
Parameterization of Light Absorption of Phytoplankton, Non-Algal Particles and Coloured Dissolved Organic Matter in the Atlantic Region of the Southern Ocean (Austral Summer of 2020)

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
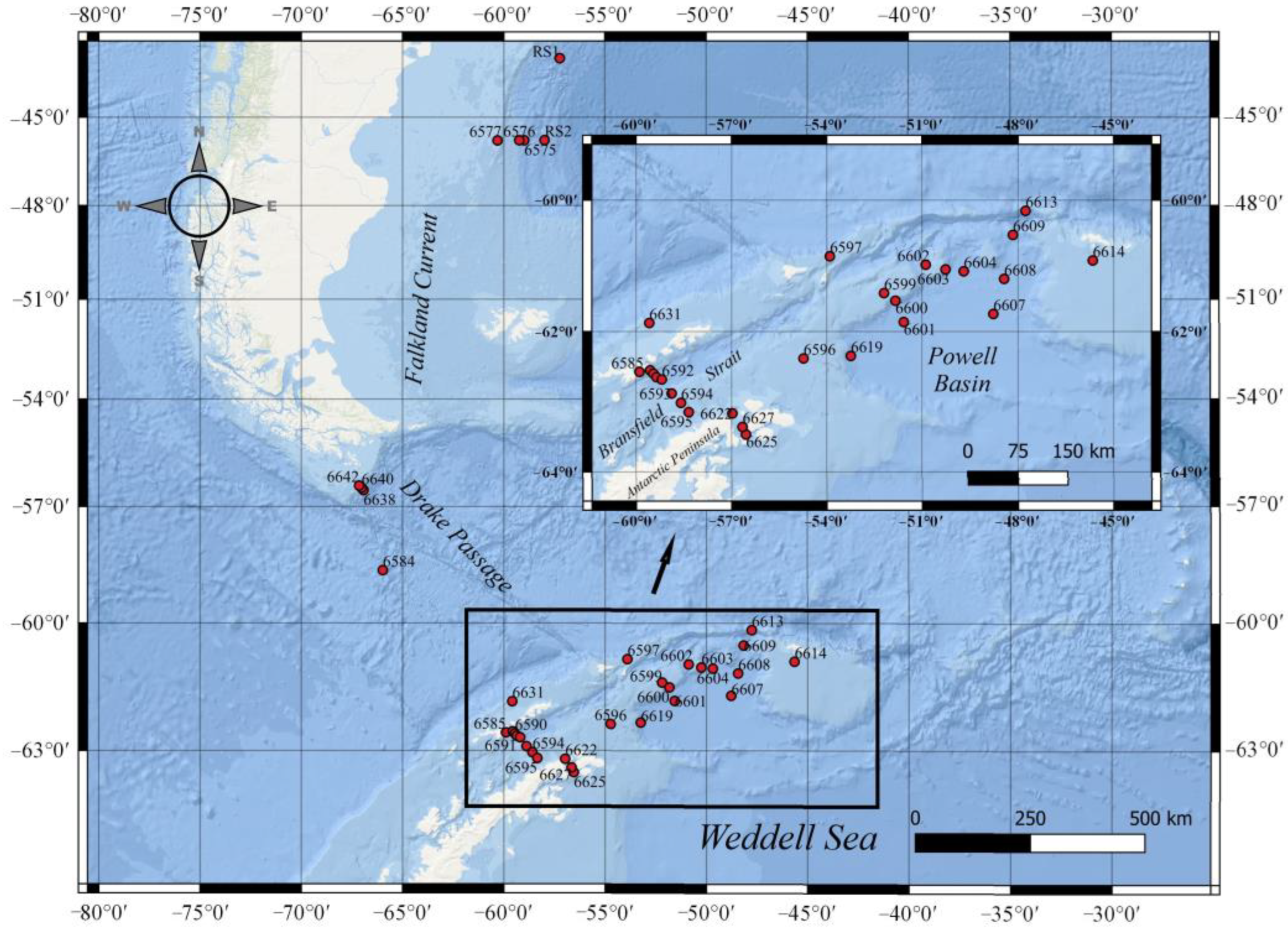
2.1. Sampling
2.2. Chlorophyll a Concentration
2.3. Particulate Absorption
2.4. CDOM Absorption
3. Results
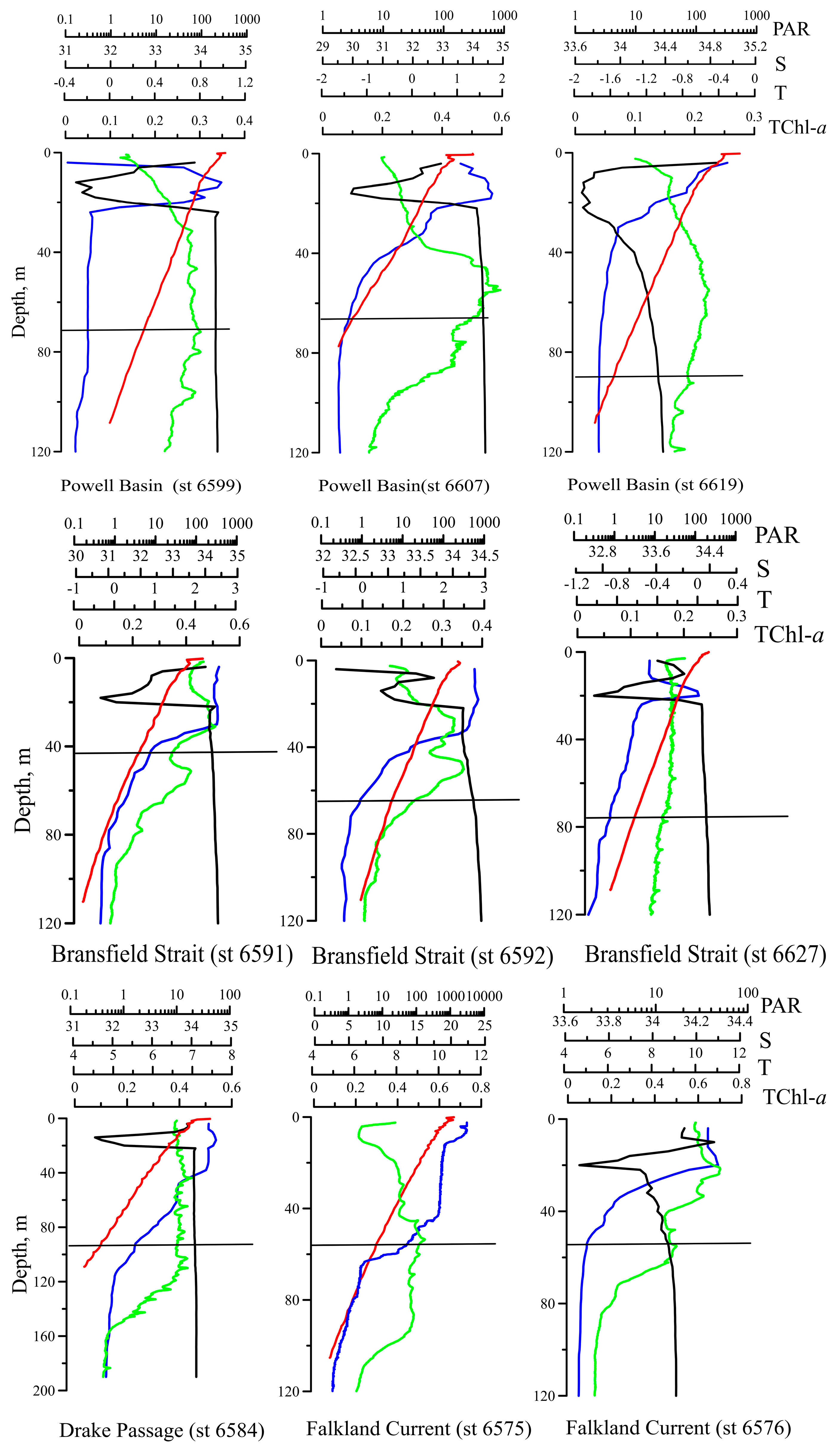
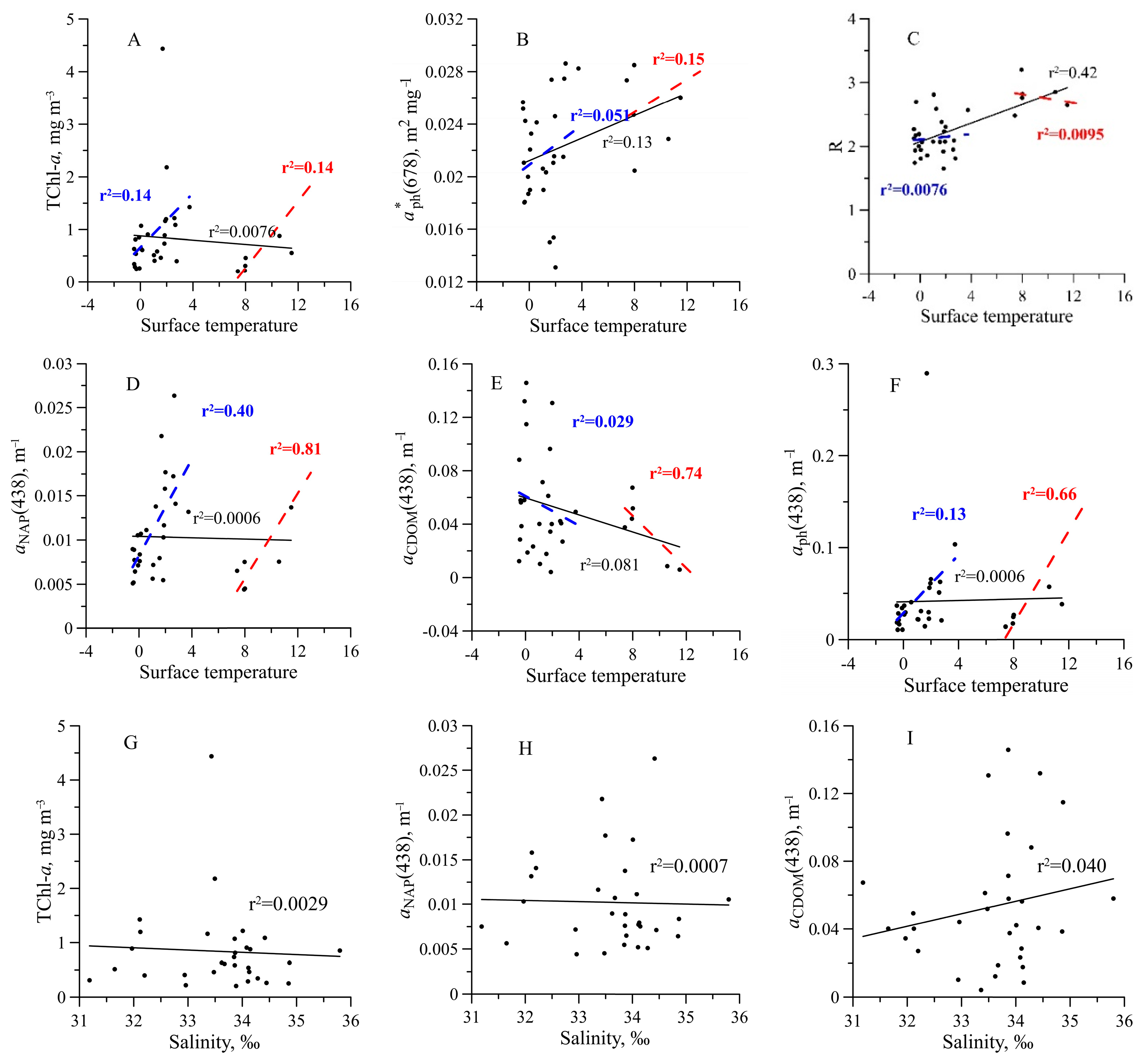
3.1. Hydrophysical Characteristics
3.2. Chlorophyll a Concentration
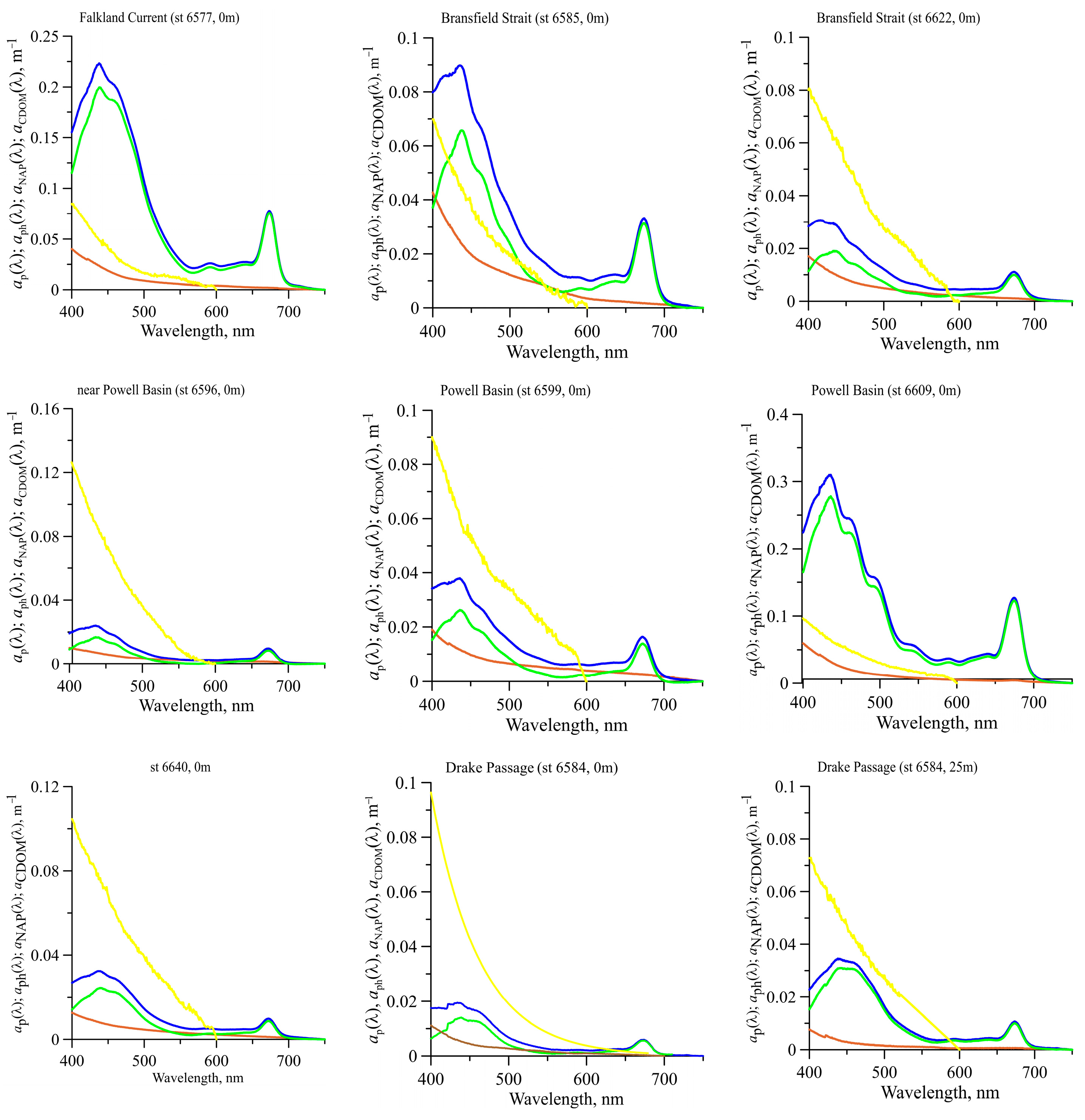
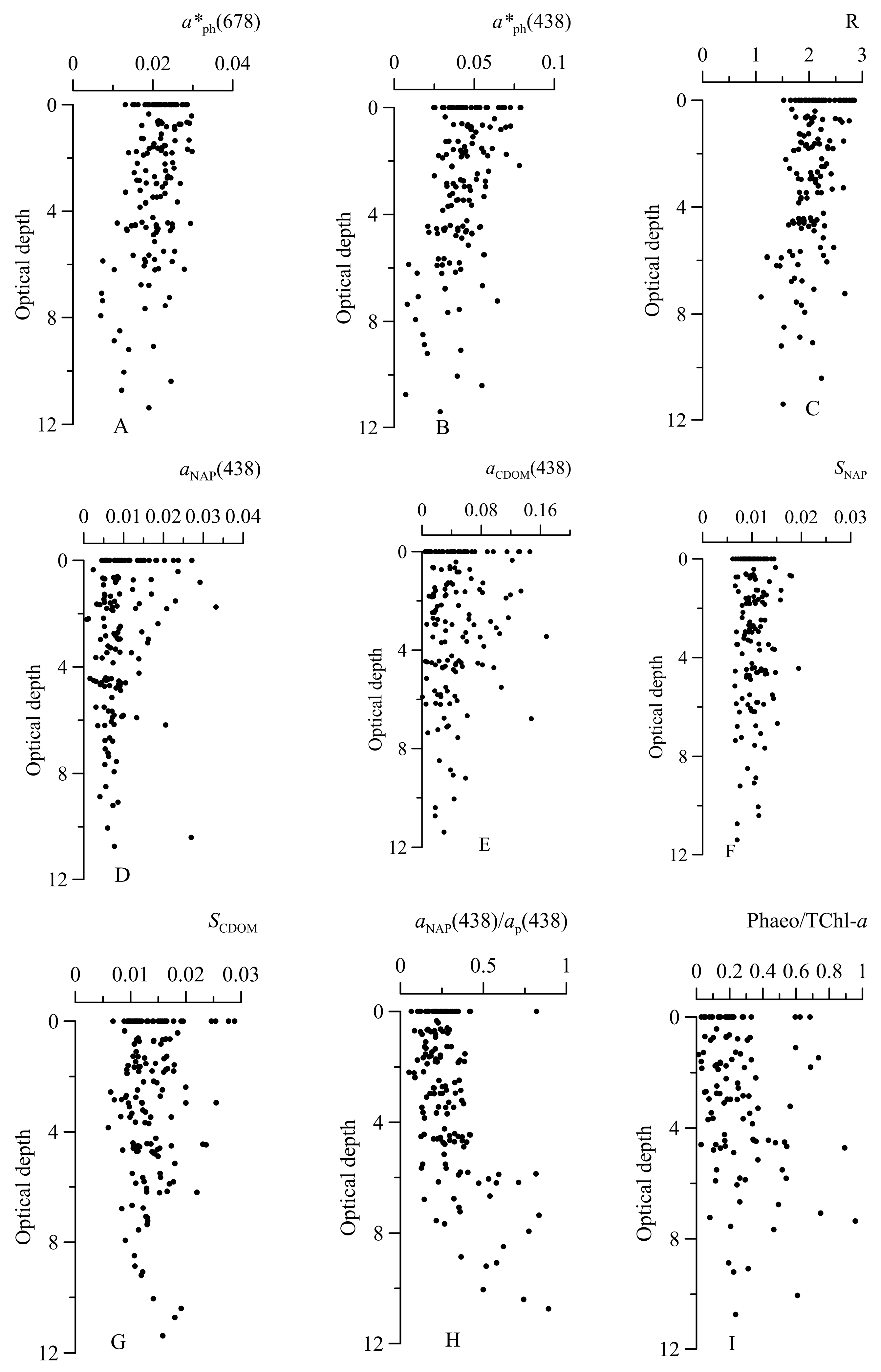
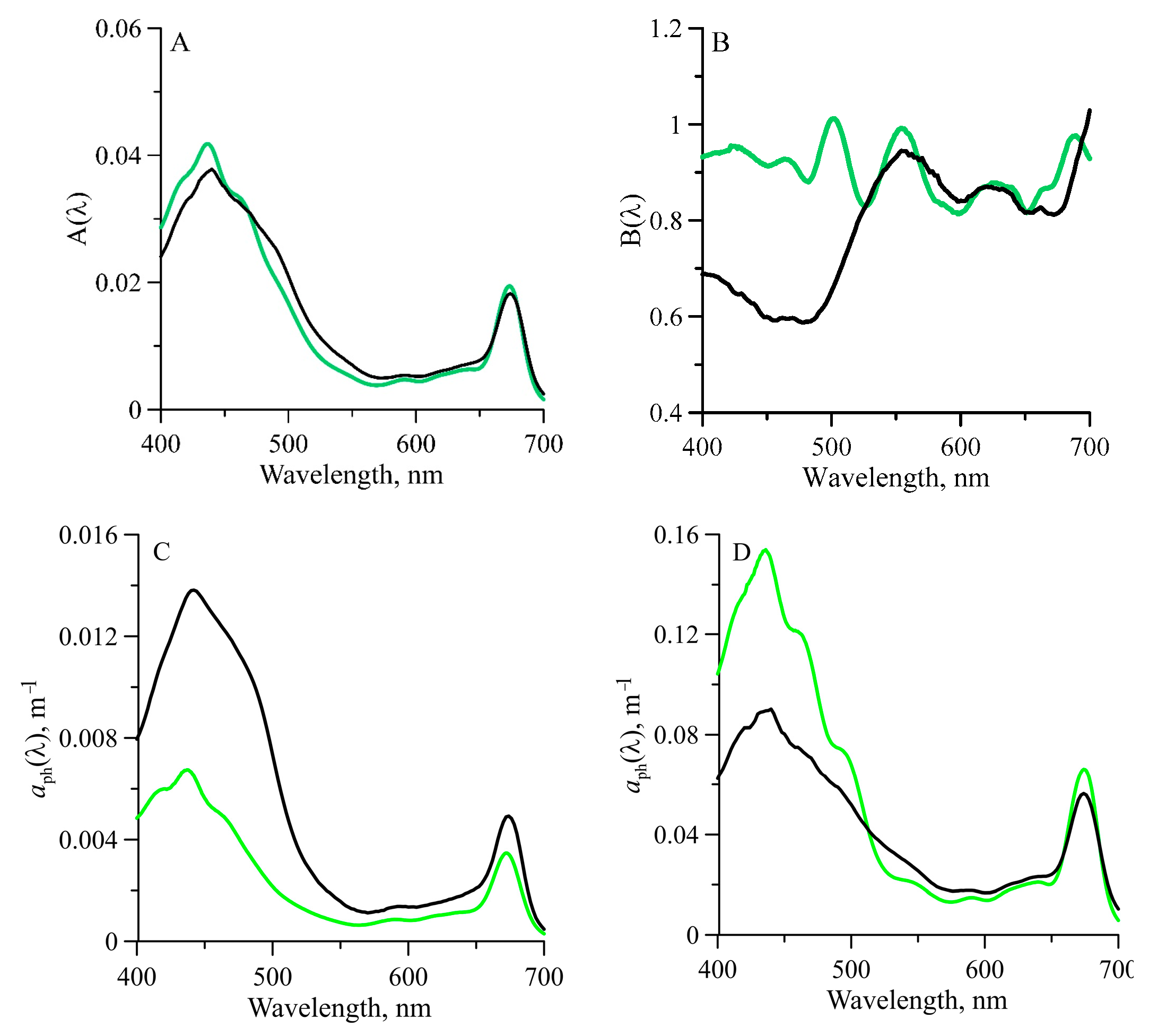
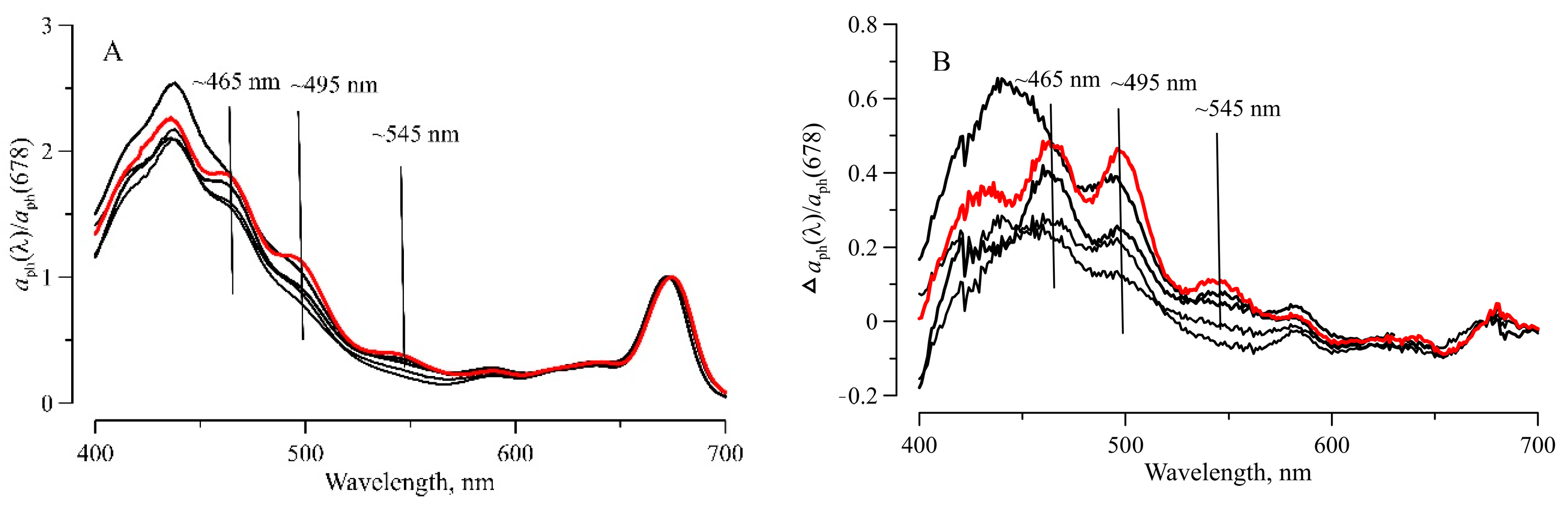
3.3. Light Absorption by Phytoplankton
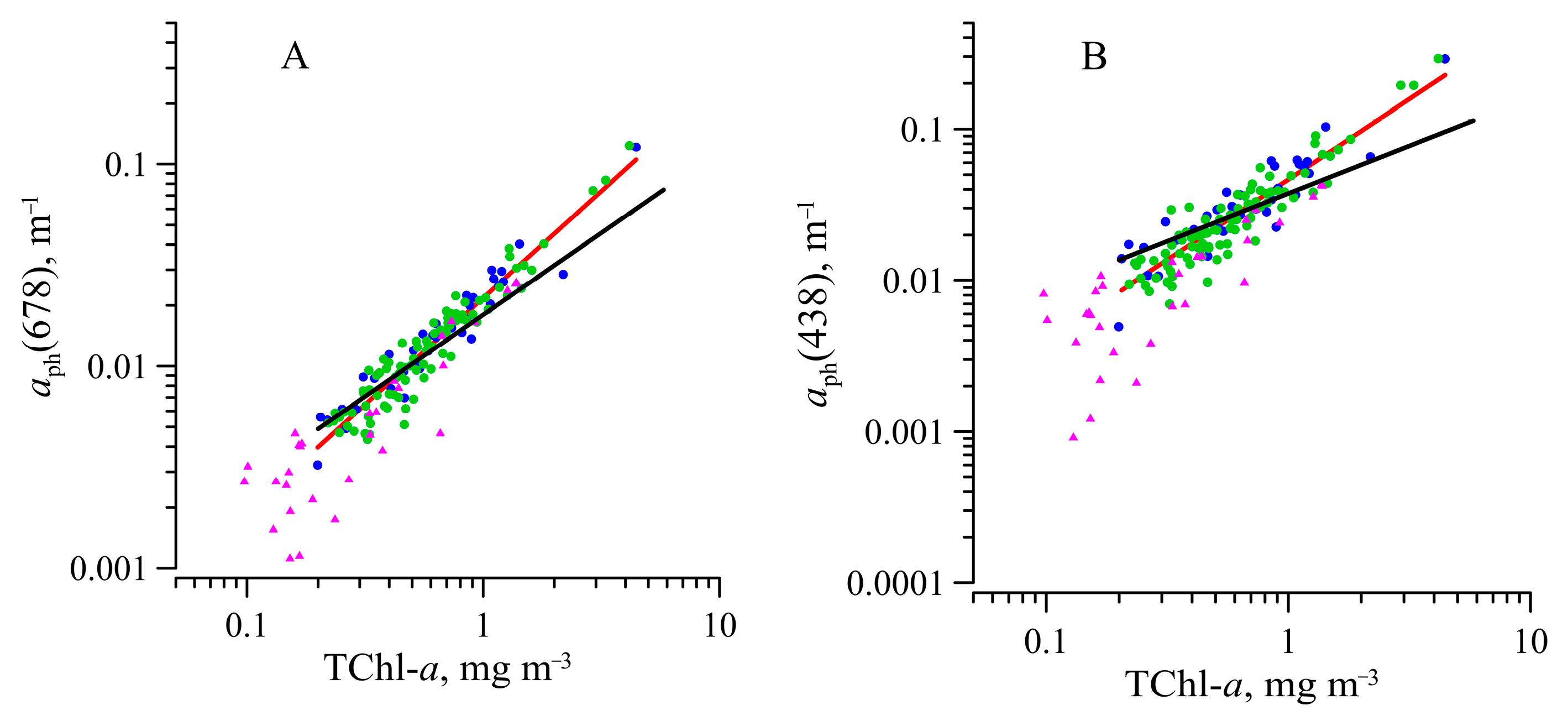
3.4. Parameterization of a Link between Phytoplankton Absorption Coefficient and Sum of Chlorophyll a and Phaeopigments Concentration
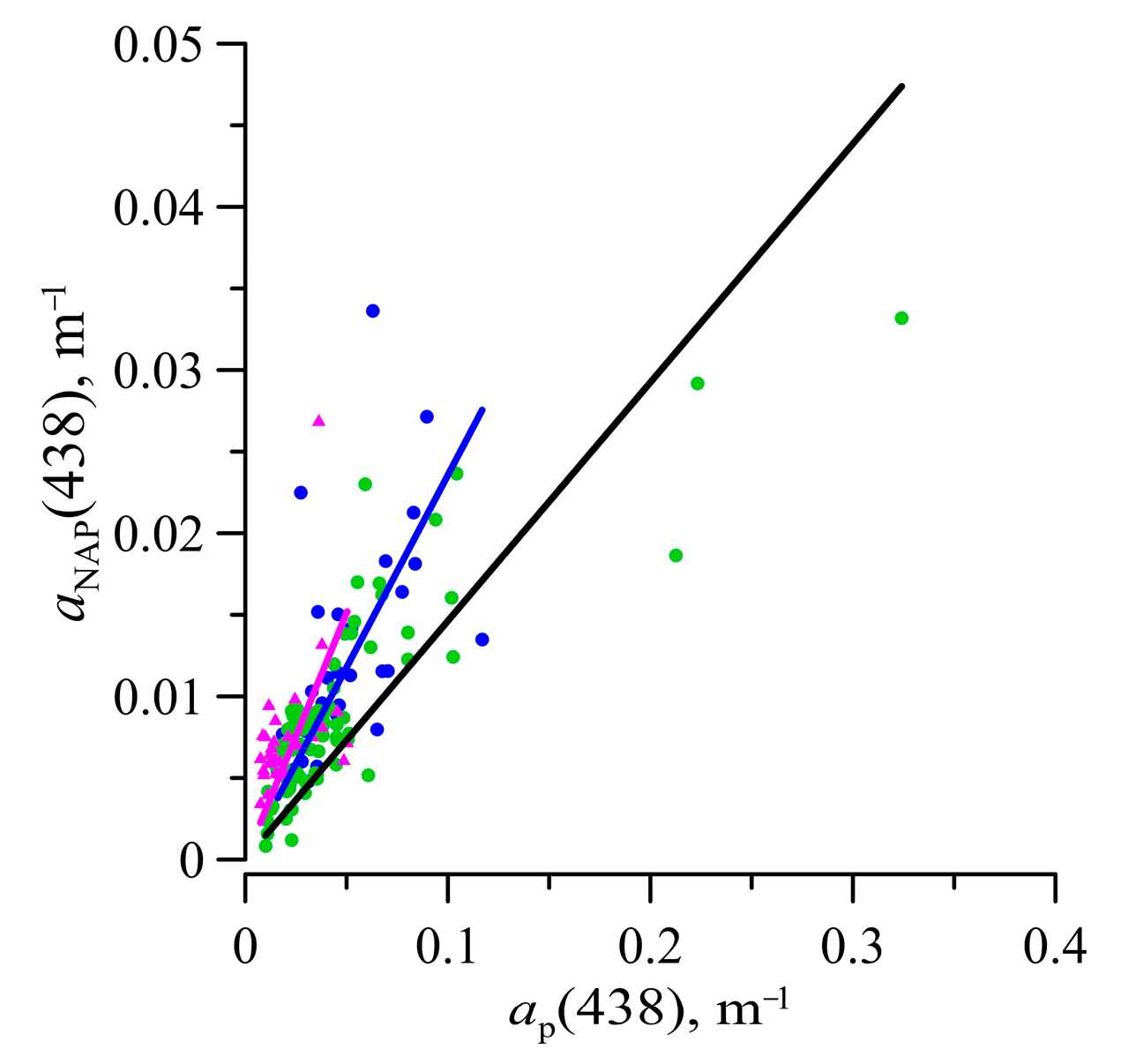
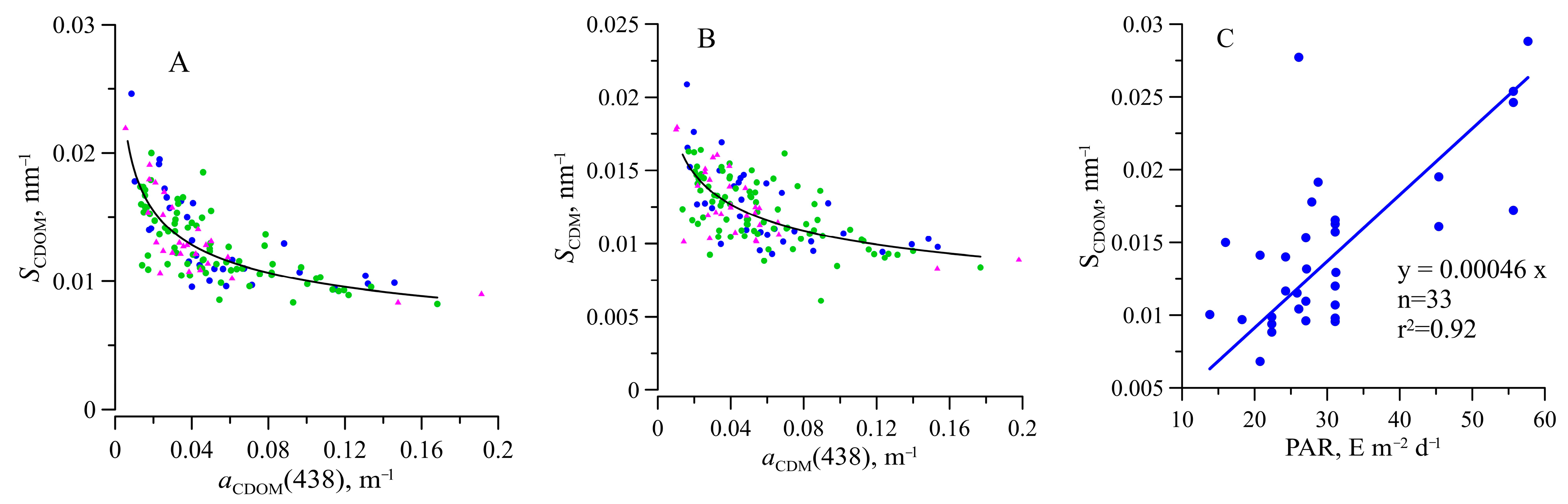
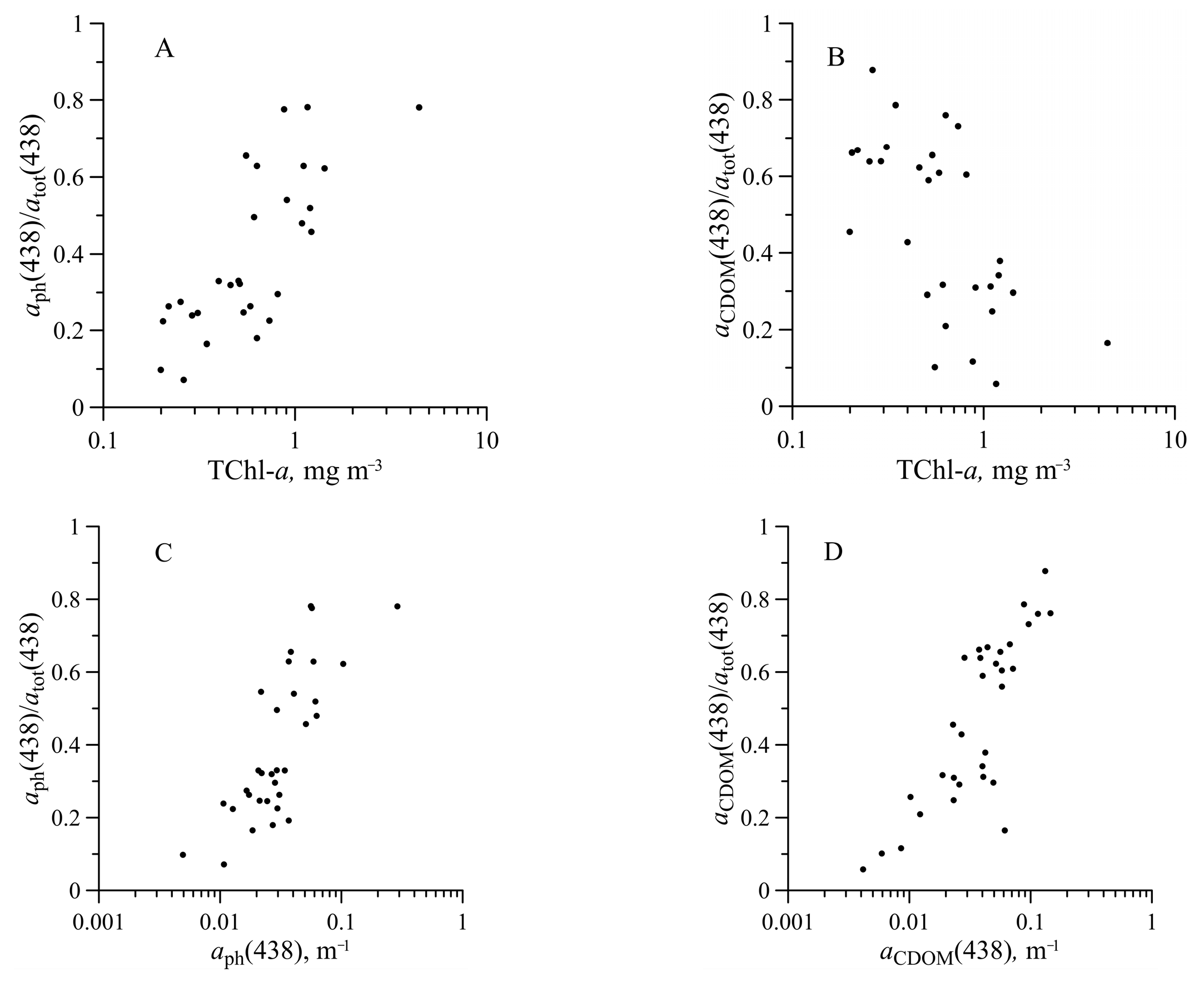
3.5. Light Absorption by NAP, CDOM, and CDM
3.6. Absorption Budget
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kerr, R.; Mata, M.M.; Mendes, C.R.B.; Secchi, E.R. Northern Antarctic Peninsula: A marine climate hotspot of rapid changes on ecosystems and ocean dynamic. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 149, 4–9. [Google Scholar] [CrossRef]
- CCAMLR. Statistical Bulletin; Commission for the Conservation of Antarctic Marine Living Resources: Hobart, Australia, 2018; p. 30. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture; Food and Agriculture Organization: Rome, Italy, 2018. [Google Scholar]
- Grant, S.M.; Hill, S.L.; Trathan, P.N.; Murphy, E.J. Ecosystem services of the Southern Ocean: Trade-offs in decision-making. Antarct. Sci. 2013, 25, 603–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, A.; Siegel, V.; Pakhomov, E.; Rothery, P. Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature 2004, 432, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.; Murphy, E.J.; Meredith, M.P.; King, J.C.; Peck, L.S.; Barnes, D.K.; Smith, R.C. Climate change and the marine ecosystem of the western Antarctic Peninsula. Phil. Trans. R. Soc. 2007, 362, 149–166. [Google Scholar] [CrossRef] [Green Version]
- Ducklow, H.W.; Baker, K.; Martinson, D.G.; Quetin, L.B.; Ross, R.M.; Smith, R.C.; Stammerjohn, S.E.; Vernet, M.; Fraser, W. Marine ecosystems: The West Antarctic Peninsula. Phil. Trans. R. Soc. 2007, 362, 67–94. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; in press. [Google Scholar]
- Pan, B.J.; Vernet, M.; Reynolds, R.A.; Mitchell, B.G. The optical and biological properties of glacial meltwater in an Antarctic fjord. PLoS ONE 2019, 14, e0211107. [Google Scholar] [CrossRef] [Green Version]
- Morley, S.A.; Abele, D.; Barnes, D.K.A.; Cárdenas, C.A.; Cotté, C.; Gutt, J.; Henley, S.F.; Höfer, J.; Hughes, K.A.; Martin., S.M.; et al. Global Drivers on Southern Ocean Ecosystems: Changing Physical Environments and Anthropogenic Pressures in an Earth System. Front. Mar. Sci. 2020, 7, 547188. [Google Scholar] [CrossRef]
- Pitchford, J.W.; Brindley, J. Iron limitation, grazing pressure and oceanic high nutrient-low chlorophyll (HNLC) regions. J. Plankton Res. 1999, 21, 525–547. [Google Scholar] [CrossRef] [Green Version]
- Pollard, R.; Tréguer, P.; Read, J. Quantifying nutrient supply to the Southern Ocean. J. Geophys. Res. Ocean. 2006, 111, C05011. [Google Scholar] [CrossRef]
- Mitchell, B.G. Predicative bio-optical relationships for polar oceans and marginal ice zones. J. Mar. Syst. 1992, 3, 91–105. [Google Scholar] [CrossRef]
- Kerkar, A.U.; Tripathy, S.C.; Minu, P.; Baranval, N.; Sabu, P.; Patra, S.; Mishra, R.K.; Sarkar, A. Variability in primary productivity and bio-optical properties in the Indian sector of the Southern Ocean during an austral summer. Polar Biol. 2020, 43, 1469–1492. [Google Scholar] [CrossRef]
- Boyd, P.W.; Jickells, T.; Law, C.S.; Blain, S.; Boyle, E.A.; Buesseler, K.O.; Coale, K.H.; Cullen, J.J.; de Baar, H.J.W.; Follows, M.; et al. Mesoscale iron enrichment experiments 1993-2005: Synthesis and future directions. Science 2007, 315, 612–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadley, M.R.; Jickells, T.D.; Heywood, K.J. The role of iron sources and transport for Southern Ocean productivity. Deep. Res. Part I Oceanogr. Res. Pap. 2014, 87, 82–94. [Google Scholar] [CrossRef]
- Yoon, J.-E.; Yoo, K.-C.; Macdonald, A.M.; Yoon, H.-I.; Park, K.-T.; Yang, E.J.; Kim, H.-C.; Lee, J.I.; Lee, M.K.; Jung, J.; et al. Reviews and syntheses: Ocean iron fertilization experiments—Past, present, and future looking to a future Korean Iron Fertilization Experiment in the Southern Ocean (KIFES) project. Biogeosciences 2018, 15, 5847–5889. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.; Jochens, A.E.; Kennicutt li, M.C.; Biggs, D.C. Spatial and temporal variability of phytoplankton biomass and community structure over the continental margin of the northeast Gulf of Mexico based on pigment analysis. Cont. Shelf Res. 2003, 23, 1–17. [Google Scholar] [CrossRef]
- Wright, S.W.; Jeffrey, S.W. Pigment markers for phytoplankton production. Handb. Environ. Chem. 2006, 2, 71–104. [Google Scholar] [CrossRef]
- Garcia, C.A.E.; Garcia, V.M.T.; McClain, C.R. Evaluation of SeaWiFS chlorophyll algorithms in the southwestern Atlantic and Southern Oceans. Remote Sens. Environ. 2005, 95, 125–137. [Google Scholar] [CrossRef]
- Dierssen, H.M.; Smith, R.C. Bio-optical properties and remote sensing ocean color algorithms for Antarctic Peninsula waters. J. Geophys. Res. 2000, 105, 26301–26312. [Google Scholar] [CrossRef]
- Szeto, M.; Werdell, P.J.; Moore, T.S.; Campbell, J.W. Are the world’s oceans optically different? J. Geophys. Res. 2011, 116, C00H04. [Google Scholar] [CrossRef] [Green Version]
- Pereira, E.S.; Garcia, C.A. Evaluation of satellite-derived MODIS chlorophyll algorithms in the northern Antarctic Peninsula. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 149, 124–137. [Google Scholar] [CrossRef]
- Ferreira, A.; Brito, A.C.; Mendes, C.R.B.; Brotas, V.; Costa, R.R.; Guerreiro, C.V.; Sá, C.; Jackson, T. OC4-SO: A New Chlorophyll-a Algorithm for theWestern Antarctic Peninsula Using Multi-Sensor Satellite Data. Remote Sens. 2022, 14, 1052. [Google Scholar] [CrossRef]
- Johnson, R.; Strutton, P.G.; Wright, S.W.; McMinn, A.; Meiners, K.M. Three improved satellite chlorophyll algorithms for the Southern Ocean. J. Geophys. Res. Ocean. 2013, 118, 3694–3703. [Google Scholar] [CrossRef]
- Ferreira, A.; Ciotti, Á.M.; Mendes, C.R.B.; Uitz, J.; Bricaud, A. Phytoplankton light absorption and the package effect in relation to photosynthetic and photoprotective pigments in the northern tip of Antarctic Peninsula. J. Geophys. Res. 2017, 122, 7344–7363. [Google Scholar] [CrossRef]
- Jena, B. The effect of phytoplankton pigment composition and packaging on the retrieval of chlorophyll-a concentration from satellite observations in the Southern Ocean. Int. J. Remote Sens. 2017, 38, 3763–3784. [Google Scholar] [CrossRef]
- Kerkar, A.U.; Tripathy, S.C.; Pandi, S.R. Bio-optical depiction of a polar ocean under global change: Exploring the regional absorption traits. Glob. Planet. Change 2022, 213, 103818. [Google Scholar] [CrossRef]
- Reynolds, R.A.; Stramski, D.; Mitchell, B.G. A chlorophyll-dependent semianalytical reflectance model derived from field measurements of absorption and backscattering coefficients within the Southern Ocean. J. Geophys. Res. Ocean. 2001, 106, 7125–7138. [Google Scholar] [CrossRef]
- Kirk, J.T.O. Light and Photosynthesis in Aquatic Ecosystems; Cambridge University Press: Cambridge, UK, 2011; p. 649. [Google Scholar]
- Sosik, H.; Vernet, M.; Mitchell, B.G. A comparison of particulate absorption properties between high- and mid-latitude surface waters. Antarct. JUS 1992, 27, 162–164. [Google Scholar]
- Robinson, C.M.; Huot, Y.; Schuback, N.; Ryan-Keogh, T.J.; Thomalla, S.J.; Antoine, D. High latitude Southern Ocean phytoplankton have distinctive bio-optical properties. Opt. Express 2021, 29, 21084–21112. [Google Scholar] [CrossRef]
- Ferreira, A.; Ciotti, Á.M.; Garcia, C.A.E. Bio-optical characterization of the northern Antarctic Peninsula waters: Absorption budget and insights on particulate backscattering. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 149, 138–149. [Google Scholar] [CrossRef]
- Suslin, V.; Churilova, T. The Black Sea regional algorithm of separation of light absorption by phytoplankton and colored detrital matter using ocean color scanner’s bands from 480-560 nm. Int. J. Remote Sens. 2016, 37, 4380–4400. [Google Scholar] [CrossRef]
- Churilova, T.; Moiseeva, N.; Efimova, T.; Suslin, V.; Krivenko, O.; Zemlianskaia, E. Annual variability in light absorption by particles and colored dissolved organic matter in coastal waters of Crimea (the Black Sea). In Proceedings of the 23rd International Symposium on Atmospheric and Ocean Optics: Atmospheric Physics, Irkutsk, Russia, 3–7 July 2017; SPIE: Bellingham, WA, USA, 2017; p. 10466. [Google Scholar] [CrossRef]
- Churilova, T.; Suslin, V. Parameterization of light absorption by all in-water optically active components in the Black Sea: Impact for underwater irradiance and primary production modeling. In Proceedings of the Coastal to Global Operational Oceanography: Achievements and Challenges: Proceedings of the Fifth International Conference on EuroGOOS, Exeter, UK, 20–22 May 2010; EuroGOOS Publication: Brussels, Belgium, 2010; pp. 199–205. [Google Scholar] [CrossRef]
- Suslin, V.V.; Churilova, T.Y.; Lee, M.; Moncheva, S.; Finenko, Z.Z. Comparison of the Black Sea chlorophyll-a algorithms for SeaWiFS and MODIS instruments. Fundamentalnaya i Prikladnaya Gidrofizika 2018, 11, 64–72. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Lorenzen, C.J. Determination of chlorophyll and pheo-pigments: Spectrophotometric equations. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- Oceancolor. Level-3 Browser. Available online: https://oceancolor.gsfc.nasa.gov/l3/ (accessed on 1 December 2022).
- Oceancolor. Chlorophyll A. Available online: https://oceancolor.gsfc.nasa.gov/atbd/chlor_a (accessed on 1 December 2022).
- O’Reilly, J.E.; McClain, C.R.; Ainsworth, E.; Barnes, R.; Eplee, R.E., Jr.; Patt, F.S.; Robinson, W.; Wang, M.; Bailey, S. SeaWiFS Postlaunch Calibration and Validation Analyses. NASA Tech. Memo. 2000, 11, 49. [Google Scholar]
- Hu, C.; Lee, Z.; Franz, B. Chlorophyll a algorithms for oligotrophic oceans: A novel approach based on three-band reflectance difference. J. Geophys. Res. 2012, 117, C1. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, J.E.; Werdell, P.J. Chlorophyll algorithms for ocean color sensors—OC4, OC5 & OC6. Remote Sens. Environ. 2019, 229, 32–47. [Google Scholar] [CrossRef]
- Seegers, B.N.; Stumpf, R.P.; Schaeffer, B.A.; Loftin, K.A.; Werdell, P.J. Performance metrics for the assessment of satellite data products: An ocean color case study. Opt. Express 2018, 26, 7404–7422. [Google Scholar] [CrossRef] [Green Version]
- Neeley, A.R.; Mannino, A. Ocean Optics and Biogeochemistry Protocols for Satellite Ocean Colour Sensor Validation (v. 1. 0); IOCCG: Dartmouth, NS, Canada, 2018. [Google Scholar]
- Kishino, M.; Takahashi, N.; Okami, N.; Ichimura, S. Estimation of the spectral absorption coefficients of phytoplankton in the sea. Bull. Mar. Sci. 1985, 37, 634–642. [Google Scholar]
- Roesler, C.S.; Perry, M.J. In situ phytoplankton absorption, fluorescence emission, and particulate backscattering spectra determined from reflectance. J. Geophys. Res. 1995, 100, 13279–13294. [Google Scholar] [CrossRef]
- Mitchell, B.G. Algorithms for determining the absorption coefficient for aquatic particulates using the quantitative filter technique. In Ocean Optics X; SPIE: Bellingham, WA, USA, 1990; Volume 1302, pp. 137–148. [Google Scholar]
- Bricaud, A.; Babin, M.; Morel, A.; Claustre, H. Variability in the chlorophyll-specific absorption coefficients of natural phytoplankton: Analysis and parameterization. J. Geophys. Res. 1995, 100, 13321–13332. [Google Scholar] [CrossRef]
- Mitchell, B.G.; Kahru, M.; Wieland, J.; Stramska, M. Determination of spectral absorption coefficients of particles, dissolved materials and phytoplankton for discrete water samples. Ocean Opt. Prot. Satel. Ocean Col. Sen. Valid. Revis. 2003, 3, 231. [Google Scholar]
- Babin, M.; Stramski, D.; Ferrari, G.M.; Claustre, H.; Bricaud, A.; Obolensky, G.; Hoepffner, N. Variations in the light absorption coefficients of phytoplankton, non-algal particles, and dissolved organic matter in coastal waters around Europe. J. Geophys. Res. 2003, 108, C7. [Google Scholar] [CrossRef]
- Heidenreich, K.M.; Richardson, T.L. Photopigment, absorption, and growth responses of marine cryptophytes to varying spectral irradiance. J. of Phycol. 2020, 56, 507–520. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.H. Handbook of Biolological Statistics; Sparky House Publishing: Baltimore, MD, USA, 2014. [Google Scholar]
- Morozov, E.G.; Krechik, V.A.; Frey, D.I.; Polukhin, A.A.; Artemiev, V.A.; Kasyan, V.V.; Sapozhnikov, P.V.; Mukhametianov, R.Z. Frontal Zone Between Relatively Warm and Cold Waters in the Northern Weddell Sea. In Antarctic Peninsula Region of the Southern Ocean Springer; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 31–53. [Google Scholar]
- Stepanova, S.V.; Polukhin, A.A.; Borisenko, G.V.; Chultsova, A.L.; Marina, E.N.; Popov, O.S.; Seliverstova, A.M.; Vidnichuk, A.V.; Tishchenko, P.P. Hydrochemical structure of waters in the Northern Weddell Sea in Austral summer 2020. In Antarctic Peninsula Region of the Southern Ocean: Oceanography and Ecology; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 159–174. [Google Scholar]
- Figueiras, F.G.; Arbones, B.; Estrada, M. Implications of bio-optical modeling of phytoplankton photosynthesis in Antarctic waters: Further evidence of no light limitation in the Bransfield Strait. Limnol. Oceanogr. 1999, 44, 1599–1608. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, F.L. Bio-optical characteristics of Gerlache and Bransfield Strait waters during an Antarctic summer cruise. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 675–691. [Google Scholar] [CrossRef]
- Smith, R.C.; Baker, K.S.; Byers, M.L.; Stammerjohn, S.E. Primary productivity of the Palmer Long Term Ecological Research Area and the Southern Ocean. J. Mar. Syst. 1998, 17, 245–259. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis, 2nd. ed.; Princeton University Press: Princeton, NJ, USA, 2007; p. 512. [Google Scholar]
- Holeton, C.L.; Nédélec, F.; Sanders, R.; Brown, L.; Moore, C.M.; Stevens, D.P.; Heywood, K.J.; Statham, P.J.; Lucas, C.H. Physiological state of phytoplankton communities in the Southwest Atlantic sector of the Southern Ocean, as measured by fast repetition rate fluorometry. Polar Biol. 2005, 29, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Hiscock, M.R.; Lance, V.P.; Apprill, A.M.; Bidigare, R.R.; Johnson, Z.I.; Mitchell, B.G.; Smith, W.O.; Barber, R.T. Photosynthetic maximum quantum yield increases are an essential component of the Southern Ocean phytoplankton response to iron. Proc. Natl. Acad. Sci. USA 2008, 105, 4775–4780. [Google Scholar] [CrossRef] [Green Version]
- Le Quéré, C.; Buitenhuis, E.T.; Moriarty, R.; Alvain, S.; Aumont, O.; Bopp, L.; Chollet, S.; Enright, C.; Franklin, D.J.; Geider, R.J.; et al. Role of zooplankton dynamics for Southern Ocean phytoplankton biomass and global biogeochemical cycles. Biogeosciences 2016, 13, 4111–4133. [Google Scholar] [CrossRef] [Green Version]
- Bricaud, A.; Morel, A.; Prieur, L. Absorption by dissolved organic matter of the sea (yellow substance) in the UV and visible domains. Limnol. Oceanogr. 1981, 26, 43–53. [Google Scholar] [CrossRef]
- Cullen, J.J.; Lewis, M.R. The kinetics of algal photoadaptation in the context of vertical mixing. J. Plank. Res. 1988, 10, 1039–1063. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; Mantoura, R.F.C.; Wright, S.W. Phytoplankton Pigments in Oceanography: Guidelines to Modern Method; UNESCO publishing: Paris, France, 1997. [Google Scholar]
- Morel, A.; Bricaud, A. Theoretical results concerning light absorption in a discrete medium and application to specific absorption of phytoplankton. Deep-Sea Res. 1981, 28, 1375–1393. [Google Scholar] [CrossRef]
- MacIntyre, H.L.; Kana, T.M.; Anning, J.; Geider, R. Photoacclimation of photosynthesis irradiance response curves and photosynthetic pigments in microalgae and cyanobacteria. J. Phycol. 2002, 38, 17–38. [Google Scholar] [CrossRef] [Green Version]
- Mukhanov, V.; Sakhon, E.; Polukhin, A.; Artemiev, V.; Morozov, E.; Tsai, A.-Y. Cryptophyte and Photosynthetic Picoeukaryote Abundances in the Bransfield Strait during Austral Summer. Water 2022, 14, 185. [Google Scholar] [CrossRef]
- Schofield, O.; Saba, G.; Coleman, K.; Carvalho, F.; Couto, N.; Ducklow, H.; Finkel, Z.; Irwin, A.; Kahl, A.; Miles, T.; et al. Decadal variability in coastal phytoplankton community composition in a changing West Antarctic Peninsula. Deep Sea Res. Part I Oceanogr. Res. Pap. 2017, 124, 42–54. [Google Scholar] [CrossRef]
- Moore, L.R.; Georicke, R.; Chisholm, S.W. Comparative physiology of Synechococcus and Prochlorococcus: Influence of light and temperature on growth, pigments, fluorescence and absorptive properties. Mar. Ecol. Prog. Ser. 1995, 116, 259–275. [Google Scholar] [CrossRef]
- Novarino, G. A companion to the identification of cryptomonad flagellates (Cryptophyceae = Cryptomonadea). Hydrobiologia 2003, 502, 225–270. [Google Scholar] [CrossRef]
- Churilova, T.Y.; Suslin, V.V.; Rylkova, O.A. Parameterization of light absorption by all optically active components in the Black Sea. Ecolog. Safety Coast. and Shelf Areas Complex Use Shelf Res. 2008, 16, 190–201. (In Russian) [Google Scholar]
- Mendes, C.R.B.; Tavano, V.M.; Dotto, T.S.; Kerr, R.; De Souza, M.S.; Garcia, C.A.E.; Secchi, E.R. New insights on the dominance of cryptophytes in Antarctic coastal waters: A case study in Gerlache Strait. Deep Sea Res. Part II 2018, 149, 161–170. [Google Scholar] [CrossRef]
- Morel, A.; Ahn, Y.-H. Optics of heterotrophic nanoflagellates and ciliates: A tentative assessment of their scattering role in oceanic waters compared to those of bacterial and algal cells. J. Mar. Res. 1991, 49, 177–202. [Google Scholar] [CrossRef]
- Stramski, D.; Kiefer, D.A. Can heterotrophic bacteria be important to marine light absorption? J. Plankton. Res. 1998, 20, 1489–1500. [Google Scholar] [CrossRef] [Green Version]
- Bricaud, A.; Morel, A.; Babin, M.; Allali, K.; Claustre, H. Variations of light absorption by suspended particles with chlorophyll a concentration in oceanic (case 1) waters: Analysis and implications for bio-optical models. J. Geophys. Res. 1998, 103, 31033–31044. [Google Scholar] [CrossRef]
- Efimova, T.; Churilova, T.; Moiseeva, N.; Zemlianskaia, E.; Dzhulay, A.; Krivenko, O. Dynamics in pigment concentration and light absorption by phytoplankton, non-algal particles and colored dissolved organic matter in the Black Sea coastal waters (near Sevastopol). In Proceedings of the 24th International Symposium on Atmospheric and Ocean Optics: Atmospheric Physics, Tomsk, Russia, 2–5 July 2018; SPIE: Bellingham, WA, USA, 2018; p. 10833. [Google Scholar] [CrossRef]
- Nelson, N.B.; Siegel, D.A. The global distribution and dynamics of chromophoric dissolved organic matter. Annu. Rev. Mar. Sci. 2013, 5, 447–476. [Google Scholar] [CrossRef] [Green Version]
- Helms, J.R.; Stubbins, A.; Ritchie, J.D.; Minor, E.C.; Kieber, D.J.; Mopper, K. Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnol. Oceanogr. 2008, 53, 955–969. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Retuerta, E.; Reche, I.; Pulido-Villena, E.; Agustí, S.; Duarte, C.M. Distribution and photoreactivity of chromophoric dissolved organic matter in the Antarctic Peninsula (Southern Ocean). Mar. Chem. 2010, 118, 129–139. [Google Scholar] [CrossRef]
- Ortega-Retuerta, E.; Frazer, T.K.; Duarte, C.M.; Ruiz-Halpern, S.; Tovar-Sánchez, A.; Arrieta, J.M.; Rechea, I. Biogeneration of chromophoric dissolved organic matter by bacteria and krill in the Southern Ocean. Limnol. Oceanogr. 2009, 54, 1941–1950. [Google Scholar] [CrossRef] [Green Version]
- Del Castillo, C.E.; Miller, R.L. Horizontal and vertical distributions of colored dissolved organic matter during the Southern Ocean Gas Exchange Experiment. J. Geophys. Res. Oceans 2011, 116, C4. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A(λ) | B(λ) | r2 | n | Region | Reference |
|---|---|---|---|---|---|
| λ = 438 nm | |||||
| 0.042 | 0.93 | 0.83 | 126 | Bransfield Strait, Falkland Current, Drake Passage, and the Powell Basin | This research |
| 0.038 | 0.63 | 0.90 | 815 | Different regions of the global ocean, not including the Southern Ocean | Bricaud et al., 1995 |
| 0.042 | 0.74 | 0.90 | 455 | Gerlache Strait, the Bransfield Strait, and the northwestern Weddell Sea | Ferreira et al., 2017 |
| λ = 678 nm | |||||
| 0.018 | 0.92 | 0.92 | 126 | Bransfield Strait, Falkland Current, Drake Passage, and the Powell Basin | This research |
| 0.017 | 0.82 | 0.94 | 815 | Different regions of the global ocean, not including the Southern Ocean | Bricaud et al., 1995 |
| 0.020 | 0.79 | 0.93 | 455 | Gerlache Strait, the Bransfield Strait, and the northwestern Weddell Sea | Ferreira et al., 2017 |
| λ | A(λ) | B(λ) | λ | A(λ) | B(λ) |
|---|---|---|---|---|---|
| 400 | 0.0286 | 0.9319 | 478 | 0.0257 | 0.8877 |
| 402 | 0.0295 | 0.9347 | 480 | 0.0246 | 0.8822 |
| 404 | 0.0304 | 0.9356 | 482 | 0.0236 | 0.8803 |
| 406 | 0.0315 | 0.9371 | 484 | 0.0228 | 0.8859 |
| 408 | 0.0326 | 0.9396 | 486 | 0.0220 | 0.8960 |
| 410 | 0.0337 | 0.9407 | 488 | 0.0212 | 0.9127 |
| 412 | 0.0346 | 0.9410 | 490 | 0.0205 | 0.9316 |
| 414 | 0.0354 | 0.9420 | 492 | 0.0198 | 0.9529 |
| 416 | 0.0360 | 0.9425 | 494 | 0.0191 | 0.9723 |
| 418 | 0.0364 | 0.9440 | 496 | 0.0183 | 0.9896 |
| 420 | 0.0367 | 0.9450 | 498 | 0.0176 | 1.0027 |
| 422 | 0.0373 | 0.9551 | 500 | 0.0167 | 1.0103 |
| 424 | 0.0377 | 0.9545 | 502 | 0.0159 | 1.0122 |
| 426 | 0.0383 | 0.9530 | 504 | 0.0151 | 1.0071 |
| 428 | 0.0390 | 0.9531 | 506 | 0.0142 | 0.9961 |
| 430 | 0.0399 | 0.9505 | 508 | 0.0134 | 0.9807 |
| 432 | 0.0408 | 0.9471 | 510 | 0.0126 | 0.9599 |
| 434 | 0.0415 | 0.9433 | 512 | 0.0119 | 0.9382 |
| 436 | 0.0418 | 0.9399 | 514 | 0.0111 | 0.9130 |
| 438 | 0.0417 | 0.9344 | 516 | 0.0105 | 0.8905 |
| 440 | 0.0412 | 0.9304 | 518 | 0.0099 | 0.8694 |
| 442 | 0.0403 | 0.9249 | 520 | 0.0093 | 0.8534 |
| 444 | 0.0391 | 0.9221 | 522 | 0.0088 | 0.8415 |
| 446 | 0.0378 | 0.9191 | 524 | 0.0084 | 0.8330 |
| 448 | 0.0367 | 0.9166 | 526 | 0.0080 | 0.8296 |
| 450 | 0.0356 | 0.9136 | 528 | 0.0076 | 0.8314 |
| 452 | 0.0349 | 0.9141 | 530 | 0.0073 | 0.8377 |
| 454 | 0.0344 | 0.9154 | 532 | 0.0070 | 0.8470 |
| 456 | 0.0341 | 0.9179 | 534 | 0.0068 | 0.8617 |
| 458 | 0.0338 | 0.9222 | 536 | 0.0066 | 0.8787 |
| 460 | 0.0336 | 0.9242 | 538 | 0.0063 | 0.8943 |
| 462 | 0.0333 | 0.9262 | 540 | 0.0061 | 0.9133 |
| 464 | 0.0329 | 0.9274 | 542 | 0.0059 | 0.9309 |
| 466 | 0.0322 | 0.9270 | 544 | 0.0057 | 0.9451 |
| 468 | 0.0314 | 0.9242 | 546 | 0.0055 | 0.9599 |
| 470 | 0.0304 | 0.9196 | 548 | 0.0053 | 0.9706 |
| 472 | 0.0293 | 0.9134 | 550 | 0.0051 | 0.9802 |
| 474 | 0.0281 | 0.9047 | 552 | 0.0049 | 0.9886 |
| 476 | 0.0269 | 0.8957 | 554 | 0.0047 | 0.9925 |
| λ | A(λ) | B(λ) | λ | A(λ) | B(λ) |
| 556 | 0.0045 | 0.9909 | 630 | 0.0059 | 0.8759 |
| 558 | 0.0043 | 0.9832 | 632 | 0.006 | 0.8739 |
| 560 | 0.0041 | 0.9802 | 634 | 0.0061 | 0.8698 |
| 562 | 0.004 | 0.968 | 636 | 0.0062 | 0.8701 |
| 564 | 0.0039 | 0.9559 | 638 | 0.0063 | 0.8684 |
| 566 | 0.0039 | 0.9371 | 640 | 0.0063 | 0.8685 |
| 568 | 0.0038 | 0.9233 | 642 | 0.0063 | 0.8621 |
| 570 | 0.0038 | 0.9064 | 644 | 0.0063 | 0.8533 |
| 572 | 0.0039 | 0.8898 | 646 | 0.0063 | 0.8409 |
| 574 | 0.0039 | 0.8744 | 648 | 0.0064 | 0.8282 |
| 576 | 0.004 | 0.8603 | 650 | 0.0065 | 0.819 |
| 578 | 0.0041 | 0.8498 | 652 | 0.0068 | 0.8172 |
| 580 | 0.0042 | 0.8411 | 654 | 0.0074 | 0.8217 |
| 582 | 0.0043 | 0.8394 | 656 | 0.0082 | 0.8305 |
| 584 | 0.0045 | 0.8355 | 658 | 0.0094 | 0.8416 |
| 586 | 0.0046 | 0.8346 | 660 | 0.0108 | 0.8514 |
| 588 | 0.0047 | 0.8307 | 662 | 0.0124 | 0.8612 |
| 590 | 0.0047 | 0.8258 | 664 | 0.0142 | 0.8649 |
| 592 | 0.0047 | 0.8229 | 666 | 0.0159 | 0.867 |
| 594 | 0.0047 | 0.8174 | 668 | 0.0175 | 0.8667 |
| 596 | 0.0046 | 0.8163 | 670 | 0.0186 | 0.869 |
| 598 | 0.0045 | 0.8135 | 672 | 0.0193 | 0.874 |
| 600 | 0.0045 | 0.8148 | 674 | 0.0194 | 0.8845 |
| 602 | 0.0044 | 0.8182 | 676 | 0.0189 | 0.8991 |
| 604 | 0.0044 | 0.8227 | 678 | 0.0177 | 0.9163 |
| 606 | 0.0045 | 0.8302 | 680 | 0.0159 | 0.935 |
| 608 | 0.0046 | 0.8398 | 682 | 0.0138 | 0.9501 |
| 610 | 0.0047 | 0.8494 | 684 | 0.0114 | 0.9656 |
| 612 | 0.0049 | 0.8552 | 686 | 0.0092 | 0.9721 |
| 614 | 0.0051 | 0.8635 | 688 | 0.0072 | 0.9745 |
| 616 | 0.0052 | 0.8664 | 690 | 0.0056 | 0.975 |
| 618 | 0.0053 | 0.8696 | 692 | 0.0042 | 0.9692 |
| 620 | 0.0055 | 0.8705 | 694 | 0.0033 | 0.9612 |
| 624 | 0.0056 | 0.8777 | 696 | 0.0025 | 0.9491 |
| 622 | 0.0056 | 0.8737 | 698 | 0.002 | 0.9383 |
| 626 | 0.0057 | 0.8775 | 700 | 0.0016 | 0.8634 |
| 628 | 0.0058 | 0.878 |
| Satellite Scanners | n | bias | MAE |
|---|---|---|---|
| Merged ATV | 12 | 0.63 | 1.9 |
| Aqua-MODIS | 17 | 0.51 | 2.4 |
| S3A-OLCI | 8 | 0.54 | 1.8 |
| SNPP-VIIRS | 14 | 0.47 | 2.2 |
| without maximum value (one point highlighted in Figure 15) | |||
| Merged ATV | 11 | 0.55 | 1.8 |
| Aqua-MODIS | 16 | 0.45 | 2.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Churilova, T.; Moiseeva, N.; Skorokhod, E.; Efimova, T.; Buchelnikov, A.; Artemiev, V.; Salyuk, P. Parameterization of Light Absorption of Phytoplankton, Non-Algal Particles and Coloured Dissolved Organic Matter in the Atlantic Region of the Southern Ocean (Austral Summer of 2020). Remote Sens. 2023, 15, 634. https://doi.org/10.3390/rs15030634
Churilova T, Moiseeva N, Skorokhod E, Efimova T, Buchelnikov A, Artemiev V, Salyuk P. Parameterization of Light Absorption of Phytoplankton, Non-Algal Particles and Coloured Dissolved Organic Matter in the Atlantic Region of the Southern Ocean (Austral Summer of 2020). Remote Sensing. 2023; 15(3):634. https://doi.org/10.3390/rs15030634
Chicago/Turabian StyleChurilova, Tatiana, Natalia Moiseeva, Elena Skorokhod, Tatiana Efimova, Anatoly Buchelnikov, Vladimir Artemiev, and Pavel Salyuk. 2023. "Parameterization of Light Absorption of Phytoplankton, Non-Algal Particles and Coloured Dissolved Organic Matter in the Atlantic Region of the Southern Ocean (Austral Summer of 2020)" Remote Sensing 15, no. 3: 634. https://doi.org/10.3390/rs15030634