
Divergence Observation in a Mesoscale Eddy during Chla Bloom Revealed in Submesoscale Satellite Currents

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chla Observations near Mesoscale Eddies
2.2. Satellite-Based Surface Currents
2.3. Model Observations
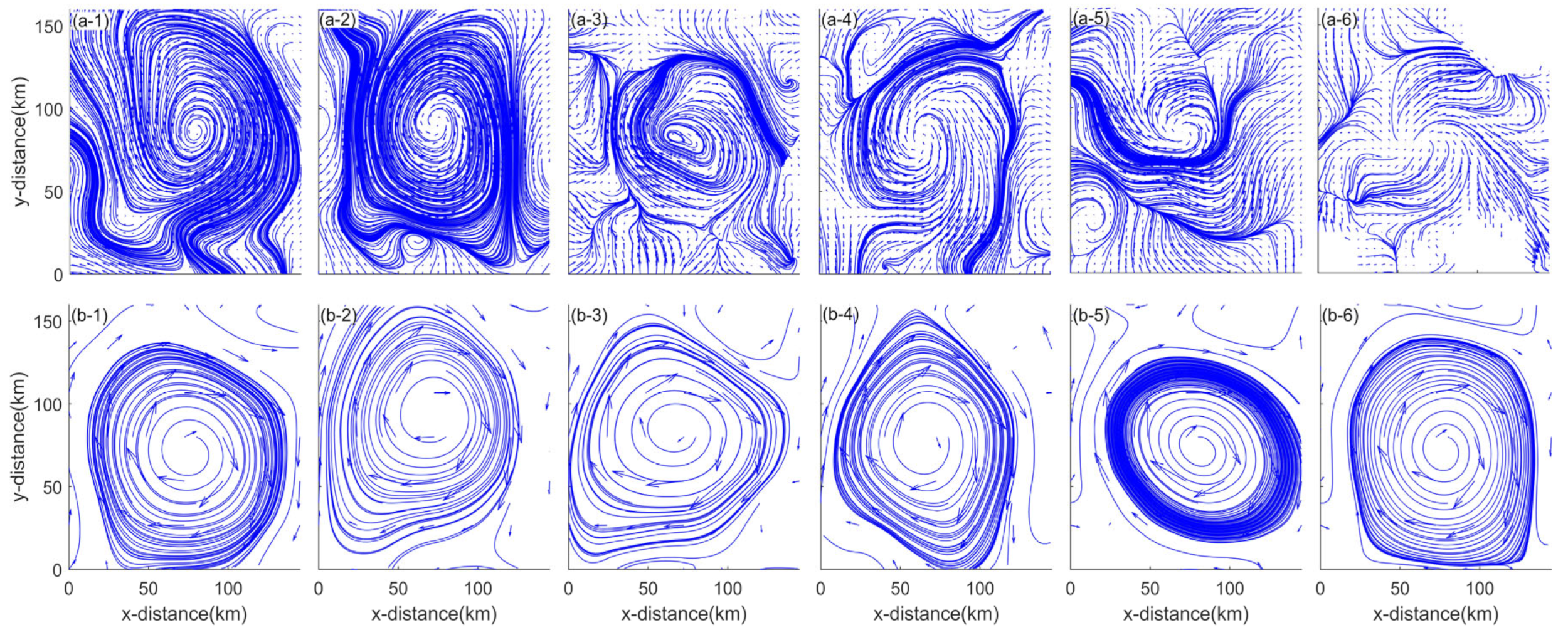
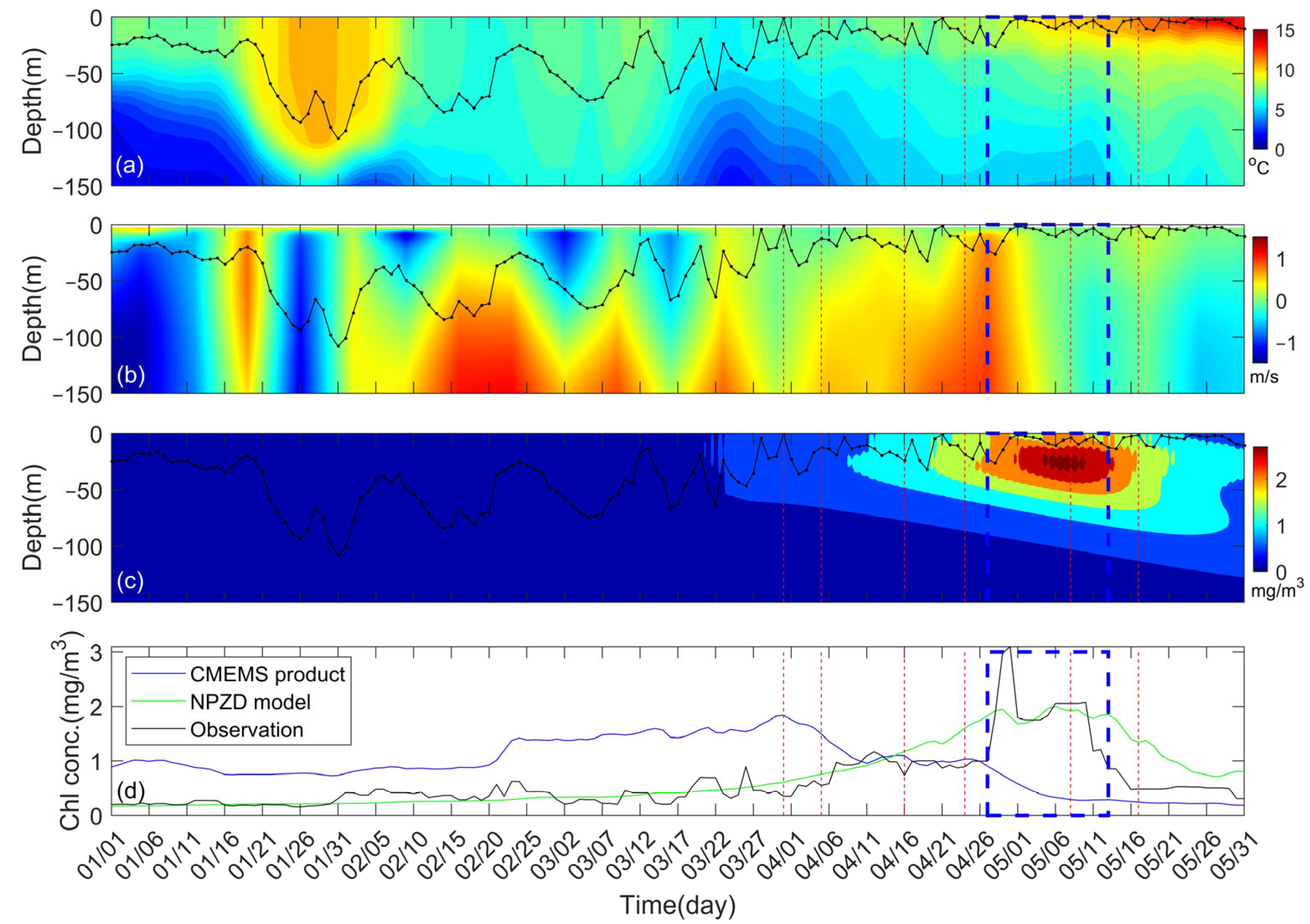
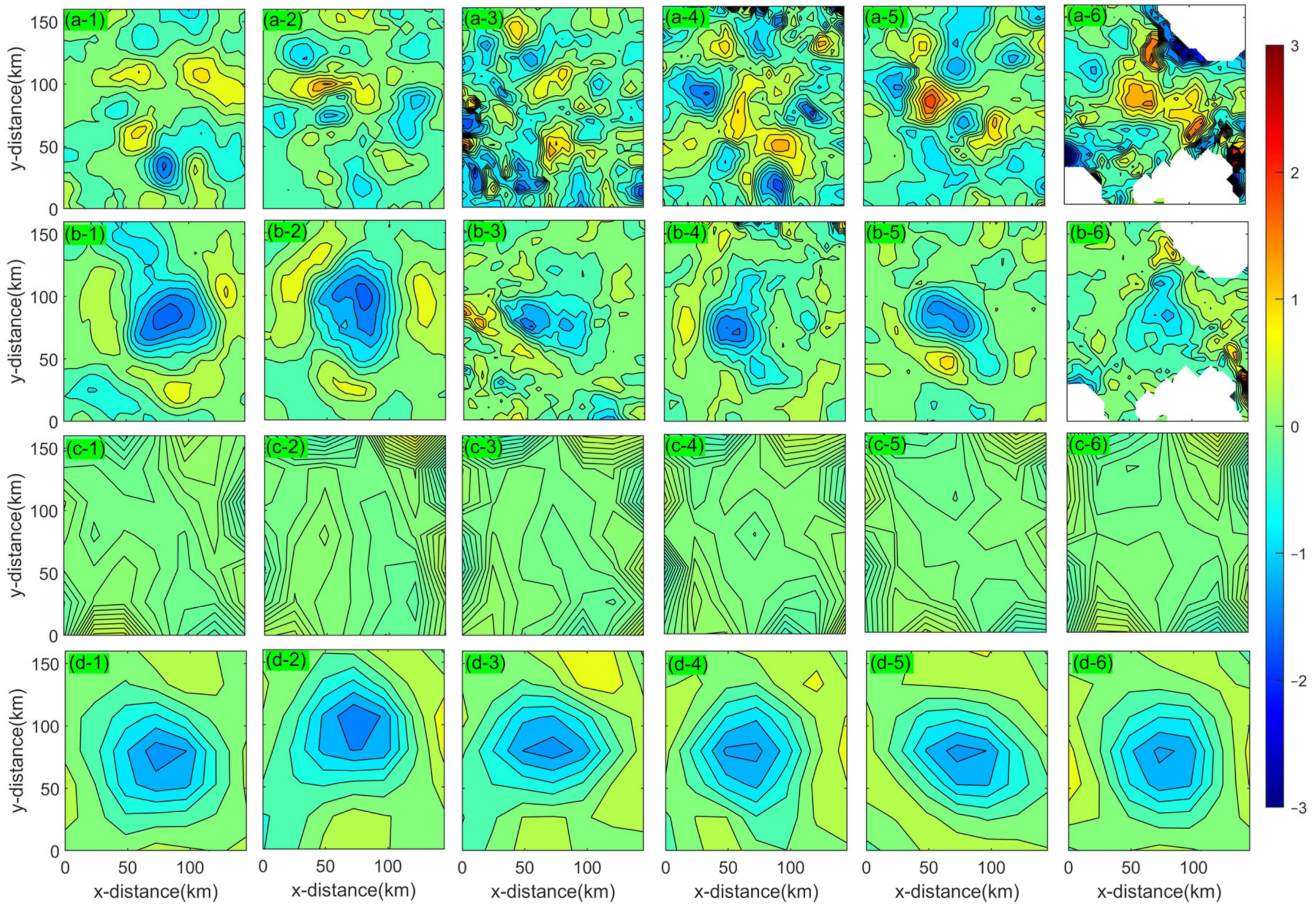
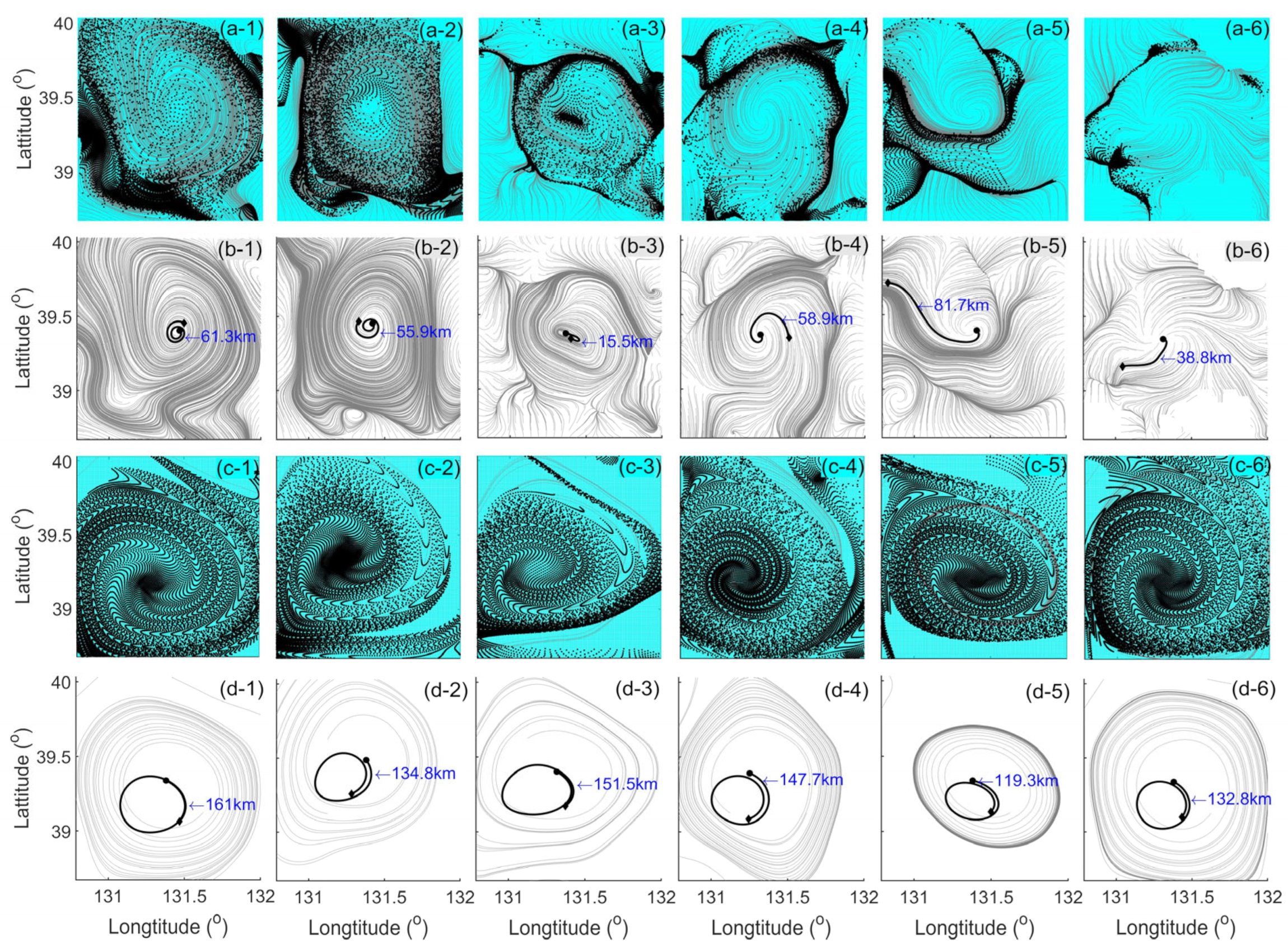
3. Results
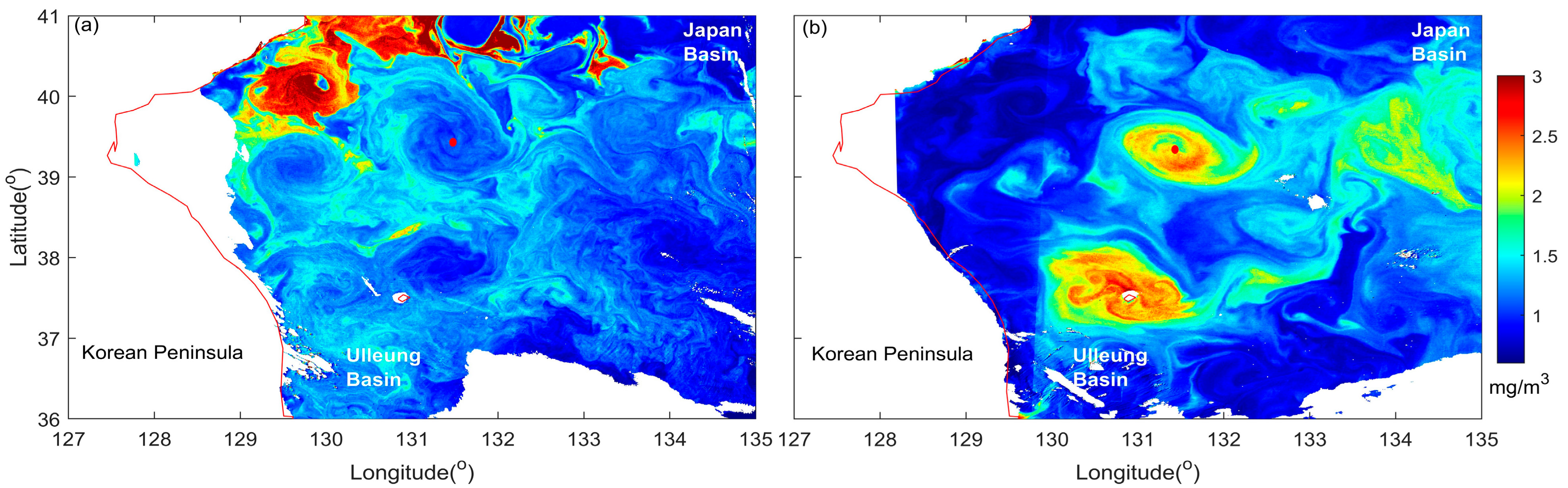
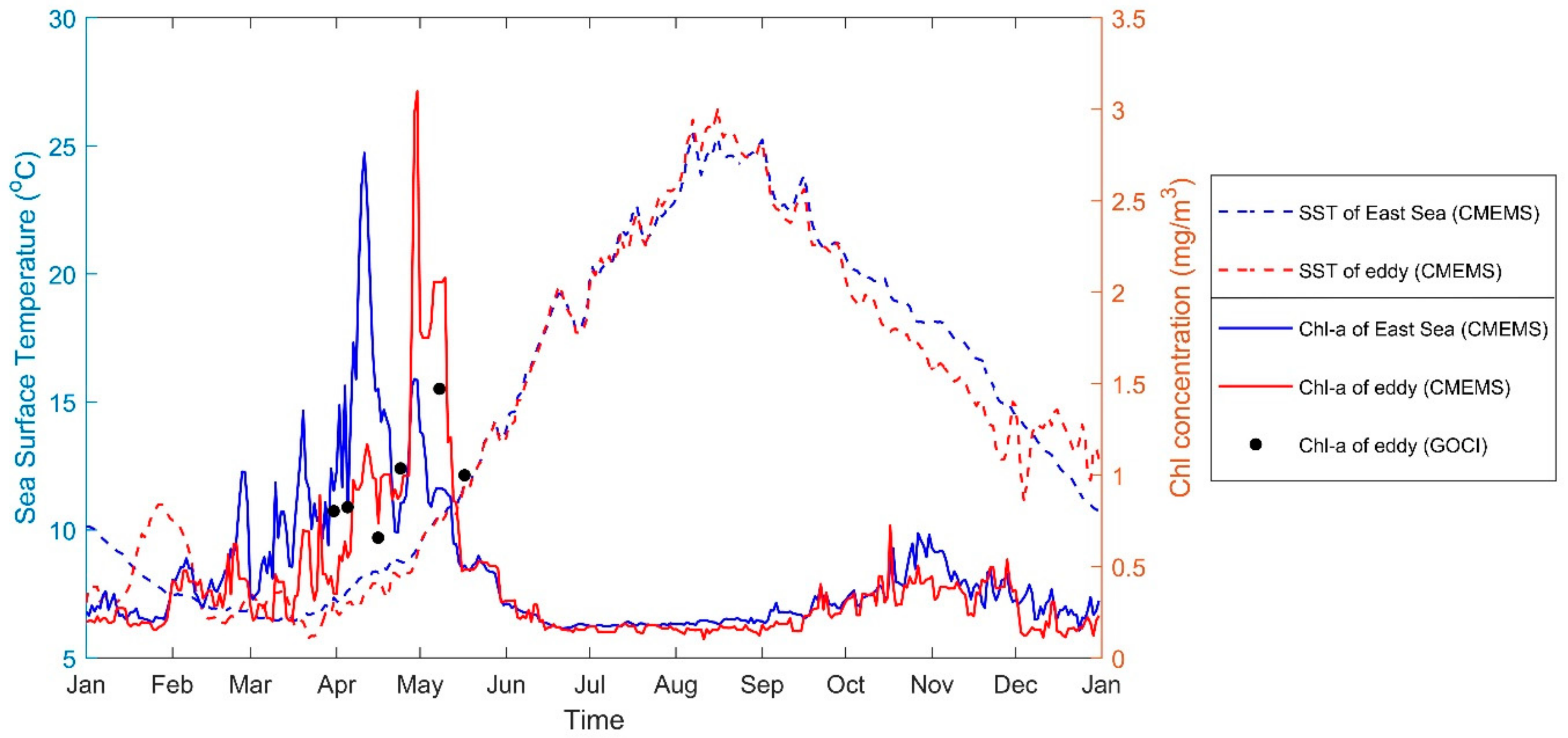
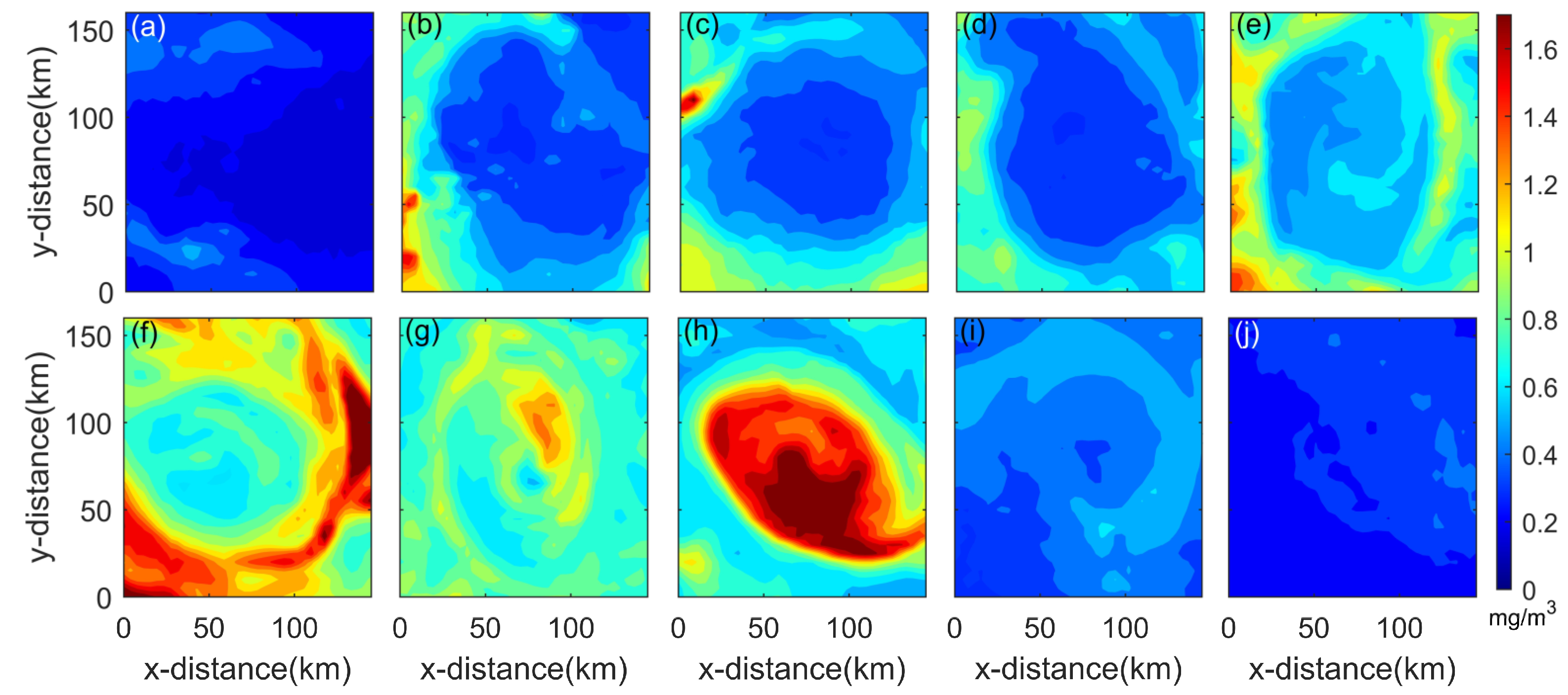
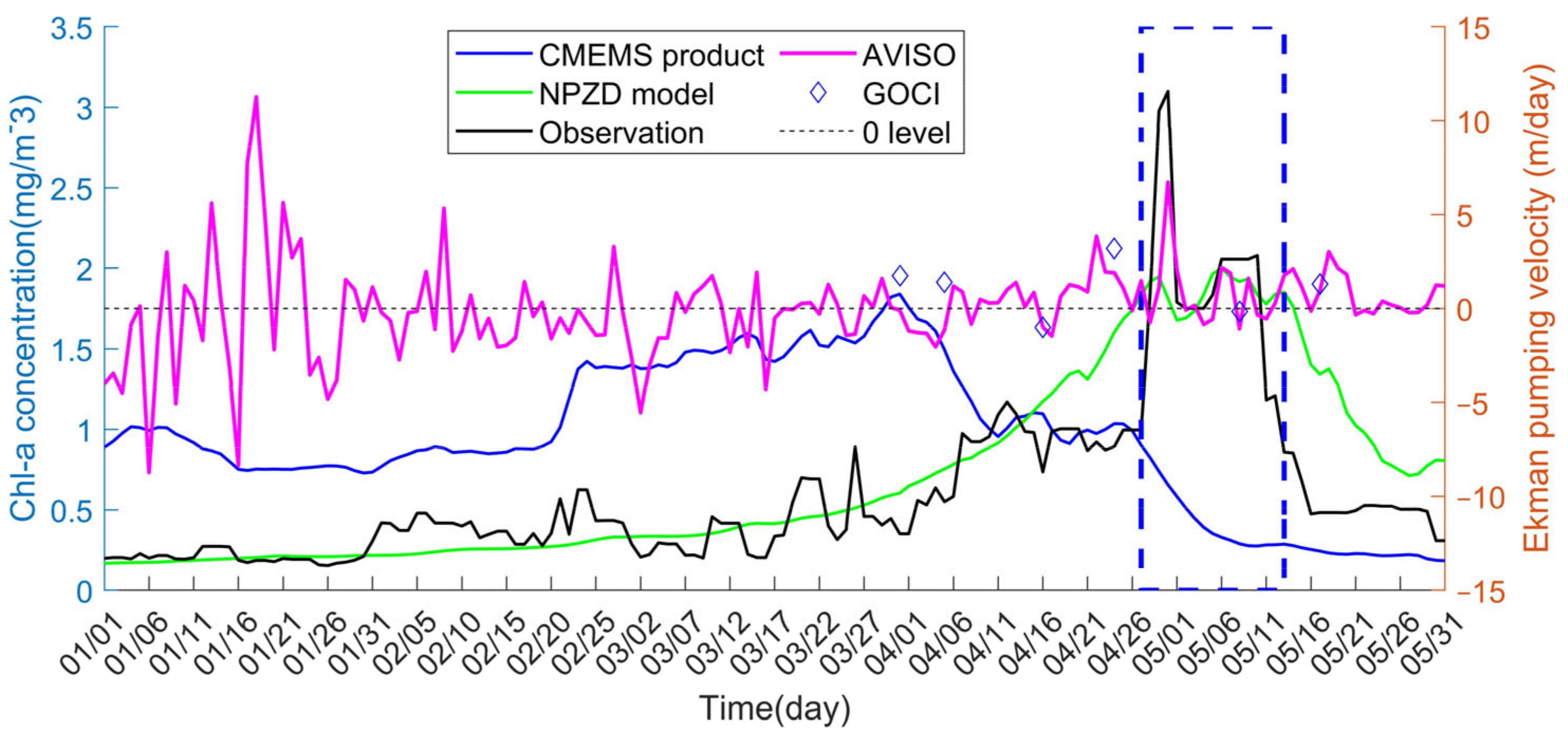
3.1. Secondary Bloom Observations
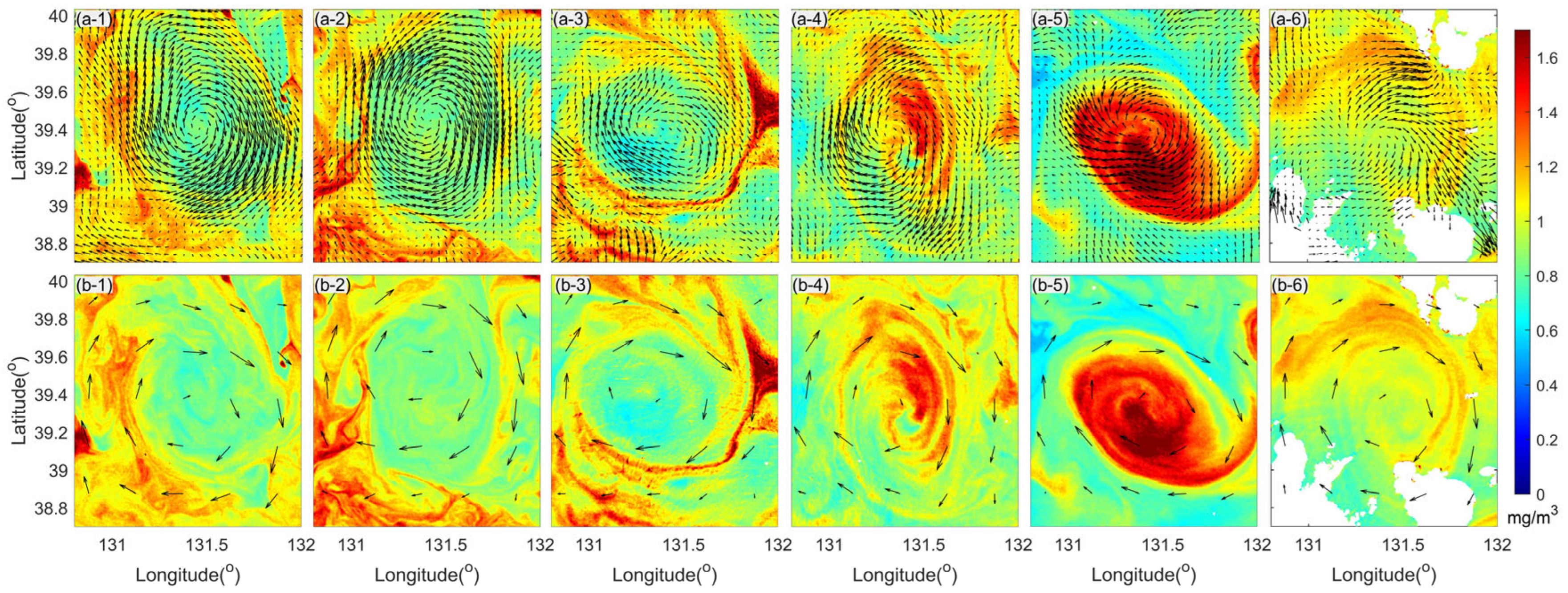
3.2. Current Observations
3.3. Model Observations
4. Discussion
4.1. Divergence and Vorticity in a Mesoscale Eddy
4.2. Horizontal Material Transport in a Mesoscale Eddy
4.3. Possible Bloom Mechanisms
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ferrari, R.; Wunsch, C. Ocean circulation kinetic energy: Reservoirs, sources, and sinks. Annu. Rev. Fluid Mech. 2009, 41, 253–282. [Google Scholar] [CrossRef]
- Kim, D.; Yang, E.J.; Kim, K.H.; Shin, C.W.; Park, J.; Yoo, S.; Hyun, J.H. Impact of an anticyclonic eddy on the summer nutrient and chlorophyll a distributions in the Ulleung Basin, East Sea (Japan Sea). ICES J. Mar. Sci. 2012, 69, 23–29. [Google Scholar] [CrossRef]
- Arostegui, M.C.; Gaube, P.; Woodworth Jefcoats, P.A.; Kobayashi, D.R.; Braun, C.D. Anticyclonic eddies eggreagate pelagic predators in a subtropical gyre. Nature 2022, 609, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Fei, J.; Liu, L.; Huang, X.; Li, Y. An investigation of the influences of mesoscale ocean eddies on tropical cyclone intensities. Mon. Weather Rev. 2017, 145, 1181–1201. [Google Scholar] [CrossRef]
- Small, R.J.; Msadek, R.; Kwon, Y.O.; Booth, J.F.; Zarzycki, C. Atmosphere surface storm track response to resolved ocean mesoscale in two sets of global climate model experiments. Clim. Dyn. 2019, 52, 2067–2089. [Google Scholar] [CrossRef]
- Korotenko, K.A. Effects of mesoscale eddies on behavior of an oil spill resulting from an accidental deepwater blowout in the Black Sea: An assessment of the environmental impacts. PeerJ 2018, 6, e5448. [Google Scholar] [CrossRef]
- McGillicuddy, D.J., Jr. Mechanisms of physical-biological-biogeochemical interaction at the oceanic mesoscale. Annu. Rev. Mar. Sci. 2016, 8, 125–159. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, D. Eddy-feature phytoplankton bloom induced by a tropical cyclone in the South China Sea. Int. J. Remote Sens. 2012, 33, 7444–7457. [Google Scholar] [CrossRef]
- Ciancia, E.; Lacava, T.; Pergola, N.; Vellucci, V.; Antoine, D.; Satriano, V.; Tramutoli, V. Quantifying the Variability of Phytoplankton Blooms in the NW Mediterranean Sea with the Robust Satellite Techniques (RST). Remote Sens. 2021, 13, 5151. [Google Scholar] [CrossRef]
- Korotenko, K.; Osadchiev, A.; Melnikov, V. Mesoscale Eddies in the Black Sea and Their Impact on River Plumes: Numerical Modeling and Satellite Observations. Remote Sens. 2022, 14, 4149. [Google Scholar] [CrossRef]
- McGillicuddy, D.J., Jr.; Kosnyrev, V.K.; Ryan, J.P.; Yoder, J.A. Covariation of mesoscale ocean color and sea-surface temperature patterns in the Sargasso Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 48, 1823–1836. [Google Scholar] [CrossRef]
- Dong, C.; Liu, L.; Nencioli, F.; Bethel, B.J.; Liu, Y.; Xu, G.; Ma, J.; Ji, J.; Sun, W.; Shan, H.; et al. The near-global ocean mesoscale eddy atmospheric-oceanic-biological interaction observational dataset. Sci. Data 2022, 9, 436. [Google Scholar] [CrossRef]
- Zhu, Y.; Liang, X. Characteristics of Eulerian Mesoscale Eddies in the Gulf of Mexico. Front. Mar. Sci. 2022, 9, 2655. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, X.; Liu, T.; Chen, D. Characteristics of Mesoscale Eddies in the Vicinity of the Kuroshio: Statistics from Satellite Altimeter Observations and OFES Reanalysis Data. J. Mar. Sci. Eng. 2022, 10, 1975. [Google Scholar] [CrossRef]
- Mkhinini, N.; Coimbra, A.L.S.; Stegner, A.; Arsouze, T.; Taupier-Letage, I.; Béranger, K. Long-lived mesoscale eddies in the eastern Mediterranean Sea: Analysis of 20 years of AVISO geostrophic velocities. J. Geophys. Res. Ocean. 2014, 119, 8603–8626. [Google Scholar] [CrossRef]
- Ioannou, A.; Stegner, A.; Tuel, A.; LeVu, B.; Dumas, F.; Speich, S. Cyclostrophic corrections of AVISO/DUACS surface velocities and its application to mesoscale eddies in the Mediterranean Sea. J. Geophys. Res. Ocean. 2019, 124, 8913–8932. [Google Scholar] [CrossRef]
- Kurczyn, J.A.; Beier, E.; Lavín, M.F.; Chaigneau, A. Mesoscale eddies in the northeastern Pacific tropical-subtropical transition zone: Statistical characterization from satellite altimetry. J. Geophys. Res. Ocean. 2012, 117, C10021. [Google Scholar] [CrossRef]
- Kang, J.J.; Jang, H.K.; Lim, J.H.; Lee, D.; Lee, J.H.; Bae, H.; Lee, C.H.; Kang, C.-K.; Lee, S.H. Characteristics of Different Size Phytoplankton for Primary Production and Biochemical Compositions in the Western East/Japan Sea. Front. Microbiol. 2020, 11, 560102. [Google Scholar] [CrossRef]
- Choi, J.; Park, Y.G.; Kim, W.; Kim, Y.H. Characterization of submesoscale turbulence in the east/japan sea using geostationary ocean color satellite images. Geophys. Res. Lett. 2019, 46, 8214–8223. [Google Scholar] [CrossRef]
- George, J.V.; Nuncio, M.; Chacko, R.; Anilkumar, N.; Noronha, S.B.; Patil, S.M.; Pavithran, S.; Alappattu, D.P.; Krishnan, K.P.; Achuthankutty, C.T. Role of physical processes in chlorophyll distribution in the western tropical Indian Ocean. J. Mar. Syst. 2013, 113, 1–12. [Google Scholar] [CrossRef]
- McWilliams, J.C. Submesoscale currents in the ocean. Proc. R. Soc. A Math. Phys. Eng. Sci. 2016, 472, 20160117. [Google Scholar] [CrossRef] [PubMed]
- Berta, M.; Griffa, A.; Haza, A.C.; Horstmann, J.; Huntley, H.S.; Ibrahim, R.; Lund, B.; Özgökmen, T.M.; Poje, A.C. Submesoscale kinematic properties in summer and winter surface flows in the Northern Gulf of Mexico. J. Geophys. Res. Ocean. 2020, 125, e2020JC016085. [Google Scholar] [CrossRef]
- Liu, G.; Bracco, A.; Sitar, A. Submesoscale Mixing Across the Mixed Layer in the Gulf of Mexico. Front. Mar. Sci. 2021, 8, 615066. [Google Scholar] [CrossRef]
- Kim, W.; Moon, J.E.; Park, Y.J.; Ishizaka, J. Evaluation of chlorophyll retrievals from Geostationary Ocean color imager (GOCI) for the north-east Asian region. Remote Sens. Environ. 2016, 184, 482–495. [Google Scholar] [CrossRef]
- Park, M.S.; Lee, S.; Ahn, J.H.; Lee, S.J.; Choi, J.K.; Ryu, J.H. Decadal Measurements of the First Geostationary Ocean Color Satellite (GOCI) Compared with MODIS and VIIRS Data. Remote Sens. 2021, 14, 72. [Google Scholar] [CrossRef]
- Park, K.A.; Park, J.E.; Kang, C.K. Satellite-Observed Chlorophyll-a Concentration Variability in the East Sea (Japan Sea): Seasonal Cycle, Long-Term Trend, and Response to Climate Index. Front. Mar. Sci. 2022, 9, 807570. [Google Scholar] [CrossRef]
- Yamada, K.; Ishizaka, J.; Yoo, S.; Kim, H.C.; Chiba, S. Seasonal and interannual variability of sea surface chlorophyll a concentration in the Japan/East Sea (JES). Prog. Oceanogr. 2004, 61, 193–211. [Google Scholar] [CrossRef]
- Chen, S.; Meng, Y.; Lin, S.; Xi, J. Remote Sensing of the Seasonal and Interannual Variability of Surface Chlorophyll-a Concentration in the Northwest Pacific over the Past 23 Years (1997–2020). Remote Sens. 2022, 14, 5611. [Google Scholar] [CrossRef]
- Ashijan, C.; Arnone, R.A.; Davis, C.; Jones, B.; Kahru, M.; Lee, C.; Mitchell, B.G. Biological Structure and Seasonality in the Japan/East Sea. Oceanography 2006, 19, 122–133. [Google Scholar] [CrossRef]
- Faghmous, J.H.; Frenger, I.; Yao, Y.; Warmka, R.; Lindell, A.; Kumar, V. A daily global mesoscale ocean eddy dataset from satellite altimetry. Sci. Data 2015, 2, 150028. [Google Scholar] [CrossRef] [Green Version]
- Emery, W.J.; Thomas, A.C.; Collins, M.J.; Crawford, W.R.; Mackas, D.L. An objective procedure to compute advection from sequential infrared satellite images. J. Geophys. Res. 1986, 91, 12–865. [Google Scholar]
- Zavialov, P.O.; Grigorieva, J.V.; Möller, O.O., Jr.; Kostianoy, A.G.; Gregoire, M. Continuity preserving modified maximum cross-correlation technique. J. Geophys. Res. Ocean. 2002, 107, 24-1. [Google Scholar] [CrossRef]
- Yang, H.; Choi, J.K.; Park, Y.J.; Han, H.J.; Ryu, J.H. Application of the Geostationary Ocean Color Imager (GOCI) to estimates of ocean surface currents. J. Geophys. Res. Ocean. 2014, 119, 3988–4000. [Google Scholar] [CrossRef]
- Yang, H.; Arnone, R.; Jolliff, J. Estimating advective near-surface currents from ocean color satellite images. Remote Sens. Environ. 2015, 158, 1–14. [Google Scholar] [CrossRef]
- Park, J.E.; Park, K.A.; Ullman, D.S.; Cornillon, P.C.; Park, Y.J. Observation of diurnal variations in mesoscale eddy sea-surface currents using GOCI data. Remote Sens. Lett. 2016, 7, 1131–1140. [Google Scholar] [CrossRef]
- Sun, H.; Song, Q.; Shao, R.; Schlicke, T. Estimation of sea surface currents based on ocean colour remote-sensing image analysis. Int. J. Remote Sens. 2016, 37, 5105–5121. [Google Scholar] [CrossRef]
- Warren, M.A.; Quartly, G.D.; Shutler, J.D.; Miller, P.I.; Yoshikawa, Y. Estimation of ocean surface currents from maximum cross correlation applied to GOCI geostationary satellite remote sensing data over the Tsushima (Korea) Straits. J. Geophys. Res. Ocean. 2016, 121, 6993–7009. [Google Scholar] [CrossRef]
- Liu, J.; Emery, W.J.; Wu, X.; Li, M.; Li, C.; Zhang, L. Computing ocean surface currents from GOCI ocean color satellite imagery. IEEE Trans. Geosci. Remote Sens. 2017, 55, 7113–7125. [Google Scholar] [CrossRef]
- Choi, J.M.; Kim, W.; Hong, T.T.M.; Park, Y.G. Derivation and Evaluation of Satellite-Based Surface Current. Front. Mar. Sci. 2021, 8, 695780. [Google Scholar] [CrossRef]
- Thielicke, W.; Stamhuis, E. PIVlab–towards user-friendly, affordable and accurate digital particle image velocimetry in MATLAB. J. Open Res. Softw. 2014, 2. [Google Scholar] [CrossRef]
- Mason, E.; Pascual, A.; McWilliams, J.C. A new sea surface height–based code for oceanic mesoscale eddy tracking. J. Atmos. Ocean. Technol. 2014, 31, 1181–1188. [Google Scholar] [CrossRef] [Green Version]
- Delepoulle, A.; Chelton, D.; Schlax, M.; Faugere, Y.; Dibarboure, G. 24 year mesoscale eddy trajectory atlas on AVISO. In Geophysical Research Abstracts; EGU2018-13690; European Geosciences Union: Vienna, Austria, 2018; Volume 20. [Google Scholar]
- Kuhn, A.M.; Fennel, K.; Mattern, J.P. Model investigations of the North Atlantic spring bloom initiation. Prog. Oceanogr. 2015, 138, 176–193. [Google Scholar] [CrossRef]
- Li, J.; Qi, Y.; Jing, Z.; Wang, J. Enhancement of eddy-Ekman pumping inside anticyclonic eddies with wind-parallel extension: Satellite observations and numerical studies in the South China Sea. J. Mar. Syst. 2014, 132, 150–161. [Google Scholar] [CrossRef]
- Jalón-Rojas, I.; Wang, X.H.; Fredj, E. A 3D numerical model to track marine plastic debris (TrackMPD): Sensitivity of microplastic trajectories and fates to particle dynamical properties and physical processes. Mar. Pollut. Bull. 2019, 141, 256–272. [Google Scholar] [CrossRef]
- Martin, A.P. Phytoplankton patchiness: The role of lateral stirring and mixing. Prog. Oceanogr. 2003, 57, 125–174. [Google Scholar] [CrossRef]
- D’Ovidio, F.; De Monte, S.; Della Penna, A.; Cotté, C.; Guinet, C. Ecological implications of eddy retention in the open ocean: A Lagrangian approach. J. Phys. A Math. Theor. 2013, 46, 254023. [Google Scholar] [CrossRef]
- Gaube, P.; McGillicuddy, D.J., Jr.; Chelton, D.B.; Behrenfeld, M.J.; Strutton, P.G. Regional variations in the influence of mesoscale eddies on near-surface chlorophyll. J. Geophys. Res. Ocean. 2014, 119, 8195–8220. [Google Scholar] [CrossRef] [Green Version]
- Stern, M.E. Interaction of a uniform wind stress with a geostrophic vortex. In Deep Sea Research and Oceanographic Abstracts; Elsevier: Amsterdam, The Netherlands, 1965; Volume 12, pp. 355–367. [Google Scholar]
- Tennekes, H.; Lumley, J.L. A First Course in Turbulence; The Massachusetts Institute of Technology: NewYork, NY, USA, 1972; Chapter 2; Volume 9. [Google Scholar]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, T.T.M.; Park, Y.-G.; Choi, J.M. Divergence Observation in a Mesoscale Eddy during Chla Bloom Revealed in Submesoscale Satellite Currents. Remote Sens. 2023, 15, 995. https://doi.org/10.3390/rs15040995
Hong TTM, Park Y-G, Choi JM. Divergence Observation in a Mesoscale Eddy during Chla Bloom Revealed in Submesoscale Satellite Currents. Remote Sensing. 2023; 15(4):995. https://doi.org/10.3390/rs15040995
Chicago/Turabian StyleHong, Tran Thi My, Young-Gyu Park, and Jun Myoung Choi. 2023. "Divergence Observation in a Mesoscale Eddy during Chla Bloom Revealed in Submesoscale Satellite Currents" Remote Sensing 15, no. 4: 995. https://doi.org/10.3390/rs15040995