
BMI, Alcohol Consumption and Gut Microbiome Species Richness Are Related to Structural and Functional Neurological Abnormalities

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Cohorts and Study Design
2.2. Anthropometric Characteristics
2.3. Medical Questionnaire
2.4. Dietary Assessment
2.5. Biochemical Analyses
2.6. Analysis of the Intestinal Microbiome
2.7. DNA Extraction, 16S rDNA Sequencing and Quality Control
2.8. Normalization and Taxonomic Classification
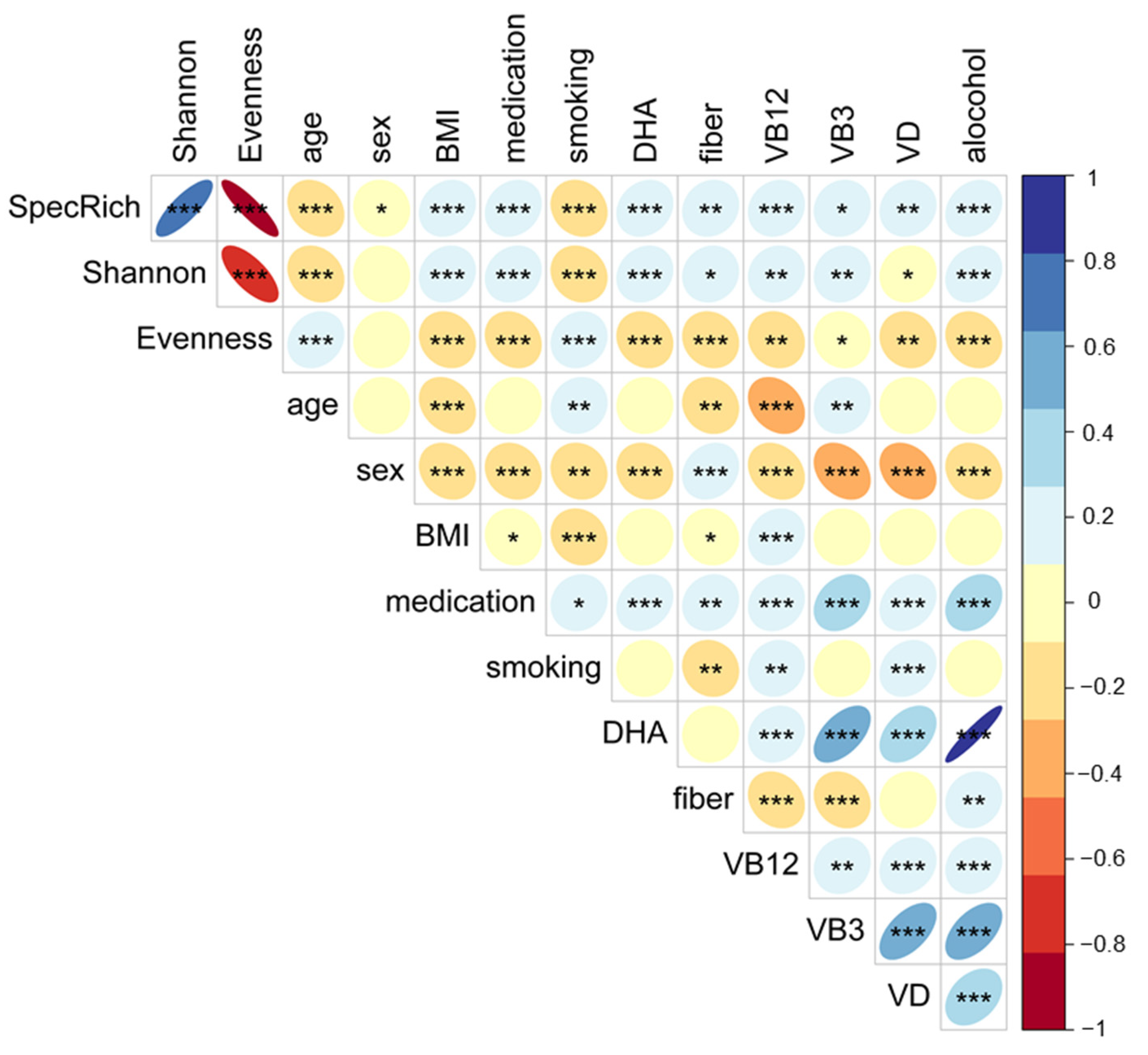
2.9. Statistical Analysis of the Microbiome Data
3. Statistics
4. Results
4.1. Characterization of Clinical Biochemistry and Anthropometric Data
4.2. Characterization of Micro- and Macronutrients
5. Neurological Cases and Microbiome
5.1. Core Measurable Microbiome (CMM)
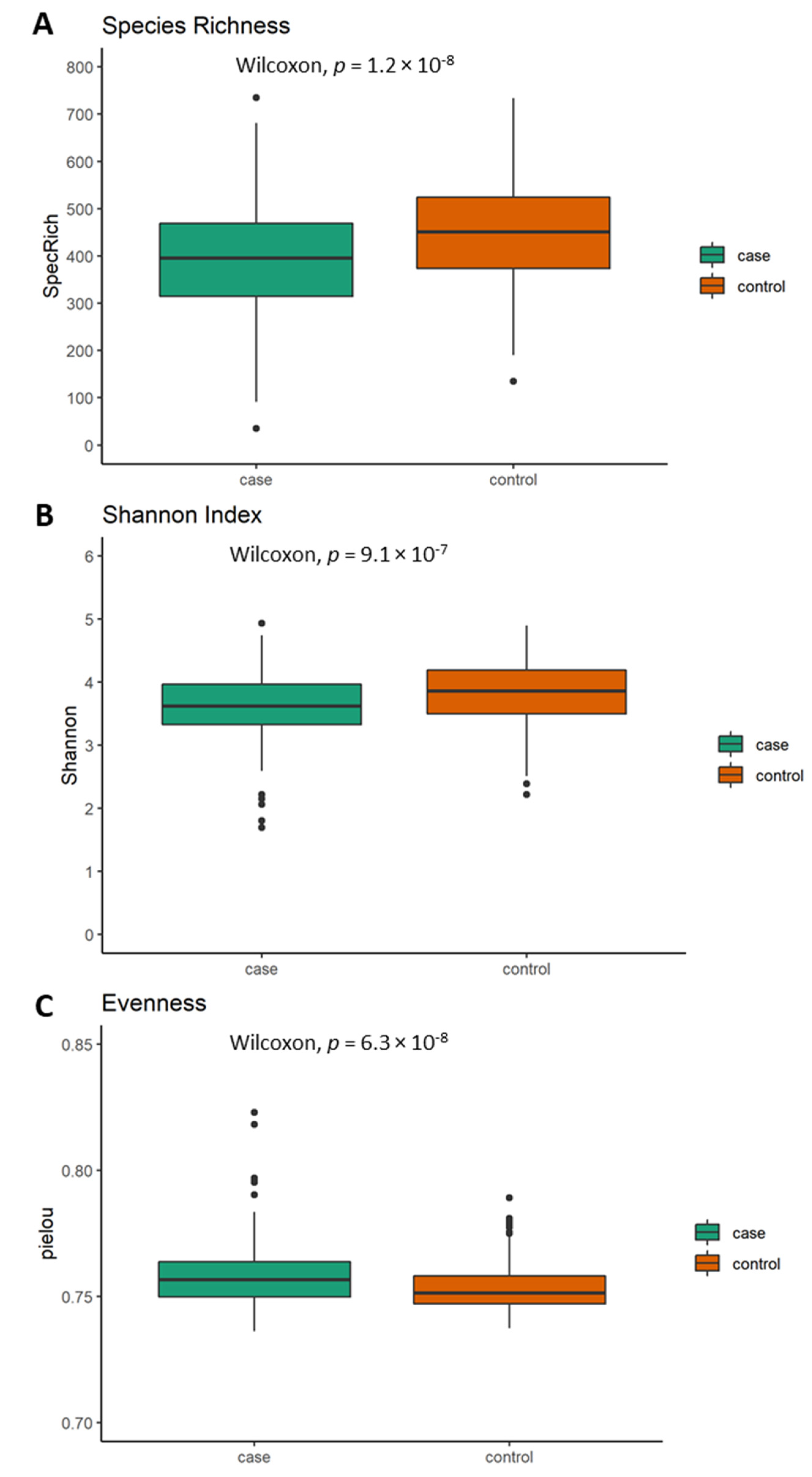
5.2. Differences in Alpha Diversity between Neurological Cases and Healthy Controls
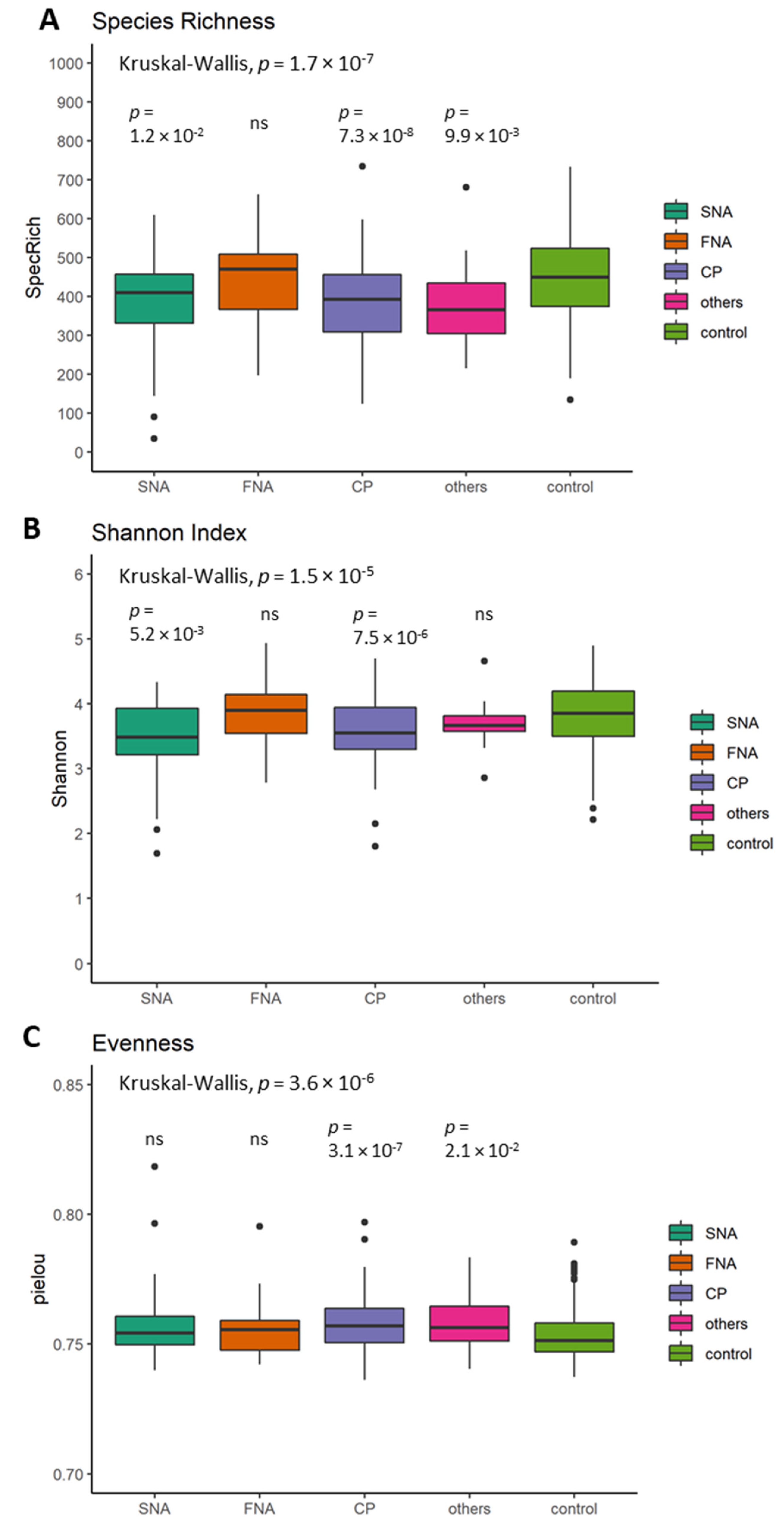
5.3. Differences in Alpha Diversity between Neurological Subgroups and Healthy Controls
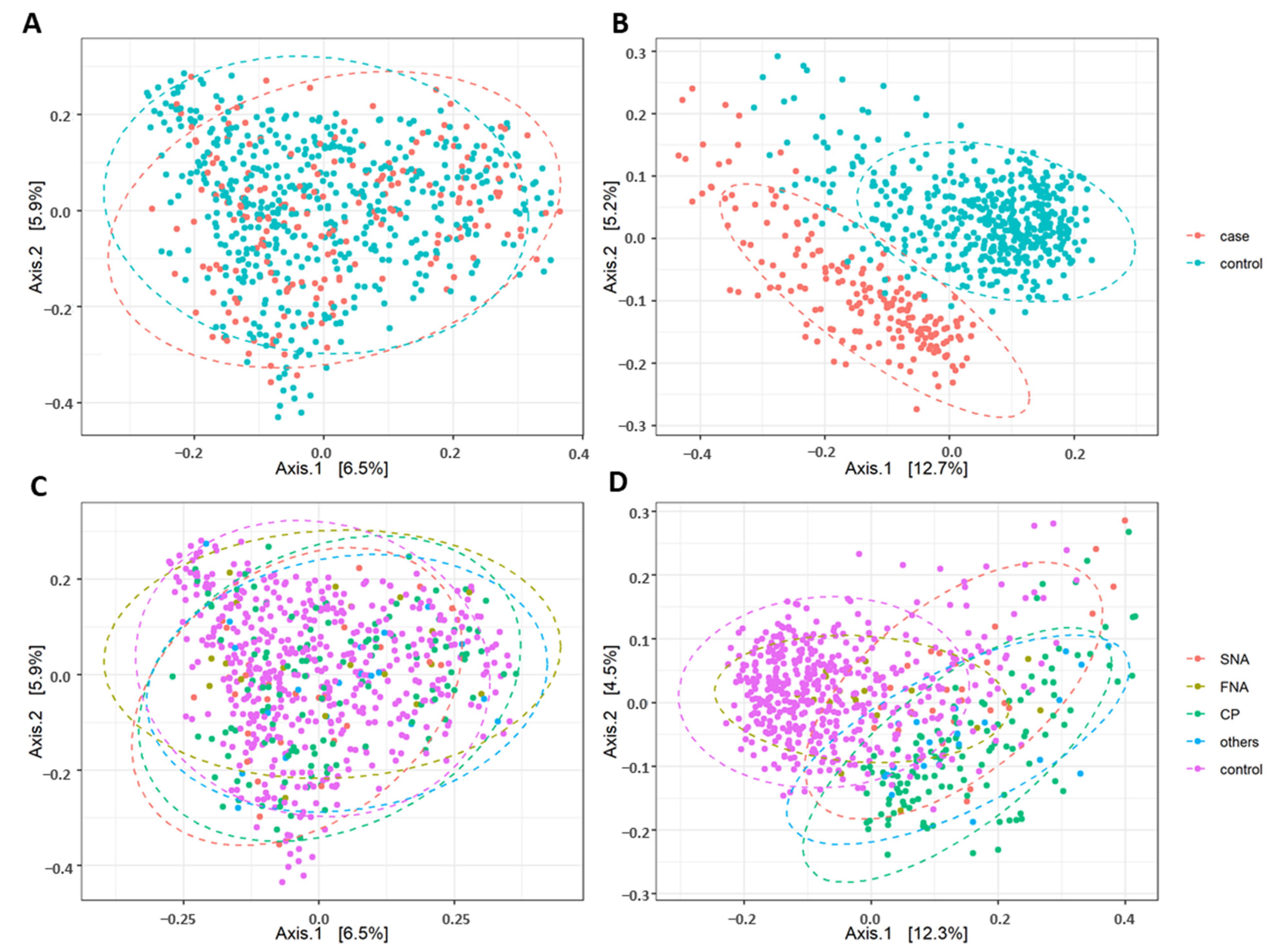
5.4. Difference in Beta Diversity between the Neurological Cases, their Subgroups and Healthy Controls
5.5. Prediction of Functional Profiles
5.6. Calculation of the Risk to Develop Neurological Diseases
6. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deuschl, G.; Beghi, E.; Fazekas, F.; Varga, T.; Christoforidi, A.K.; Sipido, E.; Bassetti, C.L.; Vos, T.; Feigin, V.L. The Burden of Neurological Diseases in Europe: An Analysis for the Global Burden of Disease Study 2017. Lancet Public Health 2020, 5, e551–e567. [Google Scholar] [CrossRef]
- Feigin, V.L.; Nichols, E.; Alam, T.; Bannick, M.S.; Beghi, E.; Blake, N.; Culpepper, W.J.; Dorsey, E.R.; Elbaz, A.; Ellenbogen, R.G.; et al. Global, Regional, and National Burden of Neurological Disorders, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef]
- Leussink, V.I. Aspekte der Ernährung zur Prävention und Therapie Chronisch Neurologischer Erkrankungen. Der Nervenarzt 2019, 90, 843–857. [Google Scholar] [CrossRef] [PubMed]
- Buttó, L.F.; Haller, D. Dysbiosis in Intestinal Inflammation: Cause or Consequence. Int. J. Med Microbiol. 2016, 306, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Sandhu, K.; Peterson, V.; Dinan, T.G. The Gut Microbiome in Neurological Disorders. Lancet Neurol. 2020, 19, 179–194. [Google Scholar] [CrossRef]
- Rhee, S.H.; Pothoulakis, C.; Mayer, E.A. Principles and Clinical Implications of the Brain–Gut–Enteric Microbiota Axis. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 306–314. [Google Scholar] [CrossRef]
- Kostic, A.; Xavier, R.J.; Gevers, D. The Microbiome in Inflammatory Bowel Disease: Current Status and the Future Ahead. Gastroenterology 2014, 146, 1489–1499. [Google Scholar] [CrossRef]
- Geuking, M.B.; Köller, Y.; Rupp, S.; McCoy, K. The Interplay Between the Gut Microbiota and the Immune System. Gut Microbes 2014, 5, 411–418. [Google Scholar] [CrossRef]
- Quigley, E.M.M. Microflora Modulation of Motility. J. Neurogastroenterol. Motil. 2011, 17, 140–147. [Google Scholar] [CrossRef]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus Strain Regulates Emotional Behavior and Central GABA Receptor Expression in a Mouse Via the Vagus Nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef]
- Collins, S.M.; Surette, M.G.; Bercik, P. The Interplay Between the Intestinal Microbiota and the Brain. Nat. Rev. Genet. 2012, 10, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, K.-A.; Foster, J.A. Effects of Gut Microbiota on the Brain: Implications for Psychiatry. J. Psychiatry Neurosci. 2009, 34, 230–231. [Google Scholar]
- Patrick, R.P.; Ames, B.N. Vitamin D and the Omega-3 Fatty Acids Control Serotonin Synthesis and Action, part 2: Relevance for ADHD, Bipolar Disorder, Schizophrenia, and Impulsive Behavior. FASEB J. 2015, 29, 2207–2222. [Google Scholar] [CrossRef] [PubMed]
- Schöndorf, D.C.; Ivanyuk, D.; Baden, P.; Sanchez-Martinez, A.; De Cicco, S.; Yu, C.; Giunta, I.; Schwarz, L.K.; Di Napoli, G.; Panagiotakopoulou, V.; et al. The NAD+Precursor Nicotinamide Riboside Rescues Mitochondrial Defects and Neuronal Loss in iPSC and Fly Models of Parkinson’s Disease. Cell Rep. 2018, 23, 2976–2988. [Google Scholar] [CrossRef]
- Müller, N.; Schulte, D.M.; Türk, K.; Freitag-Wolf, S.; Hampe, J.; Zeuner, R.; Schröder, J.O.; Gouni-Berthold, I.; Berthold, H.; Krone, W.; et al. IL-6 Blockade by Monoclonal Antibodies Inhibits Apolipoprotein (A) Expression and Lipoprotein (A) Synthesis in Humans. J. Lipid Res. 2015, 56, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Relling, I.; Akcay, G.; Fangmann, D.; Knappe, C.; Schulte, D.M.; Hartmann, K.; Müller, N.; Türk, K.; Dempfle, A.; Franke, A.; et al. Role of wnt5a in Metabolic Inflammation in Humans. J. Clin. Endocrinol. Metab. 2018, 103, 4253–4264. [Google Scholar] [CrossRef]
- Kaaks, R.; Slimani, N.; Riboli, E. Pilot Phase Studies on the Accuracy of Dietary Intake Measurements in the EPIC Project: Overall Evaluation of Results. European Prospective Investigation into Cancer and Nutrition. Int. J. Epidemiol. 1997, 26, 26S–36S. [Google Scholar] [CrossRef]
- Riboli, E.; Hunt, K.J.; Slimani, N.; Ferraria, P.; Norata, T.; Fahey, M.; Charrondierea, U.R.; Hemona, B.; Casagrandea, C.; Vignata, J.; et al. European Prospective Investigation into Cancer and Nutrition (EPIC): Study Populations and Data Collection. Public Health Nutr. 2002, 5, 1113–1124. [Google Scholar] [CrossRef]
- Deutsche Gesellschaft für Ernährung, Ö.G.f.E. , Schweizerische Gesellschaft für Ernährungsforschung, Schweizerische Vereinigung für Ernährung. Referenzwerte für die Nährstoffzufuhr, 2nd ed.; Neuer Umschau Buchverlag: Neustadt an der Weinstraße, Germany, 2019. [Google Scholar]
- Willett, W.C.; Howe, G.R.; Kushi, L. Adjustment for Total Energy Intake in Epidemiologic Studies. Am. J. Clin. Nutr. 1997, 65, 1220S–1228S. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.-C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome Definition Re-Visited: Old Concepts and New Challenges. Microbiome 2020, 8, 1–22. [Google Scholar] [CrossRef]
- Heinsen, F.-A.; Fangmann, D.; Müller, N.; Schulte, D.M.; Rühlemann, M.; Türk, K.; Settgast, U.; Lieb, W.; Baines, J.F.; Schreiber, S.; et al. Beneficial Effects of a Dietary Weight Loss Intervention on Human Gut Microbiome Diversity and Metabolism Are Not Sustained during Weight Maintenance. Obes. Facts 2016, 9, 379–391. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.; Highlander, S.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, A.W.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-High-Throughput Microbial Community Analysis on the Illumina HiSeq and MiSeq Platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. SINTAX: A Simple Non-Bayesian Taxonomy Classifier for 16S and ITS Sequences. bioRxiv 2016, 074161. [Google Scholar] [CrossRef]
- Wang, J.; Kurilshikov, A.; Radjabzadeh, D.; Turpin, W.; Croitoru, K.; Bonder, M.J.; Jackson, M.; Medina-Gomez, C.; Frost, F.; Homuth, G.; et al. Meta-Analysis of Human Genome-Microbiome Association Studies: The Mibiogen Consortium Initiative. Microbiome 2018, 6, 101. [Google Scholar] [CrossRef]
- Aßhauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting Functional Profiles from Metagenomic 16S rRNA data: Figure 1. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, I.J.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Liu, C.; Cui, Y.; Li, X.; Yao, M. Microeco: An R Package for Data Mining in Microbial Community Ecology. FEMS Microbiol. Ecol. 2021, 97, fiaa255. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Cournot, M.; Marquie, J.C.; Ansiau, D.; Martinaud, C.; Fonds, H.; Ferrieres, J.; Ruidavets, J.B. Relation between Body Mass Index and Cognitive Function in Healthy Middle-Aged Men and Women. Neurology 2006, 67, 1208–1214. [Google Scholar] [CrossRef] [PubMed]
- Bhat, Z.F.; Morton, J.D.; Mason, S.; Bekhit, A.E.-D.A.; Bhat, H. Obesity and Neurological Disorders: Dietary Perspective of a Global Menace. Crit. Rev. Food Sci. Nutr. 2019, 59, 1294–1310. [Google Scholar] [CrossRef]
- World Health Organisation. Body Mass Index. Available online: https://www.euro.who.int/en/health-topics/disease-prevention/nutrition/a-healthy-lifestyle/body-mass-index-bmi (accessed on 11 June 2020).
- Deutsche Gesellschaft für Ernährung e. V. Available online: https://www.dge.de/wissenschaft/referenzwerte/fett/ (accessed on 1 October 2020).
- Cedernaes, J.; Schönke, M.; Westholm, J.O.; Mi, J.; Chibalin, A.; Voisin, S.; Osler, M.; Vogel, H.; Hörnaeus, K.; Dickson, S.L.; et al. Acute Sleep Loss Results in Tissue-Specific Alterations in Genome-Wide DNA Methylation State and Metabolic Fuel Utilization in Humans. Sci. Adv. 2018, 4, eaar8590. [Google Scholar] [CrossRef]
- Hogg, E.; Athreya, K.; Basile, C.; Tan, E.E.; Kaminski, J.; Tagliati, M. High Prevalence of Undiagnosed Insulin Resistance in Non-Diabetic Subjects with Parkinson’s Disease. J. Park. Dis. 2018, 8, 259–265. [Google Scholar] [CrossRef]
- Lu, C.-L.; Chang, Y.-H.; Sun, Y.; Li, C.-Y. A Population-Based Study of Epilepsy Incidence in Association with Type 2 Diabetes and Severe Hypoglycaemia. Diabetes Res. Clin. Pr. 2018, 140, 97–106. [Google Scholar] [CrossRef]
- Pietrobon, D.; Moskowitz, M.A. Pathophysiology of Migraine. Annu. Rev. Physiol. 2013, 75, 365–391. [Google Scholar] [CrossRef]
- Frank-Cannon, T.C.; Alto, L.T.; McAlpine, F.E.; Tansey, M.G. Does Neuroinflammation Fan the Flame in Neurodegenerative Diseases? Mol. Neurodegener. 2009, 4, 47. [Google Scholar] [CrossRef]
- Ben-Or, O.; Zelnik, N.; Shaoul, R.; Pacht, A.; Lerner, A. The Neurologic Profile of Children and Adolescents with Inflammatory Bowel Disease. J. Child Neurol. 2015, 30, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.R.; Teles, B.C.; Brasil, F.; Souza, M.H.; Furtado, L.E.; De Castro-Costa, C.M.; Rola, F.H.; Braga, L.L.; Gondim, F.D.A. Peripheral Neuropathy and Neurological Disorders in an Unselected Brazilian Population-Based Cohort of IBD Patients. Inflamm. Bowel Dis. 2008, 14, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, T.; Jin, F. Alzheimer’s Disease and Gut Microbiota. Sci. China Life Sci. 2016, 59, 1006–1023. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.C.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean Diet Intervention Alters the Gut Microbiome in Older People Reducing Frailty and Improving Health Status: The NU-AGE 1-Year Dietary Intervention Across Five European Countries. Gut 2020, 69, 1218–1228. [Google Scholar] [CrossRef]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The Gut Microbiota Influences Blood-Brain Barrier Permeability in Mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef]
- Spadoni, I.; Fornasa, G.; Rescigno, M. Organ-Specific Protection Mediated by Cooperation Between Vascular and Epithelial Barriers. Nat. Rev. Immunol. 2017, 17, 761–773. [Google Scholar] [CrossRef]
- Li, C.; Cui, L.; Yang, Y.; Miao, J.; Zhao, X.; Zhang, J.; Cui, G.; Zhang, Y. Gut Microbiota Differs Between Parkinson’s Disease Patients and Healthy Controls in Northeast China. Front. Mol. Neurosci. 2019, 12, 171. [Google Scholar] [CrossRef]
- Li, Y.; Li, R.X.; Du, Y.T.; Xu, X.J.; Xue, Y.; Gao, D.; Gao, T.; Sheng, Z.; Zhang, L.Y.; Tuo, H.Z. Features of Gut Microbiota in Patients with Idiopathic Parkinson’s Disease. Zhonghua Yi Xue Za Zhi 2020, 100, 1017–1022. [Google Scholar] [CrossRef]
- Jacobson, K.; Gao, Z.-G. Adenosine Receptors as Therapeutic Targets. Nat. Rev. Drug Discov. 2006, 5, 247–264. [Google Scholar] [CrossRef]
- Sollevi, A. Adenosine for Pain Control. Acta Anaesthesiol. Scand. 1997, 41, 135–136. [Google Scholar] [CrossRef]
- Dresselhaus, E.C.; Meffert, M.K. Cellular Specificity of NF-κB Function in the Nervous System. Front. Immunol. 2019, 10, 1043. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Hottiger, M.O. Crosstalk between Wnt/β-Catenin and NF-κB Signaling Pathway during Inflammation. Front. Immunol. 2016, 7, 378. [Google Scholar] [CrossRef]
- Hamilton, J.L.; Nagao, M.; Levine, B.R.; Chen, D.; Olsen, B.R.; Im, H.-J. Targeting VEGF and Its Receptors for the Treatment of Osteoarthritis and Associated Pain. J. Bone Miner. Res. 2016, 31, 911–924. [Google Scholar] [CrossRef]
- Bussolati, B.; Dunk, C.; Grohman, M.; Kontos, C.; Mason, J.; Ahmed, A. Vascular Endothelial Growth Factor Receptor-1 Modulates Vascular Endothelial Growth Factor-Mediated Angiogenesis via Nitric Oxide. Am. J. Pathol. 2001, 159, 993–1008. [Google Scholar] [CrossRef]
- Straßburg, A. Ernährungserhebungen Methoden und Instrumente. In Ernährungs-Umschau; UMSCHAU ZEITSCHRIFTENVERLAG GmbH: Wiesbaden, Germany, 2010; pp. 422–430. [Google Scholar]
- Anstey, K.J.; Cherbuin, N.; Budge, M.; Young, J. Body Mass Index in Midlife and Late-Life as a Risk Factor for Dementia: A Meta-Analysis of Prospective Studies. Obes. Rev. 2011, 12, e426–e437. [Google Scholar] [CrossRef]
- O’Brien, P.D.; Hinder, L.M.; Callaghan, B.C.; Feldman, E.L. Neurological Consequences of Obesity. Lancet Neurol. 2017, 16, 465–477. [Google Scholar] [CrossRef]
- Sabia, S.; Kivimaki, M.; Shipley, M.J.; Marmot, M.G.; Singh-Manoux, A. Body Mass Index Over the Adult Life Course and Cognition in Late Midlife: The Whitehall II Cohort Study. Am. J. Clin. Nutr. 2009, 89, 601–607. [Google Scholar] [CrossRef]
- Dahl, A.K.; Hassing, L.B.; Fransson, E.; Gatz, M.; Reynolds, C.A.; Pedersen, N.L. Body Mass Index Across Midlife and Cognitive Change in Late Life. Int. J. Obes. 2013, 37, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Fitzpatrick, A.L.; Rapp, S.R.; Nahin, R.L.; Williamson, J.D.; Lopez, O.L.; DeKosky, S.T.; Kuller, L.H.; Mackey, R.H.; Mukamal, K.J.; et al. Alcohol Consumption and Risk of Dementia and Cognitive Decline Among Older Adults with or without Mild Cognitive Impairment. JAMA Netw. Open 2019, 2, e1910319. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Liu, X.; Yin, Q.; Zhu, W.; Zhang, R.; Fan, X. Alcohol Consumption and Transition of Mild Cognitive Impairment to Dementia. Psychiatry Clin. Neurosci. 2009, 63. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cases (n = 238) | Controls (n = 612) | All Subjects (n = 850) | |
|---|---|---|---|
| Women | 184 (77.3%) a | 391 (63.9%) | 575 (67.6%) |
| Men | 54 (22.7%) a | 221 (36.1%) | 275 (32.4%) |
| Neurological diseases | |||
| SNA | 15.1% | ||
| Parkinson’s disease | 1 (0.4%) | ||
| Tremor | 7 (2.9%) | ||
| Restless leg | 26 (10.9%) | ||
| Multiple sclerosis | 2 (0.8%) | ||
| FNA | 11.3% | ||
| Epilepsy | 16 (6.7%) | ||
| Anxiety | 8 (3.4%) | ||
| Depression | 2 (0.8%) | ||
| Sleep disorder | 1 (0.4%) | ||
| CP | 62.2% | ||
| Migraine | 135 (56.7%) | ||
| Pain | 12 (5.0%) | ||
| Neuropathy | 1 (0.4%) | ||
| Others | 27 (11.3%) | ||
| Age, year | 48.49 (±13.41) 1 | 46.73 (±14.68) | 47.23 (±14.34) |
| Weight, kg | 100.10 (±33.06) * | 84.82 (±26.66) | 89.10 (±29.40) |
| Height, cm | 171.18 (±8.41) * | 173.57 (±8.88) | 173.90 (±8.81) |
| BMI, kg/m2 | 34.14 (±10.85) * | 28.05 (±8.22) | 29.76 (±9.43) |
| UW | 1.8% (4) | 1.9% (11) | 1.9% (15) |
| NW | 22.4% (50) a | 44.5% (253) | 38.3% (303) |
| OW | 17.0% (38) a | 28.0% (159) | 24.9% (197) |
| OBI | 17.0% (38) a | 9.7% (55) | 11.8% (93) |
| OBII | 11.2% (25) a | 4.8% (27) | 6.6% (52) |
| OBIII | 30.6% (68) a | 11.1% (63) | 16.6% (131) |
| Glucose, mg/dL | 95.00 (88.00/105.00) *2 | 92.00 (87.00/99.00) | 93 (87.00/101.00) |
| Insulin, µU/mL | 12.20 (7.30/22.65) * | 8.50 (5.80/13.10) | 9.30 (6.20/15.30) |
| HOMA-IR | 2.93(1.61/5.79) * | 1.99 (1.30/3.08) | 2.13 (1.37/3.60) |
| Triglyceride, mg/dL | 119.00 (79.00/168.25) * | 87.00 (66.00/125.00) | 95.00 (68.75/139.00) |
| CRP, mg/L | 2.60 (0.90/6.40) * | 1.10 (0.90/2.80) | 1.30 (0.90/3.70) |
| IL-6, pg/mL | 3.65 (2.10/5.43) * | 2.60 (1.50/3.80) | 2.80 (1.60/4.35) |
| Lipoprotein-a, mg/L | 104.00 (95.00/245.00) * | 95.00 (93.10/200.00) | 95.00 (95.00/214.25) |
| Cases (n = 223) | Controls (n = 568) | All Subjects (n = 791) | |
|---|---|---|---|
| Protein total, E% | 14.52 (13.04/15.89) 1 | 14.38 (12.96/15.79) | 14.42 (12.98/15.85) |
| Essential amino acids, g/day | 34.87 (28.48/41.37) | 37.12 (33.60/41.43) | 37.23 (33.65/41.29) |
| Arginine, g/day | 4.17 (3.65/4.73) | 4.09 (3.61/4.57) | 4.11 (3.63/4.59) |
| Cysteine, g/day | 1.01 (0.92/1.10) | 0.99 (0.91/1.09) | 1.00 (0.91/1.09) |
| Tyrosine, g/d | 2.70 (2.41/2.93) | 2.67 (2.41/2.96) | 2.68 (2.41/2.95) |
| Methionine, g/d | 1.60 (1.44/1.80) | 1.61 (1.43/1.81) | 1.61 (1.44/1.81) |
| Phenylalanine, g/d | 3.18 (2.60/3.71) | 3.31 (3.04/3.61) | 3.33 (3.04/3.60) |
| Tryptophan, g/day | 0.86 (0.77/0.94) | 0.86 (0.78/0.94) | 0.86 (0.77/0.94) |
| Fat total, E% | 42.05 (37.74/45.25) | 41.29 (37.42/44.55) | 41.43 (37.60/44.82) |
| Saturated fatty acids, E% | 36.86 (28.80/44.04) | 37.74 (29.52/47.02) | 37.22 (29.25/46.20) |
| Short-chain fatty acids, E% | 0.81 (0.65/1.02) | 0.86 (0.66/1.07) | 0.84 (0.66/1.05) |
| Medium-chain fatty acids, g/d | 0.76 (0.65/0.90) | 0.80 (0.66/0.92) | 0.79 (0.66/0.91) |
| Long-chain fatty acids, E% | 37.57 (33.92/40.43) | 36.79 (33.36/39.89) | 36.90 (33.58/40.03) |
| Polyunsaturated fatty acids, E% | 7.37 (5.93/8.47) * | 7.04 (5.74/7.93) | 6.86 (5.80/8.14) |
| Octadecadienoic acid/linoleic acid, (g/day) | 14.28 (12.17/16.61) * | 13.53 (11.54/15.74) | 13.70 (11.70/16.02) |
| Octadecatrienoic acid/linolenic acid, (g/day) | 1.95 (1.79/2.28) | 1.93 (1.76/2.25) | 1.93 (1.77/2.26) |
| Eicosatetraenoic acid/arachidonic acid, (g/day) | 0.18 (0.15/0.22) | 0.18 (0.15/0.22) | 0.18 (0.15/0.22) |
| Docosahexaenoic acid, (g/day) | 0.18 (0.13/0.26) | 0.20 (0.13/0.27) | 0.19 (0.13/0.26) |
| Monounsaturated fatty acids, E% | 14.38 (13.00/16.02) | 14.37 (12.86/15.75) | 14.38 (12.91/15.79) |
| Carbohydrate total, E% | 42.99 (38.98/46.98) * | 41.46 (38.37/45.78) | 41.79 (38.55/46.08) |
| Monosaccharides, g/day | 49.00 (37.77/62.00) | 47.83 (35.91/59.79) | 48.12 (36.50/60.34) |
| Fructose, g/day | 26.33 (20.75/34.50) | 25.47 (19.75/33.59) | 25.63 (20.02/33.81) |
| Galactose, g/day | 4.23 (4.05/4.57) | 4.20 (4.05/4.46) | 4.21 (4.06/4.50) |
| Glucose, g/day | 21.85 (16.69/27.43) | 21.46 (15.89/26.13) | 21.57 (16.25/26.56) |
| Dietary fiber total, g/day | 21.62 (18.91/26.47) | 21.38 (18.67/25.22) | 21.51 (18.75/25.55) |
| Soluble fiber, g/day | 7.14 (6.03/8.47) | 6.94 (5.99/8.14) | 6.96 (6.00/8.21) |
| Insoluble fiber, g/day | 14.67 (12.69/17.61) | 14.33 (12.52/16.97) | 14.47 (12.55/17.22) |
| Alcohol, E% | 1.10 (0.42/2.80) * | 2.56 (1.02/5.10) | 2.18 (0.72/4.58) |
| Table salt, g/day | 5.54 (4.93/6.08) | 5.46 (4.97/5.99) | 5.48 (4.96/6.03) |
| Calcium, mg/day | 865.60 (749.60/1016.90) | 852.30 (737.10/989.50) | 857.50 (740.50/995.90) |
| Vitamin B1 (thiamine), mg/day | 1.76 (1.61/1.86) | 1.73 (1.61/1.85) | 1.74 (1.61/1.85) |
| Vitamin B12 (cobalamin), µg/day | 5.50 (4.40/6.40) | 5.70 (4.50/6.70) | 5.50 (4.40/6.70) |
| Vitamin B2 (riboflavin), mg/day | 1.51 (1.32/1.69) | 1.48 (1.33/1.67) | 1.49 (1.33/1.68) |
| Vitamin B3 (niacin), mg/day | 14.21 (12.13/16.75) | 14.77 (12.46/17.19) | 14.64 (12.33/17.00) |
| Vitamin B5 (pantothenic acid), mg/day | 4.55 (3.79/5.45) | 4.59 (3.81/5.63) | 4.58 (3.80/5.54) |
| Vitamin B6 (pyridoxine), mg/day | 1.55 (1.40/1.75) | 1.56 (1.40/1.73) | 1.55 (1.40/1.74) |
| Vitamin B7 (biotin), µg/day | 45.20 (40.30/49.80) | 44.70 (39.90/50.50) | 44.90 (40.00/50.50) |
| Vitamin B9 (free folic acid equivalent), µg/day | 110.30 (94.80/130.70) | 112.20 (95.30/125.70) | 110.80 (95.20/126.50) |
| Vitamin B9 (free folic acid), µg/day | 87.60 (72.10/100.00) | 84.60 (71.10/96.10) | 84.30 (71.70/96.80) |
| Vitamin B9 (total folic acid), µg/day | 275.00 (248.90/312.40) | 276.20 (249.90/305.50) | 276.00 (249.80/308.30) |
| Vitamin C (ascorbic acid), mg/day | 123.37 (98.28/162.98) | 118.09 (95.73/157.92) | 119.25 (96.36/158.86) |
| Vitamin D (calciferols), µg/day | 3.70 (2.70/4.90) * | 4.40 (2.90/5.20) | 3.90 (2.80/5.10) |
| Vitamin E (tocopherol equivalent), mg/day | 14.08 (11.61/15.80) | 13.69 (12.28/15.23) | 13.80 (12.36/15.39) |
| Vitamin K (phylloquinone), µg/day | 314.50 (274.90/357.00) | 305.30 (271.60/337.90) | 306.80 (271.90/342.50) |
| Bray–Curtis | Jaccard | |||||
|---|---|---|---|---|---|---|
| R2 | p-Value | Significance | R2 | p-Value | Significance | |
| Medication regularly | 4.04 × 10−3 | 9.99 × 10−4 | *** | 6.74 × 10−3 | 9.99 × 10−4 | *** |
| Smoking habits | 3.26 × 10−3 | 9.99 × 10−4 | *** | 3.77 × 10−3 | 9.99 × 10−4 | *** |
| Alcohol | 1.78 × 10−3 | 1.47 × 10−1 | - | 1.96 × 10−3 | 1.28 × 10−1 | - |
| Dietary fiber | 3.44 × 10−3 | 9.99 × 10−4 | *** | 3.35 × 10−3 | 2.99 × 10−3 | ** |
| Docosahexaenoic acid | 2.12 × 10−3 | 3.10 × 10−2 | * | 2.20 × 10−3 | 8.19 × 10−2 | - |
| Vitamin B12 | 2.57 × 10−3 | 9.99 × 10−4 | *** | 2.77 × 10−3 | 1.19 × 10−2 | * |
| Vitamin B3 | 2.06 × 10−3 | 3.99 × 10−2 | * | 2.12 × 10−3 | 1.08 × 10−1 | - |
| Vitamin D | 1.69 × 10−3 | 2.01 × 10−1 | - | 1.79 × 10−3 | 2.00 × 10−1 | - |
| BMI | 4.13 × 10−3 | 9.99 × 10−4 | *** | 4.61 × 10−3 | 9.99 × 10−4 | *** |
| Age | 4.10 × 10−3 | 9.99 × 10−4 | *** | 4.99 × 10−3 | 9.99 × 10−4 | *** |
| Sex | 2.63 × 10−3 | 2.99 × 10−3 | ** | 2.42 × 10−3 | 3.40 × 10−2 | * |
| Group membership | 3.31 × 10−3 | 9.99 × 10−4 | *** | 1.03 × 10−2 | 9.99 × 10−4 | *** |
| Bray–Curtis | Jaccard | |||||
|---|---|---|---|---|---|---|
| R2 | p-Value | Significance | R2 | p-Value | Significance | |
| Medication regularly | 3.95 × 10−3 | 9.99 × 10−4 | *** | 5.99 × 10−3 | 9.99 × 10−4 | *** |
| Smoking habits | 3.26 × 10−3 | 9.99 × 10−4 | *** | 3.69 × 10−3 | 9.99 × 10−4 | *** |
| Alcohol | 1.76 × 10−3 | 1.64 × 10−1 | - | 1.82 × 10−3 | 1.99 × 10−1 | - |
| Dietary fiber | 3.65 × 10−3 | 9.99 × 10−4 | *** | 3.67 × 10−3 | 1.99 × 10−3 | ** |
| Docosahexaenoic acid | 2.10 × 10−3 | 2.99 × 10−2 | * | 2.29 × 10−3 | 5.39 × 10−2 | |
| Vitamin B12 | 2.56 × 10−3 | 3.99 × 10−3 | ** | 2.67 × 10−3 | 2.49 × 10−2 | * |
| Vitamin B3 | 2.16 × 10−3 | 2.59 × 10−2 | * | 2.37 × 10−3 | 3.49 × 10−2 | * |
| Vitamin D | 1.69 × 10−3 | 2.16 × 10−1 | - | 1.74 × 10−3 | 2.32 × 10−1 | - |
| BMI | 4.29 × 10−3 | 9.99 × 10−4 | *** | 4.79 × 10−3 | 9.99 × 10−4 | *** |
| Age | 4.07 × 10−3 | 9.99 × 10−4 | *** | 4.92 × 10−3 | 9.99 × 10−4 | *** |
| Sex | 2.77 × 10−3 | 1.99 × 10−3 | ** | 2.52 × 10−3 | 3.29 × 10−2 | * |
| Group membership | 8.09 × 10−3 | 9.99 × 10−4 | *** | 1.71 × 10−2 | 9.99 × 10−4 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geisler, C.; Pankoke, J.; Schlicht, K.; Knappe, C.; Rohmann, N.; Hartmann, K.; Settgast, U.; Türk, K.; Seoudy, A.K.; Franke, A.; et al. BMI, Alcohol Consumption and Gut Microbiome Species Richness Are Related to Structural and Functional Neurological Abnormalities. Nutrients 2021, 13, 3743. https://doi.org/10.3390/nu13113743
Geisler C, Pankoke J, Schlicht K, Knappe C, Rohmann N, Hartmann K, Settgast U, Türk K, Seoudy AK, Franke A, et al. BMI, Alcohol Consumption and Gut Microbiome Species Richness Are Related to Structural and Functional Neurological Abnormalities. Nutrients. 2021; 13(11):3743. https://doi.org/10.3390/nu13113743
Chicago/Turabian StyleGeisler, Corinna, Jil Pankoke, Kristina Schlicht, Carina Knappe, Nathalie Rohmann, Katharina Hartmann, Ute Settgast, Kathrin Türk, Anna Katharina Seoudy, Andre Franke, and et al. 2021. "BMI, Alcohol Consumption and Gut Microbiome Species Richness Are Related to Structural and Functional Neurological Abnormalities" Nutrients 13, no. 11: 3743. https://doi.org/10.3390/nu13113743