
Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases Targeting Gut Microbiota: A Narrative Review


 ,
,
Abstract
:1. Introduction
2. Bioactive Effects of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases
2.1. Obesity
2.2. Type 2 and Gestational Diabetes Mellitus
2.3. Other Metabolic Diseases
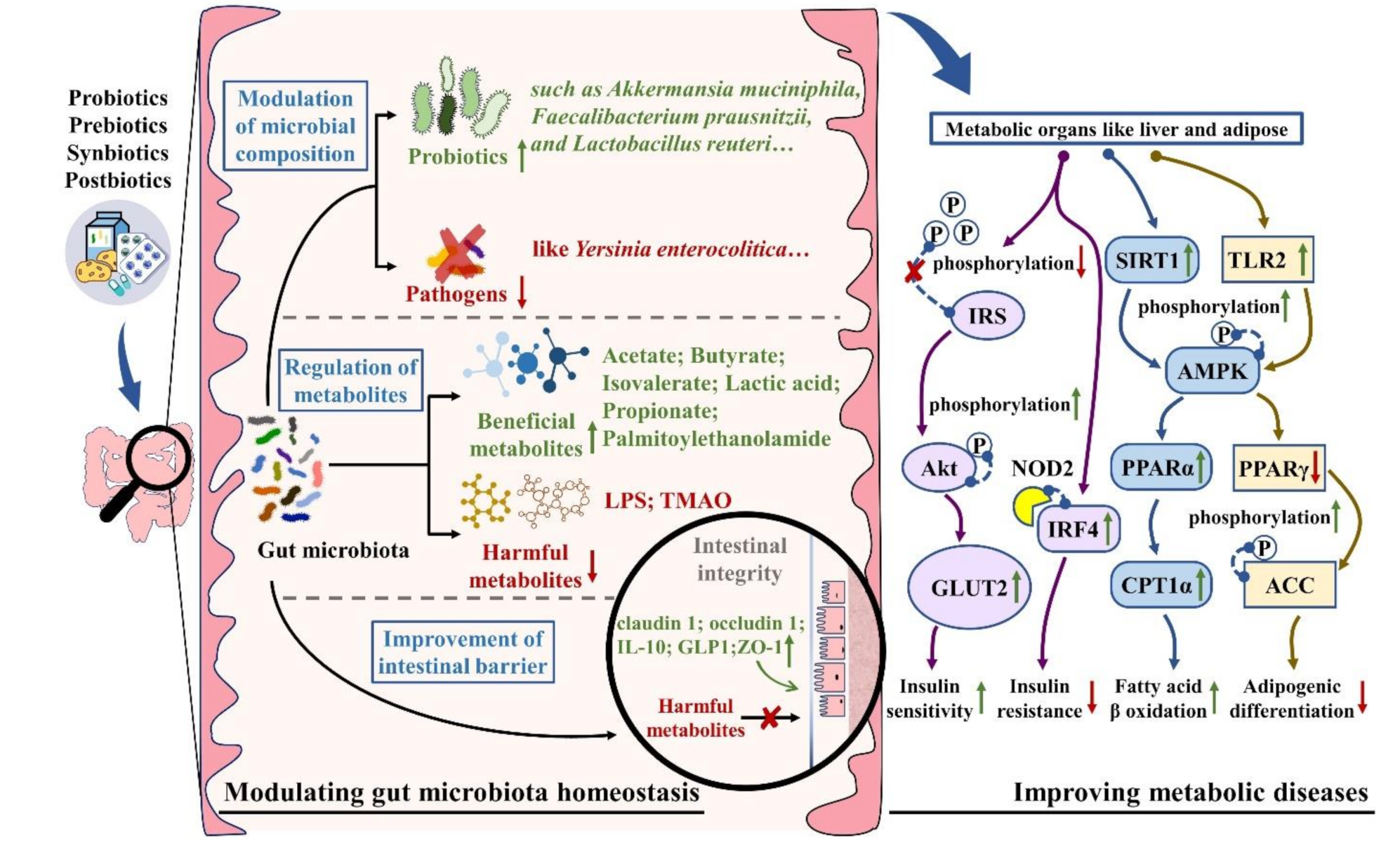
3. Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases by Targeting Gut Microbiota
3.1. The Modulation of Gut Microbiota Composition
3.2. The Regulation of Gut Microbial Metabolites
3.3. The Improvement of Intestinal Barrier Function
4. Clinical Effects of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases
4.1. Obesity
4.2. Type 2 and Gestational Diabetes Mellitus
4.3. Other Metabolic Diseases
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gowd, V.; Karim, N.; Shishir, M.R.I.; Xie, L.H.; Chen, W. Dietary polyphenols to combat the metabolic diseases via altering gut microbiota. Trends Food Sci. Technol. 2019, 93, 81–93. [Google Scholar] [CrossRef]
- Li, H.Y.; Gan, R.Y.; Shang, A.; Mao, Q.Q.; Sun, Q.C.; Wu, D.T.; Geng, F.; He, X.Q.; Li, H.B. Plant-Based Foods and Their Bioactive Compounds on Fatty Liver Disease: Effects, Mechanisms, and Clinical Application. Oxidative Med. Cell Longev. 2021, 2021, 6621644. [Google Scholar] [CrossRef]
- Wu, T.R.; Lin, C.S.; Chang, C.J.; Lin, T.L.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Lu, C.C.; Young, J.D.; Lai, H.C. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut 2019, 68, 248–262. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Li, S.; Gan, R.Y.; Zhou, T.; Xu, D.P.; Li, H.B. Impacts of Gut Bacteria on Human Health and Diseases. Int. J. Mol. Sci. 2015, 16, 7493–7519. [Google Scholar] [CrossRef]
- Stewart, C.J.; Embleton, N.D.; Marrs, E.C.; Smith, D.P.; Nelson, A.; Abdulkadir, B.; Skeath, T.; Petrosino, J.F.; Perry, J.D.; Berrington, J.E.; et al. Temporal bacterial and metabolic development of the preterm gut reveals specific signatures in health and disease. Microbiome 2016, 4, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Chassaing, B.; Singh, V.; Pellizzon, M.; Ricci, M.; Fythe, M.D.; Kumar, M.V.; Gewirtz, A.T. Fiber-mediated nourishment of gut microbiota protects against diet-induced obesity by restoring IL-22-mediated colonic health. Cell Host Microbe 2018, 23, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Regnier, M.; Van Hul, M.; Knauf, C.; Cani, P.D. Gut microbiome, endocrine control of gut barrier function and metabolic diseases. J. Endocrinol. 2021, 248, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Holmes, Z.C.; Silverman, J.D.; Dressman, H.K.; Wei, Z.; Dallow, E.P.; Armstrong, S.C.; Seed, P.C.; Rawls, J.F.; David, L.A. Short-chain fatty acid production by gut microbiota from children with obesity differs according to prebiotic choice and bacterial community composition. MBio 2020, 11, e00914-20. [Google Scholar] [CrossRef]
- Jansma, J.; Brinkman, F.; van Hemert, S.; El Aidy, S. Targeting the endocannabinoid system with microbial interventions to improve gut integrity. Prog. Neuro-Psychopharmacol. Biol. 2021, 106, 110169. [Google Scholar] [CrossRef] [PubMed]
- da Silva, T.F.; Casarotti, S.N.; de Oliveira, G.L.V.; Penna, A.L.B. The impact of probiotics, prebiotics, and synbiotics on the biochemical, clinical, and immunological markers, as well as on the gut microbiota of obese hosts. Crit. Rev. Food Sci. Nutr. 2021, 61, 337–355. [Google Scholar] [CrossRef]
- Vallianou, N.; Stratigou, T.; Christodoulatos, G.S.; Tsigalou, C.; Dalamaga, M. Probiotics, prebiotics, synbiotics, postbiotics, and obesity: Current evidence, controversies, and perspectives. Curr. Obes. Rep. 2020, 9, 179–192. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, in press. [Google Scholar] [CrossRef]
- Wu, J.Y.; Wang, K.; Wang, X.M.; Pang, Y.L.; Jiang, C.T. The role of the gut microbiome and its metabolites in metabolic diseases. Protein Cell 2021, 12, 360–373. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Thonusin, C.; Chattipakorn, N.; Chattipakorn, S.C. Impacts of gut microbiota on gestational diabetes mellitus: A comprehensive review. Eur. J. Nutr. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Walia, S.; Kondepudi, K.K.; Shukla, G. Administration of indigenous probiotics modulate high-fat diet-induced metabolic syndrome in Sprague Dawley rats. Antonie Van Leeuwenhoek 2020, 113, 1345–1359. [Google Scholar] [CrossRef]
- Kobyliak, N.; Falalyeyeva, T.; Beregova, T.; Spivak, M. Probiotics for experimental obesity prevention: Focus on strain dependence and viability of composition. Endokrynol. Pol. 2017, 68, 659–667. [Google Scholar] [CrossRef]
- Celik, M.N.; Unlu Sogut, M. Probiotics improve chemerin levels and metabolic syndrome parameters in obese rats. Balk Med. J. 2019, 36, 270–275. [Google Scholar] [CrossRef]
- Kobyliak, N.; Falalyeyeva, T.; Tsyryuk, O.; Eslami, M.; Kyriienko, D.; Beregova, T.; Ostapchenko, L. New insights on strain-specific impacts of probiotics on insulin resistance: Evidence from animal study. J. Diabetes Metab. Disord. 2020, 19, 289–296. [Google Scholar] [CrossRef]
- Luo, Q.; Zhang, J.R.; Li, H.B.; Wu, D.T.; Geng, F.; Corke, H.; Wei, X.L.; Gan, R.Y. Green Extraction of Antioxidant Polyphenols from Green Tea (Camellia sinensis). Antioxidants 2020, 9, 785. [Google Scholar] [CrossRef]
- Cao, S.Y.; Li, B.Y.; Gan, R.Y.; Mao, Q.Q.; Wang, Y.F.; Shang, A.; Meng, J.M.; Xu, X.Y.; Wei, X.L.; Li, H.B. The in vivo antioxidant and hepatoprotective actions of selected Chinese teas. Foods 2020, 9, 262. [Google Scholar] [CrossRef] [Green Version]
- Vo, T.D.; Lynch, B.S.; Roberts, A. Dietary exposures to common emulsifiers and their impact on the gut microbiota: Is there a cause for concern? Compr. Rev. Food Sci. Food Saf. 2019, 18, 31–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, A.; Luo, M.; Gan, R.Y.; Xu, X.Y.; Xia, Y.; Guo, H.; Liu, Y.; Li, H.B. Effects of Microwave-Assisted Extraction Conditions on Antioxidant Capacity of Sweet Tea (Lithocarpus polystachyus Rehd.). Antioxidants 2020, 9, 678. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.S.; Lee, B.H.; Shin, D.U.; Lim, M.Y.; Chung, W.H.; Park, C.S.; Baik, M.Y.; Nam, Y.D.; Seo, D.H. Amelioration of obesity in high-fat diet-fed mice by chestnut starch modified by amylosucrase from Deinococcus geothermalis. Food Hydrocoll. 2018, 75, 22–32. [Google Scholar] [CrossRef]
- Amabebe, E.; Robert, F.O.; Agbalalah, T.; Orubu, E.S.F. Microbial dysbiosis-induced obesity: Role of gut microbiota in homoeostasis of energy metabolism. Br. J. Nutr. 2020, 123, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.T.; Parajuli, N.; Sung, M.M.; Bairwa, S.C.; Levasseur, J.; Soltys, C.-L.M.; Wishart, D.S.; Madsen, K.; Schertzer, J.D.; Dyck, J.R.B. Fecal transplant from resveratrol-fed donors improves glycaemia and cardiovascular features of the metabolic syndrome in mice. Am. J. Physiol.-Endocrinol. Metab. 2018, 315, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, N.; Bauman, C.; Rodriguez Ayala, F.; Grau, R. Two cases of type 2 diabetes mellitus successfully treated with probiotics. Clin. Case Rep. 2020, 8, 3120–3125. [Google Scholar] [CrossRef]
- Li, B.Y.; Xu, X.Y.; Gan, R.Y.; Sun, Q.C.; Meng, J.M.; Shang, A.; Mao, Q.Q.; Li, H.B. Targeting Gut Microbiota for the Prevention and Management of Diabetes Mellitus by Dietary Natural Products. Foods 2019, 8, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, J.; Jang, J.; Park, K. Effects of consuming calcium-rich foods on the incidence of type 2 diabetes mellitus. Nutrients 2018, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Gijsbers, L.; Ding, E.L.; Malik, V.S.; de Goede, J.; Geleijnse, J.M.; Soedamah-Muthu, S.S. Consumption of dairy foods and diabetes incidence: A dose-response meta-analysis of observational studies. Am. J. Clin. Nutr. 2016, 103, 1111–1124. [Google Scholar] [CrossRef] [Green Version]
- Ban, Q.; Cheng, J.; Sun, X.; Jiang, Y.; Guo, M. Effect of feeding type 2 diabetes mellitus rats with synbiotic yogurt sweetened with monk fruit extract on serum lipid levels and hepatic AMPK (5’ adenosine monophosphate-activated protein kinase) signaling pathway. Food Funct. 2020, 11, 7696–7706. [Google Scholar] [CrossRef]
- Gao, H.; Wen, J.J.; Hu, J.L.; Nie, Q.X.; Chen, H.H.; Xiong, T.; Nie, S.P.; Xie, M.Y. Fermented Momordica charantia L. juice modulates hyperglycemia, lipid profile, and gut microbiota in type 2 diabetic rats. Food Res. Int. 2019, 121, 367–378. [Google Scholar] [CrossRef]
- Tang, R.; Li, L. Modulation of short-chain fatty acids as potential therapy method for type 2 diabetes mellitus. Can. J. Infect. Dis. Med. Microbiol. 2021, 2021, 6632266. [Google Scholar] [CrossRef]
- Bahmani, F.; Tajadadi-Ebrahimi, M.; Kolahdooz, F.; Mazouchi, M.; Hadaegh, H.; Jamal, A.S.; Mazroii, N.; Asemi, S.; Asemi, Z. The consumption of synbiotic bread containing Lactobacillus sporogenes and inulin affects nitric oxide and malondialdehyde in patients with type 2 diabetes mellitus: Randomized, double-blind, placebo-controlled trial. J. Am. Coll. Nutr. 2016, 35, 506–513. [Google Scholar] [CrossRef]
- Farrokhian, A.; Raygan, F.; Soltani, A.; Tajabadi-Ebrahimi, M.; Sharifi Esfahani, M.; Karami, A.A.; Asemi, Z. The effects of synbiotic supplementation on carotid intima-media thickness, biomarkers of inflammation, and oxidative stress in people with overweight, diabetes, and coronary heart disease: A randomized, double-blind, placebo-controlled trial. Probiotics Antimicrob Proteins 2019, 11, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Pellonpera, O.; Mokkala, K.; Houttu, N.; Vahlberg, T.; Koivuniemi, E.; Tertti, K.; Ronnemaa, T.; Laitinen, K. Efficacy of fish oil and/or probiotic intervention on the incidence of gestational diabetes mellitus in an at-risk group of overweight and obese women: A randomized, placebo-controlled, double-blind clinical trial. Diabetes Care 2019, 42, 1009–1017. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Li, B.Y.; Meng, J.M.; Gan, R.Y.; Xu, X.Y.; Gu, Y.Y.; Wang, X.H.; Li, H.B. Effects of several tea extracts on nonalcoholic fatty liver disease in mice fed with a high-fat diet. Food Sci. Nutr. 2021, 9, 2954–2967. [Google Scholar] [CrossRef]
- Syauqy, A.; Hsu, C.Y.; Rau, H.H.; Chao, J.C.J. Association of dietary patterns with components of metabolic syndrome and inflammation among middle-aged and older adults with metabolic syndrome in Taiwan. Nutrients 2018, 10, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hann, M.; Zeng, Y.; Zong, L.; Sakurai, T.; Taniguchi, Y.; Takagaki, R.; Watanabe, H.; Mitsuzumi, H.; Mine, Y. Anti-inflammatory activity of isomaltodextrin in a C57bl/6NCrl mouse model with lipopolysaccharide-induced low-grade chronic inflammation. Nutrients 2019, 11, 2791. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Wan, Z.; Ou, A.; Liang, X.; Guo, X.; Zhang, Z.; Wu, L.; Xue, X. Monofloral honey from a medical plant, Prunella Vulgaris, protected against dextran sulfate sodium-induced ulcerative colitis via modulating gut microbial populations in rats. Food Funct. 2019, 10, 3828–3838. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Jin, X.; Li, Q.; Sawaya, A.C.H.F.; Le Leu, R.K.; Conlon, M.A.; Wu, L.; Hu, F. Propolis from Different Geographic Origins Decreases Intestinal Inflammation and Bacteroides spp. Populations in a Model of DSS-Induced Colitis. Mol. Nutr. Food Res. 2018, 62, e1800080. [Google Scholar] [CrossRef]
- Wu, T.; Shen, M.; Guo, X.; Huang, L.; Yang, J.; Yu, Q.; Chen, Y.; Xie, J. Cyclocarya paliurus polysaccharide alleviates liver inflammation in mice via beneficial regulation of gut microbiota and TLR4/MAPK signaling pathways. Int. J. Biol. Macromol. 2020, 160, 164–174. [Google Scholar] [CrossRef]
- Li, S.; Wang, L.; Liu, B.; He, N. Unsaturated alginate oligosaccharides attenuated obesity-related metabolic abnormalities by modulating gut microbiota in high-fat-diet mice. Food Funct. 2020, 11, 4773–4784. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, P.; Cen, Y.; Li, W. Effects of preoperative oral administration of glucose solution combined with postoperative probiotics on inflammation and intestinal barrier function in patients after colorectal cancer surgery. Oncol. Lett. 2019, 18, 694–698. [Google Scholar] [CrossRef] [Green Version]
- Asad, F.; Anwar, H.; Yassine, H.M.; Ullah, M.I.; Rahman, A.; Kamran, Z.; Sohail, M.U. White button mushroom, Agaricus bisporus (Agaricomycetes), and a probiotics mixture supplementation correct dyslipidemia without influencing the colon microbiome profile in hypercholesterolemic rats. Int. J. Med. Mushrooms 2020, 22, 235–244. [Google Scholar] [CrossRef]
- Xavier-Santos, D.; Lima, E.D.; Simao, A.N.C.; Bedani, R.; Saad, S.M.I. Effect of the consumption of a synbiotic diet mousse containing Lactobacillus acidophilus La-5 by individuals with metabolic syndrome: A randomized controlled trial. J. Funct. Foods 2018, 41, 55–61. [Google Scholar] [CrossRef]
- Cornejo-Pareja, I.; Munoz-Garach, A.; Clemente-Postigo, M.; Tinahones, F.J. Importance of gut microbiota in obesity. Eur. J. Clin. Nutr. 2019, 72, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Alard, J.; Lehrter, V.; Rhimi, M.; Mangin, I.; Peucelle, V.; Abraham, A.L.; Mariadassou, M.; Maguin, E.; Waligora-Dupriet, A.J.; Pot, B.; et al. Beneficial metabolic effects of selected probiotics on diet-induced obesity and insulin resistance in mice are associated with improvement of dysbiotic gut microbiota. Environ. Microbiol. 2016, 18, 1484–1497. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Zheng, C.; Zheng, J.H.; Xu, S.C. The relationship between intestinal flora changes and osteoporosis in rats with inflammatory bowel disease and the improvement effect of probiotics. Eur. Rev. Med. Pharmaco. 2020, 24, 5697–5702. [Google Scholar] [CrossRef]
- Kamdar, K.; Khakpour, S.; Chen, J.; Leone, V.; Brulc, J.; Mangatu, T.; Antonopoulos, D.A.; Chang, E.B.; Kahn, S.A.; Kirschner, B.S.; et al. Genetic and metabolic signals during acute enteric bacterial infection alter the microbiota and drive progression to chronic inflammatory disease. Cell Host Microbe 2016, 19, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheithauer, T.P.M.; Rampanelli, E.; Nieuwdorp, M.; Vallance, B.A.; Verchere, C.B.; van Raalte, D.H.; Herrema, H. Gut Microbiota as a Trigger for Metabolic Inflammation in Obesity and Type 2 Diabetes. Front. Immunol. 2020, 11, 571731. [Google Scholar] [CrossRef]
- Hsu, C.N.; Hou, C.Y.; Chan, J.Y.H.; Lee, C.T.; Tain, Y.L. Hypertension programmed by perinatal high-fat diet: Effect of maternal gut microbiota-targeted therapy. Nutrients 2019, 11, 2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, S.; Singh, T.P.; Malik, R.K. In vivo implications of potential probiotic Lactobacillus reuteri lr6 on the gut and immunological parameters as an adjuvant against protein energy malnutrition. Probiotics Antimicrob. Proteins 2020, 12, 517–534. [Google Scholar] [CrossRef]
- Barczynska, R.; Kapusniak, J.; Litwin, M.; Slizewska, K.; Szalecki, M. Dextrins from maize starch as substances activating the growth of Bacteroidetes and Actinobacteria simultaneously inhibiting the growth of firmicutes, responsible for the occurrence of obesity. Plant. Food Hum. Nutr. 2016, 71, 190–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siva, N.; Johnson, C.R.; Richard, V.; Jesch, E.D.; Whiteside, W.; Abood, A.A.; Thavarajah, P.; Duckett, S.; Thavarajah, D. Lentil (Lens culinaris Medikus) diet affects the gut microbiome and obesity markers in rat. J. Agric. Food Chem. 2018, 66, 8805–8813. [Google Scholar] [CrossRef]
- Panichsillaphakit, E.; Chongpison, Y.; Saengpanit, P.; Kwanbunbumpen, T.; Uaariyapanichkul, J.; Chomtho, S.; Pancharoen, C.; Visuthranukul, C. Children’s eating behavior questionnaire correlated with body compositions of thai children and adolescents with obesity: A pilot study. J. Nutr. Biochem. 2021, 2021, 6496134. [Google Scholar] [CrossRef]
- Hu, F.; Niu, Y.; Xu, X.; Hu, Q.; Su, Q.; Zhang, H. Resistant dextrin improves high-fat-high-fructose diet induced insulin resistance. Nutr. Metab. 2020, 17, 36. [Google Scholar] [CrossRef]
- Chen, K.; Xie, K.; Liu, Z.; Nakasone, Y.; Sakao, K.; Hossain, A.; Hou, D.X. Preventive effects and mechanisms of garlic on dyslipidemia and gut microbiome dysbiosis. Nutrients 2019, 11, 1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.A.; Ward, L.C.; Brown, L. Inulin oligofructose attenuates metabolic syndrome in high-carbohydrate, high-fat diet-fed rats. Br. J. Nutr. 2016, 116, 1502–1511. [Google Scholar] [CrossRef] [Green Version]
- de Cossio, L.F.; Fourrier, C.; Sauvant, J.; Everard, A.; Capuron, L.; Cani, P.D.; Laye, S.; Castanon, N. Impact of prebiotics on metabolic and behavioral alterations in a mouse model of metabolic syndrome. Brain Behav. Immun. 2017, 64, 33–49. [Google Scholar] [CrossRef]
- Klancic, T.; Laforest-Lapointe, I.; Choo, A.; Nettleton, J.E.; Chleilat, F.; Noye Tuplin, E.W.; Alukic, E.; Cho, N.A.; Nicolucci, A.C.; Arrieta, M.C.; et al. Prebiotic oligofructose prevents antibiotic-induced obesity risk and improves metabolic and gut microbiota profiles in rat dams and offspring. Mol. Nutr. Food Res. 2020, 64, e2000288. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, M.; Zhang, P.; Fan, S.; Huang, J.; Yu, S.; Zhang, C.; Li, H. Fucoidan and galactooligosaccharides ameliorate high-fat diet-induced dyslipidemia in rats by modulating the gut microbiota and bile acid metabolism. Nutrition 2019, 65, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Nagpal, R.; Wang, S.; Gagliano, J.; Kitzman, D.W.; Soleimanian-Zad, S.; Sheikh-Zeinoddin, M.; Read, R.; Yadav, H. Prebiotics from acorn and sago prevent high-fat-diet-induced insulin resistance via microbiome-gut-brain axis modulation. J. Nutr. Biochem. 2019, 67, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Falalyeyeva, T.; Bodnar, P.; Beregova, T. Probiotics supplemented with omega-3 fatty acids are more effective for hepatic steatosis reduction in an animal model of obesity. Probiotics Antimicrob. Proteins 2017, 9, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Falalyeyeva, T.; Boyko, N.; Tsyryuk, O.; Beregova, T.; Ostapchenko, L. Probiotics and nutraceuticals as a new frontier in obesity prevention and management. Diabetes Res. Clin. Pract. 2018, 141, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, M.; Liu, H.; Wei, X.; Su, X.; Li, M.; Yuan, J. Oral supplements of combined Bacillus licheniformis Zhengchangsheng(R) and Xylooligosaccharides improve high-fat diet-induced obesity and modulate the gut microbiota in rats. Biomed. Res. Int. 2020, 2020, 9067821. [Google Scholar] [CrossRef]
- Oh, Y.J.; Kim, H.J.; Kim, T.S.; Yeo, I.H.; Ji, G.E. Effects of Lactobacillus plantarum PMO 08 alone and combined with chia seeds on metabolic syndrome and parameters related to gut health in high-fat diet-induced obese mice. J. Med. Food 2019, 22, 1199–1207. [Google Scholar] [CrossRef]
- Jiminez, J.A.; Uwiera, T.C.; Abbott, D.W.; Uwiera, R.R.E.; Inglis, G.D. Butyrate supplementation at high concentrations alters enteric bacterial communities and reduces intestinal inflammation in mice infected with citrobacter rodentium. mSphere 2017, 2, e00243-00217. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Pan, Q.; Xin, F.Z.; Zhang, R.N.; He, C.X.; Chen, G.Y.; Liu, C.; Chen, Y.W.; Fan, J.G. Sodium butyrate attenuates high-fat diet-induced steatohepatitis in mice by improving gut microbiota and gastrointestinal barrier. World J. Gastroenterol. 2017, 23, 60–75. [Google Scholar] [CrossRef]
- Rizzetto, L.; Fava, F.; Tuohy, K.M.; Selmi, C. Connecting the immune system, systemic chronic inflammation and the gut microbiome: The role of sex. J. Autoimmun. 2018, 92, 12–34. [Google Scholar] [CrossRef] [PubMed]
- Alard, J.; Cudennec, B.; Boutillier, D.; Peucelle, V.; Descat, A.; Decoin, R.; Kuylle, S.; Jablaoui, A.; Rhimi, M.; Wolowczuk, I.; et al. Multiple selection criteria for probiotic strains with high potential for obesity management. Nutrients 2021, 13, 713. [Google Scholar] [CrossRef]
- Kim, D.E.; Kim, J.K.; Han, S.K.; Jang, S.E.; Han, M.J.; Kim, D.H. Lactobacillus plantarum NK3 and Bifidobacterium longum NK49 Alleviate Bacterial Vaginosis and Osteoporosis in Mice by Suppressing NF-kappaB-Linked TNF-alpha Expression. J. Med. Food 2019, 22, 1022–1031. [Google Scholar] [CrossRef]
- Archer, A.C.; Muthukumar, S.P.; Halami, P.M. Lactobacillus fermentum MCC2759 and MCC2760 alleviate inflammation and intestinal function in high-fat diet-fed and streptozotocin-induced diabetic rats. Probiotics Antimicrob. Proteins 2021, in press. [Google Scholar] [CrossRef]
- Robles-Vera, I.; Toral, M.; de la Visitacion, N.; Sanchez, M.; Gomez-Guzman, M.; Romero, M.; Yang, T.; Izquierdo-Garcia, J.L.; Jimenez, R.; Ruiz-Cabello, J.; et al. Probiotics prevent dysbiosis and the rise in blood pressure in genetic hypertension: Role of short-chain fatty acids. Mol. Nutr. Food Res. 2020, 64, e1900616. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Pesce, M.; Seguella, L.; Lu, J.; Corpetti, C.; Del Re, A.; De Palma, F.D.E.; Esposito, G.; Sanseverino, W.; Sarnelli, G. Engineered Lactobacillus paracasei producing palmitoylethanolamide (PEA) prevents colitis in mice. Int. J. Mol. Sci. 2021, 22, 2945. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, M.; Morimoto, M.; Suto, A.; Nakatani, A.; Hayakawa, T.; Hara, K.; Kimura, I. Synthetic dietary inulin, Fuji FF, delays development of diet-induced obesity by improving gut microbiota profiles and increasing short-chain fatty acid production. PeerJ 2020, 8, e8893. [Google Scholar] [CrossRef] [Green Version]
- Mistry, R.H.; Liu, F.; Borewicz, K.; Lohuis, M.A.M.; Smidt, H.; Verkade, H.J.; Tietge, U.J.F. Long-term beta-galacto-oligosaccharides supplementation decreases the development of obesity and insulin resistance in mice fed a western-type diet. Mol. Nutr. Food Res. 2020, 64, e1900922. [Google Scholar] [CrossRef]
- Kundi, Z.M.; Lee, J.C.; Pihlajamaki, J.; Chan, C.B.; Leung, K.S.; So, S.S.Y.; Nordlund, E.; Kolehmainen, M.; El-Nezami, H. Dietary fiber from oat and rye brans ameliorate western diet-induced body weight gain and hepatic inflammation by the modulation of short-chain fatty acids, bile acids, and tryptophan metabolism. Mol. Nutr. Food Res. 2021, 65, e1900580. [Google Scholar] [CrossRef]
- Regalado-Renteria, E.; Aguirre-Rivera, J.R.; Godinez-Hernandez, C.I.; Garcia-Lopez, J.C.; Oros-Ovalle, A.C.; Martinez-Gutierrez, F.; Martinez-Martinez, M.; Ratering, S.; Schnell, S.; Ruiz-Cabrera, M.A.; et al. Effects of agave fructans, inulin, and starch on metabolic syndrome aspects in healthy wistar rats. ACS Omega 2020, 5, 10740–10749. [Google Scholar] [CrossRef]
- Gu, Z.; Zhu, Y.; Jiang, S.; Xia, G.; Li, C.; Zhang, X.; Zhang, J.; Shen, X. Tilapia head glycolipids reduce inflammation by regulating the gut microbiota in dextran sulphate sodium-induced colitis mice. Food Funct. 2020, 11, 3245–3255. [Google Scholar] [CrossRef]
- Ke, X.; Walker, A.; Haange, S.B.; Lagkouvardos, I.; Liu, Y.; Schmitt-Kopplin, P.; von Bergen, M.; Jehmlich, N.; He, X.; Clavel, T.; et al. Synbiotic-driven improvement of metabolic disturbances is associated with changes in the gut microbiome in diet-induced obese mice. Mol. Metab. 2019, 22, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Thiennimitr, P.; Yasom, S.; Tunapong, W.; Chunchai, T.; Wanchai, K.; Pongchaidecha, A.; Lungkaphin, A.; Sirilun, S.; Chaiyasut, C.; Chattipakorn, N.; et al. Lactobacillus paracasei HII01, xylooligosaccharides, and synbiotics reduce gut disturbance in obese rats. Nutrition 2018, 54, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Yao, F.; Jia, R.; Huang, H.; Yu, Y.; Mei, L.; Bai, L.; Ding, Y.; Zheng, P. Effect of Lactobacillus paracasei N1115 and fructooligosaccharides in nonalcoholic fatty liver disease. Arch. Med. Sci. 2019, 15, 1336–1344. [Google Scholar] [CrossRef]
- Wang, W.; Xu, A.L.; Li, Z.C.; Li, Y.; Xu, S.F.; Sang, H.C.; Zhi, F. Combination of probiotics and Salvia miltiorrhiza polysaccharide alleviates hepatic steatosis via gut microbiota modulation and insulin resistance improvement in high fat-induced NAFLD mice. Diabetes Metab. J. 2020, 44, 336–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Sun, J.; Chen, X.; Nie, C.; Zhao, J.; Guan, W.; Lei, L.; He, T.; Chen, Y.; Johnston, L.J.; et al. Combination of Clostridium butyricum and corn bran optimized intestinal microbial fermentation using a weaned pig model. Front. Microbiol. 2018, 9, 3091. [Google Scholar] [CrossRef]
- Lee, J.; Park, S.; Oh, N.; Park, J.; Kwon, M.; Seo, J.; Roh, S. Oral intake of Lactobacillus plantarum L-14 extract alleviates TLR2- and AMPK-mediated obesity-associated disorders in high-fat-diet-induced obese C57BL/6J mice. Cell Prolif. 2021, 54, e13039. [Google Scholar] [CrossRef]
- Cavallari, J.F.; Fullerton, M.D.; Duggan, B.M.; Foley, K.P.; Denou, E.; Smith, B.K.; Desjardins, E.M.; Henriksbo, B.D.; Kim, K.J.; Tuinema, B.R.; et al. Muramyl dipeptide-based postbiotics mitigate obesity-induced insulin resistance via IRF4. Cell Metab. 2017, 25, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Cavallari, J.F.; Barra, N.G.; Foley, K.P.; Lee, A.; Duggan, B.M.; Henriksbo, B.D.; Anhe, F.F.; Ashkar, A.A.; Schertzer, J.D. Postbiotics for NOD2 require nonhematopoietic RIPK2 to improve blood glucose and metabolic inflammation in mice. Am. J. Physiol.-Endocrinol. Metab. 2020, 318, 579–585. [Google Scholar] [CrossRef]
- Cheng, D.; Xu, J.H.; Li, J.Y.; Wang, S.Y.; Wu, T.F.; Chen, Q.K.; Yu, T. Butyrate ameliorated-NLRC3 protects the intestinal barrier in a GPR43-dependent manner. Exp. Cell Res. 2018, 368, 101–110. [Google Scholar] [CrossRef]
- Matheus, V.A.; Monteiro, L.; Oliveira, R.B.; Maschio, D.A.; Collares-Buzato, C.B. Butyrate reduces high-fat diet-induced metabolic alterations, hepatic steatosis and pancreatic beta cell and intestinal barrier dysfunctions in prediabetic mice. Exp. Biol. Med. 2017, 242, 1214–1226. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Lv, L.; Wu, W.; Li, Y.; Shi, D.; Fang, D.; Guo, F.; Jiang, H.; Yan, R.; Ye, W.; et al. Butyrate protects mice against methionine-choline-deficient diet-induced non-alcoholic steatohepatitis by improving gut barrier function, attenuating inflammation and reducing endotoxin levels. Front. Microbiol. 2018, 9, 1967. [Google Scholar] [CrossRef] [Green Version]
- Isozaki, S.; Konishi, H.; Fujiya, M.; Tanaka, H.; Murakami, Y.; Kashima, S.; Ando, K.; Ueno, N.; Moriichi, K.; Okumura, T. Probiotic-derived polyphosphate accelerates intestinal epithelia wound healing through inducing platelet-derived mediators. Mediat. Inflamm. 2021, 2021, 5582943. [Google Scholar] [CrossRef]
- Wicinski, M.; Gebalski, J.; Golebiewski, J.; Malinowski, B. Probiotics for the treatment of overweight and obesity in humans-a review of clinical trials. Microorganisms 2020, 8, 1148. [Google Scholar] [CrossRef]
- Overby, H.B.; Ferguson, J.F. Gut microbiota-derived short-chain fatty acids facilitate microbiota: Host cross talk and modulate obesity and hypertension. Curr. Hypertens. Rep. 2021, 23, 8. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, L.; Ma, S.; Ye, J.; Zhang, H.; Li, Y.; Sair, A.T.; Pan, J.; Liu, X.; Li, X.; et al. High-dietary fiber intake alleviates antenatal obesity-induced postpartum depression: Roles of gut microbiota and microbial metabolite short-chain fatty acid involved. J. Agric. Food Chem. 2020, 68, 13697–13710. [Google Scholar] [CrossRef]
- Wilkins, A.T.; Reimer, R.A. Obesity, early life gut microbiota, and antibiotics. Microorganisms 2021, 9, 413. [Google Scholar] [CrossRef]
- Ferrarese, R.; Ceresola, E.R.; Preti, A.; Canducci, F. Probiotics, prebiotics and synbiotics for weight loss and metabolic syndrome in the microbiome era. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7588–7605. [Google Scholar] [CrossRef] [PubMed]
- Saez-Lara, M.J.; Robles-Sanchez, C.; Ruiz-Ojeda, F.J.; Plaza-Diaz, J.; Gil, A. Effects of probiotics and synbiotics on obesity, insulin resistance syndrome, type 2 diabetes and non-alcoholic fatty liver disease: A review of human clinical trials. Int. J. Mol. Sci. 2016, 17, 928. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Luo, X.; Tang, J.; Mo, Q.; Zhong, H.; Zhang, H.; Feng, F. A bridge for short-chain fatty acids to affect inflammatory bowel disease, type 1 diabetes, and non-alcoholic fatty liver disease positively: By changing gut barrier. Eur. J. Nutr. 2021, in press. [Google Scholar] [CrossRef]
- Bernini, L.J.; Simao, A.N.; Alfieri, D.F.; Lozovoy, M.A.; Mari, N.L.; de Souza, C.H.; Dichi, I.; Costa, G.N. Beneficial effects of Bifidobacterium lactis on lipid profile and cytokines in patients with metabolic syndrome: A randomized trial. Effects of probiotics on metabolic syndrome. Nutrition 2016, 32, 716–719. [Google Scholar] [CrossRef]
- De Lorenzo, A.; Costacurta, M.; Merra, G.; Gualtieri, P.; Cioccoloni, G.; Marchetti, M.; Varvaras, D.; Docimo, R.; Di Renzo, L. Can psychobiotics intake modulate psychological profile and body composition of women affected by normal weight obese syndrome and obesity? A double blind randomized clinical trial. J. Transl. Med. 2017, 15, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, L.; Roager, H.M.; Astrup, A.; Hjorth, M.F. Microbial enterotypes in personalized nutrition and obesity management. Am. J. Clin. Nutr. 2018, 108, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Song, E.J.; Han, K.; Lim, T.J.; Lim, S.; Chung, M.J.; Nam, M.H.; Kim, H.; Nam, Y.D. Effect of probiotics on obesity-related markers per enterotype: A double-blind, placebo-controlled, randomized clinical trial. EPMA J. 2020, 11, 31–51. [Google Scholar] [CrossRef]
- Shirvani-Rad, S.; Tabatabaei-Malazy, O.; Mohseni, S.; Hasani-Ranjbar, S.; Soroush, A.R.; Hoseini-Tavassol, Z.; Ejtahed, H.S.; Larijani, B. Probiotics as a complementary therapy for management of obesity: A systematic review. Evid.-Based Complement. Altern. Med. 2021, 2021, 6688450. [Google Scholar] [CrossRef]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Kotler, E.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.; et al. Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features. Cell 2018, 174, 1388–1405. [Google Scholar] [CrossRef] [Green Version]
- Shang, A.; Gan, R.Y.; Xu, X.Y.; Mao, Q.Q.; Zhang, P.Z.; Li, H.B. Effects and mechanisms of edible and medicinal plants on obesity: An updated review. Crit. Rev. Food Sci. Nutr. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.M.; Silva, N.; Freitas, R.M.P.; Freitas, M.B.D.; Chaves, J.B.P.; Oliveira, L.L.; Martino, H.S.D.; Alfenas, R.C.G. Effects of yacon flour associated with an energy restricted diet on intestinal permeability, fecal short chain fatty acids, oxidative stress and inflammation markers levels in adults with obesity or overweight: A randomized, double blind, placebo controlled clinical trial. Arch. Endocrinol. Metab. 2020, 64, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Adriano, L.S.; Dionisio, A.P.; Pinto de Abreu, F.A.; Wurlitzer, N.J.; Cordeiro de Melo, B.R.; Ferreira Carioca, A.A.; de Carvalho Sampaio, H.A. Acute postprandial effect of yacon syrup ingestion on appetite: A double blind randomized crossover clinical trial. Food Res. Int. 2020, 137, 109648. [Google Scholar] [CrossRef]
- Hume, M.P.; Nicolucci, A.C.; Reimer, R.A. Prebiotic supplementation improves appetite control in children with overweight and obesity: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 105, 790–799. [Google Scholar] [CrossRef] [Green Version]
- Parnell, J.A.; Klancic, T.; Reimer, R.A. Oligofructose decreases serum lipopolysaccharide and plasminogen activator inhibitor-1 in adults with overweight/obesity. Obesity 2017, 25, 510–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimer, R.A.; Willis, H.J.; Tunnicliffe, J.M.; Park, H.; Madsen, K.L.; Soto-Vaca, A. Inulin-type fructans and whey protein both modulate appetite but only fructans alter gut microbiota in adults with overweight/obesity: A randomized controlled trial. Mol. Nutr. Food Res. 2017, 61, e1700484. [Google Scholar] [CrossRef]
- Wall, C.R.; Hill, R.J.; Lovell, A.L.; Matsuyama, M.; Milne, T.; Grant, C.C.; Jiang, Y.; Chen, R.X.; Wouldes, T.A.; Davies, P.S.W. A multicenter, double-blind, randomized, placebo-controlled trial to evaluate the effect of consuming Growing Up Milk “Lite” on body composition in children aged 12-23 mo. Am. J. Clin. Nutr. 2019, 109, 576–585. [Google Scholar] [CrossRef]
- Raji Lahiji, M.; Zarrati, M.; Najafi, S.; Yazdani, B.; Cheshmazar, E.; Razmpoosh, E.; Janani, L.; Raji Lahiji, M.; Shidfar, F. Effects of synbiotic supplementation on serum adiponectin and inflammation status of overweight and obese breast cancer survivors: A randomized, triple-blind, placebo-controlled trial. Supportive Care Cancer 2021, 29, 4147–4157. [Google Scholar] [CrossRef]
- Raji Lahiji, M.; Najafi, S.; Janani, L.; Yazdani, B.; Razmpoosh, E.; Zarrati, M. The effect of synbiotic on glycemic profile and sex hormones in overweight and obese breast cancer survivors following a weight-loss diet: A randomized, triple-blind, controlled trial. Clin. Nutr. 2021, 40, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Hadi, A.; Sepandi, M.; Marx, W.; Moradi, S.; Parastouei, K. Clinical and psychological responses to synbiotic supplementation in obese or overweight adults: A randomized clinical trial. Complement. Ther. Med. 2019, 47, 102216. [Google Scholar] [CrossRef] [PubMed]
- Krumbeck, J.A.; Rasmussen, H.E.; Hutkins, R.W.; Clarke, J.; Shawron, K.; Keshavarzian, A.; Walter, J. Probiotic Bifidobacterium strains and galactooligosaccharides improve intestinal barrier function in obese adults but show no synergism when used together as synbiotics. Microbiome 2018, 6, 121. [Google Scholar] [CrossRef] [PubMed]
- Hadi, A.; Alizadeh, K.; Hajianfar, H.; Mohammadi, H.; Miraghajani, M. Efficacy of synbiotic supplementation in obesity treatment: A systematic review and meta-analysis of clinical trials. Crit. Rev. Food Sci. Nutr. 2020, 60, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Primec, M.; Klemenak, M.; Di Gioia, D.; Aloisio, I.; Bozzi Cionci, N.; Quagliariello, A.; Gorenjak, M.; Micetic-Turk, D.; Langerholc, T. Clinical intervention using Bifidobacterium strains in celiac disease children reveals novel microbial modulators of TNF-alpha and short-chain fatty acids. Clin. Nutr. 2019, 38, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Crovesy, L.; El-Bacha, T.; Rosado, E.L. Modulation of the gut microbiota by probiotics and symbiotics is associated with changes in serum metabolite profile related to a decrease in inflammation and overall benefits to metabolic health: A double-blind randomized controlled clinical trial in women with obesity. Food Funct. 2021, 12, 2161–2170. [Google Scholar] [CrossRef]
- Banasiewicz, T.; Domagalska, D.; Borycka-Kiciak, K.; Rydzewska, G. Determination of butyric acid dosage based on clinical and experimental studies—A literature review. Gastroenterol. Rev. 2020, 15, 119–125. [Google Scholar] [CrossRef]
- Tonucci, L.B.; Olbrich Dos Santos, K.M.; Licursi de Oliveira, L.; Rocha Ribeiro, S.M.; Duarte Martino, H.S. Clinical application of probiotics in type 2 diabetes mellitus: A randomized, double-blind, placebo-controlled study. Clin. Nutr. 2017, 36, 85–92. [Google Scholar] [CrossRef]
- Hsieh, M.C.; Tsai, W.H.; Jheng, Y.P.; Su, S.L.; Wang, S.Y.; Lin, C.C.; Chen, Y.H.; Chang, W.W. The beneficial effects of Lactobacillus reuteri ADR-1 or ADR-3 consumption on type 2 diabetes mellitus: A randomized, double-blinded, placebo-controlled trial. Sci. Rep. 2018, 8, 16791. [Google Scholar] [CrossRef]
- Sato, J.; Kanazawa, A.; Azuma, K.; Ikeda, F.; Goto, H.; Komiya, K.; Kanno, R.; Tamura, Y.; Asahara, T.; Takahashi, T.; et al. Probiotic reduces bacterial translocation in type 2 diabetes mellitus: A randomised controlled study. Sci. Rep. 2017, 7, 12115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickens, K.L.; Barthow, C.A.; Murphy, R.; Abels, P.R.; Maude, R.M.; Stone, P.R.; Mitchell, E.A.; Stanley, T.V.; Purdie, G.L.; Kang, J.M.; et al. Early pregnancy probiotic supplementation with Lactobacillus rhamnosus HN001 may reduce the prevalence of gestational diabetes mellitus: A randomised controlled trial. Br. J. Nutr. 2017, 117, 804–813. [Google Scholar] [CrossRef] [Green Version]
- Callaway, L.K.; McIntyre, H.D.; Barrett, H.L.; Foxcroft, K.; Tremellen, A.; Lingwood, B.E.; Tobin, J.M.; Wilkinson, S.; Kothari, A.; Morrison, M.; et al. Probiotics for the prevention of gestational diabetes mellitus in overweight and obese women: Findings from the SPRING double-blind randomized controlled trial. Diabetes Care 2019, 42, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kijmanawat, A.; Panburana, P.; Reutrakul, S.; Tangshewinsirikul, C. Effects of probiotic supplements on insulin resistance in gestational diabetes mellitus: A double-blind randomized controlled trial. J. Diabetes Investig. 2019, 10, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Dawe, J.P.; McCowan, L.M.E.; Wilson, J.; Okesene-Gafa, K.A.M.; Serlachius, A.S. Probiotics and maternal mental health: A randomised controlled trial among pregnant women with obesity. Sci. Rep. 2020, 10, 1291. [Google Scholar] [CrossRef] [Green Version]
- Akbari, E.; Asemi, Z.; Daneshvar Kakhaki, R.; Bahmani, F.; Kouchaki, E.; Tamtaji, O.R.; Hamidi, G.A.; Salami, M. Effect of probiotic supplementation on cognitive function and metabolic status in alzheimer’s disease: A randomized, double-blind and controlled trial. Front. Aging Neurosci. 2016, 8, 256. [Google Scholar] [CrossRef] [Green Version]
- Akkasheh, G.; Kashani-Poor, Z.; Tajabadi-Ebrahimi, M.; Jafari, P.; Akbari, H.; Taghizadeh, M.; Memarzadeh, M.R.; Asemi, Z.; Esmaillzadeh, A. Clinical and metabolic response to probiotic administration in patients with major depressive disorder: A randomized, double-blind, placebo-controlled trial. Nutrition 2016, 32, 315–320. [Google Scholar] [CrossRef]
- Dehghan, P.; Farhangi, M.A.; Tavakoli, F.; Aliasgarzadeh, A.; Akbari, A.M. Impact of prebiotic supplementation on T-cell subsets and their related cytokines, anthropometric features and blood pressure in patients with type 2 diabetes mellitus: A randomized placebo-controlled Trial. Complement. Ther. Med. 2016, 24, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Vaghef-Mehrabany, E.; Ranjbar, F.; Asghari-Jafarabadi, M.; Hosseinpour-Arjmand, S.; Ebrahimi-Mameghani, M. Calorie restriction in combination with prebiotic supplementation in obese women with depression: Effects on metabolic and clinical response. Nutr. Neurosci. 2021, 24, 339–353. [Google Scholar] [CrossRef]
- Javadi, L.; Ghavami, M.; Khoshbaten, M.; Safaiyan, A.; Barzegari, A.; Pourghassem Gargari, B. The effect of probiotic and/or prebiotic on liver function tests in patients with nonalcoholic fatty liver disease: A double blind randomized clinical trial. Iran. Red Crescent Med. J. 2017, 19, e46017. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Fogacci, F.; Bove, M.; Giovannini, M.; Borghi, C. Impact of a short-term synbiotic supplementation on metabolic syndrome and systemic inflammation in elderly patients: A randomized placebo-controlled clinical trial. Eur. J. Nutr. 2021, 60, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Djaja, N.; Permadi, I.; Witjaksono, F.; Soewondo, P.; Abdullah, M.; Agustina, R.; Ali, S. The effect of Job’s tears-enriched yoghurt on GLP-1, calprotectin, blood glucose levels and weight of patients with type 2 diabetes mellitus. Mediterr. J. Nutr. Metab. 2019, 12, 163–171. [Google Scholar] [CrossRef]
- Horvath, A.; Leber, B.; Feldbacher, N.; Tripolt, N.; Rainer, F.; Blesl, A.; Trieb, M.; Marsche, G.; Sourij, H.; Stadlbauer, V. Effects of a multispecies synbiotic on glucose metabolism, lipid marker, gut microbiome composition, gut permeability, and quality of life in diabesity: A randomized, double-blind, placebo-controlled pilot study. Eur. J. Nutr. 2020, 59, 2969–2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimi, Z.S.; Nasli-Esfahani, E.; Nadjarzade, A.; Mozaffari-Khosravi, H. Effect of symbiotic supplementation on glycemic control, lipid profiles and microalbuminuria in patients with non-obese type 2 diabetes: A randomized, double-blind, clinical trial. J. Diabetes Metab. Disord. 2017, 16, 23. [Google Scholar] [CrossRef] [Green Version]
- Kassaian, N.; Feizi, A.; Rostami, S.; Aminorroaya, A.; Yaran, M.; Amini, M. The effects of six months supplementation with probiotics and synbiotic on gut microbiota in the adults with prediabetes: A double blind randomized clinical trial. Nutrition 2020, 79–80, 110854. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Rodriguez, J.; Taminiau, B.; Amadieu, C.; Herpin, F.; Allaert, F.A.; Cani, P.D.; Daube, G.; Bindels, L.B.; Delzenne, N.M. Improvement of gastrointestinal discomfort and inflammatory status by a synbiotic in middle-aged adults: A double-blind randomized placebo-controlled trial. Sci. Rep. 2021, 11, 2627. [Google Scholar] [CrossRef] [PubMed]
- Cakir, M.; Aksel Isbilen, A.; Eyupoglu, I.; Sag, E.; Orem, A.; Mazlum Sen, T.; Kaklikkaya, N.; Kaya, G. Effects of long-term synbiotic supplementation in addition to lifestyle changes in children with obesity-related non-alcoholic fatty liver disease. Turk. J. Gastroenterol. 2017, 28, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Tamtaji, O.R.; Heidari-Soureshjani, R.; Mirhosseini, N.; Kouchaki, E.; Bahmani, F.; Aghadavod, E.; Tajabadi-Ebrahimi, M.; Asemi, Z. Probiotic and selenium co-supplementation, and the effects on clinical, metabolic and genetic status in Alzheimer’s disease: A randomized, double-blind, controlled trial. Clin. Nutr. 2019, 38, 2569–2575. [Google Scholar] [CrossRef]
- Maykish, A.; Sikalidis, A.K. Utilization of hydroxyl-methyl butyrate, leucine, glutamine and arginine supplementation in nutritional management of sarcopenia-implications and clinical considerations for type 2 diabetes mellitus risk modulation. J. Pers. Med. 2020, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Raqib, R.; Sarker, P.; Mily, A.; Alam, N.H.; Arifuzzaman, A.S.; Rekha, R.S.; Andersson, J.; Gudmundsson, G.H.; Cravioto, A.; Agerberth, B. Efficacy of sodium butyrate adjunct therapy in shigellosis: A randomized, double-blind, placebo-controlled clinical trial. BMC Infect. Dis. 2012, 12, 111. [Google Scholar] [CrossRef] [Green Version]
- Yao, K.; Zeng, L.; He, Q.; Wang, W.; Lei, J.; Zou, X. Effect of probiotics on glucose and lipid metabolism in type 2 diabetes mellitus: A meta-analysis of 12 randomized controlled trials. Med. Sci. Monitor. 2017, 23, 3044–3053. [Google Scholar] [CrossRef] [Green Version]
- Kocsis, T.; Molnar, B.; Nemeth, D.; Hegyi, P.; Szakacs, Z.; Balint, A.; Garami, A.; Soos, A.; Marta, K.; Solymar, M. Probiotics have beneficial metabolic effects in patients with type 2 diabetes mellitus: A meta-analysis of randomized clinical trials. Sci. Rep. 2020, 10, 11787. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yue, R.; Zhang, B.; Li, Z.; Shui, J.; Huang, X. Effects of probiotics on blood glucose, biomarkers of inflammation and oxidative stress in pregnant women with gestational diabetes mellitus: A meta-analysis of randomized controlled trials. Med. Clin. 2020, 154, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Lin, C.; Huang, S.; Xu, Y. Traditional Chinese medicine and intestinal microbiota: A complementary and integrative health approach to ameliorate obesity-related diseases. Holist. Nurs. Pract. 2019, 33, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Song, Z.; Weng, J.; Fantus, I.G. Curcumin and other dietary polyphenols: Potential mechanisms of metabolic actions and therapy for diabetes and obesity. Am. J. Physiol.-Endocrinol. Metab. 2018, 314, 201–205. [Google Scholar] [CrossRef]
- Daniel, S.; Phillippi, D.; Schneider, L.J.; Nguyen, K.N.; Mirpuri, J.; Lund, A.K. Exposure to diesel exhaust particles results in altered lung microbial profiles, associated with increased reactive oxygen species/reactive nitrogen species and inflammation, in C57Bl/6 wildtype mice on a high-fat diet. Part. Fibre Toxicol. 2021, 18, 3. [Google Scholar] [CrossRef]
- Tan, Q.; Orsso, C.E.; Deehan, E.C.; Kung, J.Y.; Tun, H.M.; Wine, E.; Madsen, K.L.; Zwaigenbaum, L.; Haqq, A.M. Probiotics, prebiotics, synbiotics, and fecal microbiota transplantation in the treatment of behavioral symptoms of autism spectrum disorder: A systematic review. Autism Res. 2021, 1–17, in press. [Google Scholar] [CrossRef]
- Dong, Y.; Xu, M.; Chen, L.; Bhochhibhoya, A. Probiotic foods and supplements interventions for metabolic syndromes: A systematic review and meta-analysis of recent clinical trials. Ann. Nutr. Metab. 2019, 74, 224–241. [Google Scholar] [CrossRef]
- Sitkin, S.; Pokrotnieks, J. Clinical potential of anti-inflammatory effects of Faecalibacterium prausnitzii and butyrate in inflammatory bowel disease. Inflamm. Bowel. Dis. 2019, 25, 40–41. [Google Scholar] [CrossRef]
- Magno, A.L.; Herat, L.Y.; Kiuchi, M.G.; Schlaich, M.P.; Ward, N.C.; Matthews, V.B. The influence of hypertensive therapies on circulating factors: Clinical implications for SCFAs, FGF21, TNFSF14 and TNF-alpha. J. Clin. Med. 2020, 9, 2764. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Supplements | Models | Doses | Target Disease | Main Effects and Mechanisms | Ref. |
|---|---|---|---|---|---|
| Probiotics | |||||
| Bifidobacterium lactis LMG P-28149, and Lactobacillus rhamnosus LMG S-28148 | in vitro, in vivo | 108 CFU | Obesity and insulin resistance | Restoring Akkermansia muciniphila and Rikenellaceae. Upregulating PPARγ and lipoprotein lipase expression. Enhancing insulin sensitivity and TG clearance Decreasing Lactobacillaceae. | [47] |
| Bifidobacterium longum PI10 and Ligilactobacillus salivarius PI2 | in vitro, in vivo | 5 × 108 CFU | Obesity | Upregulating GLP1 and IL-10 expression. | [70] |
| Lactobacillus plantarum NK3 and Bifidobacterium longum NK49 | in vivo | 1 × 109 CFU | Obesity and osteoporosis | Improving intestinal barrier. Suppressing LPS production. Downregulating NF-κB-linked TNF-α expression. | [71] |
| Lactobacillus fermentum MCC2760 | in vivo | 109 CFU | Type 2 diabetes mellitus | Improving intestinal barrier. Upregulating GLUT4, GLP1, and ZO-1 expression. | [72] |
| Lactobacillus casei | in vivo | 2 × 108 CFU | Hypertension | Increasing Akkermansia and Lactobacillus. Decreasing Firmicutes to Bacteroidetes ratio and ACE expression. | [51] |
| Bifidobacterium breve CECT7263 and Lactobacillus fermentum CECT5716 | in vivo | 109 CFU | Hypertension | Increasing butyrate-related bacteria. Elevating the plasma level of butyrate. Reducing LPS production. | [73] |
| Lactobacillus johnsonii, and Lactobacillus reuteri | in vivo | 1010 CFU | Inflammatory bowel disease and Osteoporosis | Improving tetrathionate metabolism. Decreasing Yersinia enterocolitica. | [49] |
| Lactobacillus paracasei | in vivo | 0.8–1.2 × 109 CFU | Colitis with metabolic disorder | Producing palmitoylethanolamide to maintain intestinal function. | [74] |
| Lactobacillus reuteri LR6 | in vivo | 1 × 109 CFU | Protein-energy malnutrition | Increasing Bifidobacteria, Firmicutes, and Lactobacilli. | [52] |
| Prebiotics | |||||
| Maize starch dextrin and Lentil | in vitro in vivo | 70.8% red lentil diet | Obesity | Increasing Actinobacteria and Bacteroidetes. Decreasing Firmicutes. | [53,54] |
| Inulin oligofructose | in vivo | 5% in diet, 0.6 g/day | Obesity | Increasing Bifidobacterium. Decreasing Firmicutes to Bacteroidetes ratio. | [58,59] |
| Chicory oligofructose | in vivo | 10% in diet | Obesity | Increasing Bifidobacterium and Collinsella. | [60] |
| Amylosucrase-modified chestnut starch | in vivo | 1500 mg/kg | Obesity | Upregulating SCFAs-GPR43-mediated pathway. | [23] |
| Fuji FF | in vivo | 10% in diet | Obesity | Increasing acetic, propionic, and butyric acids production. | [75] |
| Unsaturated alginate oligosaccharides | in vivo | 400 mg/kg | Obesity | Upregulating ZO-1 and occludin expression. | [42] |
| Acorn and sago polysaccharides | in vitro, in vivo | 1% v/v, 5% in diet | Obesity and type 2 diabetes mellitus | Reducing gut hyperpermeability and mucosal inflammatory biomarkers. | [62] |
| Galacto-oligosaccharides | in vivo | 7% w/w | Obesity and Insulin resistance | Increasing GLP1 expression. Decreasing fecal bile acid excretion. | [76] |
| Resistant dextrin from wheat and corn starch | in vivo | 5 g/kg | Type 2 diabetes mellitus | Increasing Akkermansia and Prevotella abundances. Upregulating IRS1-Akt-GLUT2 and SIRT1-AMPK-PPARα-CPR1α pathways. | [56] |
| Isomaltodextrin | in vivo | 1, 2.5, 5% in drinking water | Insulin resistance | Increasing acetic and butyric acids production. Improving intestinal barrier. Reducing circulation endotoxin level. | [38] |
| Whole garlic | in vivo | 5% in diet | Dyslipidemia | Increasing Lachnospiraceae. Decreasing Prevotella. | [57] |
| Fucoidan and Galacto-oligosaccharides | in vitro, in vivo | 100, 800 mg/kg | Dyslipidemia | Increasing Bacteroidetes, Proteobacteria, and the bile salt hydrolase activity of Lactobacillus casei DM8121. Decreasing Actinobacteria and Firmicutes. | [61] |
| Oat and rye brans dietary fiber | in vivo | 10% in diet | Metabolic disorder | Regulating bile acids and tryptophan–serotonin metabolic pathways. | [77] |
| Agave salmiana fructan | in vivo | 10% in diet | Metabolic disorder | Increasing lactic acid production. | [78] |
| Glycolipids from tilapia heads | in vivo | 30 mg/kg | Colitis with metabolic disorder | Increasing Akkermansia, Allobaculum, Bifidobacterium, Coprococcus, Oscillospira, and Prevotellaceae. | [79] |
| Long-chain inulin | in vivo | 5% in diet | Hypertension | Decreasing the fecal levels of acetate and propionate, and the plasma level of TMAO. | [51] |
| Synbiotics | |||||
| Bifidobacterium, Lactobacillus, Lactococcus, Propionibacterium plus omega-3 fatty acids | in vivo | 2.5 mL/kg | Obesity | Revealing positively synergistic effect on reducing hepatic steatosis and lipid accumulation compared to probiotics alone. | [63,64] |
| Bacillus licheniformis plus xylo-oligosaccharides | in vivo | 7.5 × 108 CFU/mL and 2 g/mL | Obesity | Revealing positively synergistic effect on improving body weight gain and lipid metabolism. Decreasing Desulfovibrionaceae and Ruminococcaceae | [65] |
| Lactobacillus plantarum PMO 08 plus chia seeds | in vivo | 1 × 109 CFU/mL and 4% in diet | Obesity | Revealing positively synergistic effect on improving obesity. Increasing Lactobacillus plantarum | [66] |
| Bifidobacterium lactis, Lactobacillus paracasei DSM 4633, plus oat b-glucan | in vivo | 108 CFU and 1 g/kg | Obesity | Increasing fecal acetate, propionate, and butyrate levels. Decreasing the bile acid pools. | [80] |
| Lactobacillus paracasei HII01 plus xylo-oligosaccharides | in vivo | 1 × 108 CFU and 10% in PBS | Obesity | Inhibiting metabolic endotoxemia. Decreasing Firmicutes to Bacteroidetes ratio and Enterobacteriaceae | [81] |
| Lactobacillus paracasei N1115 plus fructo-oligosaccharides | in vivo | 2.2 × 109 CFU/mL and 4 g/kg/day | NAFLD | Decreasing LPS production. Downregulating TLR4 and NF-κB expression. Upregulating p38 MAPK pathway and the expression of occludin 1 and claudin 1. | [82] |
| Bifidobacterium bifidum V, Lactobacillus plantarum X plus Salvia miltiorrhiza polysaccharide | in vivo | 1–2 × 108 CFU and 50 mg/kg | NAFLD | Alleviating hepatic steatosis and insulin resistance improvement. Decreasing LPS level. | [83] |
| Clostridium butyricum plus corn bran | in vivo | 1 × 108 CFU/g and 5% in diet | Intestinal impairment with metabolic disorder | Increasing the growth of acetate-produced bacteria and the production of acetate and isovalerate. Decreasing pathogen abundances. | [84] |
| Postbiotics | |||||
| Exopolysaccharide from Lactobacillus plantarum L-14 | in vitro, in vivo | 100 μM and 500 mg/kg | Obesity | Upregulating TLR2-AMPK pathway. | [85] |
| Muramyl dipeptide | in vivo | 100 μg | Obesity | Upregulating NOD2-IRF4 pathway | [86,87] |
| Butyrate | in vivo | 200 mg/kg | Type 2 diabetes mellitus | Upregulating GPR43-NLCR3-TRAF6 pathway. | [88] |
| Butyrate | in vivo | 5% in diet | Type 2 diabetes mellitus | Promoting insulin secretion without impairing pancreatic beta cells. | [89] |
| Butyrate | in vivo | 140 mM | Metabolic disorder | Increasing Lachnospiraceae and Proteobacteria. Decreasing Clostridiaceae. | [67] |
| Butyrate | in vivo | 200 mg/kg | NAFLD | Increasing Blautia, Christensenellaceae, and Lactobacillus. | [68] |
| Butyrate | in vivo | 0.6 g/kg | NAFLD | Increasing ZO-1 expression. Decreasing the levels of endotoxin. | [90] |
| Long-chain polyphosphate from Lactobacillus brevis | in vivo | 0.05 μg/μL | Colitis with metabolic disorder | Upregulating ERK pathway. | [91] |
| Supplements | Doses | Duration | Sample Size | Target Disease | Main Effects | Ref. |
|---|---|---|---|---|---|---|
| Probiotics | ||||||
| Bifidobacterium animalis SGB06, Bifidobacterium bifidum SGB02, Lactobacillus acidophilus SGL11, Lactobacillus delbrueckii DSM 20081, Lactococcus lactis SGLc01, Lactobacillus plantarum SGL07, Lactobacillus reuteri SGL01, Streptococcus thermophiles, and Streptococcus thermophilus | 1.5 × 1010 CFU | 3 weeks | 60 | Obesity | Improving body composition, bacterial composition, and psychopathological status. | [100] |
| Bifidobacterium lactis | 2.72 × 1010 CFU | 45 days | 45 | Obesity | Improving obesity, blood lipids, and inflammatory markers such as TNF-α and IL-6. | [99] |
| Bifidobacterium breve CBT BR3 and Lactobacillus plantarum CBT LP3 | 15 × 1010 CFU | 12 weeks | 50 | Obesity | More effectively improve obese biomarkers in patients with Prevotella-rich enterotype than Bacteroides-rich enterotype. | [102] |
| Bifidobacterium lactis BB-12 and Lactobacillus acidophilus La-5 | 109 CFU | 6 weeks | 50 | Type 2 diabetes mellitus | Improving fructosamine, HbA1c and IL-10 levels. | [120] |
| Lactobacillus reuteri strain ADR-1 and ADR-3 | 4 × 109 CFU for ADR-1 2 × 1010 CFU for ADR-3 | 12 weeks | 68 | Type 2 diabetes mellitus | Decreasing HbA1c and cholesterol levels. | [121] |
| Fermented milk processed by Lactobacillus casei strain Shirota | 80 mL | 16 weeks | 70 | Type 2 diabetes mellitus | Increase Clostridium coccoides, Clostridium leptum, and Lactobacillus. Decreasing of translocated gut bacteria. | [122] |
| Lactobacillus rhamnosus HN001 | 6 × 109 CFU | 14–16 weeks’ gestation | 423 | Gestational diabetes mellitus | Decreasing the relapse prevalence of diabetes mellitus. | [123] |
| Bifidobacterium lactis and Lactobacillus rhamnosus | 1 × 109 CFU | 16–28 weeks’ gestation | 411 | Gestational diabetes mellitus | Not preventing gestational diabetes mellitus. | [124] |
| Bifidobacterium and Lactobacillus | 1 × 109 CFU | 24–28 weeks’ gestation | 28 | Gestational diabetes mellitus | Improving glucose metabolism. Not affecting weight gain. | [125] |
| Bifidobacterium lactis BB12 and Lactobacillus rhamnosus GG | 6.5 × 109 CFU | 17–36 weeks’ gestation | 230 | Gestational diabetes mellitus | Not improving depression, anxiety, and physical well-being status. | [126] |
| Bifidobacterium bifidum, Lactobacillus acidophilus, Lactobacillus casei, and Lactobacillus fermentum | 2 × 109 CFU/g | 12 weeks | 60 | Alzheimer’s disease with metabolic disorder | Improving cognitive function and metabolic status. | [127] |
| Bifidobacterium bifidum, Lactobacillus acidophilus, and Lactobacillus casei | 2 × 109 CFU/g | 8 weeks | 40 | Major depressive with metabolic disorder | Improving insulin resistance, C-reactive protein, and total glutathione level. | [128] |
| Prebiotics | ||||||
| Yacon | 25 g | 6 weeks | 26–40 | Obesity | Increasing the plasma antioxidant capacity. Decreasing oxidative stress and fecal SCFAs levels. | [106,107] |
| Inulin oligofructose | 8 g/day | 16 weeks | 42 | Obesity | Decreasing energy intake by modifying appetite. | [108] |
| Oligofructose | 21 g | 12 weeks | 37 | Obesity | Improving metabolic endotoxemia. Decreasing PAI1 level. | [109] |
| Inulin-type fructan and whey protein | 5–8 g | 12 weeks | 125 | Obesity | Regulating appetite. Increase Bifidobacterium. | [110] |
| Inulin oligofructose | 10 g/day | 2 months | 46 | Type 2 diabetes mellitus | Improving glycemic status, lipid profiles, and immune markers | [129] |
| Inulin | 10 g/day | 8 weeks | 45 | Obesity-related major depressive | Enhancing beneficial effects of calorie-restricted diet on fat mass and TC level. | [130] |
| Inulin | 10 g/day | 3 months | 75 | NAFLD | Improving the grade of fatty liver and the serum levels of aminotransferase enzymes. | [131] |
| Synbiotics | ||||||
| Growing up milk | 300 mL | 12 months | 160 | Obesity | Improving body fat gain. | [111] |
| Bifidobacterium breve, Bifidobacterium longum, Lactobacillus acidophilus, Lactobacillus bulgaricus, Lactobacillus casei, Lactobacillus rhamnosus, Streptococcus thermophiles, plus fructo-oligosaccharides | 109 CFU/g and 35 mg | 8 weeks | 76 | Obesity | Improving serum insulin level and insulin resistance | [112,113] |
| Bifidobacterium bifidum, Lactobacillus acidophilus, Lactobacillus casei, plus inulin | 2 × 109 CFU/g and 0.8 g | 8 weeks | 59 | Obesity | Improving lipid profiles and psychological status. Not benefiting BMI, blood pressure, glucose metabolism, and waist circumference. | [114] |
| Bifidobacterium adolescentis IVS-1, Bifidobacterium lactis BB-12, plus galacto-oligosaccharides | 1 × 109 CFU and 5 g | 3 weeks | 114 | Obesity | Improving intestinal barrier function as single agents. No synergistic effects. | [115] |
| Lactobacillus acidophilus PBS066, Lactobacillus plantarum PBS067, and Lactobacillus reuteri PBS072, plus inulin and fructo-oligosaccharides | 1 × 109 CFU and 12.97% in liquid | 2 months | 60 | Metabolic disorder | Decreasing metabolic syndrome prevalence | [132] |
| Probiotic yogurt plus Coix lacryma-jobi | 100 mL and 25 g | 12 weeks | 60 | Type 2 diabetes mellitus | Reducing body weight and fasting blood glucose | [133] |
| Bifidobacterium bifidum W23, Bifidobacterium lactis W51, Bifidobacterium lactis W52, Lactobacillus acidophilus W37, Lactobacillus casei W56, Lactobacillus brevis W63, Lactobacillus salivarius W24, Lactococcus lactis W58, and Lactococcus lactis W19, plus galacto-oligosaccharides P11 and fructo-oligosaccharides P6 | 1.5 × 1010 CFU and 8 g | 6 months | 26 | Type 2 diabetes mellitus | Improving hip circumference, zonulin and lipoprotein. Not affecting glucose metabolism. | [134] |
| Bifidobacterium, Lactobacillus, and Streptococus thermophilus plus fructo-oligosaccharide | 500 mg/day | 9 weeks | 70 | Type 2 diabetes mellitus | Improving HbA1c, BMI, and microalbuminuria. Not affecting fasting blood glucose, lipid profiles, and creatinine. | [135] |
| Bifidobacterium bifidum, Bifidobacterium lactis, Bifidobacterium longum, and Lactobacillus acidophilus plus inulin | 1.5 × 109 CFU and 6 g | 6 months | 120 | Prediabetes mellitus | Not restoring the balance of gut microbiota. | [136] |
| Bifidobacterium lactis plus fructo-oligosaccharides | 5 × 109 CFU/bag and 4.95 g/bag | 30 days | 27 | Digestive disorder | Improving intestinal function. Decreasing IL-6, IL-8, IL-17α, and IFN-γ levels. | [137] |
| Bifidobacterium lactis, Lactobacillus acidophilus, and Lactobacillus casei plus chicory inulin | 7 × 109 CFU and 100 mg | 4 months | 28 | NAFLD | Improving fatty liver grade, and inflammatory and antioxidative status. | [138] |
| Bifidobacterium bifidum, Bifidobacterium longum, and Lactobacillus acidophilus plus selenium | 2 × 109 CFU and 200 mg | 12 weeks | 79 | Alzheimer’s disease with metabolic disorder | Improving the cognitive function and metabolic function. | [139] |
| Postbiotics | ||||||
| Hydroxyl-methyl butyrate | 3 g | 1 week | 34 | Diabetic-related sarcopenia | Inhibiting catabolic effect on skeletal muscle. | [140] |
| Butyrate | 80 mM | 3 days | 80 | Bacillary dysentery with metabolic disorder | Improving alimentary canal function. Alleviating pathological impairment of colonic mucosa barrier Enhancing antimicrobial peptides release. | [141] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.-Y.; Zhou, D.-D.; Gan, R.-Y.; Huang, S.-Y.; Zhao, C.-N.; Shang, A.; Xu, X.-Y.; Li, H.-B. Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases Targeting Gut Microbiota: A Narrative Review. Nutrients 2021, 13, 3211. https://doi.org/10.3390/nu13093211
Li H-Y, Zhou D-D, Gan R-Y, Huang S-Y, Zhao C-N, Shang A, Xu X-Y, Li H-B. Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases Targeting Gut Microbiota: A Narrative Review. Nutrients. 2021; 13(9):3211. https://doi.org/10.3390/nu13093211
Chicago/Turabian StyleLi, Hang-Yu, Dan-Dan Zhou, Ren-You Gan, Si-Yu Huang, Cai-Ning Zhao, Ao Shang, Xiao-Yu Xu, and Hua-Bin Li. 2021. "Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases Targeting Gut Microbiota: A Narrative Review" Nutrients 13, no. 9: 3211. https://doi.org/10.3390/nu13093211