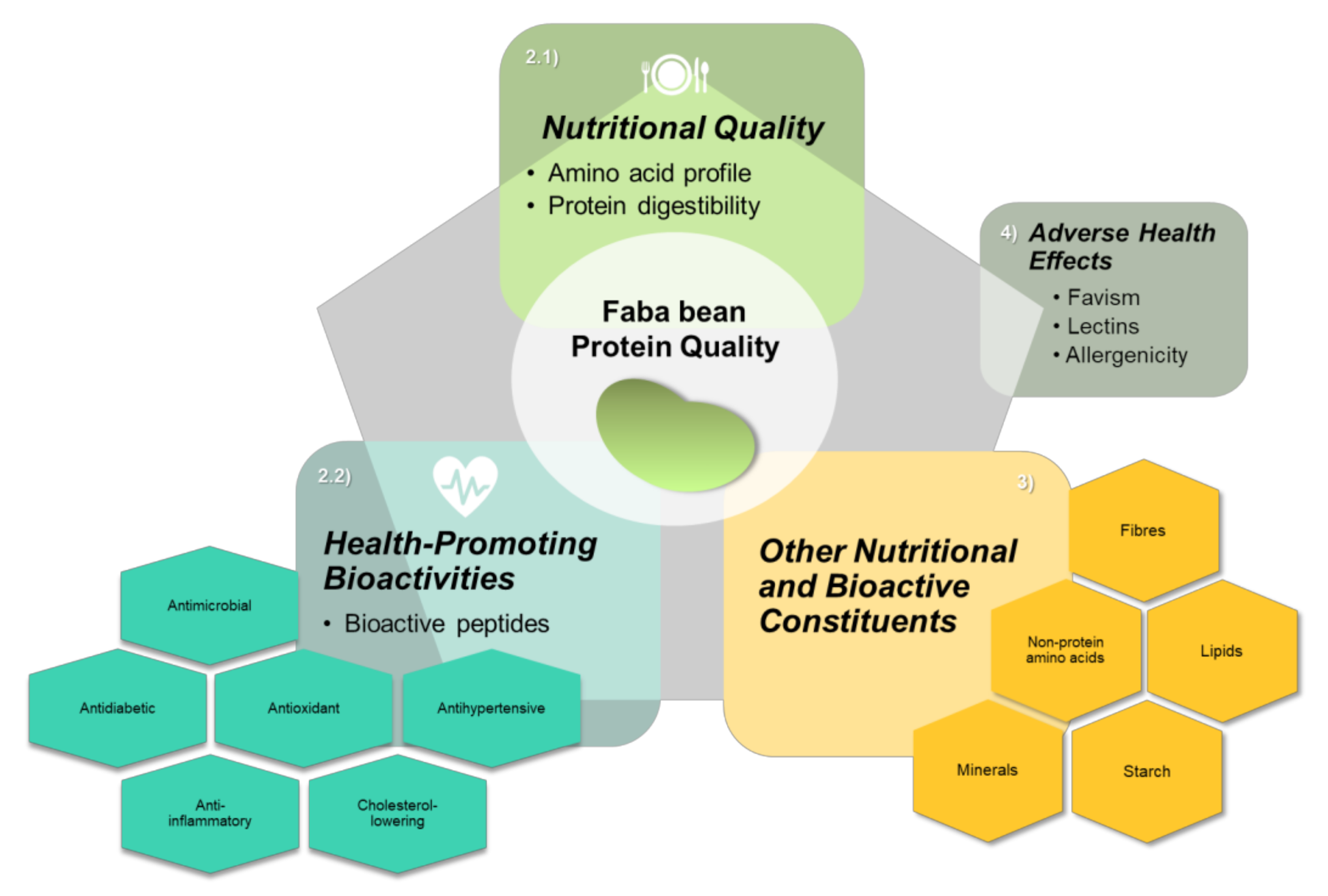
:1. Introduction
2. Faba Bean Proteins
2.1. Nutritional Quality
2.1.1. Assessment of Protein Nutritional Quality
2.1.2. Amino Acid Profile
2.1.3. Protein Digestibility
2.1.4. Effect of Processing on Protein Digestibility and Quality
Thermal Treatments
Biochemical Processes
2.1.5. Effects of Anti-Nutritional Content on Protein Digestibility and Quality
Tannins
Phytic Acid
Trypsin Inhibitor
2.2. Faba Bean Health-Promoting Bioactive Properties
2.2.1. Faba Bean Gastrointestinal Hydrolysates with Potential Beneficial Health-Related Bioactivities
2.2.2. Anti-Microbial Bioactivities of Faba Bean Enzymatic Hydrolysates
3. Other Nutritional and Bioactive Constituents of Faba Beans
3.1. Starch
3.2. Fibres
3.3. Lipids
3.4. Minerals
3.5. Non-Protein Amino Acids
4. Faba Bean Adverse Health Effects
4.1. Favism
4.2. Presence of Lectins
4.3. Allergenicity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Tilman, D.; Clark, M. Global diets link environmental sustainability and human health. Nature 2014, 515, 518. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Beydoun, M.A. Meat consumption is associated with obesity and central obesity among US adults. Int. J. Obes. 2009, 33, 621–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Franco, O.H.; Lamballais, S.; Ikram, M.A.; Schoufour, J.D.; Muka, T.; Voortman, T. Associations of specific dietary protein with longitudinal insulin resistance, prediabetes and type 2 diabetes: The Rotterdam Study. Clin. Nutr. 2020, 39, 242–249. [Google Scholar] [CrossRef]
- Abete, I.; Romaguera, D.; Vieira, A.R.; Lopez de Munain, A.; Norat, T. Association between total, processed, red and white meat consumption and all-cause, CVD and IHD mortality: A meta-analysis of cohort studies. Br. J. Nutr. 2014, 112, 762–775. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. Consumption of Red Meat and Processed Meat; IARC Publication: Lyon, France, 2015; Volume 114. [Google Scholar]
- Sinha, R.; Cross, A.J.; Graubard, B.I.; Leitzmann, M.F.; Schatzkin, A. Meat intake and mortality: A prospective study of over half a million people. Arch. Intern. Med. 2009, 169, 562–571. [Google Scholar] [CrossRef]
- Erbersdobler, H.F.; Barth, C.A.; Jahreis, G. Legumes in human nutrition: Nutrient content and protein quality of pulses. Ernaehrungs Umsch. 2017, 64, 134–139. [Google Scholar] [CrossRef]
- Technavio. Global Plant Protein Market 2017–2021. Available online: https://www.technavio.com/report/global-plant-protein-market?utm_source=t4&utm_medium=bw&utm_campaign=businesswire (accessed on 30 March 2020).
- Witcombe, J.R. Genetic Resources of Faba Beans. In Faba Bean Improvement, Proceedings of the Faba Bean Conference, Cairo, Egypt, 7–11 March 1981; Hawtin, G., Webb, C., Eds.; Springer: Dordrecht, The Netherlands, 1982; pp. 1–13. [Google Scholar]
- FAOSTAT. FAOSTAT Database. Available online: http://www.fao.org/faostat/en/#home (accessed on 29 November 2019).
- Lizarazo, C.I.; Lampi, A.; Jingwei, L.; Sontag-Strohm, T.; Piironen, V.; Stoddard, F.L. Nutritive quality and protein production from grain legumes in a boreal climate. J. Sci. Food Agric. 2015, 95, 2053–2064. [Google Scholar] [CrossRef] [Green Version]
- Oomah, B.D.; Luc, G.; Leprelle, C.; Drover, J.C.G.; Harrison, J.E.; Olson, M. Phenolics, Phytic Acid, and Phytase in Canadian-Grown Low-Tannin Faba Bean (Vicia faba L.) Genotypes. J. Agric. Food Chem. 2011, 59, 3763–3771. [Google Scholar] [CrossRef]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Köpke, U.; Nemecek, T. Ecological services of faba bean. Field Crops Res. 2010, 115, 217–233. [Google Scholar] [CrossRef]
- Aschi, A.; Aubert, M.; Riah-Anglet, W.; Nélieu, S.; Dubois, C.; Akpa-Vinceslas, M.; Trinsoutrot-Gattin, I. Introduction of Faba bean in crop rotation: Impacts on soil chemical and biological characteristics. Appl. Soil Ecol. 2017, 120, 219–228. [Google Scholar] [CrossRef]
- Dubova, L.; Alsiņa, I.; Ruža, A.; Šenberga, A. Impact of faba bean (Vicia faba, L.) cultivation on soil microbiological activity. Agron. Res. 2018, 16, 2016–2025. [Google Scholar] [CrossRef]
- Mouradi, M.; Farissi, M.; Makoudi, B.; Bouizgaren, A.; Ghoulam, C. Effect of faba bean (Vicia faba L.)–rhizobia symbiosis on barley’s growth, phosphorus uptake and acid phosphatase activity in the intercropping system. Ann. Agrar. Sci. 2018, 16, 297–303. [Google Scholar] [CrossRef]
- Xu, Y.; Qiu, W.; Sun, J.; Müller, C.; Lei, B. Effects of wheat/faba bean intercropping on soil nitrogen transformation processes. J. Soils Sediments 2018, 19, 1724–1734. [Google Scholar] [CrossRef]
- Xiao, J.; Yin, X.; Ren, J.; Zhang, M.; Tang, L.; Zheng, Y. Complementation drives higher growth rate and yield of wheat and saves nitrogen fertilizer in wheat and faba bean intercropping. Field Crops Res. 2018, 221, 119–129. [Google Scholar] [CrossRef]
- Zhang, C.; Dong, Y.; Tang, L.; Zheng, Y.; Makowski, D.; Yu, Y.; Zhang, F.; van der Werf, W. Intercropping cereals with faba bean reduces plant disease incidence regardless of fertilizer input; a meta-analysis. Eur. J. Plant Pathol. 2019, 154, 931–942. [Google Scholar] [CrossRef]
- Calabrò, S.; Cutrignelli, M.I.; Gonzalez, O.J.; Chiofalo, B.; Grossi, M.; Tudisco, R.; Panetta, C.; Infascelli, F. Meat quality of buffalo young bulls fed faba bean as protein source. Meat Sci. 2014, 96, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Health Canada. Common Food Allergens. Available online: https://www.canada.ca/en/health-canada/services/food-nutrition/food-safety/food-allergies-intolerances/food-allergies.html (accessed on 30 March 2020).
- Wunderlich, S.; Gatto, K.A. Consumer perception of genetically modified organisms and sources of information. Adv. Nutr. 2015, 6, 842–851. [Google Scholar] [CrossRef] [Green Version]
- Raikos, V.; Neacsu, M.; Russell, W.; Duthie, G. Comparative study of the functional properties of lupin, green pea, fava bean, hemp, and buckwheat flours as affected by pH. Food Sci. Nutr. 2014, 2, 802–810. [Google Scholar] [CrossRef] [Green Version]
- Estefania, C.M.C.; Ivan, V.T.J.; Maria, C.T.L.; Rosa-Millan, J.D.L.; Othona, S.S.S. Physicochemical characteristics, ATR-FTIR molecular interactions and in vitro starch and protein digestion of thermally-treated whole pulse flours. Food Res. Int. 2018, 105, 371–383. [Google Scholar] [CrossRef]
- Mattila, P.; Pihlava, J.-M.; Hellström, J.; Nurmi, M.; Eurola, M.; Mäkinen, S.; Jalava, T.; Pihlanto, A. Contents of phytochemicals and antinutritional factors in commercial protein-rich plant products. Food Qual. Saf. 2018, 2, 213–219. [Google Scholar] [CrossRef]
- Shi, L.; Mu, K.; Arntfield, S.D.; Nickerson, M.T. Changes in levels of enzyme inhibitors during soaking and cooking for pulses available in Canada. J. Food Sci. Technol. 2017, 54, 1014–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Arntfield, S.D.; Nickerson, M. Changes in levels of phytic acid, lectins and oxalates during soaking and cooking of Canadian pulses. Food Res. Int. 2018, 107, 660–668. [Google Scholar] [CrossRef]
- Chiremba, C.; Vandenberg, A.; Smits, J.; Samaranayaka, A.; Lam, R.; Hood-Niefer, S. New opportunities for faba bean. Cereal Foods World 2018, 63, 221–222. [Google Scholar] [CrossRef] [Green Version]
- Belsey, M.A. The epidemiology of favism. Bull. World Health Organ. 1973, 48, 1–13. [Google Scholar]
- Khazaei, H.; Purves, R.W.; Hughes, J.; Link, W.; O’Sullivan, D.M.; Schulman, A.H.; Björnsdotter, E.; Geu-Flores, F.; Nadzieja, M.; Andersen, S.U.; et al. Eliminating vicine and convicine, the main anti-nutritional factors restricting faba bean usage. Trends Food Sci. Technol. 2019, 91, 549–556. [Google Scholar] [CrossRef]
- Hou, W.; Zhang, X.; Yan, Q.; Li, P.; Sha, W.; Tian, Y.; Liu, Y. Linkage map of a gene controlling zero tannins (zt-1) in faba bean (Vicia faba L.) with SSR and ISSR markers. Agronomy 2018, 8, 80. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, D.; Pasqualone, A.; Costantini, M.; Ricciardi, L.; Lotti, C.; Pavan, S.; Summo, C. Data on the proximate composition, bioactive compounds, physicochemical and functional properties of a collection of faba beans (Vicia faba L.) and lentils (Lens culinaris Medik.). Data Brief 2021, 34, 106660. [Google Scholar] [CrossRef]
- Kumar, S.R.; Sadiq, M.B.; Anal, A.K. Comparative study of physicochemical and functional properties of soaked, germinated and pressure cooked Faba bean. J. Food Sci. Technol. 2022, 59, 257–267. [Google Scholar] [CrossRef]
- Setia, R.; Dai, Z.; Nickerson, M.T.; Sopiwnyk, E.; Malcolmson, L.; Ai, Y. Impacts of short-term germination on the chemical compositions, technological characteristics and nutritional quality of yellow pea and faba bean flours. Food Res. Int. 2019, 122, 263–272. [Google Scholar] [CrossRef]
- Bhatty, R.S. Chemical composition of some faba bean cultivars. Can. J. Plant Sci. 1974, 54, 413–421. [Google Scholar] [CrossRef]
- Mattila, P.; Mäkinen, S.; Eurola, M.; Jalava, T.; Pihlava, J.M.; Hellström, J.; Pihlanto, A. Nutritional Value of Commercial Protein-Rich Plant Products. Plant Foods Hum. Nutr. 2018, 73, 108–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamidou, S.; Nengas, I.; Grigorakis, K.; Nikolopoulou, D.; Jauncey, K. Chemical Composition and Antinutritional Factors of Field Peas (Pisum sativum), Chickpeas (Cicer arietinum), and Faba Beans (Vicia faba) as Affected by Extrusion Preconditioning and Drying Temperatures. Cereal Chem. 2011, 88, 80–86. [Google Scholar] [CrossRef]
- Mayer Labba, I.C.; Frøkiær, H.; Sandberg, A.S. Nutritional and antinutritional composition of fava bean (Vicia faba L., var. minor) cultivars. Food Res. Int. 2021, 140, 110038. [Google Scholar] [CrossRef] [PubMed]
- Ivarsson, E.; Neil, M. Variations in nutritional and antinutritional contents among faba bean cultivars and effects on growth performance of weaner pigs. Livest. Sci. 2018, 212, 14–21. [Google Scholar] [CrossRef]
- Hood-Niefer, S.D.; Warkentin, T.D.; Chibbar, R.N.; Vandenberg, A.; Tyler, R.T. Effect of genotype and environment on the concentrations of starch and protein in, and the physicochemical properties of starch from, field pea and fababean. J. Sci. Food. Agric. 2012, 92, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Moussou, N.N.N. Nutrients and non-nutrients composition and in vitro starch digestibility of five Algerian legume seed flours. Int. Food Res. J. 2019, 26, 1339–1349. [Google Scholar]
- Grela, E.R.; Kiczorowska, B.; Samolińska, W.; Matras, J.; Kiczorowski, P.; Rybiński, W.; Hanczakowska, E. Chemical composition of leguminous seeds: Part I—content of basic nutrients, amino acids, phytochemical compounds, and antioxidant activity. Eur. Food Res. Technol. 2017, 243, 1385–1395. [Google Scholar] [CrossRef]
- Abdel-Aal, E.-S.M.; Ragaee, S.; Rabalski, I.; Warkentin, T.; Vandenberg, A. Nutrient content and viscosity of Saskatchewan-grown pulses in relation to their cooking quality. Can. J. Plant Sci. 2018, 99, 67–77. [Google Scholar] [CrossRef]
- Njoumi, S.; Josephe Amiot, M.; Rochette, I.; Bellagha, S.; Mouquet-Rivier, C. Soaking and cooking modify the alpha-galacto-oligosaccharide and dietary fibre content in five Mediterranean legumes. Int. J. Food Sci. Nutr. 2019, 70, 551–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Jin, Z.; Simsek, S.; Hall, C.; Rao, J.; Chen, B. Effect of germination on the chemical composition, thermal, pasting, and moisture sorption properties of flours from chickpea, lentil, and yellow pea. Food Chem. 2019, 295, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Kotlarz, A.; Sujak, A.; Strobel, W.; Grzesiak, W. Chemical Composition and Nutritive Value of Protein of the Pea Seeds-Effect of Harvesting Year and Variety. Veg. Crops Res. Bull. 2011, 75, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Frias, J.; Giacomino, S.; Peñas, E.; Pellegrino, N.; Ferreyra, V.; Apro, N.; Carrión, M.O.; Vidal-Valverde, C. Assessment of the nutritional quality of raw and extruded Pisum sativum L. var. laguna seeds. LWT Food Sci. Technol. 2011, 44, 1303–1308. [Google Scholar] [CrossRef] [Green Version]
- Martín-Cabrejas, M.A.; Ariza, N.; Esteban, R.; Mollá, E.; Waldron, K.; López-Andréu, F.J. Effect of Germination on the Carbohydrate Composition of the Dietary Fiber of Peas (Pisum sativum L.). J. Agric. Food Chem. 2003, 51, 1254–1259. [Google Scholar] [CrossRef] [PubMed]
- Boye, J.; Zare, F.; Pletch, A. Pulse proteins: Processing, characterization, functional properties and applications in food and feed. Food Res. Int. 2010, 43, 414–431. [Google Scholar] [CrossRef]
- Cipollone, M.A.; Tironi, V.A. Yellow pea flour and protein isolate as sources of antioxidant peptides after simulated gastrointestinal digestion. Legum. Sci. 2020, 2, e59. [Google Scholar] [CrossRef]
- Stoughton-Ens, M.D.; Hatcher, D.W.; Wang, N.; Warkentin, T.D. Influence of genotype and environment on the dietary fiber content of field pea (Pisum sativum L.) grown in Canada. Food Res. Int. 2010, 43, 547–552. [Google Scholar] [CrossRef]
- Agume, A.S.; Njintang, N.Y.; Mbofung, C.M. Effect of Soaking and Roasting on the Physicochemical and Pasting Properties of Soybean Flour. Foods 2017, 6, 12. [Google Scholar] [CrossRef]
- Singh, R.K.; Bohra, N.; Sharma, L. Valorizing faba bean for animal feed supplements via biotechnological approach: Opinion. Biocatal. Agric. Biotechnol. 2019, 17, 366–368. [Google Scholar] [CrossRef]
- Redondo-Cuenca, A.; Villanueva-Suárez, M.J.; Rodríguez-Sevilla, M.D.; Mateos-Aparicio, I. Chemical composition and dietary fibre of yellow and green commercial soybeans (Glycine max). Food Chem. 2007, 101, 1216–1222. [Google Scholar] [CrossRef]
- Maeta, A.; Katsukawa, M.; Hayase, Y.; Takahashi, K. Comparisons of Soybean and Wheat; in the Focus on the Nutritional Aspects and Acute Appetite Sensation. Foods 2022, 11, 389. [Google Scholar] [CrossRef] [PubMed]
- Ukwuru, M.U. Effect of processing on the chemical qualities and functional properties of soy flour. Plant Foods Hum. Nutr. 2003, 58, 1–11. [Google Scholar] [CrossRef]
- Grieshop, C.M.; Kadzere, C.T.; Clapper, G.M.; Flickinger, E.A.; Bauer, L.L.; Frazier, R.L.; Fahey, G.C. Chemical and Nutritional Characteristics of United States Soybeans and Soybean Meals. J. Agric. Food Chem. 2003, 51, 7684–7691. [Google Scholar] [CrossRef]
- Grela, E.R.; Günter, K.D. Fatty acid composition and tocopherol content of some legume seeds. Anim. Feed Sci. Technol. 1995, 52, 325–331. [Google Scholar] [CrossRef]
- Nti, C.A.; Plahar, W.A.; Annan, N.T. Development and quality characteristics of shelf-stable soy-agushie: A residual by-product of soymilk production. Food Sci. Nutr. 2016, 4, 315–321. [Google Scholar] [CrossRef]
- Grieshop, C.M.; Fahey, G.C., Jr. Comparison of quality characteristics of soybeans from Brazil, China, and the United States. J. Agric. Food Chem. 2001, 49, 2669–2673. [Google Scholar] [CrossRef]
- Pisarikova, B.; Zraly, Z. Dietary fibre content in lupine (Lupinus albus L.) and soya (Glycine max L.) seeds. Acta Vet. Brno 2010, 79, 211–216. [Google Scholar] [CrossRef]
- Kahlon, T.S.; Shao, Q. In vitro binding of bile acids by soy bean (Glycine max), black eye bean (Vigna unguiculata), garbanzo (Cicer arietinum) and lima bean (Phaseolus lunatus). Food Chem. 2004, 86, 435–440. [Google Scholar] [CrossRef]
- Jiménez-Escrig, A.; Serra, M.-T.; Rupérez, P. Non-digestible carbohydrates in Brazilian soybean seeds [Glycine max (L.) Merril]. Int. J. Food Sci. Technol. 2010, 45, 2524–2530. [Google Scholar] [CrossRef]
- Wilson, L.A.; Birmingham, V.A.; Moon, D.P.; Snyder, H.E. Isolation and characterization of starch from mature soybeans. Cereal Chem. 1978, 55, 661–670. [Google Scholar]
- Alghamdi, S.S.; Khan, M.A.; El-Harty, E.H.; Ammar, M.H.; Farooq, M.; Migdadi, H.M. Comparative phytochemical profiling of different soybean (Glycine max (L.) Merr) genotypes using GC-MS. Saudi J. Biol. Sci. 2018, 25, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Amarowicz, R.; Shahidi, F. Antioxidant activity of broad bean seed extract and its phenolic composition. J. Funct. Foods 2017, 38, 656–662. [Google Scholar] [CrossRef]
- Li, L.; Yuan, T.Z.; Setia, R.; Raja, R.B.; Zhang, B.; Ai, Y. Characteristics of pea, lentil and faba bean starches isolated from air-classified flours in comparison with commercial starches. Food Chem. 2019, 276, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Çalışkantürk Karataş, S.; Günay, D.; Sayar, S. In vitro evaluation of whole faba bean and its seed coat as a potential source of functional food components. Food Chem. 2017, 230, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Purves, R.W.; Zhang, H.; Khazaei, H.; Vandenberg, A. Rapid analysis of medically relevant compounds in faba bean seeds using FAIMS and mass spectrometry. Int. J. Ion Mobil. Spectrom. 2017, 20, 125–135. [Google Scholar] [CrossRef]
- Coda, R.; Varis, J.; Verni, M.; Rizzello, C.G.; Katina, K. Improvement of the protein quality of wheat bread through faba bean sourdough addition. LWT Food Sci. Technol. 2017, 82, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Jakubczyk, A.; Karaś, M.; Złotek, U.; Szymanowska, U.; Baraniak, B.; Bochnak, J. Peptides obtained from fermented faba bean seeds (Vicia faba) as potential inhibitors of an enzyme involved in the pathogenesis of metabolic syndrome. LWT 2019, 105, 306–313. [Google Scholar] [CrossRef]
- Karkouch, I.; Tabbene, O.; Gharbi, D.; Ben Mlouka, M.A.; Elkahoui, S.; Rihouey, C.; Coquet, L.; Cosette, P.; Jouenne, T.; Limam, F. Antioxidant, antityrosinase and antibiofilm activities of synthesized peptides derived from Vicia faba protein hydrolysate: A powerful agents in cosmetic application. Ind. Crops Prod. 2017, 109, 310–319. [Google Scholar] [CrossRef]
- Nosworthy, M.G.; Medina, G.; Franczyk, A.J.; Neufeld, J.; Appah, P.; Utioh, A.; Frohlich, P.; House, J.D. Effect of processing on the in vitro and in vivo protein quality of beans (Phaseolus vulgaris and Vicia Faba). Nutrients 2018, 10, 671. [Google Scholar] [CrossRef] [Green Version]
- Dugardin, C.; Cudennec, B.; Tourret, M.; Caron, J.; Guérin-Deremaux, L.; Behra-Miellet, J.; Lefranc-Millot, C.; Ravallec, R. Explorative Screening of Bioactivities Generated by Plant-Based Proteins after In Vitro Static Gastrointestinal Digestion. Nutrients 2020, 12, 3746. [Google Scholar] [CrossRef] [PubMed]
- León-Espinosa, E.B.; Sánchez-Chino, X.; Garduño-Siciliano, L.; Álvarez-González, R.I.; Dávila-Ortiz, G.; Madrigal-Bujaidar, E.; Téllez-Medina, D.I.; Jiménez-Martínez, C. Hypocholesterolemic and Anticarcinogenic Effect of Vicia faba Protein Hydrolyzates. Nutr. Cancer 2016, 68, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Altaf Khan, M.; Megahed, H.A.; Hussein, M.M.; El-Harty, E.H.; Magdi, A.O.; Muhammad, F.; Salem, S.A. Comparative nutritional profiles of various faba bean and chickpea genotypes. Int. J. Agric. Biol. 2015, 17, 449–457. [Google Scholar] [CrossRef]
- Multari, S.; Neacsu, M.; Scobbie, L.; Cantlay, L.; Duncan, G.; Vaughan, N.; Stewart, D.; Russell, W.R. Nutritional and Phytochemical Content of High-Protein Crops. J. Agric. Food Chem. 2016, 64, 7800–7811. [Google Scholar] [CrossRef]
- Maalouf, F.; Hu, J.; O’Sullivan, D.M.; Zong, X.; Hamwieh, A.; Kumar, S.; Baum, M. Breeding and genomics status in faba bean (Vicia faba). Plant Breed. 2019, 138, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Wanasundara, P.; Shand, P. Short-term germination of faba bean (Vicia faba L.) and the effect on selected chemical constituents. Appl. Food Res. 2021, 2, 100030. [Google Scholar] [CrossRef]
- Barłóg, P.; Grzebisz, W.; Łukowiak, R. The Effect of Potassium and Sulfur Fertilization on Seed Quality of Faba Bean (Vicia faba L.). Agronomy 2019, 9, 209. [Google Scholar] [CrossRef] [Green Version]
- Senberga, A.; Dubova, L.; Alsina, I.; Strauta, L. Rhizobium sp.—A potential tool for improving protein content in peas and faba beans. Rural Sustain. Res. 2017, 37, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Ntatsi, G.; Gutiérrez-Cortines, M.E.; Karapanos, I.; Barros, A.; Weiss, J.; Balliu, A.; Rosa, E.A.d.S.; Savvas, D. The quality of leguminous vegetables as influenced by preharvest factors. Sci. Horticult. 2018, 232, 191–205. [Google Scholar] [CrossRef]
- Osborne, T.B. The Proteins of The Wheat Kernel; Carnegie institution of Washington: Washington, DC, USA, 1907; p. 119. [Google Scholar]
- Alghamdi, S.S. Chemical composition of faba bean (Vicia faba L.) genotypes under various water regimes. Pak. J. Nutr. 2009, 8, 477–482. [Google Scholar] [CrossRef] [Green Version]
- Utsumi, S. Plant Food Protein Engineering. In Advances in Food and Nutrition Research; Kinsella, J.E., Ed.; Academic Press: Kyoto, Japan, 1992; Volume 36, pp. 89–208. [Google Scholar]
- Müntz, K.; Horstmann, C.; Schlesier, B. Vicia globulins. In Seed Proteins; Shewry, P.R., Casey, R., Eds.; Springer: Dordrecht, The Netherlands, 1999; pp. 259–284. [Google Scholar]
- Singhal, A.; Stone, A.K.; Vandenberg, A.; Tyler, R.; Nickerson, M.T. Effect of genotype on the physicochemical and functional attributes of faba bean (Vicia faba L.) protein isolates. Food Sci. Biotechnol. 2016, 25, 1513–1522. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.; Stone, A.K.; Yovchev, A.G.; Peter, R.; Vandenberg, A.; Nickerson, M.T. Effect of genotype and environment on the surface characteristics and functionality of air-classified faba bean protein concentrates. Eur. Food Res. Technol. 2016, 242, 1903–1911. [Google Scholar] [CrossRef]
- Warsame, A.O.; Michael, N.; O’Sullivan, D.M.; Tosi, P. Identification and Quantification of Major Faba Bean Seed Proteins. J. Agric. Food Chem. 2020, 68, 8535–8544. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Takako, F.; Meili, Z.; Shiori, M.; Maruyama, N.; Utsumi, S. Comparison of physicochemical properties of 7S and 11S globulins from pea, fava bean, cowpea, and French bean with those of soybean-french bean 7S globulin exhibits excellent properties. J. Agric. Food Chem. 2008, 56, 10273–10279. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, X.; Hou, W.; Li, P.; Sha, W.; Tian, Y. Structure and function of seed storage proteins in faba bean (Vicia faba L.). 3 Biotech 2017, 7, 74. [Google Scholar] [CrossRef] [Green Version]
- Rahate, K.A.; Madhumita, M.; Prabhakar, P.K. Nutritional composition, anti-nutritional factors, pretreatments-cum-processing impact and food formulation potential of faba bean (Vicia faba L.): A comprehensive review. LWT 2021, 138, 110796. [Google Scholar] [CrossRef]
- Sharan, S.; Zanghelini, G.; Zotzel, J.; Bonerz, D.; Aschoff, J.; Saint-Eve, A.; Maillard, M.N. Fava bean (Vicia faba L.) for food applications: From seed to ingredient processing and its effect on functional properties, antinutritional factors, flavor, and color. Compr. Rev. Food Sci. Food Saf. 2021, 20, 401–428. [Google Scholar] [CrossRef]
- Wang, D.; Lin, H.; Kan, J.; Liu, L.; Zeng, X.; Shen, G. Food Chemistry; Nova Science Publishers, Incorporated: Hauppauge, NY, USA, 2012. [Google Scholar]
- Canadian Food Inspection Agency. Proteins. Available online: https://www.inspection.gc.ca/food-label-requirements/labelling/industry/nutrition-labelling/elements-within-the-nutrition-facts-table/eng/1389206763218/1389206811747?chap=7 (accessed on 30 March 2020).
- Health Canada. Method FO-1. Available online: https://www.hc-sc.gc.ca/fn-an/alt_formats/hpfb-dgpsa/pdf/res-rech/fo-1-eng.pdf (accessed on 30 March 2020).
- Marinangeli, C.P.F.; House, J.D. Potential impact of the digestible indispensable amino acid score as a measure of protein quality on dietary regulations and health. Nutr. Rev. 2017, 75, 658–667. [Google Scholar] [CrossRef] [Green Version]
- FAO. Dietary Protein Quality Evaluation in Human Nutrition: Report of An FAO Expert Consultation, Auckland, New Zealand, 31 March–2 April 2011; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Schaafsma, G. Advantages and limitations of the protein digestibility-corrected amino acid score (PDCAAS) as a method for evaluating protein quality in human diets. Br. J. Nutr. 2012, 108, S333–S336. [Google Scholar] [CrossRef] [Green Version]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Bohn, T.; Carriere, F.; Day, L.; Deglaire, A.; Egger, L.; Freitas, D.; Golding, M.; Le Feunteun, S.; Macierzanka, A.; Menard, O.; et al. Correlation between in vitro and in vivo data on food digestion. What can we predict with static in vitro digestion models? Crit. Rev. Food Sci. Nutr. 2018, 58, 2239–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, K.A.; Gallagher, E.; Burke, R.; McCarthy, S.; Barry-Ryan, C. Proximate composition and anti-nutritional factors of fava-bean (Vicia faba), green-pea and yellow-pea (Pisum sativum) flour. J. Food Comp. Anal. 2019, 82, 103233. [Google Scholar] [CrossRef]
- Małgorzata, Ś.; Jerzy, K.; Ewa, H. The Effect of Feeding Native Faba Bean Seeds (Vicia faba L.) to Sows and Supplemented With Enzymes to Piglets and Growing Pigs. Ann. Anim. Sci. 2018, 18, 1007–1027. [Google Scholar] [CrossRef] [Green Version]
- Carbonaro, M.; Grant, G.; Cappelloni, M. Heat-induced denaturation impairs digestibility of legume (Phaseolus vulgaris L. and Vicia faba L.) 7S and 11S globulins in the small intestine of rat. Sci. Food Agric. 2005, 85, 65–72. [Google Scholar] [CrossRef]
- Laleg, K.; Salles, J.; Berry, A.; Giraudet, C.; Patrac, V.; Guillet, C.; Denis, P.; Tessier, F.J.; Guilbaud, A.; Howsam, M.; et al. Nutritional evaluation of mixed wheat-faba bean pasta in growing rats: Impact of protein source and drying temperature on protein digestibility and retention. Br. J. Nutr. 2019, 121, 496–507. [Google Scholar] [CrossRef]
- Leterme, P.; Monmart, T.; Baudart, E. Amino Acid Composition of Pea (Pisum sativum) Proteins and Protein Profile of Pea Flour. J. Sci. Food Agric. 1990, 53, 107–110. [Google Scholar] [CrossRef]
- Kashyap, S.; Varkey, A.; Shivakumar, N.; Devi, S.; Reddy, B.H.R.; Thomas, T.; Preston, T.; Sreeman, S.; Kurpad, A.V. True ileal digestibility of legumes determined by dual-isotope tracer method in Indian adults. Am. J. Clin. Nutr. 2019, 110, 873–882. [Google Scholar] [CrossRef]
- Roux, L.L.; Chacon, R.; Dupont, D.; Jeantet, R.; Deglaire, A.; Nau, F. In vitro static digestion reveals how plant proteins modulate model infant formula digestibility. Food Res. Int. 2020, 130, 108917. [Google Scholar] [CrossRef]
- Hamosh, M. Digestion in the Newborn. Clin. Perinatol. 1996, 23, 191–209. [Google Scholar] [CrossRef]
- Gilani, G.S.; Sepehr, E. Protein Digestibility and Quality in Products Containing Antinutritional Factors Are Adversely Affected by Old Age in Rats. J. Nutr. 2003, 133, 220–225. [Google Scholar] [CrossRef]
- Carbonaro, M.; Maselli, P.; Nucara, A. Relationship between digestibility and secondary structure of raw and thermally treated legume proteins: A Fourier transform infrared (FT-IR) spectroscopic study. Amino Acids 2012, 43, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Verni, M.; Koivula, H.; Montemurro, M.; Seppa, L.; Kemell, M.; Katina, K.; Coda, R.; Gobbetti, M. Influence of fermented faba bean flour on the nutritional, technological and sensory quality of fortified pasta. Food Funct. 2017, 8, 860–871. [Google Scholar] [CrossRef] [PubMed]
- Polanowska, K.; Grygier, A.; Kuligowski, M.; Rudzińska, M.; Nowak, J. Effect of tempe fermentation by three different strains of Rhizopus oligosporus on nutritional characteristics of faba beans. LWT 2020, 122, 109024. [Google Scholar] [CrossRef]
- Sozer, N.; Melama, L.; Silbir, S.; Rizzello, C.G.; Flander, L.; Poutanen, K. Lactic acid fermentation as a pre-treatment process for faba bean flour and its effect on textural, structural and nutritional properties of protein-enriched gluten-free faba bean breads. Foods 2019, 8, 431. [Google Scholar] [CrossRef] [Green Version]
- Berrazaga, I.; Mession, J.L.; Laleg, K.; Salles, J.; Guillet, C.; Patrac, V.; Giraudet, C.; Le Bacquer, O.; Boirie, Y.; Micard, V.; et al. Formulation, process conditions, and biological evaluation of dairy mixed gels containing fava bean and milk proteins: Effect on protein retention in growing young rats. J. Dairy Sci. 2018, 102, 1066–1082. [Google Scholar] [CrossRef] [Green Version]
- Chandra-Hioe, M.V.; Wong, C.H.M.; Arcot, J. The Potential Use of Fermented Chickpea and Faba Bean Flour as Food Ingredients. Plant Foods Hum. Nutr. 2016, 71, 90–95. [Google Scholar] [CrossRef]
- Ortiz, L.T.; Centeno, C.; Treviño, J. Tannins in faba bean seeds: Effects on the digestion of protein and amino acids in growing chicks. Anim. Feed Sci. Technol. 1993, 41, 271–278. [Google Scholar] [CrossRef]
- Zduńczyk, Z.; Juśkiewicz, J.; Wróblewska, M.; Flis, M. Effect of faba bean seeds with different content of proanthocyanidins on growth of rats, caecal enzyme activity and metabolism indices. Acta Aliment. 2003, 32, 161–168. [Google Scholar] [CrossRef]
- Kosińska, A.; Karamać, M.; Penkacik, K.; Urbalewicz, A.; Amarowicz, R. Interactions between tannins and proteins isolated from broad bean seeds (Vicia faba Major) yield soluble and non-soluble complexes. Eur. Food Res. Technol. 2011, 233, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Weihua, X.; Miao, Z.; Jing, L.; Chuanxiu, X.; Yuwei, L. Effects of phytase and tannase on in vivo nutruitive utilisation of faba bean (Vicia faba L.) flour. Int. Food Res. J. 2015, 22, 1550–1556. [Google Scholar]
- Yu-Wei, L.; Wei-Hua, X. Effect of different processing methods on certain antinutritional factors and protein digestibility in green and white faba bean (Vicia faba L.). CyTA-J. Food 2013, 11, 43–49. [Google Scholar]
- Alonso, R.; Aguirre, A.; Marzo, F. Effects of extrusion and traditional processing methods on antinutrients and in vitro digestibility of protein and starch in faba and kidney beans. Food Chem. 2000, 68, 159–165. [Google Scholar] [CrossRef]
- Lafarga, T.; Villaró, S.; Bobo, G.; Simó, J.; Aguiló-Aguayo, I. Bioaccessibility and antioxidant activity of phenolic compounds in cooked pulses. Int. J. Food Sci. Technol. 2019, 54, 1816–1823. [Google Scholar] [CrossRef] [Green Version]
- Erba, D.; Angelino, D.; Marti, A.; Manini, F.; Faoro, F.; Morreale, F.; Pellegrini, N.; Casiraghi, M.C. Effect of sprouting on nutritional quality of pulses. Int. J. Food Sci. Nutr. 2019, 70, 30–40. [Google Scholar] [CrossRef]
- Zdunczyk, Z.; Frejnagel, S.; Wróblewska, M.; Juskiewicz, J.; Oszmiański, J.; Estrella, I. Biological activity of polyphenol extracts from different plant sources. Food Res. Int. 2002, 35, 183–186. [Google Scholar] [CrossRef]
- Turco, I.; Ferretti, G.; Bacchetti, T. Review of the health benefits of faba bean (Vicia faba L.) polyphenols. J. Food Nutr. Res. 2016, 55, 283–293. [Google Scholar]
- Konietzny, U.; Greiner, R. PHYTIC ACID | Nutritional Impact. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 4555–4563. [Google Scholar]
- Luo, Y.; Xie, W.; Jin, X.; Zhang, B.; Wang, Q.; He, Y. The impact of processing on phytic acid, in vitro soluble zinc and Phy/Zn molar ratio of faba bean (Vicia faba L.). Int. Food Res. J. 2013, 20, 1285–1291. [Google Scholar]
- Rosa-Sibakov, N.; Re, M.; Karsma, A.; Laitila, A.; Nordlund, E. Phytic Acid Reduction by Bioprocessing as a Tool to Improve the in Vitro Digestibility of Faba Bean Protein. J. Agric. Food Chem. 2018, 66, 10394–10399. [Google Scholar] [CrossRef]
- Vidal-Valverde, C.; Frias, J.; Sotomayor, C.; Diaz-Pollan, C.; Fernandez, M.; Urbano, G. Nutrients and antinutritional factors in faba beans as affected by processing. Eur. Food Res. Technol. 1998, 207, 140–145. [Google Scholar] [CrossRef]
- Hejdysz, M.; Kaczmarek, S.A.; Rutkowski, A. Extrusion cooking improves the metabolisable energy of faba beans and the amino acid digestibility in broilers. Anim. Feed Sci. Technol. 2016, 212, 100–111. [Google Scholar] [CrossRef]
- Evandro Fei, F.; Abdallah Abd Elazeem, H.; Jack Ho, W.; Clara Shui Fern, B.; Saeed Saad, S.; Tzi Bun, N. Isolation of a New Trypsin Inhibitor from the Faba Bean (Vicia faba cv. Giza 843) with Potential Medicinal Applications. Protein Pept. Lett. 2011, 18, 64–72. [Google Scholar] [CrossRef]
- Ye, X.Y.; Ng, T.B.; Rao, P.F. A Bowman-Birk-type trypsin-chymotrypsin inhibitor from broad beans. Biochem. Biophys. Res. Commun. 2001, 289, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Zambrowicz, A.; Pokora, M.; Setner, B.; Dąbrowska, A.; Szołtysik, M.; Babij, K.; Szewczuk, Z.; Trziszka, T.; Lubec, G.; Chrzanowska, J. Multifunctional peptides derived from an egg yolk protein hydrolysate: Isolation and characterization. Amino Acids 2015, 47, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Manzanares, P.; Gandía, M.; Garrigues, S.; Marcos, J.F. Improving Health-Promoting Effects of Food-Derived Bioactive Peptides through Rational Design and Oral Delivery Strategies. Nutrients 2019, 11, 2545. [Google Scholar] [CrossRef] [Green Version]
- Karami, Z.; Akbari-Adergani, B. Bioactive food derived peptides: A review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef]
- Lopez-Barrios, L.; Gutierrez-Uribe, J.A.; Serna-Saldivar, S.O. Bioactive peptides and hydrolysates from pulses and their potential use as functional ingredients. J. Food Sci. 2014, 79, R273–R283. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP Database and Other Programs for Processing Bioactive Peptide Sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [CrossRef] [Green Version]
- Rizzello, C.G.; Verni, M.; Bordignon, S.; Gramaglia, V.; Gobbetti, M. Hydrolysate from a mixture of legume flours with antifungal activity as an ingredient for prolonging the shelf-life of wheat bread. Food Microbiol. 2017, 64, 72–82. [Google Scholar] [CrossRef]
- Xiao, P.; Liu, Y.; Rizwan ur, R.; Kang, R.; Wang, Y. A novel lactic acid bacteria growth-stimulating peptide from broad bean (Vicia faba L.) protein hydrolysates. Adv. J. Food Sci. Technol. 2015, 7, 697–703. [Google Scholar] [CrossRef]
- Corrochano, A.R.; Cal, R.; Kennedy, K.; Wall, A.; Murphy, N.; Trajkovic, S.; O’Callaghan, S.; Adelfio, A.; Khaldi, N. Characterising the efficacy and bioavailability of bioactive peptides identified for attenuating muscle atrophy within a Vicia faba-derived functional ingredient. Curr. Res. Food Sci. 2021, 4, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Felix, M.; Cermeño, M.; FitzGerald, R.J. Assessment of the microstructural characteristics and the in vitro bioactive properties of sunflower oil-based emulsions stabilized by fava bean (Vicia faba) protein. Food Hydrocoll. 2019, 97, 105220. [Google Scholar] [CrossRef]
- Parya Samaei, S.; Ghorbani, M.; Tagliazucchi, D.; Martini, S.; Gotti, R.; Themelis, T.; Tesini, F.; Gianotti, A.; Gallina Toschi, T.; Babini, E. Functional, nutritional, antioxidant, sensory properties and comparative peptidomic profile of faba bean (Vicia faba, L.) seed protein hydrolysates and fortified apple juice. Food Chem. 2020, 330, 127120. [Google Scholar] [CrossRef]
- Ashraf, J.; Awais, M.; Liu, L.; Khan, M.I.; Tong, L.-T.; Ma, Y.; Wang, L.; Zhou, X.; Zhou, S. Effect of thermal processing on cholesterol synthesis, solubilisation into micelles and antioxidant activities using peptides of Vigna angularis and Vicia faba. LWT 2020, 129, 109504. [Google Scholar] [CrossRef]
- Raveschot, C.; Cudennec, B.; Coutte, F.; Flahaut, C.; Fremont, M.; Drider, D.; Dhulster, P. Production of Bioactive Peptides by Lactobacillus Species: From Gene to Application. Front. Microbiol. 2018, 9, 2354. [Google Scholar] [CrossRef] [Green Version]
- Jawed, A.; Singh, G.; Kohli, S.; Sumera, A.; Haque, S.; Prasad, R.; Paul, D. Therapeutic role of lipases and lipase inhibitors derived from natural resources for remedies against metabolic disorders and lifestyle diseases. S. Afr. J. Bot. 2019, 120, 25–32. [Google Scholar] [CrossRef]
- Neels, J.G. A role for 5-lipoxygenase products in obesity-associated inflammation and insulin resistance. Adipocyte 2013, 2, 262–265. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on Mechanisms of In Vitro Antioxidant Activity of Polysaccharides. Oxid. Med. Cell Longev. 2016, 2016, 5692852. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.S.; Alvarenga Brizola, V.R.; Granato, D. High-throughput assay comparison and standardization for metal chelating capacity screening: A proposal and application. Food Chem. 2017, 214, 515–522. [Google Scholar] [CrossRef]
- Cal, R.; Davis, H.; Kerr, A.; Wall, A.; Molloy, B.; Chauhan, S.; Trajkovic, S.; Holyer, I.; Adelfio, A.; Khaldi, N. Preclinical Evaluation of a Food-Derived Functional Ingredient to Address Skeletal Muscle Atrophy. Nutrients 2020, 12, 2274. [Google Scholar] [CrossRef]
- Sun, X.; Acquah, C.; Aluko, R.E.; Udenigwe, C.C. Considering food matrix and gastrointestinal effects in enhancing bioactive peptide absorption and bioavailability. J. Funct. Foods 2020, 64, 103680. [Google Scholar] [CrossRef]
- Iwaniak, A.; Hrynkiewicz, M.; Bucholska, J.; Minkiewicz, P.; Darewicz, M. Understanding the nature of bitter-taste di- and tripeptides derived from food proteins based on chemometric analysis. J. Food Biochem. 2019, 43, e12500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramdath, D.; Renwick, S.; Duncan, A.M. The Role of Pulses in the Dietary Management of Diabetes. Can. J. Diabetes 2016, 40, 355–363. [Google Scholar] [CrossRef] [PubMed]
- DeMartino, P.; Cockburn, D.W. Resistant starch: Impact on the gut microbiome and health. Curr. Opin. Biotechnol. 2020, 61, 66–71. [Google Scholar] [CrossRef]
- Dong, H.; Vasanthan, T. Amylase resistance of corn, faba bean, and field pea starches as influenced by three different phosphorylation (cross-linking) techniques. Food Hydrocoll. 2020, 101, 105506. [Google Scholar] [CrossRef]
- Martens, B.M.J.; Gerrits, W.J.J.; Bruininx, E.M.A.M.; Schols, H.A. Amylopectin structure and crystallinity explains variation in digestion kinetics of starches across botanic sources in an in vitro pig model. J. Anim. Sci. Biotechnol. 2018, 9, 91. [Google Scholar] [CrossRef] [Green Version]
- Pérez, S.; Bertoft, E. The molecular structures of starch components and their contribution to the architecture of starch granules: A comprehensive review. Starch Stärke 2010, 62, 389–420. [Google Scholar] [CrossRef]
- Bertoft, E. Understanding Starch Structure: Recent Progress. Agronomy 2017, 7, 56. [Google Scholar] [CrossRef]
- Greffeuille, V.; Marsset-Baglieri, A.; Molinari, N.; Cassan, D.; Sutra, T.; Avignon, A.; Micard, V. Enrichment of pasta with faba bean does not impact glycemic or insulin response but can enhance satiety feeling and digestive comfort when dried at very high temperature. Food Funct. 2015, 6, 2996–3005. [Google Scholar] [CrossRef]
- Tazrart, K.; Lamacchia, C.; Zaidi, F.; Haros, M. Nutrient composition and in vitro digestibility of fresh pasta enriched with Vicia faba. J. Food Compos. Anal. 2016, 47, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Copeland, L. Molecular disassembly of starch granules during gelatinization and its effect on starch digestibility: A review. Food Funct. 2013, 4, 1564–1580. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, H.; Waheed, R.; Nawaz, M.; Shahwar, D. Physical and Chemical Modifications in Starch Structure and Reactivity; Intechopen: London, UK, 2020. [Google Scholar]
- Mudryj, A.N.; Yu, N.; Aukema, H.M. Nutritional and health benefits of pulses. Appl. Physiol. Nutr. Metab. 2014, 39, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.M.H.; Gyles, C.L.; Marinangeli, C.P.F.; Carlberg, J.G.; Jones, P.J.H. Dietary fibre intakes and reduction in functional constipation rates among Canadian adults: A cost-of-illness analysis. Food Nutr. Res. 2015, 59, 28646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bélanger, M.; Poirier, M.; Jbilou, J.; Scarborough, P. Modelling the impact of compliance with dietary recommendations on cancer and cardiovascular disease mortality in Canada. Pub. Health 2014, 128, 222–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesch, E.D.; Carr, T.P. Food Ingredients That Inhibit Cholesterol Absorption. Prev. Nutr. Food Sci. 2017, 22, 67–80. [Google Scholar] [CrossRef]
- Gullón, P.; Gullón, B.; Tavaria, F.; Vasconcelos, M.; Gomes, A.M. In vitro fermentation of lupin seeds (Lupinus albus) and broad beans (Vicia faba): Dynamic modulation of the intestinal microbiota and metabolomic output. Food Funct. 2015, 6, 3316–3322. [Google Scholar] [CrossRef]
- Fan, P.-H.; Zang, M.-T.; Xing, J. Oligosaccharides composition in eight food legumes species as detected by high-resolution mass spectrometry. J. Sci. Food Agric. 2015, 95, 2228–2236. [Google Scholar] [CrossRef]
- Lunn, J.; Theobald, H.E. The health effects of dietary unsaturated fatty acids. Nutr. Bull. 2006, 31, 178–224. [Google Scholar] [CrossRef]
- Yang, Z.; Piironen, V.; Lampi, A.-M. Lipid-modifying enzymes in oat and faba bean. Food Res. Int. 2017, 100, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Zhong-qing, J.; Pulkkinen, M.; Yu-jie, W.; Lampi, A.M.; Stoddard, F.L.; Salovaara, H.; Piironen, V.; Sontag-Strohm, T. Faba bean flavour and technological property improvement by thermal pre-treatments. LWT Food Sci. Technol. 2016, 68, 295–305. [Google Scholar] [CrossRef]
- Grela, E.R.; Samolińska, W.; Kiczorowska, B.; Klebaniuk, R.; Kiczorowski, P. Content of Minerals and Fatty Acids and Their Correlation with Phytochemical Compounds and Antioxidant Activity of Leguminous Seeds. Biol. Trace Elem. Res. 2017, 180, 338–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farquhar, W.B.; Edwards, D.G.; Jurkovitz, C.T.; Weintraub, W.S. Dietary sodium and health: More than just blood pressure. J. Am. Coll. Cardiol. 2015, 65, 1042–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hever, J. Plant-Based Diets: A Physician’s Guide. Perm. J. 2016, 20, 15-082. [Google Scholar] [CrossRef] [PubMed]
- Toledo, N.M.V.; Brigide, P.; López-Nicolás, R.; Frontela, C.; Ros, G.; Canniatti-Brazaca, S.G. Higher inositol phosphates and total oxalate of cookies containing fruit by-products and their influence on calcium, iron, and zinc bioavailability by Caco-2 cells. Cereal Chem. 2019, 96, 456–464. [Google Scholar] [CrossRef]
- Gad, S.S.; el-Zalaki, M.E.; Mohamed, M.S.; Mohasseb, S.Z. Oxalate content of some leafy vegetables and dry legumes consumed widely in Egypt. Food Chem. 1982, 8, 169–177. [Google Scholar] [CrossRef]
- Xie, W.-H.; Jin, X.-X.; Wang, Q.; He, Y.-J. Effects of germination on iron, zinc, calcium, manganese, and copper availability from cereals and legumes AU-Luo, Yu-Wei. CyTA J. Food 2014, 12, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Xie, W. Effect of phytase treatment on iron bioavailability in faba bean (Vicia faba L.) flour. Food Chem. 2012, 134, 1251–1255. [Google Scholar] [CrossRef]
- Liao, X.; Yun, S.; Zhao, G. Structure, Function, and Nutrition of Phytoferritin: A Newly Functional Factor for Iron Supplement. Crit. Rev. Food Sci. Nutr. 2014, 54, 1342–1352. [Google Scholar] [CrossRef]
- Perfecto, A.; Rodriguez-Ramiro, I.; Rodriguez-Celma, J.; Sharp, P.; Balk, J.; Fairweather-Tait, S. Pea Ferritin Stability under Gastric pH Conditions Determines the Mechanism of Iron Uptake in Caco-2 Cells. J. Nutr. 2018, 148, 1229–1235. [Google Scholar] [CrossRef]
- Shaojun, Y.; Senpei, Y.; Luyao, H.; Xin, Q.; Peng, M.; Guanghua, Z. Isolation and characterization of a new phytoferritin from broad bean (Vicia faba) seed with higher stability compared to pea seed ferritin. Food Res. Int. 2012, 48, 271–276. [Google Scholar]
- Waller, D.G.; Sampson, A.P. 24-Extrapyramidal movement disorders and spasticity. In Medical Pharmacology and Therapeutics, 5th ed.; Waller, D.G., Sampson, A.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 325–336. [Google Scholar]
- Mohseni Mehran, S.M.; Golshani, B. Simultaneous determination of levodopa and carbidopa from fava bean, green peas and green beans by high performance liquid gas chromatography. J. Clin. Diagn. Res. JCDR 2013, 7, 1004–1007. [Google Scholar] [CrossRef]
- Rabey, J.M.; Vered, Y.; Shabtai, H.; Graff, E.; Korczyn, A.D. Improvement of parkinsonian features correlate with high plasma levodopa values after broad bean (Vicia faba) consumption. J. Neurol Neurosurg. Psychiatr. 1992, 55, 725–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etemadi, F.; Hashemi, M.; Randhir, R.; ZandVakili, O.; Ebadi, A. Accumulation of l-DOPA in various organs of faba bean and influence of drought, nitrogen stress, and processing methods on l-DOPA yield. Crop J. 2018, 6, 426–434. [Google Scholar] [CrossRef]
- Topal, N.; Bozoglu, H. Determination of L-dopa (L-3, 4-dihydroxyphenylalanine) content of some faba bean (Vicia faba L.) genotypes. Tarim Bilimleri. Dergisi. 2016, 22, 145–151. [Google Scholar]
- Polanowska, K.; Łukasik, R.M.; Kuligowski, M.; Nowak, J. Development of a sustainable, simple, and robust method for efficient L-DOPA extraction. Molecules 2019, 24, 2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardador-Martínez, A.; Maya-Ocaña, K.; Ortiz-Moreno, A.; Herrera-Cabrera, B.E.; Dávila-Ortiz, G.; Múzquiz, M.; Martín-Pedrosa, M.; Burbano, C.; Cuadrado, C.; Jiménez-Martínez, C. Effect of Roasting and Boiling on the Content of Vicine, Convicine and L-3,4-dihydroxyphenylalanine in Vicia faba L. J. Food Qual. 2012, 35, 419–428. [Google Scholar] [CrossRef]
- Abdel-Sattar, E.; Mahrous, E.A.; Thabet, M.M.; Elnaggar, D.M.Y.; Youssef, A.M.; Elhawary, R.; Zaitone, S.A.; Celia Rodríguez, P.; Segura-Carretero, A.; Mekky, R.H. Methanolic extracts of a selected Egyptian Vicia faba cultivar mitigate the oxidative/inflammatory burden and afford neuroprotection in a mouse model of Parkinson’s disease. Inflammopharmacology 2021, 29, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Li, T.; Ji, F.; Wang, H.; Pang, J. Effect of GABA on blood pressure and blood dynamics of anesthetic rats. Int. J. Clin. Exp. Med. 2015, 8, 14296–14302. [Google Scholar]
- Yang, R.; Hui, Q.; Gu, Z. Effects of ABA and CaCl2 on GABA accumulation in fava bean germinating under hypoxia-NaCl stress. Biosci. Biotechnol. Biochem. 2016, 80, 540–546. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Wang, S.; Yin, Y.; Gu, Z. Hypoxia treatment on germinating faba bean (Vicia faba L.) seeds enhances GABA-related protection against salt stress. Chil. J. Agric. Res. 2015, 75, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Yin, Y.; Gu, Z. Polyamine degradation pathway regulating growth and GABA accumulation in germinating fava bean under hypoxia-nacl stress. J. Agric. Sci. Technol. 2015, 17, 311–320. [Google Scholar]
- Yang, R.; Yin, Y.; Guo, L.; Han, Y.; Gu, Z. Sequence analysis of diamine oxidase gene from fava bean and its expression related to γ-aminobutyric acid accumulation in seeds germinating under hypoxia-NaCl stress. J. Sci. Food Agric. 2014, 94, 1585–1591. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bai, Q.; Jin, X.; Wen, H.; Gu, Z. Effects of cultivar and culture conditions on γ-aminobutyric acid accumulation in germinated fava beans (Vicia faba L.). J. Sci. Food Agric. 2010, 90, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Melama, L.; Rizzello, C.G.; Curiel, J.A.; Sibakov, J.; Holopainen, U.; Pulkkinen, M.; Sozer, N. Effect of air classification and fermentation by Lactobacillus plantarum VTT E-133328 on faba bean (Vicia faba L.) flour nutritional properties. Int. J. Food Microbiol. 2015, 193, 34–42. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Losito, I.; Facchini, L.; Katina, K.; Palmisano, F.; Gobbetti, M.; Coda, R. Degradation of vicine, convicine and their aglycones during fermentation of faba bean flour. Sci. Rep. 2016, 6, 32452. [Google Scholar] [CrossRef] [Green Version]
- Luzzatto, L.; Arese, P. Favism and Glucose-6-Phosphate Dehydrogenase Deficiency. N. Engl. J. Med. 2018, 378, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Nkhoma, E.T.; Poole, C.; Vannappagari, V.; Hall, S.A.; Beutler, E. The global prevalence of glucose-6-phosphate dehydrogenase deficiency: A systematic review and meta-analysis. Blood Cells Mol. Dis. 2009, 42, 267–278. [Google Scholar] [CrossRef]
- Hagag, A.A.; Badraia, I.M.; Elfarargy, M.S.; Elmageed, M.M.A.; Abo-Ali, E.A. Study of glucose-6-phosphate dehydrogenase deficiency: 5 years retrospective Egyptian study. Endocr. Metab. Immune Disord. Drug Targets 2018, 18, 155–162. [Google Scholar] [CrossRef]
- Khazaei, H.; Purves, R.W.; Song, M.; Stonehouse, R.; Bett, K.E.; Stoddard, F.L.; Vandenberg, A. Development and validation of a robust, breeder-friendly molecular marker for the vc- locus in faba bean. Mol. Breed. 2017, 37, 140. [Google Scholar] [CrossRef]
- Gallo, V.; Skorokhod, O.A.; Simula, L.F.; Marrocco, T.; Tambini, E.; Schwarzer, E.; Marget, P.; Duc, G.; Arese, P. No red blood cell damage and no hemolysis in G6PD-deficient subjects after ingestion of low vicine/convicine Vicia faba seeds. Blood 2018, 131, 1621–1625. [Google Scholar] [CrossRef] [Green Version]
- Getachew, F.; Vandenberg, A.; Smits, J. A practical toxicity bioassay for vicine and convicine levels in faba bean (Vicia faba). J. Sci. Food Agric. 2018, 98, 5105–5111. [Google Scholar] [CrossRef] [PubMed]
- Purves, R.W.; Khazaei, H.; Vandenberg, A. Toward a high-throughput method for determining vicine and convicine levels in faba bean seeds using flow injection analysis combined with tandem mass spectrometry. Food Chem. 2018, 256, 219–227. [Google Scholar] [CrossRef] [PubMed]
- El-Araby, M.M.; El-Shatoury, E.H.; Soliman, M.M.; Shaaban, H.F. Characterization and antimicrobial activity of lectins purified from three Egyptian leguminous seeds. AMB Express 2020, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Alatorre-Cruz, J.M.; Pita-López, W.; López-Reyes, R.G.; Ferriz-Martínez, R.A.; Cervantes-Jiménez, R.; de Jesús Guerrero Carrillo, M.; Vargas, P.J.A.; López-Herrera, G.; Rodríguez-Méndez, A.J.; Zamora-Arroyo, A.; et al. Effects of intragastrically-administered Tepary bean lectins on digestive and immune organs: Preclinical evaluation. Toxicol. Rep. 2018, 5, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Gendel, S.M. Comparison of international food allergen labeling regulations. Regul. Toxicol. Pharmacol. 2012, 63, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Smits, M.; Le, T.M.; Welsing, P.; Houben, G.; Knulst, A.; Verhoeckx, K. Legume protein consumption and the prevalence of legume sensitization. Nutrients 2018, 10, 1545. [Google Scholar] [CrossRef] [Green Version]
- Smits, M.; Verhoeckx, K.; Knulst, A.; Welsing, P.; de Jong, A.; Houben, G.; Le, T.M. Ranking of 10 legumes according to the prevalence of sensitization as a parameter to characterize allergenic proteins. Toxicol. Rep. 2021, 8, 767–773. [Google Scholar] [CrossRef]
- Mur Gimeno, P.; Feo Brito, F.; Martín Iglesias, A.; Lombardero Vega, M.; Bautista Martínez, P. Allergic reaction caused by a new hidden food, broad bean flour. Allergy Eur. J. Allergy Clin. Immunol. 2007, 62, 1340–1341. [Google Scholar] [CrossRef]
- Rodríguez-Mazariego, M.E.; Fuentes Aparicio, V.; Bartolomé Zavala, B.; Acevedo Matos, M.; Zapatero Remón, L. Broad bean-induced anaphylaxis: A case report. J. Investig. Allergol. Clin. Immunol. 2016, 26, 374–375. [Google Scholar] [CrossRef] [Green Version]
- Damiani, E.; Aloia, A.M.; Priore, M.G.; Pastore, A.; Nardulli, S.; Lippolis, C.; Macchia, L.; Ferrannini, A. Vicia faba Hypersensitivity and ASA Intolerance in a Farmer: A Case Report. J. Allergy 2011, 2011, 191787. [Google Scholar] [CrossRef] [Green Version]
- Remington, B.; Broekman, H.C.H.; Blom, W.M.; Capt, A.; Crevel, R.W.R.; Dimitrov, I.; Faeste, C.K.; Fernandez-Canton, R.; Giavi, S.; Houben, G.F.; et al. Approaches to assess IgE mediated allergy risks (sensitization and cross-reactivity) from new or modified dietary proteins. Food Chem. Toxicol. 2018, 112, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Kumar, S.; Verma, A.K.; Sharma, A.; Tripathi, A.; Chaudhari, B.P.; Kant, S.; Das, M.; Jain, S.K.; Dwivedi, P.D. Hypersensitivity linked to exposure of broad bean protein(s) in allergic patients and BALB/c mice. Nutrition 2014, 30, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.M. Current Concepts and Unresolved Questions in Dietary Protein Requirements and Supplements in Adults. Front. Nutr. 2017, 4, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariotti, F.; Gardner, D.C. Dietary Protein and Amino Acids in Vegetarian Diets—A Review. Nutrients 2019, 11, 2661. [Google Scholar] [CrossRef] [Green Version]
- Katz, D.L.; Doughty, K.N.; Geagan, K.; Jenkins, D.A.; Gardner, C.D. Perspective: The Public Health Case for Modernizing the Definition of Protein Quality. Adv. Nutr. 2019, 10, 755–764. [Google Scholar] [CrossRef]


{kind=link}
| Legume | Proteins | Carbohydrates | Ash | Fat | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TCH 1 | Starch | Amylose 2 | TDF 3 | IDF 4 | SDF 5 | |||||
| Faba bean | Mean | 27.6 | 66.0 | 40.0 | 34.0 | 12.9 | 15.1 | 1.4 | 3.4 | 1.4 |
| SD | 3.0 | 5.1 | 3.4 | 6.4 | 9.0 | 4.6 | 1.8 | 0.4 | 0.4 | |
| Min | 22.7 | 55.2 | 28.1 | 18.6 | 6.4 | 10.7 | 0.6 | 2.6 | 0.7 | |
| Max | 34.7 | 71.4 | 47.5 | 44.4 | 34.9 | 30.3 | 7.6 | 4.4 | 3.2 | |
| n1 6 | 106 | 57 | 46 | 24 | 17 | 18 | 18 | 94 | 80 | |
| n2 7 | 13 | 6 | 7 | 3 | 6 | 4 | 4 | 11 | 11 | |
| Pea | Mean | 23.4 | 63.5 | 44.9 | 29.6 | 14.7 | 11.0 | 2.5 | 3.0 | 1.6 |
| SD | 2.4 | 7.1 | 1.2 | 3.5 | 2.6 | 0.9 | 1.4 | 0.3 | 0.5 | |
| Min | 18.1 | 52.8 | 42.2 | 19.1 | 12.2 | 9.7 | 1.7 | 2.4 | 1.0 | |
| Max | 27.5 | 70.0 | 46.6 | 31.6 | 19.4 | 12.9 | 5.6 | 3.7 | 2.9 | |
| n1 | 34 | 5 | 18 | 12 | 11 | 8 | 8 | 23 | 23 | |
| n2 | 12 | 4 | 6 | 3 | 6 | 3 | 3 | 10 | 10 | |
| Soy | Mean | 40.0 | 28.6 | 2.7 | - | 21.9 | 24.8 | 2.6 | 5.2 | 19.7 |
| SD | 3.0 | 3.0 | 2.7 | - | 8.3 | 8.6 | 2.3 | 0.6 | 2.2 | |
| Min | 31.5 | 19.7 | 0.2 | - | 13.7 | 15.4 | 0.6 | 3.0 | 14.0 | |
| Max | 46.8 | 33.2 | 6.7 | - | 35.5 | 32.6 | 6.1 | 6.3 | 23.6 | |
| n1 | 48 | 31 | 19 | - | 9 | 5 | 5 | 40 | 60 | |
| n2 | 12 | 5 | 2 | - | 5 | 4 | 4 | 8 | 12 | |
| Faba Bean | Pea | Soy | Amino Acid Scoring Pattern 1 | ||
|---|---|---|---|---|---|
| cv. Bobas | cv. Kasztelan | cv. Solara | ND | ||
| (High tannin) | (Low tannin) | ||||
| Histidine * | 2.41 | 2.29 | 2.52 | 2.91 | 2 |
| Isoleucine * | 3.94 | 3.91 | 3.33 | 4.6 | 3.2 |
| Leucine * | 7.47 | 7.01 | 6.58 | 7.76 | 6.6 |
| Lysine * | 7.08 | 6.71 | 6.84 | 7.08 | 5.7 |
| Methionine * | 0.87 | 1.06 | 1.03 | 1.29 | 2.7 2 |
| Cysteine | 1.33 | 0.85 | 1.55 | 1.19 | |
| Phenylalanine * | 4.19 | 4.12 | 4.19 | 5.87 | 5.2 3 |
| Tyrosine | 2.78 | 2.59 | 3.16 | 3.65 | |
| Threonine * | 3.40 | 3.40 | 3.59 | 3.69 | 3.1 |
| Tryptophan * | 0.87 | 0.85 | 0.94 | 1.38 | 0.85 |
| Valine * | 4.31 | 4.12 | 3.89 | 4.64 | 4.3 |
| Arginine | 9.46 | 9.04 | 6.84 | 8.86 | . |
| Alanine | 4.15 | 4.03 | 4.27 | 4.39 | . |
| Aspartic acid | 10.74 | 10.4 | 10.68 | 11.98 | . |
| Glutamic acid | 16.51 | 16.26 | 16.92 | 17.88 | . |
| Glycine | 4.73 | 4.25 | 4.32 | 4.20 | . |
| Proline | 3.94 | 3.86 | 3.76 | 4.92 | . |
| Serine | 4.69 | 4.76 | 4.79 | 4.77 | . |
| References | [105] | [105] | [108] | [105] | [100] |
| Amino Acids Sequence | Bioactive Properties | Starting Material | Protein Hydrolysis Method | Protein Precursor | Hydrophobic Residue (%) | References | |
|---|---|---|---|---|---|---|---|
| 1 | GGQHQQEEESEEQK | Antioxidant (DPPH assay) Antibiofilm (biofilm inhibition of Pseudomonas aeruginosa PA14) | Faba bean protein isolate | Trypsin hydrolysis (18 h 37 °C) | Legumin | 0 | [74] |
| 2 | GPLVHPQSQSQSN | Antioxidant (DPPH assay) Antityrosinase (tyrosinase inhibition assay) | Legumin | 15 | |||
| 3 | LSPGDVLVIPAGYPVAIK | Antioxidant (DPPH, FRAP, and ferrous ion-chelating assays) Antibiofilm (biofilm inhibition of Pseudomonas aeruginosa PA14) | Vicilin | 56 | |||
| 4 | VESEAGLTETWNPNHPELR | Antioxidant (DPPH assay), Antityrosinase (tyrosinase inhibition assay) Antibiofilm (biofilm inhibition of Pseudomonas aeruginosa PA14) | Legumin | 26 | |||
| 5 | EEYDEEKEQGEEEIR | Antioxidant (DPPH assay) Antibiofilm (biofilm inhibition of Pseudomonas aeruginosa PA14) | Vicilin | 13 | |||
| 6 | ELAFPGSAQEVDTLLENQK | Fungicide | Lentil, pea and faba bean flours mixed (1:1:1) | Veron® PS (6 h 30 °C) | Vicilin | 36 | [142] |
| 7 | LSPGDVLVIPAGYPVAIK | Fungicide | Vicilin | 50 | |||
| 8 | SAQ | Promoting lactic acid bacteria growth | Faba bean protein isolate | Alcalase (1 h 37 °C) | ND | ND | [143] |
| 9 | Peptide enriched fraction | ||||||
| DALEPDNRIESEGGLIETWNPNNRQ | Antioxidant (ABTS assay) Antihypertensive (ACE inhibition) Anti-inflammatory (LOX inhibition) Reduction of lipids absorption (pancreatic lipase inhibition) | Fermented faba bean flour | In vitro simulated gastrointestinal digestion | Legumin | ND | [73] | |
| FEEPQQSEQGEGR | |||||||
| GSRQEEDEDEDE | |||||||
| WMNYNDQIPVINNQLDQMPR | |||||||
| RGEDEDDKEKRHSQKGES | |||||||
| RLNIGSSSSDIYNPQAGR | |||||||
| 10 | HLPSYSPSPQ | Promote muscle protein synthesis (increased phosphorylation S6 in skeletal muscle cells) | Faba bean protein powder | Food grade endopeptidase | ND | ND | [144] |
| 11 | TIKIPAGT | Anti-inflammatory (Reduced TNF-α in macrophages) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martineau-Côté, D.; Achouri, A.; Karboune, S.; L’Hocine, L. Faba Bean: An Untapped Source of Quality Plant Proteins and Bioactives. Nutrients 2022, 14, 1541. https://doi.org/10.3390/nu14081541
Martineau-Côté D, Achouri A, Karboune S, L’Hocine L. Faba Bean: An Untapped Source of Quality Plant Proteins and Bioactives. Nutrients. 2022; 14(8):1541. https://doi.org/10.3390/nu14081541
Chicago/Turabian StyleMartineau-Côté, Delphine, Allaoua Achouri, Salwa Karboune, and Lamia L’Hocine. 2022. "Faba Bean: An Untapped Source of Quality Plant Proteins and Bioactives" Nutrients 14, no. 8: 1541. https://doi.org/10.3390/nu14081541