Preventive Role of L-Carnitine and Balanced Diet in Alzheimer’s Disease

 ,
, 
Abstract
:1. Introduction
2. AD Epidemiology
3. Prevention of Alzheimer’s Disease—Mediterranean and MIND Diets
4. Physiological Properties of L-Carnitine
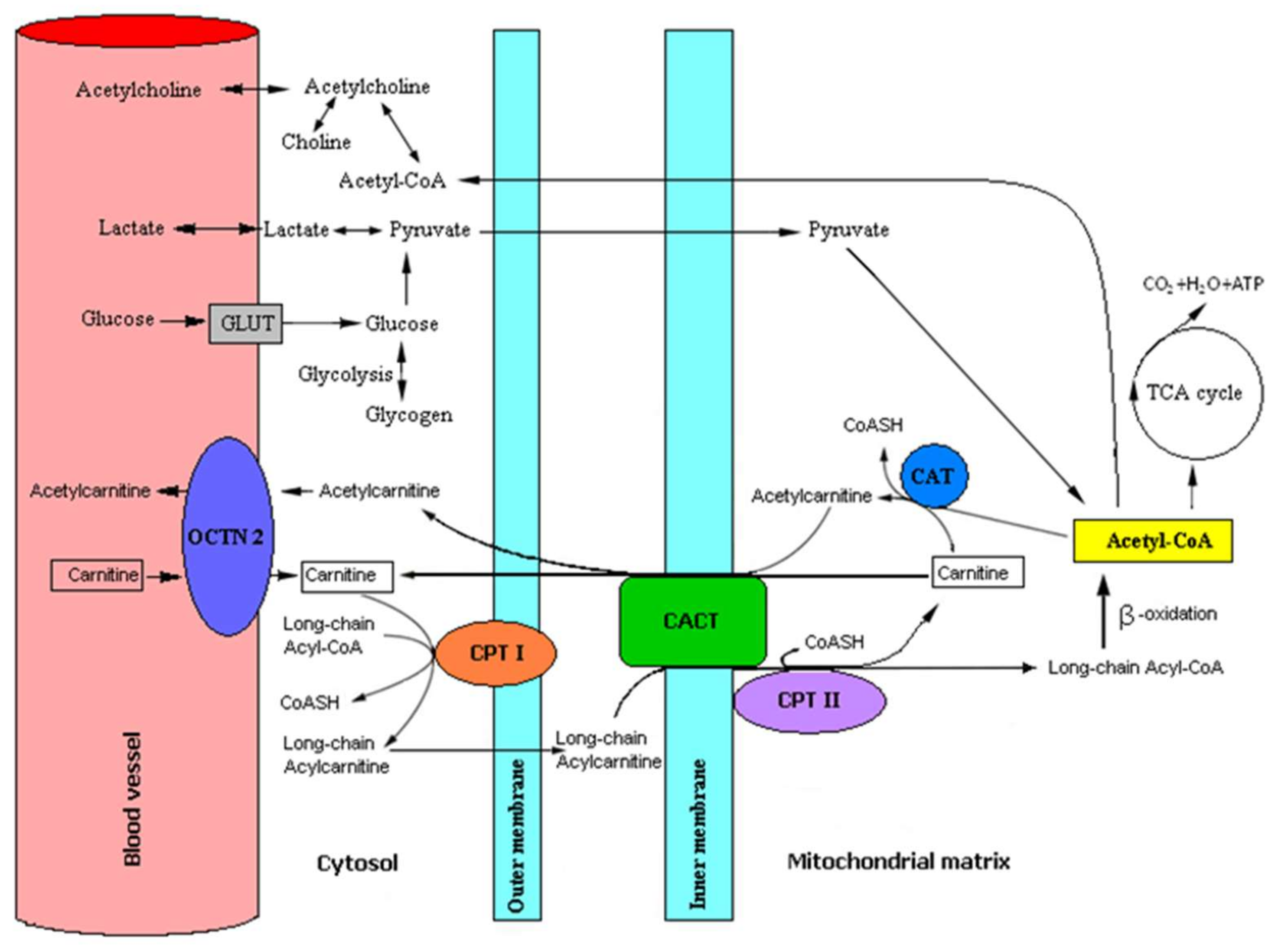
4.1. Role of L-Carnitine and Acetyl-L-Carnitine in Human Brain Metabolism
4.2. Recommendations for L-Carnitine Content in the Diet

{kind=link}
| Type of Food | Total L-Carnitine Content |
|---|---|
| Ruminant meat | (mg/100 g) |
| kangaroo meat | 637 |
| horseflesh | 423 |
| beef | 98.2–139 |
| beef steak | 232 |
| beef kidneys | 31.0 |
| beef liver | 15.6 |
| lamb | 106–113 |
| goat meat | 95.0–99.0 |
| pork | 20.0–30.0 |
| pork liver | 10.7 |
| Poultry, bird meat | (mg/100 g) |
| duck | 73.0 |
| pigeon | 52.8 |
| turkey | 51.0 |
| chicken | 34.0 |
| quail | 29.1 |
| pheasant | 13.5 |
| Fish | (mg/100 g) |
| salmon | 5.96 |
| zebrafish | 2.80–8.95 |
| yellow catfish | 5.93 |
| Milk | (mg/100 mL) |
| sheep | 10.2–12.7 |
| goat | 4.50–7.50 |
| cow | 7.80–9.60 |
| Milk products | (mg/100 g) |
| yoghurt | 40.0 |
| buttermilk | 38.0 |
| cottage cheese | 22.5–26.6 |
| sour cream | 19.7 |
| coffee cream | 16.6 |
| cheese | 14.0–28.0 |
| Mushrooms | (mg/100 g) |
| Pleureotus ostreatus—oyster mushrooms | 53.0 |
| champignon | 29.8 |
| Cantharellus cibarius—chanterelle | 13.3 |
| other mushrooms | 1.00–6.00 |
| Vegetables | (mg/100 g) |
| cucumber | 4.45 |
| cauliflower | 3.26 |
| carrot | 3.73 |
| maize | 0.68 |
| peas | 0.60 |
| Fruits | (mg/100 g) |
| avocado | 1.72 |
| guava | 0.82 |
| bananas | 0.39 |
| apples | 0.29 |
| orange | 0.22 |
4.3. Supplementation with L-Carnitine and Its Derivatives
4.4. Choosing Proper Form of Carnitine Supplements
4.4.1. Pure L-Carnitine (LC)
4.4.2. Acetyl L-Carnitine (ALC)
4.4.3. Propionyl L-Carnitine (PLCAR)
4.4.4. Acetyl L-Carnitine Arginate (ALCA)
4.4.5. Glycine-Propionyl- L-carnitine (GPLC)
4.4.6. L-Carnitine-L-Tartate (LCLT)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Strafella, C.; Caputo, V.; Galota, M.R.; Zampatti, S.; Marella, G.; Mauriello, S.; Cascella, R.; Giardina, E. Application of precision medicine in neurodegenerative diseases. Front. Neurol. 2018, 9, 701:1–701:9. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, C.; Bates, T.E.; Butterfield, D.A.; Calafato, S.; Cornelius, C.; De Lorenzo, A.; Dinkova Kostova, A.T.; Calabrese, V. Natural antioxidants in Alzheimer’s disease. Expert Opin. Investig. Drugs 2007, 16, 1921–1931. [Google Scholar] [CrossRef] [PubMed]
- Winiarska-Mieczan, A.; Baranowska-Wójcik, E.; Kwiecień, M.; Grela, E.R.; Szwajgier, D.; Kwiatkowska, K.; Kiczorowska, B. The role of dietary antioxidants in the pathogenesis of neurodegenerative diseases and their impact on cerebral oxidoreductive balance. Nutrients 2020, 12, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenk, G.L. Neuropathologic changes in Alzheimer’s disease. J. Clin. Psychiatr. 2003, 64, 7–10. [Google Scholar]
- Schliebs, R.; Arendt, T. The cholinergic system in aging and neuronal degeneration. Behav. Brain Res. 2011, 221, 555–563. [Google Scholar] [CrossRef]
- Ouerfurth, H.W.; LaFerla, F.M. Alzheimer’s disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [Green Version]
- Kaźmierczak, A.; Adamczyk, A.; Benigna-Strosznajder, J. The role of extracellular α-synuclein in molecular mechanisms of cell heath. Post. Hig. Med. Dosw. 2013, 67, 1047–1057. [Google Scholar] [CrossRef]
- Dean, D.C., 3rd; Jerskey, B.A.; Chen, K.; Protas, H.; Thiyyagura, P.; Roontiva, A.; O’Muircheartaigh, J.; Dirks, H.; Waskiewicz, N.; Lehman, K.; et al. Brain differences in infants at differential genetic risk for late-onset Alzheimer disease: A cross-sectional imaging study. JAMA Neurol. 2014, 71, 11–22. [Google Scholar] [CrossRef]
- Ng, S.; Lin, C.C.; Hwang, Y.H.; Hsieh, W.S.; Liao, H.F.; Chen, P.C. Mercury, APOE, and children’s neurodevelopment. Neurotoxicology 2013, 37, 85–92. [Google Scholar] [CrossRef]
- Dehghani, C.; Frost, S.; Jayasena, R.; Masters, C.L.; Kanagasingam, Y. Ocular biomarkers of Alzheimer’s disease: The role of anterior eye and potential future directions. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3554–3563. [Google Scholar] [CrossRef] [Green Version]
- Nunes-Tavares, N.; Santos, L.E.; Stutz, B.; Brito-Moreira, J.; Klein, W.L.; Ferreira, S.T.; de Mello, F.G. Inhibition of choline acetyltransferase as a mechanism for cholinergic dysfunction induced by amyloid-β peptide oligomers. J. Biol. Chem. 2012, 287, 19377–19385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szutowicz, A.; Bielarczyk, H.; Jankowska-Kulawy, A.; Pawełczyk, T.; Ronowska, A. Acetyl-CoA the key factor for survival or death of cholinergic neurons in course of neurodegenerative diseases. Neurochem. Res. 2013, 38, 1523–1542. [Google Scholar] [CrossRef] [Green Version]
- Lodeiro, M.; Ibáñez, C.; Cifuentes, A.; Simó, C.; Cedazo-Mínguez, Á. Decreased cerebrospinal fluid levels of L-carnitine in non-apolipoprotein E4 carriers at early stages of Alzheimer’s disease. J. Alzheimers Dis. 2014, 41, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Nasaruddin, M.B.; Elliott, C.T.; McGuinness, B.; Passmore, A.P.; Kehoe, P.G.; Hölscher, C.; McClean, P.L.; Graham, S.F.; Green, B.D. Alzheimer’s disease-like pathology has transient effects on the brain and blood metabolome. Neurobiol. Aging 2016, 38, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.C.; Alhasawi, A.; Appanna, V.P.; Auger, C.; Appanna, V.D. Brain metabolism and Alzheimer’s disease: The prospect of a metabolite-based therapy. J. Nutr. Health Aging 2015, 19, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Cristofano, A.; Sapere, N.; La Marca, G.; Angiolillo, A.; Vitale, M.; Corbi, G.; Scapagnini, G.; Intrieri, M.; Russo, C.; Corso, G.; et al. Serum levels of acyl-carnitines along the continuum from normal to Alzheimer’s dementia. PLoS ONE 2016, 11, e0155694. [Google Scholar] [CrossRef]
- White, H.L.; Scates, P.W. Acetyl-L-carnitine as a precursor of acetylcholine. Neurochem. Res. 1990, 15, 597–601. [Google Scholar] [CrossRef]
- Traina, G. The neurobiology of acetyl-L-carnitine. Front Biosci. (Landmark Ed) 2016, 21, 1314–1329. [Google Scholar] [CrossRef] [Green Version]
- Calvani, M.; Carta, A.; Caruso, G.; Benedetti, N.; Iannuccelli, M. Action of acetyl-L-carnitine in neurodegeneration and Alzheimer’s disease. Ann. N. Y. Acad. Sci. 1992, 663, 483–486. [Google Scholar] [CrossRef]
- Juliet, P.A.; Balasubramaniam, D.; Balasubramaniam, N.; Panneerselvam, C. Carnitine: A neuromodulator in aged rats. J. Gerontol. A- Biol. Sci. Med. Sci. 2003, 58, 970–974. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Chen, Z.; Zhao, N.; Liu, D.; Guo, Z.Y.; Tan, L.; Hu, J.; Wang, Q.; Wang, J.Z.; Zhu, L.Q. Acetyl-L-carnitine attenuates homocysteine-induced Alzheimer-like histopathological and behavioral abnormalities. Rejuvenation Res. 2011, 14, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Suchy, J.; Chan, A.; Shea, T.B. Dietary supplementation with a combination of alpha-lipoic acid, acetyl-L-carnitine, glycerophosphocholine, docosahexaenoic acid, and phosphatidylserine reduces oxidative damage to murine brain and improves cognitive performance. Nutr. Res. 2009, 29, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Pettegrew, J.W.; Klunk, W.E.; Panchalingam, K.; Kanfer, J.N.; McClure, R.J. Clinical and neurochemical effects of acetyl-L-carnitine in Alzheimer’s disease. Neurobiol. Aging 1995, 16, 1–4. [Google Scholar] [CrossRef]
- Onofrj, M.; Ciccocioppo, F.; Varanese, S.; di Muzio, A.; Calvani, M.; Chiechio, S.; Osio, M.; Thomas, A. Acetyl-L-carnitine: From a biological curiosity to a drug for the peripheral nervous system and beyond. Expert Rev. Neurother. 2013, 13, 925–936. [Google Scholar] [CrossRef]
- Grodzicki, W.; Dziendzikowska, K. The Role of selected Bioactive Compounds in the Prevention of Alzheimer’s Disease. Antioxidants 2020, 9, 229. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Muniz, F.J.; Macho-González, A.; Garcimartín, A.; Santos-López, J.A.; Benedí, J.; Bastida, S.; González-Muñoz, M.J. The nutritional components of beer and its relationship with neurodegeneration and Alzheimer’s disease. Nutrients 2019, 11, 1558. [Google Scholar] [CrossRef] [Green Version]
- De Boer, B.; Hamers, J.P.H.; Zwakhalen, S.M.G.; Tan, F.E.S.; Verbeek, H. Quality of care and quality of life of people with dementia living at green care farms: A cross-sectional study. BMC Geriatr. 2017, 17, 155. [Google Scholar] [CrossRef]
- Gonçalves-Pereira, M.; Cardoso, A.; Verdelho, A.; Alves da Silva, J.; Caldas de Almeida, M.; Fernandes, A.; Raminhos, C.; Ferri, C.P.; Prina, A.M.; Prince, M.; et al. The prevalence of dementia in a Portuguese community sample: A 10/66 Dementia Research Group study. BMC Geriatr. 2017, 17, 261. [Google Scholar] [CrossRef]
- World Health Organization. Neurological Disorders: Public Health Challenges; World Health Organization: Geneva, Switzerland, 2006; pp. 204–207. [Google Scholar]
- Ferri, C.P.; Prince, M.; Brayne, C.; Brodaty, H.; Fratiglioni, L.; Ganguli, M.; Hall, K.; Hasegawa, K.; Hendrie, H.; Huang, Y.; et al. Global prevalence of dementia: A Delphi consensus study. Lancet 2005, 366, 2112–2117. [Google Scholar] [CrossRef]
- Fillit, H.M. The pharmacoeconomics of Alzheimer’s disease. Am. J. Manag. Care 2000, 6, S1139–S1144. [Google Scholar]
- Moore, K.; Hughes, C.F.; Ward, M.; Hoey, L.; McNulty, H. Diet, nutrition and the ageing brain: Current evidence and new directions. Proc. Nutr. Soc. 2018, 77, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Gabryelewicz, T. Prevalence of dementia syndromes among the residents of the Mokotow district of Warsaw, aged 65–84 (in Polish). Psychiatr. Pol. 1999, 3, 353–366. [Google Scholar]
- Bdzan, L.B.; Turczyński, J.; Szabert, K. Prevalence of dementia in a rural population. Psychiatr. Pol. 2007, 41, 181–188. [Google Scholar] [PubMed]
- Principal Polish Statistical Office. Demographic Yearbook of Poland; Principal Polish Statistical Office: Warsaw, Poland, 2004; p. 207. (In Polish)
- Szczudlik, A.; Barcikowska-Kotowicz, M.; Gabryelewicz, T.; Opala, G.; Parnowski, T.; Kuźnicki, J.; Rossa, A.; Sadowska, A. The Situation of People with Alzheimer’s Disease in Poland; RPO Report; Szczudlik, A., Ed.; RPO: Warsaw, Poland, 2016. (In Polish)
- Shah, H.; Albanese, E.; Duggan, C.; Rudan, I.; Langa, K.M.; Carrillo, M.C.; Chan, K.Y.; Joanette, Y.; Prince, M.; Rossor, M.; et al. Research priorities to reduce the global burden of dementia by 2025. Lancet Neurol. 2016, 15, 1285–1294. [Google Scholar] [CrossRef]
- Perrone, L.; Grant, W.B. Observational and ecological studies of dietary advanced glycation end products in national diets and Alzheimer’s disease incidence and prevalence. J. Alzheimers Dis. 2015, 45, 965–979. [Google Scholar] [CrossRef]
- Lubitz, I.; Ricny, J.; Atrakchi-Baranes, D.; Shemesh, C.; Kravitz, E.; Liraz-Zaltsman, S.; Maksin-Matveev, A.; Cooper, I.; Leibowitz, A.; Uribarri, J.; et al. High dietary advanced glycation end products are associated with poorer spatial learning and accelerated Aβ deposition in an Alzheimer mouse model. Aging Cell 2016, 15, 309–316. [Google Scholar] [CrossRef]
- Solfrizzi, V.; Panza, F.; Frisardi, V.; Seripa, D.; Logroscino, G.; Imbimbo, B.P.; Pilotto, A. Diet and Alzheimer’s disease risk factors or prevention: The current evidence. Expert Rev. Neurother. 2011, 11, 677–708. [Google Scholar] [CrossRef]
- Anastasiou, C.A.; Yannakoulia, M.; Kosmidis, M.H.; Dardiotis, E.; Hadjigeorgiou, G.M.; Sakka, P.; Arampatzi, X.; Bougea, A.; Labropoulos, I.; Scarmeas, N. Mediterranean diet and cognitive health: Initial results from the hellenic longitudinal investigation of ageing and diet. PLoS ONE 2017, 12, e0182048. [Google Scholar] [CrossRef] [Green Version]
- GBD 2016 DALYs and HALE Collaborators. Global, regional, and national disability-adjusted life-years (DALYs) for 333 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990-2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1260–1344. [Google Scholar] [CrossRef] [Green Version]
- Yusufov, M.; Weyandt, L.L.; Piryatinsky, I. Alzheimer’s disease and diet: A systematic review. Int. J. Neurosci. 2017, 127, 161–175. [Google Scholar] [CrossRef]
- Singh, B.; Parsaik, A.K.; Mielke, M.M.; Erwin, P.J.; Knopman, D.S.; Petersen, R.C.; Roberts, R.O. Association of mediterranean diet with mild cognitive impairment and Alzheimer’s disease: A systematic review and meta-analysis. J. Alzheimers Dis. 2014, 39, 271–822. [Google Scholar] [CrossRef] [Green Version]
- Dochniak, M.; Ekiert, K. Nutrition in prevention and treatment of Alzheimer’s and Parkinson’s diseases. Nurs. Public Health 2015, 5, 199–208. (In Polish) [Google Scholar]
- Parnowski, T. Alzheimer’s Disease; Wydawnictwo Lekarskie PZWL: Warsaw, Poland, 2010. (In Polish) [Google Scholar]
- Abbatecola, A.M.; Russo, M.; Barbieri, M. Dietary patterns and cognition in older persons. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Tangney, C.C.; Wang, Y.; Sacks, F.M.; Bennett, D.A.; Aggarwal, N.T. MIND diet associated with reduced incidence of Alzheimer’s disease. Alzheimers Dement. 2015, 11, 1007–1014. [Google Scholar] [CrossRef] [Green Version]
- Stefani, M.; Rigacci, S. Beneficial properties of natural phenols: Highlight on protection against pathological conditions associated with amyloid aggregation. Biofactors 2014, 40, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Kępka, A.; Szajda, S.D.; Waszkiewicz, N.; Płudowski, P.; Chojnowska, S.; Rudy, M.; Szulc, A.; Ładny, J.R.; Zwierz, K. Carnitine: Function, metabolism and value in hepatic failure during chronic alcohol intoxication. Post. Hig. Med. Dosw. 2011, 65, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Czeczot, H.; Ścibior, D. Role of L-carnitine in metabolism, nutrition and therapy. Post. Hig. Med. Dosw. 2005, 59, 9–19. [Google Scholar]
- Kępka, A.; Chojnowska, S.; Okungbowa, O.E.; Zwierz, K. The role of carnitine in the perinatal period. Dev. Period Med. 2014, 18, 417–425. [Google Scholar]
- Bourdin, B.; Adenier, H.; Perrin, Y. Carnitine is associated with fatty acid metabolism in plants. Plant. Physiol. Biochem. 2007, 45, 926–931. [Google Scholar] [CrossRef]
- Hurot, J.M.; Cucherat, M.; Haugh, M.; Fouque, D. Effects of L-carnitine supplementation in maintenance hemodialysis patients: A systematic review. J. Am. Soc. Nephrol. 2002, 13, 708–714. [Google Scholar]
- Oey, N.A.; van Vlies, N.; Wijburg, F.A.; Wanders, R.J.; Attie-Bitach, T.; Vaz, F.M. L-carnitine is synthesized in the human fetal-placental unit: Potential roles in placental and fetal metabolism. Placenta 2006, 27, 841–846. [Google Scholar] [CrossRef]
- Rebouche, C.J. Kinetics, pharmacokinetics, and regulation of L-carnitine and acetyl-L-carnitine metabolism. Ann. N. Y. Acad. Sci. 2004, 1033, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Bain, M.A.; Fornasini, G.; Evans, A.M. Trimethylamine: Metabolic, pharmacokinetic and safety aspects. Curr. Drug Metab. 2005, 6, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Zimetti, F.; Caffarra, P.; Tassotti, M.; Bernini, F.; Brighenti, F.; Zini, A.; Zanotti, I. The gut microbial metabolite trimethylamine-N-oxide is present in human cerebrospinal fluid. Nutrients 2017, 9, 1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, A.M.; Fornasini, G. Pharmacokinetics of L-carnitine. Clin. Pharmacokinet. 2003, 42, 941–967. [Google Scholar] [CrossRef]
- Lavon, L. Perturbation of serum carnitine levels in human adults by chronic renal disease and dialysis therapy. Am. J. Clin. Nutr. 1981, 34, 1314–1320. [Google Scholar]
- Bodkowski, R.; Patkowska-Sokoła, B.; Nowakowski, P.; Jamroz, D.; Janczak, M. Products of animal origin—The most important L-carnitine source in human diet (in Polish). Prz. Hod. 2011, 10, 22–25. [Google Scholar]
- Jones, L.L.; McDonald, D.A.; Borum, P.R. Acylcarnitines: Role in brain. Prog. Lipid Res. 2010, 49, 61–75. [Google Scholar] [CrossRef]
- Calvani, M.; Benatti, P.; Mancinelli, A.; D’Iddio, S.; Giordano, V.; Koverech, A.; Amato, A.; Brass, E.P. Carnitine replacement in end-stage renal disease and hemodialysis. Ann. N. Y. Acad. Sci. 2004, 1033, 52–66. [Google Scholar] [CrossRef]
- Jacob, C.; Belleville, F. L-carnitine: Metabolism, function and value in pathology. Pathol. Biol. (Paris) 1992, 40, 910–919. [Google Scholar]
- Hedayati, S.S. Dialysis-related carnitine disorder. Semin. Dial. 2006, 19, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Ribas, G.S.; Vargas, C.R.; Wajner, M. L-carnitine supplementation as a potential antioxidant therapy for inherited neurometabolic disorders. Gene 2014, 533, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Valero, T. Mitochondrial biogenesis: Pharmacological approaches. Curr. Pharm. Des. 2014, 20, 5507–5509. [Google Scholar] [CrossRef] [PubMed]
- Zammit, V.A.; Ramsay, R.R.; Bonomini, M.; Arduini, A. Carnitine, mitochondrial function and therapy. Adv. Drug Deliv. Rev. 2009, 61, 1353–1362. [Google Scholar] [CrossRef] [PubMed]
- Aureli, T.; Miccheli, A.; Ricciolini, R.; Di Cocco, M.E.; Ramacci, M.T.; Angelucci, L.; Ghirardi, O.; Conti, F. Aging brain: Effect of acetyl-L-carnitine treatment on rat brain energy and phospholipid metabolism. A study by 31P and 1H NMR spectroscopy. Brain Res. 1990, 526, 108–112. [Google Scholar] [CrossRef]
- Aureli, T.; Di Cocco, M.E.; Puccetti, C.; Ricciolini, R.; Scalibastri, M.; Miccheli, A.; Manetti, C.; Conti, F. Acetyl-L-carnitine modulates glucose metabolism and stimulates glycogen synthesis in rat brain. Brain Res. 1998, 796, 75–81. [Google Scholar] [CrossRef]
- Ferreira, G.C.; McKenna, M.C. L-Carnitine and acetyl-L-carnitine roles and neuroprotection in developing brain. Neurochem. Res. 2017, 42, 1661–1675. [Google Scholar] [CrossRef]
- Gorini, A.; D’Angelo, A.; Villa, R.F. Action of L-acetylcarnitine on different cerebral mitochondrial populations from cerebral cortex. Neurochem. Res. 1998, 23, 1485–1491. [Google Scholar] [CrossRef]
- Calabrese, V.; Giuffrida Stella, A.M.; Calvani, M.; Butterfield, D.A. Acetylcarnitine and cellular stress response: Roles in nutritional redox homeostasis and regulation of longevity genes. J. Nutr. Biochem. 2006, 17, 73–88. [Google Scholar] [CrossRef]
- Madiraju, P.; Pande, S.V.; Prentki, M.; Madiraju, S.R. Mitochondrial acetylcarnitine provides acetyl groups for nuclear histone acetylation. Epigenetics 2009, 4, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Martignoni, E.; Facchinetti, F.; Sances, G.; Petraglia, F.; Nappi, G.; Genazzani, A.R. Acetyl-L-carnitine acutely administered raises beta-endorphin and cortisol plasma levels in humans. Clin. Neuropharmacol. 1988, 11, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Marszałek, M. Alzheimer’s disease against peptides products of enzymatic cleavage of APP protein. Forming and variety of fibrillating peptides—Some aspects (in Polish). Post. Hig. Med. Dosw. 2016, 70, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Nałęcz, K.A.; Miecz, D.; Berezowski, V.; Cecchelli, R. Carnitine: Transport and physiological functions in the brain. Mol. Aspects Med. 2004, 25, 551–567. [Google Scholar] [CrossRef] [PubMed]
- Respondek, M.; Buszman, E. Regulation of neurogenesis: Factors affecting of new neurons formation in adult mammals brain (in Polish). Post. Hig. Med. Dosw. 2015, 69, 1451–1461. [Google Scholar]
- Kazak, F.; Yarim, G.F. Neuroprotective effects of acetyl-l-carnitine on lipopolysaccharide-induced neuroinflammation in mice: Involvement of brain-derived neurotrophic factor. Neurosci Lett. 2017, 658, 32–36. [Google Scholar] [CrossRef]
- Abdul, H.M.; Calabrese, V.; Calvani, M.; Butterfield, D.A. Acetyl-L-carnitine-induced up-regulation of heat shock proteins protects cortical neurons against amyloid-beta peptide 1-42-mediated oxidative stress and neurotoxicity: Implications for Alzheimer’s disease. J. Neurosci. Res. 2006, 82, 398–408. [Google Scholar] [CrossRef]
- Head, K.A. Peripheral neuropathy: Pathogenic mechanisms and alternative therapies. Altern. Med. Rev. 2006, 11, 294–329. [Google Scholar]
- Hudson, S.; Tabet, N. Acetyl-L-carnitine for dementia. Cochrane Database Syst. Rev. 2003, 2, CD003158:1–CD003158:38. [Google Scholar] [CrossRef]
- Scheggi, S.; Rauggi, R.; Nanni, G.; Tagliamonte, A.; Gambarana, C. Repeated acetyl-l-carnitine administration increases phospho-Thr34 DARPP-32 levels and antagonizes cocaine-induced increase in Cdk5 and phospho-Thr75 DARPP-32 levels in rat striatum. Eur. J. Neurosci. 2004, 19, 1609–1620. [Google Scholar] [CrossRef]
- Smeland, O.B.; Meisingset, T.W.; Borges, K.; Sonnewald, U. Chronic acetyl-L-carnitine alters brain energy metabolism and increases noradrenaline and serotonin content in healthy mice. Neurochem. Int. 2012, 61, 100–107. [Google Scholar] [CrossRef]
- Robinson, B.L.; Dumas, M.; Cuevas, E.; Gu, Q.; Paule, M.G.; Ali, S.F.; Kanungo, J. Distinct effects of ketamine and acetyl L-carnitine on the dopamine system in zebrafish. Neurotoxicol. Teratol. 2016, 54, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evangeliou, A.; Vlassopoulos, D. Carnitine metabolism and deficit-when supplementation is necessary? Curr. Pharm. Biotechnol. 2003, 4, 211–219. [Google Scholar] [CrossRef]
- Rigault, C.; Mazué, F.; Bernard, A.; Demarquoy, J.; Le Borgne, F. Changes in l-carnitine content of fish and meat during domestic cooking. Meat Sci. 2008, 78, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Rospond, B.; Chłopicka, J. The biological function of L-carnitine and its content in the particular food examples (in Polish). Prz. Lek. 2013, 70, 85–91. [Google Scholar]
- Kelly, G.S. L-Carnitine: Therapeutic applications of a conditionally essential amino acid. Altern. Med. Rev. 1998, 3, 345–360. [Google Scholar]
- Pękala, J.; Patkowska-Sokoła, B.; Bodkowski, R.; Jamroz, D.; Nowakowski, P.; Lochyński, S.; Librowski, T. L-carnitine-metabolic functions and meaning in humans life. Curr. Drug Metab. 2011, 12, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Yang, M.; Zhou, M.; Xiao, J.; Guo, J.; He, L. L-carnitine for cognitive enhancement in people without cognitive impairment. Cochrane Database Syst. Rev. 2017, 3, CD009374:1–CD009374:28. [Google Scholar] [CrossRef] [PubMed]
- Wollen, K.A. Alzheimer’s disease: The pros and cons of pharmaceutical, nutritional, botanical, and stimulatory therapies, with a discussion of treatment strategies from the perspective of patients and practitioners. Altern. Med. Rev. 2010, 15, 223–244. [Google Scholar] [PubMed]
- Gavrilova, S.I.; Kalyn, I.B.; Kolykhalov, I.V.; Roshchina, I.F.; Selezneva, N.D. Acetyl-L-carnitine (carnicetine) in the treatment of early stages of Alzheimer’s disease and vascular dementia. Zh. Nevrol. Psikhiatr. Im. S. S. Korsakova 2011, 111, 16–22. [Google Scholar]
- Parnetti, L.; Gaiti, A.; Mecocci, P.; Cadini, D.; Senin, U. Pharmacokinetics of i.v. and oral acetyl-L-carnitine in a multiple dose regimen in patients with senile dementia of Alzheimer type. Eur. J. Clin. Pharmacol. 1992, 42, 89–93. [Google Scholar] [CrossRef]
- Wang, S.M.; Han, C.; Lee, S.J.; Patkar, A.A.; Masand, P.S.; Pae, C.U. A review of current evidence for acetyl-l-carnitine in the treatment of depression. J. Psychiatr. Res. 2014, 53, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Gaby, A.R. Natural approaches to epilepsy. Altern. Med. Rev. 2007, 12, 9–24. [Google Scholar] [PubMed]
- Bain, M.A.; Milne, R.W.; Evans, A.M. Disposition and metabolite kinetics of oral L-carnitine in humans. J. Clin. Pharmacol. 2006, 46, 1163–1170. [Google Scholar] [CrossRef]
- Ferrari, R.; Merli, E.; Cicchitelli, G.; Mele, D.; Fucili, A.; Ceconi, C. Therapeutic effects of L-carnitine and propionyl-L-carnitine on cardiovascular diseases: A review. Ann. N. Y. Acad. Sci. 2004, 1033, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Deng, R.; Li, J.; Chi, W.; Su, Z.; Lin, J.; Pflugfelder, S.C.; Li, D.Q. Protective effects of L-carnitine against oxidative injury by hyperosmolarity in human corneal epithelial cells. Invest. Ophthalmol. Vis. Sci. 2015, 56, 5503–5511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seen, S.; Tong, L. Dry eye disease and oxidative stress. Acta Ophthalmol. 2017, 96, e412–e420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tashiro, K.; Kaida, Y.; Yamagishi, S.I.; Tanaka, H.; Yokoro, M.; Yano, J.; Sakai, K.; Kurokawa, Y.; Taguchi, K.; Nakayama, Y.; et al. L-Carnitine supplementation improves self-rating depression scale scores in uremic male patients undergoing hemodialysis. Lett. Drug Des. Discov. 2017, 14, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Cruciani, R.A.; Revuelta, M.; Dvorkin, E.; Homel, P.; Lesage, P.; Esteban-Cruciani, N. L-carnitine supplementation in patients with HIV/AIDS and fatigue: A double-blind, placebo-controlled pilot study. HIV AIDS (Auckl) 2015, 7, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benvenga, S. Effects of L-carnitine on thyroid hormone metabolism and on physical exercise tolerance. Horm. Metab. Res. 2005, 37, 566–571. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Fan, Z.K.; Cao, Y.; Jia, Z.Q.; Li, G.; Zhi, X.D.; Yu, D.S.; Lv, G. Acetyl-L-carnitine ameliorates mitochondrial damage and apoptosis following spinal cord injury in rats. Neurosci. Lett. 2015, 604, 18–23. [Google Scholar] [CrossRef]
- Bigford, G.E.; Del Rossi, G. Supplemental substances derived from foods as adjunctive therapeutic agents for treatment of neurodegenerative diseases and disorders. Adv. Nutr. 2014, 5, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Chiechio, S.; Canonico, P.L.; Grilli, M. L-Acetylcarnitine: A mechanistically distinctive and potentially rapid-acting antidepressant drug. Int. J. Mol. Sci. 2017, 19, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, A.M.; Mancinelli, A.; Longo, A. Excretion and metabolism of propionyl-L-carnitine in the isolated perfused rat kidney. J. Pharmacol. Exp. Ther. 1997, 281, 1071–1076. [Google Scholar] [PubMed]
- Ścibior, D.; Czeczot, H. Arginine-metabolism and functions in the human organism. Post. Hig. Med. Dosw. 2004, 58, 321–332. [Google Scholar]
- Bloomer, R.J.; Fisher-Wellman, K.H.; Tucker, P.S. Effect of oral acetyl L-carnitine arginate on resting and postprandial blood biomarkers in pre-diabetics. Nutr. Metab. (Lond.) 2009, 6, 25:1–25:11. [Google Scholar] [CrossRef] [Green Version]
- Aliev, G.; Liu, J.; Shenk, J.C.; Fischbach, K.; Pacheco, G.J.; Chen, S.G.; Obrenovich, M.E.; Ward, W.F.; Richardson, A.G.; Smith, M.A.; et al. Neuronal mitochondrial amelioration by feeding acetyl-L-carnitine and lipoic acid to aged rats. J. Cell. Mol. Med. 2009, 13, 320–333. [Google Scholar] [CrossRef]
- Bescós, R.; Sureda, A.; Tur, J.A.; Pons, A. The effect of nitric-oxide-related supplements on human performance. Sports Med. 2012, 42, 99–117. [Google Scholar] [CrossRef]
- Ganai, A.A.; Jahan, S.; Ahad, A.; Abdin, M.Z.; Farooqi, H. Glycine propionyl L-carnitine attenuates D-galactosamine induced fulminant hepatic failure in Wistar rats. Chem. Biol. Interact. 2014, 214, 33–40. [Google Scholar] [CrossRef]
- Jacobs, P.L.; Goldstein, E.R. Long-term glycine propionyl-l-carnitine supplemention and paradoxical effects on repeated anaerobic sprint performance. J. Int Soc. Sports Nutr. 2010, 7, 35:1–35:8. [Google Scholar] [CrossRef] [Green Version]
- Bloomer, R.J.; Smith, W.A.; Fisher-Wellman, K.H. Oxidative stress in response to forearm ischemia-reperfusion with and without carnitine administration. Int. J. Vitam. Nutr. Res. 2010, 80, 12–23. [Google Scholar] [CrossRef]
- Spiering, B.A.; Kraemer, W.J.; Hatfield, D.L.; Vingren, J.L.; Fragala, M.S.; Ho, J.Y.; Thomas, G.A.; Häkkinen, K.; Volek, J.S. Effects of L-carnitine L-tartrate supplementation on muscle oxygenation responses to resistance exercise. J. Strength. Cond. Res. 2008, 22, 1130–1135. [Google Scholar] [CrossRef] [PubMed]
- Spiering, B.A.; Kraemer, W.J.; Vingren, J.L.; Hatfield, D.L.; Fragala, M.S.; Ho, J.Y.; Maresh, C.M.; Anderson, J.M.; Volek, J.S. Responses of criterion variables to different supplemental doses of L-carnitine L-tartrate. J. Strength. Cond. Res. 2007, 21, 259–264. [Google Scholar] [CrossRef]
- Kraemer, W.J.; Spiering, B.A.; Volek, J.S.; Ratamess, N.A.; Sharman, M.J.; Rubin, M.R.; French, D.N.; Silvestre, R.; Hatfield, D.L.; Van Heest, J.L.; et al. Androgenic responses to resistance exercise: Effects of feeding and L-carnitine. Med. Sci. Sports Exerc. 2006, 38, 1288–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramowicz, W.N.; Galloway, S.D. Effects of acute versus chronic L-carnitine L-tartrate supplementation on metabolic responses to steady state exercise in males and females. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 386–400. [Google Scholar] [CrossRef]
- Volek, J.S.; Kraemer, W.J.; Rubin, M.R.; Gómez, A.L.; Ratamess, N.A.; Gaynor, P. L-Carnitine L-tartrate supplementation favorably affects markers of recovery from exercise stress. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E474–E482. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| No. | Type of Food | Components | Intake Categories | Characteristics |
|---|---|---|---|---|
| 1. | green, leafy vegetables | kale, spinach, kohlrabi, different varieties of lettuce, cooked greens and salads | ≥6 servings/week | vegetables rich in vitamins C and A |
| 2. | all other vegetables | celery, cabbage, beets, cucumbers, cauliflower, zucchini, tomatoes, leeks, garlic and onion | 1 serving/day | to choose non-starchy vegetables with a lot of nutrients and a low number of calories |
| 3. | berries | strawberries, blueberries, raspberries, blackberries | ≥2 servings/week | a source of antioxidants |
| 4. | nuts and almonds | pineapple, pistachios, macadamia, pecans, peanuts and Brazilian walnut | ≥5 servings/week | are a source of unsaturated fatty acids and antioxidants; contain vitamins E, B1 and PP; and reduce the level of “bad” cholesterol |
| 5. | whole grains | oatmeal, quinoa, brown rice, whole-wheat pasta and 100% whole-wheat bread | ≥3 servings/day | a source of fiber, folic acid, vitamin B3, iron, zinc, magnesium and phosphorus |
| 6. | fish | salmon, sardines, trout, tuna and mackerel | ≥1 serving/week | high amounts of omega-3 fatty acids |
| 7. | poultry | chicken or turkey | ≥2 servings/week | fried chicken is not encouraged on the MIND diet |
| 8. | beans | lentils, soybeans, string beans, broad beans, green peas, chickpeas and white beans | ≥4 servings/week | a source of fiber, protein, vitamins and minerals |
| 9. | olive oil | cold pressed oils | a source of vitamins A, D, E, and K; polyunsaturated fatty acids; use fat for long frying at a high smoke point | |
| 10. | milk, dairy products | low-fat: milk, cheese, yoghurt, buttermilk, kefir and cottage cheese | ≥2 glasses/day; 280–400 g semi-skimmed cheese or 1 slice (30 g) of yellow cheese. | a source of protein and minerals: calcium, potassium, phosphorus, magnesium, zinc, manganese, iron; high in vitamins: B2, B12, A, D, E, and K and probiotics |
| 11. | wine | red and white | ≤1 glass serving/day | both red and white wine may benefit the brain; red wine is recommended because a lot of research has focused on the red wine compound resveratrol |
| beer | non-alcoholic beer | regular beer consumption is not recommended for some risk-group populations (pregnant, children, people affected by liver diseases) | source: carbohydrates, protein/amino acids (proline, glutamic and aspartic acid, glycine, alanine), minerals (fluoride, potassium, phosphorus, calcium, sodium, magnesium, silicon), vitamins (B1–B6, folic acid), and other compounds, such as polyphenols |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kepka, A.; Ochocinska, A.; Borzym-Kluczyk, M.; Skorupa, E.; Stasiewicz-Jarocka, B.; Chojnowska, S.; Waszkiewicz, N. Preventive Role of L-Carnitine and Balanced Diet in Alzheimer’s Disease. Nutrients 2020, 12, 1987. https://doi.org/10.3390/nu12071987
Kepka A, Ochocinska A, Borzym-Kluczyk M, Skorupa E, Stasiewicz-Jarocka B, Chojnowska S, Waszkiewicz N. Preventive Role of L-Carnitine and Balanced Diet in Alzheimer’s Disease. Nutrients. 2020; 12(7):1987. https://doi.org/10.3390/nu12071987
Chicago/Turabian StyleKepka, Alina, Agnieszka Ochocinska, Małgorzata Borzym-Kluczyk, Ewa Skorupa, Beata Stasiewicz-Jarocka, Sylwia Chojnowska, and Napoleon Waszkiewicz. 2020. "Preventive Role of L-Carnitine and Balanced Diet in Alzheimer’s Disease" Nutrients 12, no. 7: 1987. https://doi.org/10.3390/nu12071987