
Variability of Chl a Concentration of Priority Marine Regions of the Northwest of Mexico
 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
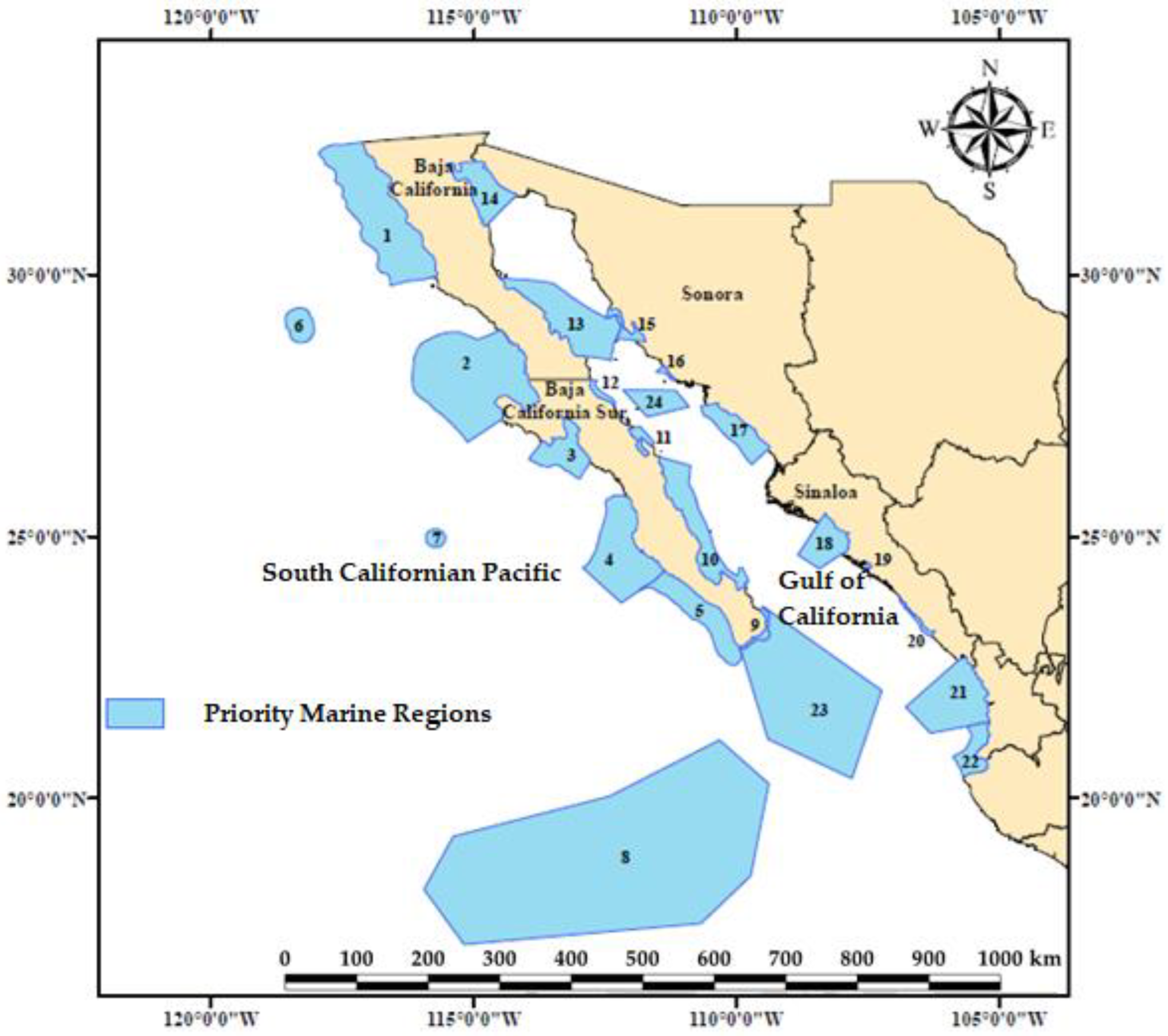
2.1. Study Area
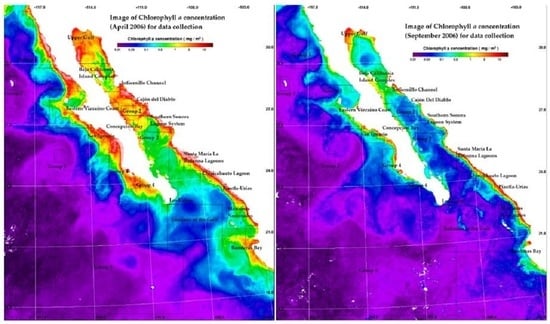
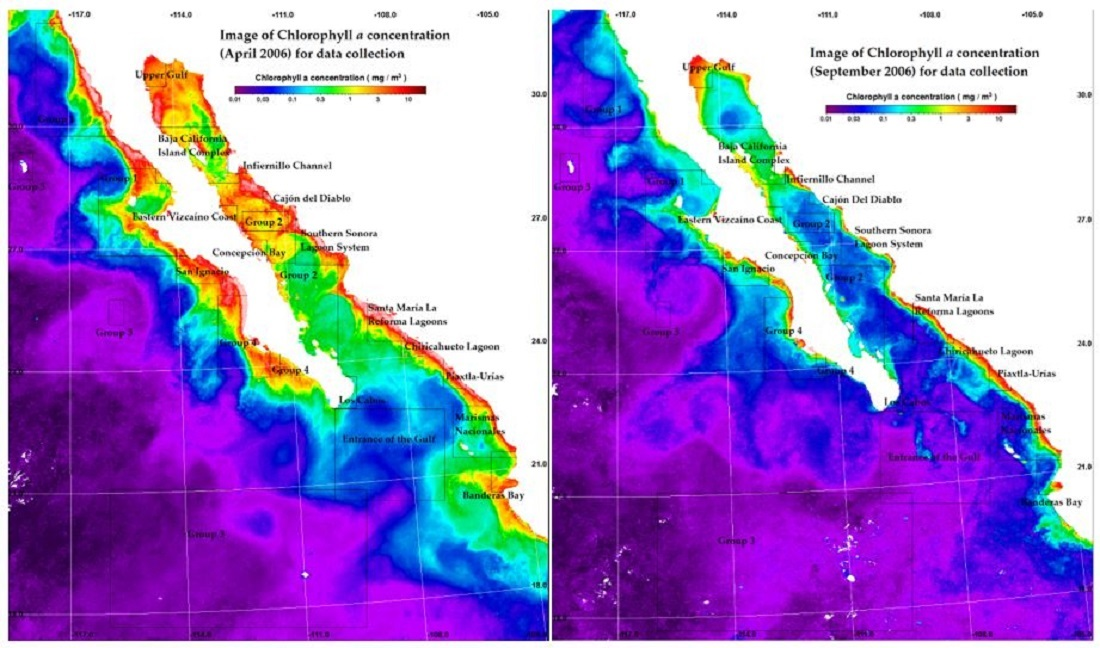
2.2. Oceanographic Characterization and Monthly Chlorophyll a Data
2.3. Processing and Statistical Analyses of Monthly Chlorophyll a Data
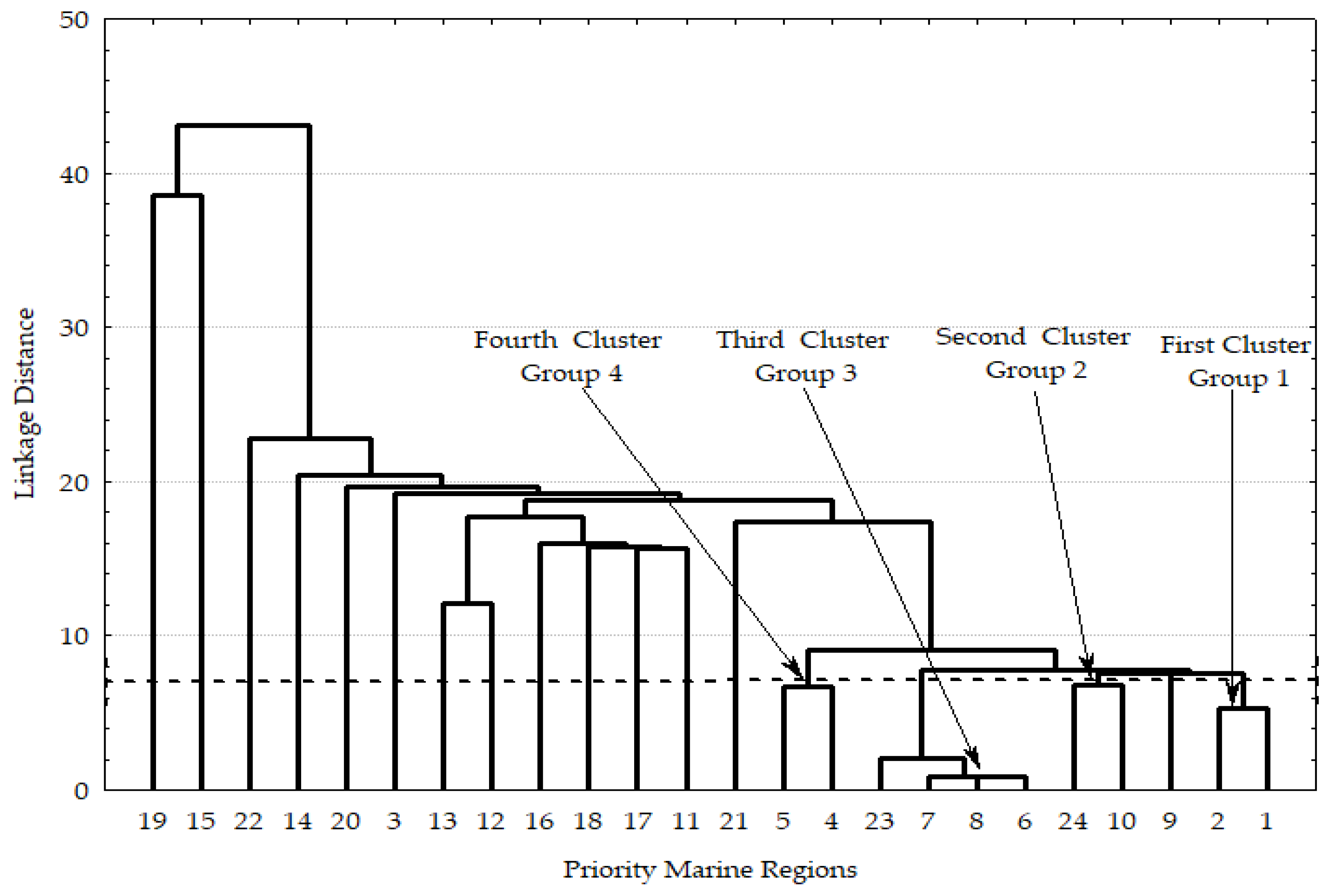
3. Results
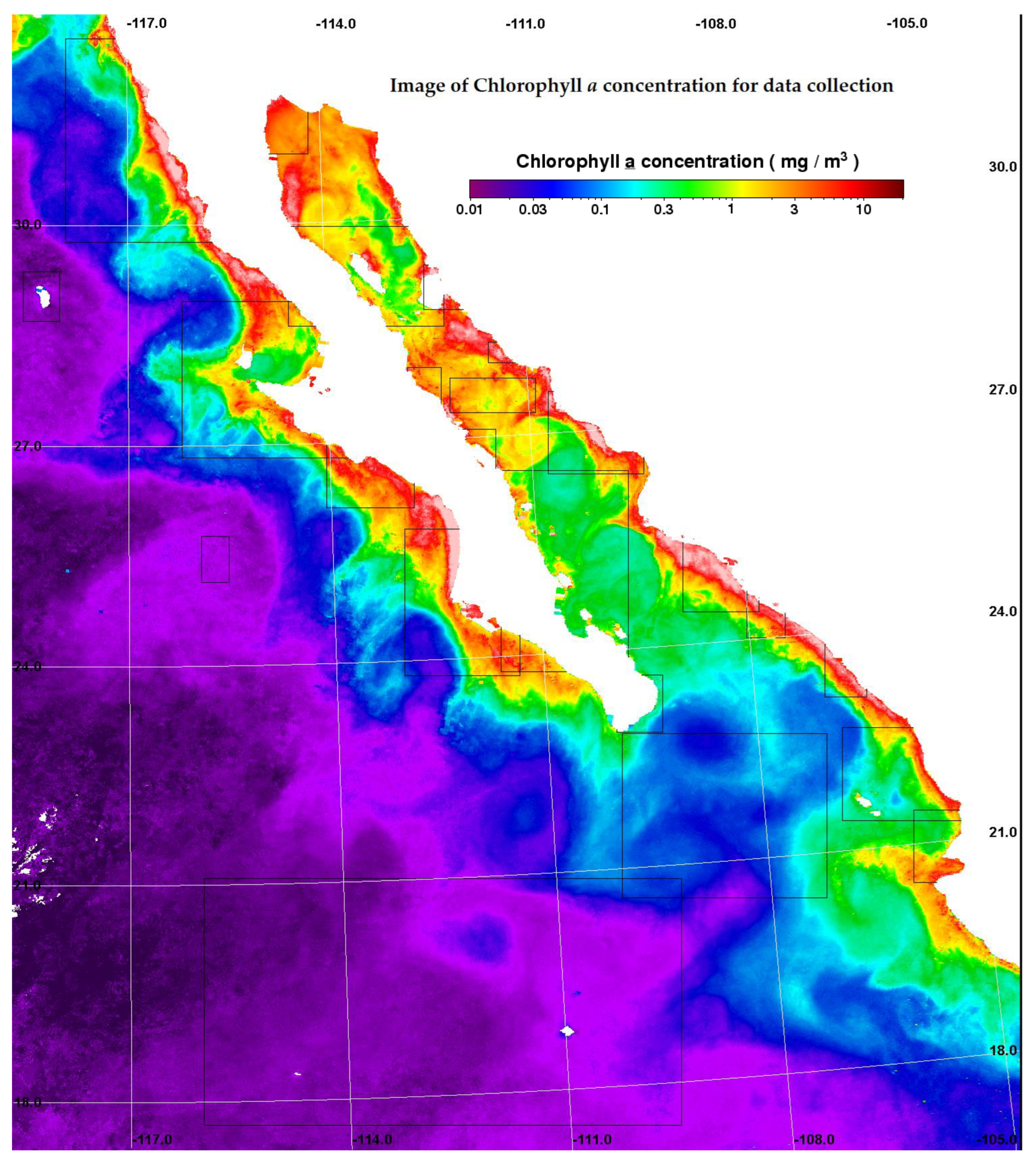
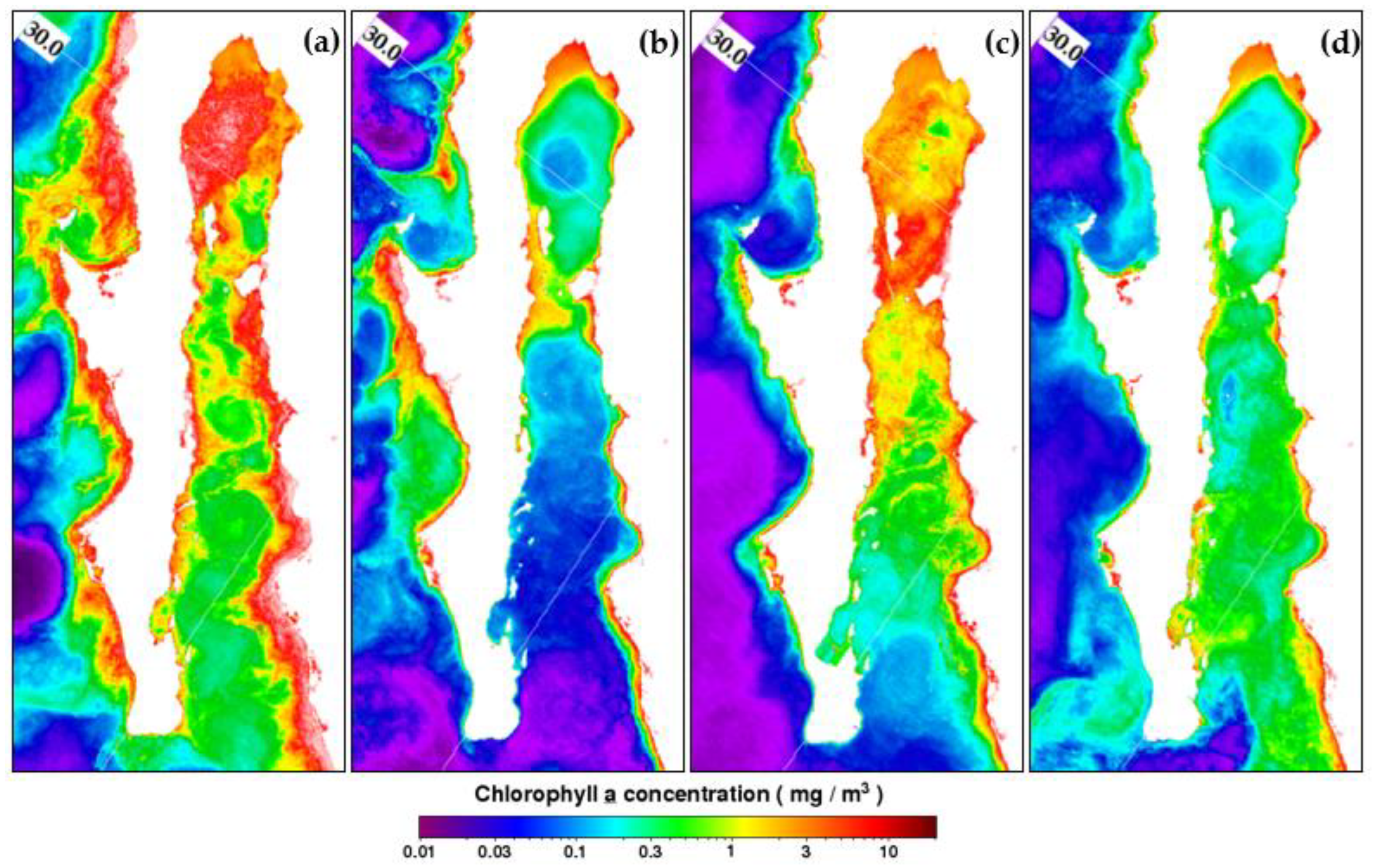
3.1. Regional Characterization
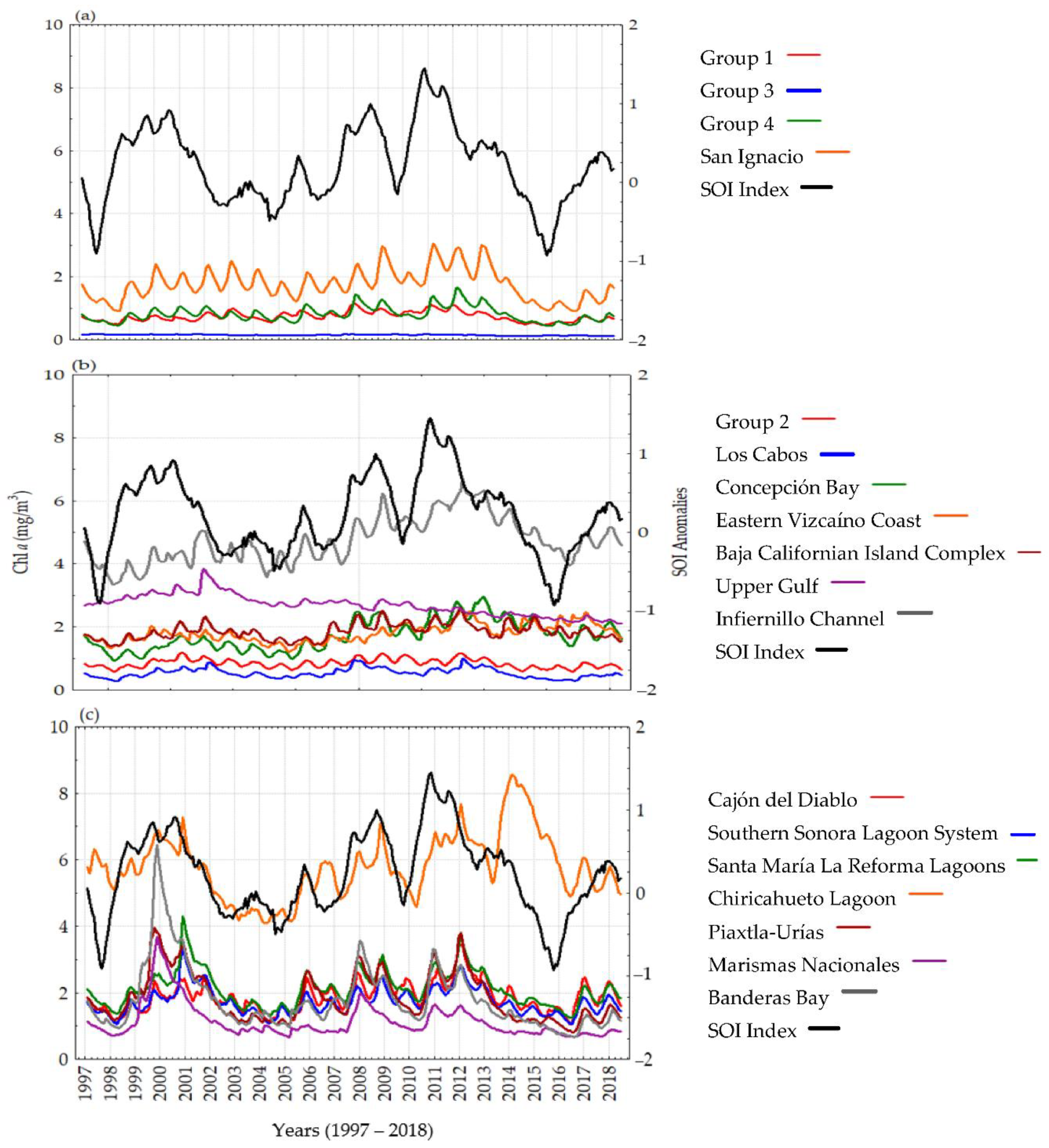
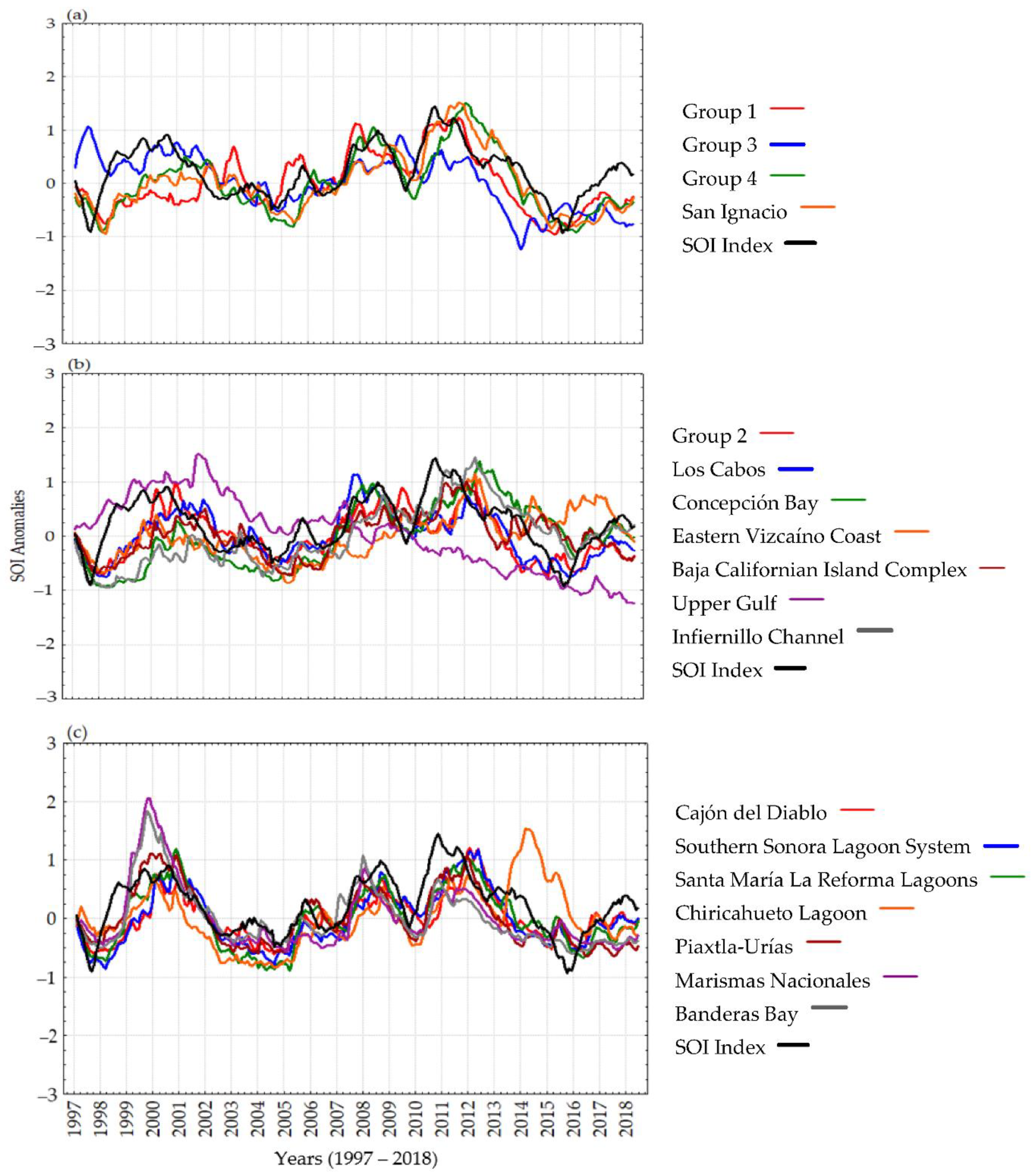
3.2. Time Series Analyses and Anomalies
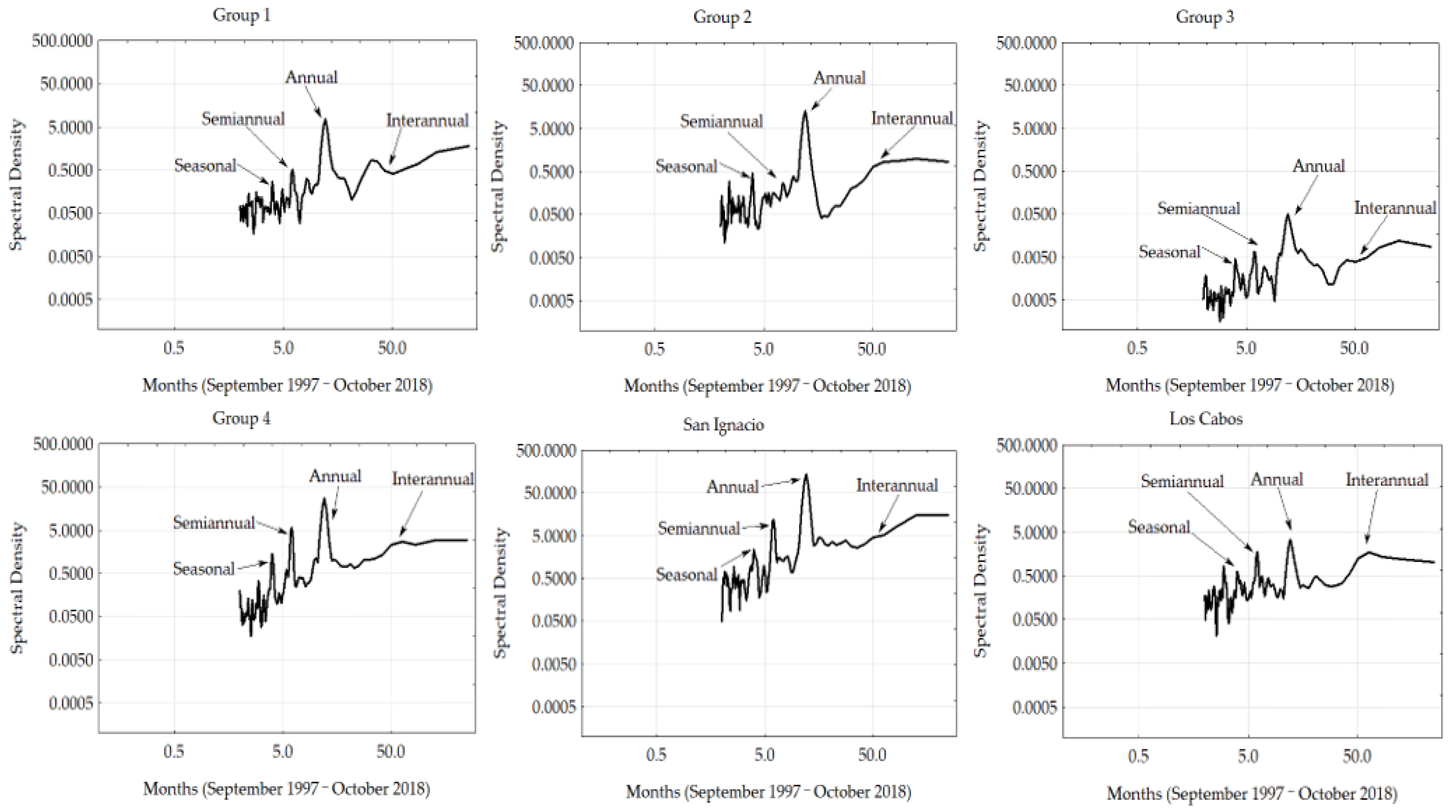
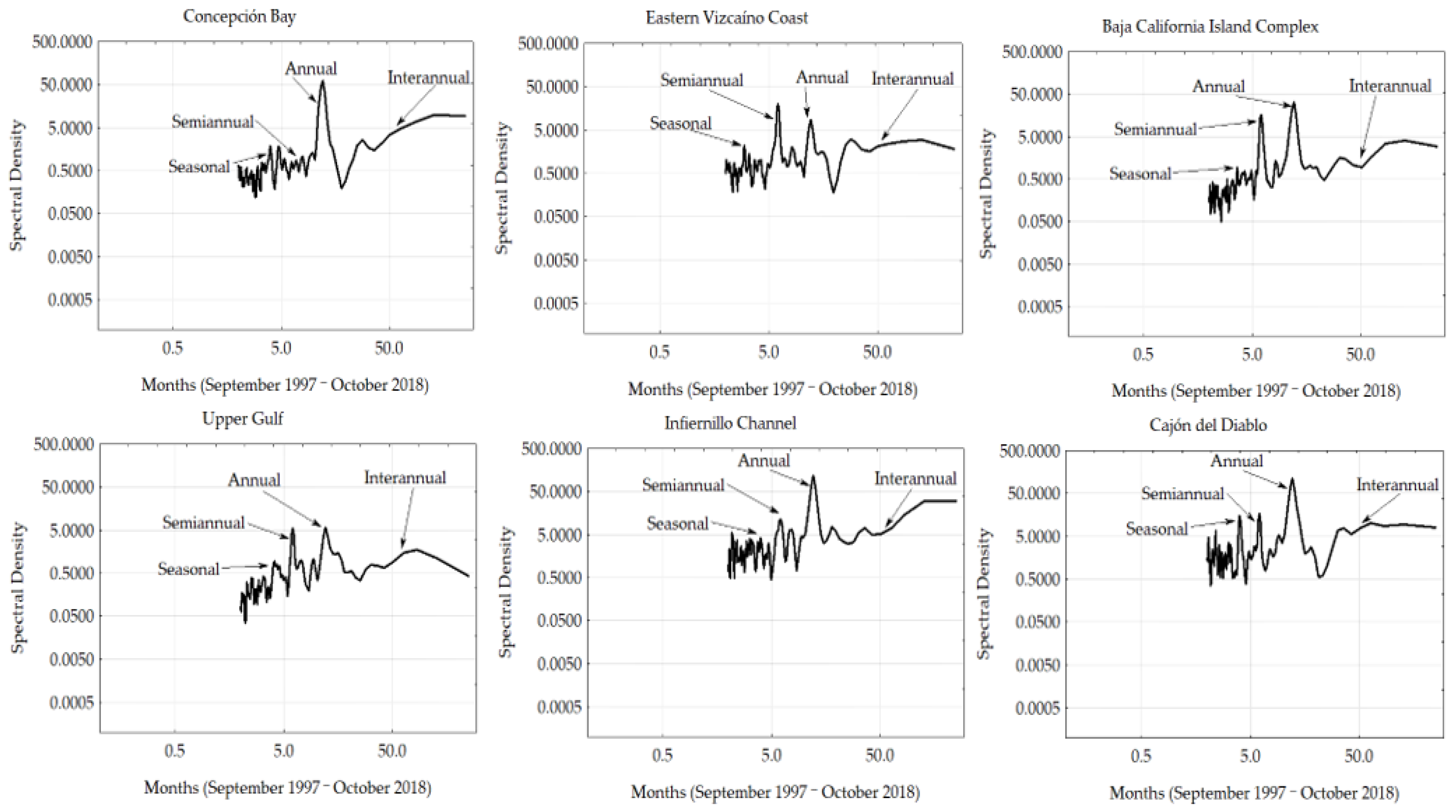
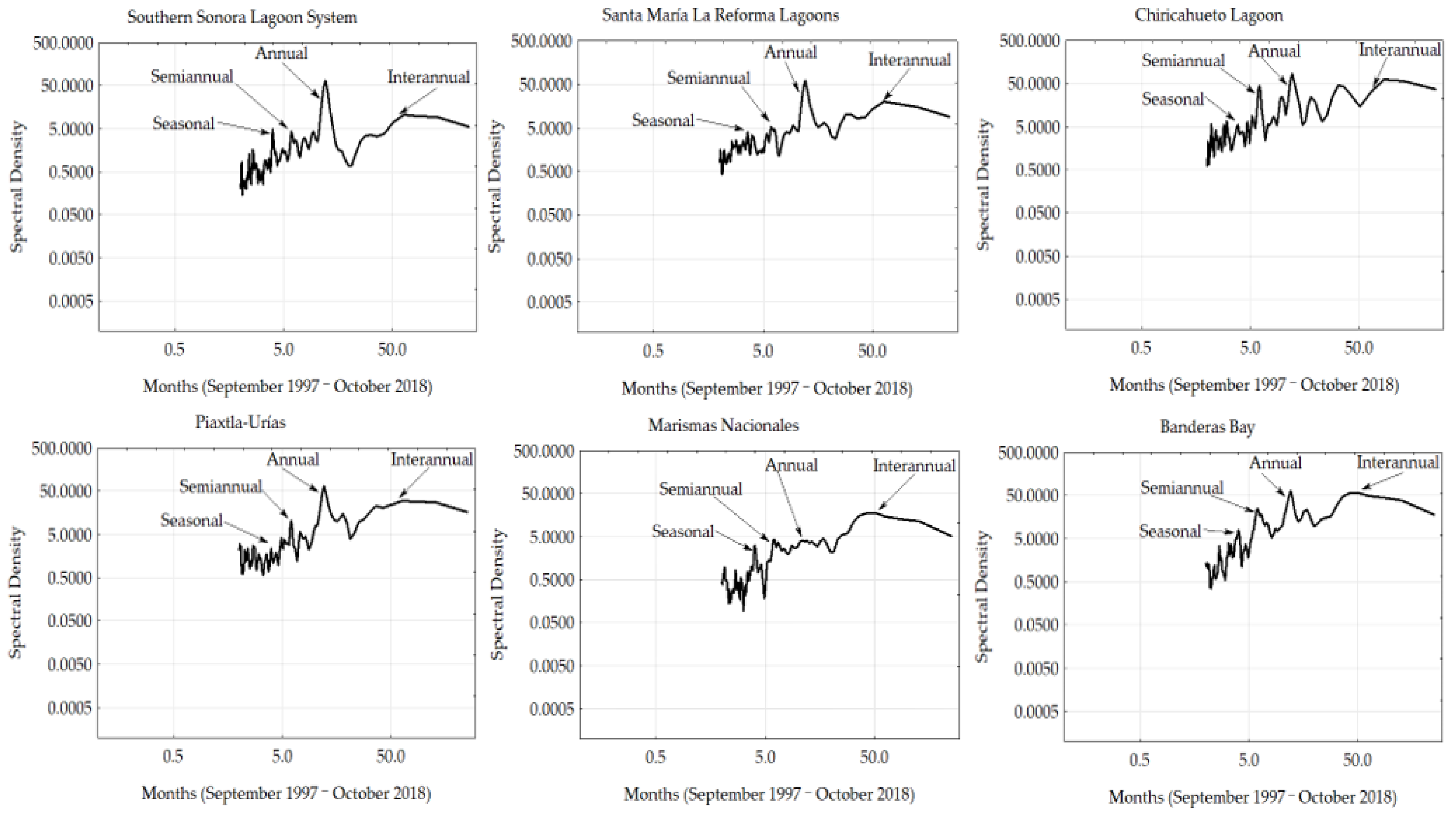
3.3. Fourier Analyses
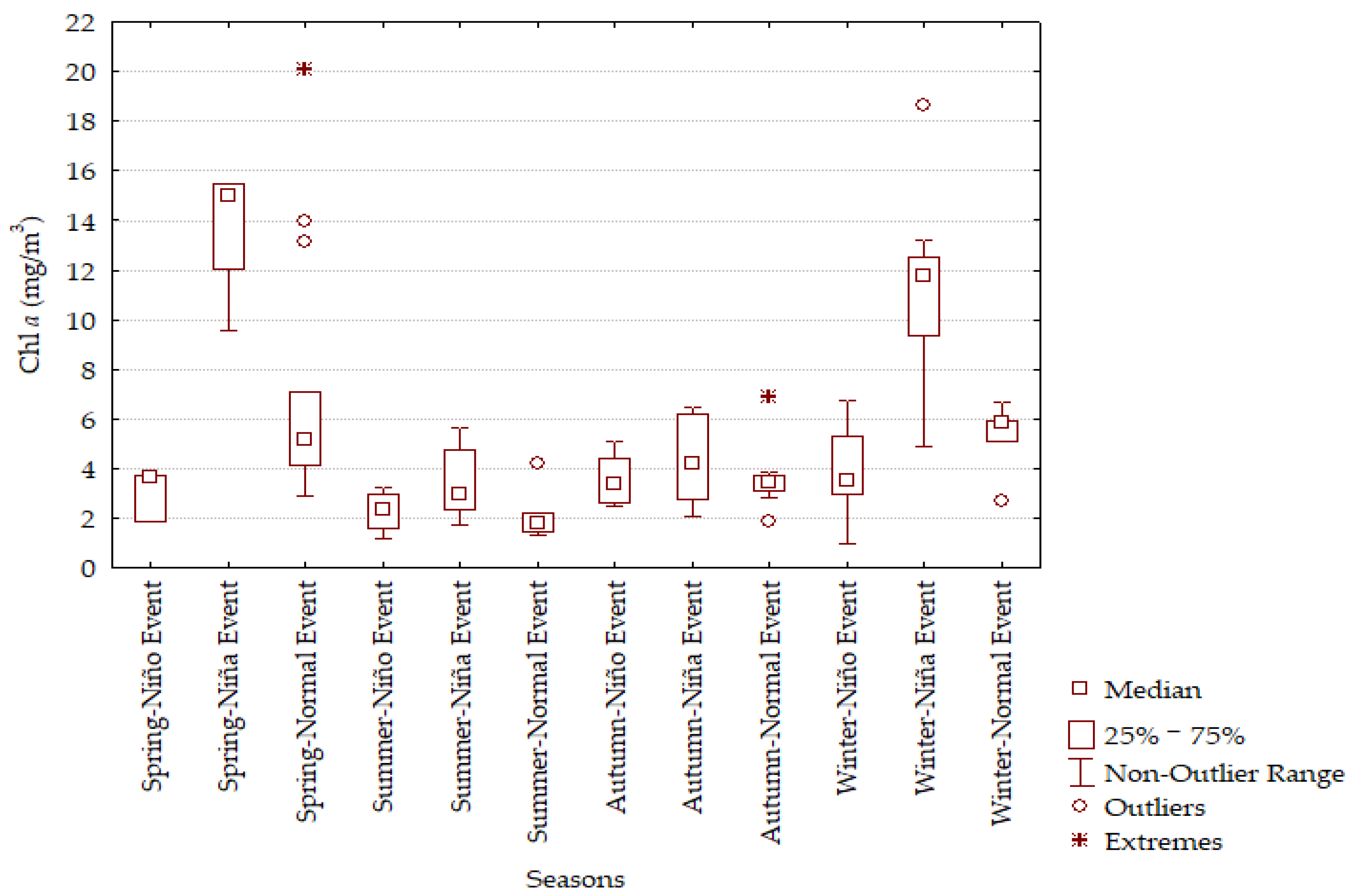
3.4. Statistical Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lara-Lara, J.R.; Arenas-Fuentes, V.; Bazán-Guzmán, C.; Díaz-Castañeda, V.; Escobar-Briones, E.; García-Abad, M.; Espe-jel-Carvajal, M.I.; Guzmán-Arroyo, M.; Ladah, L.B.; López-Hernández, M.; et al. Los Ecosistemas Marinos. In Capital Natural de México. Vol. I: Conocimiento Actual de la Biodiversidad, 1st ed.; Soberón, J., Halffter, G., y Llorente, J., Eds.; Comisión Nacional para el Conoci-miento y Uso de la Biodiversidad, CONABIO: Ciudad de México, México, 2008; Volume 1, pp. 135–159. [Google Scholar]
- Rosales-Nanduca, H.; Gerrodette, T.; Urbán-R, J.; Cárdenas-Hinojosa, G.; Medrano-González, L. Macroecology of marine mammal species in the Mexican Pacific Ocean: Diversity and distribution. Mar. Ecol. Prog. Ser. 2011, 431, 281–291. [Google Scholar] [CrossRef]
- Saldaña-Ruiz, L.E.; García-Rodríguez, E.; Pérez-Jiménez, J.; Tovar-Ávila, J.; Rivera-Téllez, E. Chapter Two—Biodiversity and Conservation of Sharks in Pacific Mexico. In Advances in Marine Biology, 1st ed.; Larson, S.E., Lowry, D., Eds.; The University of Warwick: Coventry, UK, 2019; Volume 83, pp. 11–60. [Google Scholar]
- Lluch-Cota, S.E.; Aragón-Noriega, E.A.; Arreguín-Sánchez, F.; Aurioles-Gamboa, D.; Bautista-Romero, J.J.; Brusca, R.C.; Cervantes-Duarte, R.; Cortés-Altamirano, R.; Del-Monte-Luna, P.; Esquivel-Herrera, A.; et al. The Gulf of California: Review of ecosystems status and sustainability challenges. Pro. Oce. 2007, 73, 1–26. [Google Scholar] [CrossRef]
- Arreguín-Sánchez, F.; del Monte-Luna, P.; Zetina-Rejón, M.J.; Albáñez-Lucero, M.O. The Gulf of California Large Marine Ecosystem: Fisheries and other natural resources. Environ. Dev. 2017, 22, 71–77. [Google Scholar] [CrossRef]
- Espinosa, H. El Pacífico Mexicano. Ciencias 2004, 76, 14–21. [Google Scholar]
- Lluch-Cota, S.E.; Parés-Sierra, A.; Magaña-Rueda, V.O.; Arreguín-Sánchez, F.; Bazzino, G.; Herrera-Cervantes, H.; Lluch-Belda, D. Changing climate in the Gulf of California. Prog. Oceanogr. 2010, 87, 114–126. [Google Scholar] [CrossRef]
- Arriaga-Cabrera, L.; Aguilar, A.; Espinoza, J.M. Regiones Marinas y Planeación Para la Conservación de la Biodiversidad. In Capital Natural de México. Vol. II: Estado de Conservación y Tendencias de Cambio, 1st ed.; Dirzo, R., González, R., March, I.J., Eds.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, CONABIO: Ciudad de México, México, 2009; Volume 2, pp. 433–457. [Google Scholar]
- Wilkinson, T.A.C.; Wilken, E.; Bezaury-Creel, J.; Hourigan, T.F.; Agardy, T.; Herrmann, H.; Janishevski, L.; Madden, C.; Morgan, L.; Padilla, M. Ecorregiones Marinas de América del Norte; Comisión para la Cooperación Ambiental: Montreal, QC, Canada, 2009; pp. 107–108. [Google Scholar]
- Gaxiola-Castro, G.; Cepeda-Morales, J.C.A.; Nájera-Martínez, S.; Espinosa-Carreón, T.L.; De la Cruz-Orozco, M.E.; So-sa-Avalos, R.; Aguirre-Hernández, E.; Cantú-Ontiveros, J.P. Biomasa y Producción de Fitoplancton. In Dinámica del Ecosistema Pelágico Frente a Baja California, 1997–2007. In Diez Años de Investigaciones Mexicanas de la Corriente de California, 1st ed.; Gaxiola-Castro, G., Durazo-Arvizu, R., Eds.; Secretaría del Medio Ambiente y Recursos Naturales, Instituto Nacional de Ecología, Centro de Investigación Científica y de Educación Superior de Ensenada, Universidad Autónoma de Baja California: Ensenada, México, 2010; pp. 59–85. [Google Scholar]
- Valiela, I. Marine Ecological Processes, 2nd ed.; Springer: New York, NY, USA, 1995; p. 645. [Google Scholar]
- Ramírez, D.G.; Giraldo, A.; Tovar, J. Primary production, biomass, and taxonomic composition of coastal and oceanic phy-toplankton in the Colombian Pacific (September–October 2004). Lat. Ame. Aqua. Res. 2006, 34, 211–216. [Google Scholar]
- Morales-Hernández, J.C.; Carrillo-González, F.M.; Farfán-Molina, L.M.; Cornejo-López, V.M. Vegetation change cover in the coastal region of Bahia de Banderas. Mexico. Cal. 2016, 38, 17–29. [Google Scholar]
- Rohli, R.V.; Vega, A. Climatology, 4th ed.; Jones & Bartlett Learning: Burlington, MA, USA, 2018; p. 60. [Google Scholar]
- Soto-Mardones, L.; Marinone, S.; Parés-Sierra, A. Time and spatial variability of sea surface temperature in the Gulf of California. Cienc. Mar. 1999, 25, 1–30. [Google Scholar] [CrossRef]
- Henson, S.A.; Sarmiento, J.L.; Dunne, J.P.; Bopp, L.; Lima, I.; Doney, S.C.; John, J.; Beaulieu, C. Detection of anthropogenic climate change in satellite records of ocean chlorophyll and productivity. Biogeosciences 2010, 7, 621–640. [Google Scholar] [CrossRef]
- Gregg, W.W.; Rousseaux, C.S. Decadal trends in global pelagic ocean chlorophyll: A new assessment integrating multiple satellites, in situ data, and models. J. Geophys. Res. Oceans 2014, 119, 5921–5933. [Google Scholar] [CrossRef]
- Nayak, R.K.; Mishra, S.K.; Satyesh, G.; Nagamani, P.V.; Choudhury, S.B.; Seshasai, M.V.R. Remote sensing application in satellite oceanography. Ind. Geo. J. 2018, 93, 156–165. [Google Scholar]
- Longhurst, A.R. Ecological Geography of the Sea, 2nd ed.; Elsevier Academic Press: San Diego, CA, USA, 2007; p. 5. [Google Scholar]
- Devi, G.K.; Ganasri, B.; Dwarakish, G. Applications of Remote Sensing in Satellite Oceanography: A Review. Aquat. Procedia 2015, 4, 579–584. [Google Scholar] [CrossRef]
- Gregg, W.W.; Conkright, M.E. Global seasonal climatologies of ocean chlorophyll: Blending in situ and satellite data for the Coastal Zone Color Scanner era. J. Geophys. Res. Oceans. 2001, 106, 2499–2515. [Google Scholar] [CrossRef]
- Dutkiewicz, S.; Hickman, A.E.; Jahn, O.; Henson, S.; Beaulieu, C.; Monier, E. Ocean Color Signature of Climate Change. Nat. Com. 2019, 10, 1–13. [Google Scholar]
- Krug, L.A.; Platt, T.; Sathyendranath, S.; Barbosa, A.B. Ocean surface partitioning strategies using ocean colour remote Sensing: A review. Prog. Oceanogr. 2017, 155, 41–53. [Google Scholar] [CrossRef]
- Schwarz, J.N. Dynamic partitioning of tropical Indian Ocean surface waters using ocean colour data—Management and modelling applications. J. Environ. Manag. 2020, 276, 111308. [Google Scholar] [CrossRef]
- Krug, L.A.; Platt, T.; Sathyendranath, S.; Barbosa, A.B. Patterns and drivers of phytoplankton phenology off SW Iberia: A phenoregion based perspective. Prog. Oceanogr. 2018, 165, 233–256. [Google Scholar] [CrossRef]
- Heras-Sánchez, M.C.; Valdez-Holguín, J.E.; Garatuza-Payán, J.; Cisneros-Mata, M.A.; Díaz-Tenorio, L.M.; Robles-Morua, A.; Ha-zas-Izquierdo, R.G. Regiones del Golfo de California determinadas por la distribución de temperatura superficial del mar y la clorofila-a. Biotecnia 2019, 21, 13–21. [Google Scholar] [CrossRef]
- Santamaría-del-Angel, E.; González-Silvera, A.; Millán-Núñez, R.; Callejas-Jiménez, M.E.; Cajal-Medrano, R. Determining Dynamic Biogeographic Regions Using Remote Sensing Data. In Handbook of Satellite Remote Sensing Image Interpretation: Applications for Marine Living Resources Conservation and Management, 1st ed.; Morales, J., Stuart, V., Platt, T., Sathyendranath, S., Eds.; EU PRESPO and IOCCG: Dartmouth, NS, Canada, 2011; Volume 1, pp. 273–293. [Google Scholar]
- Millán-Núñez, R.; Alvarez-Borrego, S.; Trees, C.C. Modeling the vertical distribution of chlorophyll in the California Current System. J. Geo. Res. 1997, 102, 8587–8595. [Google Scholar] [CrossRef]
- Thomas, A.C.; Mendelssohn, R.; Weatherbee, R. Background trends in California Current surface chlorophyll concentrations: A state-space view. J. Geophys. Res. Oceans 2013, 118, 5296–5311. [Google Scholar] [CrossRef]
- Hernández-de la Torre, B.; Aguirre-Gómez, R.; Gaxiola-Castro, G.; Álvarez-Borrego, S.; Gallegos-García, A.; Rosete-Vergés, F.; Bocco-Verdinelli, G. Ordenamiento Ecológico Marino en el Pacífico Norte mexicano: Propuesta metodológica. Hidrobiológica 2015, 25, 151–163. [Google Scholar]
- Santamaría-Del-Angel, E.; Alvarez-Borrego, S.; Muller-Karger, F.E. Gulf of California biogeographic regions based on coastal zone color scanner imagery. J. Geophys. Res. Earth Surf. 1994, 99, 7411–7421. [Google Scholar] [CrossRef]
- Kahru, M.; Marinone, S.; Lluch-Cota, S.; Parés-Sierra, A.; Mitchell, B.G. Ocean-color variability in the Gulf of California: Scales from days to ENSO. Deep-Sea Res. II 2004, 51, 139–146. [Google Scholar] [CrossRef]
- García-Morales, R.; López-Martínez, J.; Valdez-Holguin, J.E.; Herrera-Cervantes, H.; Espinosa-Chaurand, L.D. Environmental Variability and Oceanographic Dynamics of the Central and Southern Coastal Zone of Sonora in the Gulf of California. Remote Sens. 2017, 9, 925. [Google Scholar] [CrossRef]
- García-Morales, R.; Pérez-Lezama, E.L.; Shirasago-Germán, B. Influence of environmental variability of baleen whales (suborden mysticeti) in the Gulf of California. Mar. Eco. 2017, 38, 10. [Google Scholar]
- Gaxiola-Castro, G.; Durazo, R. Introducción. In Dinámica del Ecosistema Pelágico Frente a Baja California, 1997–2007. Diez años de Investigaciones Mexicanas de la Corriente de California, 1st ed.; Gaxiola-Castro, G., Durazo-Arvizu, R., Eds.; Secretaría del Medio Ambiente y Recursos Naturales, Instituto Nacional de Ecología, Centro de Investigación Científica y de Educación Superior de Ensenada, Universidad Autónoma de Baja California: Ensenada, México, 2010; pp. 13–24. [Google Scholar]
- Espinosa-Carreón, T.L.; Gaxiola-Castro, G.; Beier, E.; Strub, P.T.; Kurczyn, J.A. Effects of mesoscale processes on phytoplankton chlorophyll off Baja California. J. Geo. Res. Oce. 2012, 117, C04005. [Google Scholar] [CrossRef]
- Alvarez-Borrego, S. Physical, Chemical and Biological Oceanography of the Gulf of California. In The Gulf of California: Biodiversity and Conservation, 2nd ed.; Brusca, R., Ed.; The University of Arizona Press: Tucson, AZ, USA, 2010; pp. 24–48. [Google Scholar]
- Alvarez-Borrego, S.; Lara-Lara, J.R. The Physical Environment and Primary Productivity of the Gulf of California. In The Gulf of California, Province of the Californias, 1st ed.; Dauphin, J.P., Simoneit, B.R.T., Eds.; American Association of Petroleum Geologist: Tulsa, OK, USA, 1991; pp. 555–567. [Google Scholar]
- Alvarez-Borrego, S. Phytoplankton biomass and production in the Gulf of California: A review. Bot. Mar. 2012, 55, 119–128. [Google Scholar] [CrossRef]
- Kahru, M. The California Current Merged Satellite-Derived 4-km Dataset. 2020. Available online: http://www.wimsoft.com/CC4km.htm (accessed on 29 September 2020).
- Kahru, M.; Kudela, R.M.; Manzano-Sarabia, M.; Mitchell, B.G. Trends in the surface chlorophyll of the California Current: Merging data from multiple ocean color satellites. Deep Sea Res. Part II Top. Stud. Oceanogr. 2012, 77–80, 89–98. [Google Scholar] [CrossRef]
- Kahru, M. Windows Image Manager, WIM Software (Ver. 9.06) and User’s Manual. 2016. Available online: http://www.wimsoft.com/ (accessed on 29 September 2020).
- Everitt, B.S.; Landau, S.; Leese, M.; Stahl, D. Cluster Analysis, 5th ed.; John Wiley & Sons Ltd.: Chichester, UK, 2011; p. 13. [Google Scholar]
- Kahru, M.; Mitchell, G. Influence of the 1997–1998 El Niño on surface chlorophyll in the California Current. Geo. Res. Let. 2000, 27, 2937–2940. [Google Scholar] [CrossRef]
- Kahru, M.; Mitchell, G. Seasonal and nonseasonal variability of satellite-derived chlorophyll and colored dissolved organic matter concentration in the California Current. J. Geo. Res. 2001, 102, 2517–2529. [Google Scholar] [CrossRef]
- García-Morales, R. Variabilidad Oceanográfica del Hábitat de Los Stocks de Sardinops Sagax (Jenyns, 1842) (Clupeiformes: Clupeidae) en el Sistema de Corriente de California (1981–2005). Ph.D. Thesis, Centro Interdisciplinario de Ciencias Mari-nas-Instituto Politécnico Nacional, La Paz, Baja California Sur, México, 2012. [Google Scholar]
- Zaitsev, O.; Cervantes-Duarte, R.; Montante, O.; Gallegos-Garcia, A. Coastal Upwelling Activity on the Pacific Shelf of the Baja California Peninsula. J. Oceanogr. 2003, 59, 489–502. [Google Scholar] [CrossRef]
- Pérez-Brunius, P.; López, M.; Páres-Sierra, A.; Pineda, J. Comparison of upwelling indices off Baja California derived from three different wind data sources. CalCOFI Rep. 2007, 48, 204–212. [Google Scholar]
- Durazo, R.; Ramírez-Manguilar, A.M.; Miranda, L.E.; Soto-Mardones, L.A. Climatología de variables oceanográficas. In Di-námica del Ecosistema Pelágico Frente a Baja California, 1997–2007. Diez años de Investigaciones Mexicanas de la Corriente de California, 1st ed.; Gaxiola-Castro, G., Durazo-Arvizu, R., Eds.; Secretaría del Medio Ambiente y Recursos Naturales, Instituto Nacional de Ecología, Centro de Investigación Científica y de Educación Superior de Ensenada, Universidad Autónoma de Baja California: Ensenada, México, 2010; pp. 25–57. [Google Scholar]
- Durazo, R. Climate and upper ocean variability off Baja California, Mexico: 1997–2008. Prog. Oceanogr. 2009, 83, 361–368. [Google Scholar] [CrossRef]
- Durazo, R. Seasonality of the transitional region of the California Current System off Baja California. J. Geophys. Res. Oceans 2015, 120, 1173–1196. [Google Scholar] [CrossRef]
- Castro, R.; Martínez, J.A. Variabilidad espacial y temporal del campo del viento. In Dinámica del Ecosistema Pelágico Frente a Baja California, 1997–2007. Diez Años de Investigaciones Mexicanas de la Corriente de California, 1st ed.; Gaxiola-Castro, G., Durazo-Arvizu, R., Eds.; Secretaría del Medio Ambiente y Recursos Naturales, Instituto Nacional de Ecología, Centro de Investigación Científica y de Educación Superior de Ensenada, Universidad Autónoma de Baja California: Ensenada, México, 2010; pp. 129–147. [Google Scholar]
- Durazo, R.; Baumgartner, T. Evolution of oceanographic conditions off Baja California: 1997–1999. Prog. Oceanogr. 2002, 54, 7–31. [Google Scholar] [CrossRef]
- Round, F. The Gulf of California. Part I. Its composition, distribution and contribution to the sediments. J. Exp. Mar. Biol. Ecol. 1967, 1, 76–97. [Google Scholar] [CrossRef]
- Escalante, F.; Valdez-Holguín, J.E.; Álvarez-Borrego, S.; Lara-Lara, J.R. Temporal and spatial variation of sea surface temperature, chlorophyll a, and primary productivity in the Gulf of California. Cie. Mar. 2013, 39, 203–215. [Google Scholar] [CrossRef]
- Alvarez-Borrego, S. Gulf of California. In Ecosystems of the World, 26: Estuaries and Enclosed Seas, 1st ed.; Ketchum, B.H., Ed.; Elsevier Scientific: New York, NY, USA, 1983; pp. 427–429. [Google Scholar]
- Torres-Orozco, E. Análisis Volumétrico de las Masas de Agua del Golfo de California. Master’s Thesis, Centro de Investigación Científica y de Educación Superior de Ensenada, Ensenada, Baja California, México, 1993. [Google Scholar]
- Santamaría-del-Angel, E.; Álvarez-Borrego, S.; Millán-Núñez, R.; Muller-Karger, F.E. Sobre el efecto de las surgencias de verano en la biomasa fitoplanctónica del Golfo de California. Rev. Soc. Mex. Hist. Nat. 1999, 49, 207–212. [Google Scholar]
- Espinosa-Carreón, T.L.; Valdez-Holguín, J.E. Variabilidad interanual de clorofila en el Golfo de California. Eco. Apl. 2007, 6, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Lavín, M.F.; Beier, E.; Badan, A. Estructura hidrográfica y circulación del Golfo de California: Escalas estacional e interanual. In Contribuciones a la Oceanografía Física en México, 1st ed.; Lavín, M.F., Ed.; Unión Geofísica Mexicana: Ensenada, Baja California, México, 1997; pp. 141–172. [Google Scholar]
- Lavín, M.F.; Marinone, S.G. An overview of the physical Oceanography of the Gulf of California. In Nonlinear Processes in Geophysical Fluid Dynamics, 1st ed.; Velasco-Fuentes, O.I., Sheinbaum, J., Ochoa- de la Torre, J.L., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 173–204. [Google Scholar]
- Stevenson, M.R. On the physical and biological oceanography near to the entrance of the Gulf of California, October 1966–August 1967. Bul. Int. Tro. Tun. Com. 1970, 14, 389–504. [Google Scholar]
- Millán-Nuñez, R.; Santamaría-Del-Ángel, E.; Cajal-Medrano, R.; Barocio-León, O. The Colorado River Delta: A high primary productivity ecosystem. Cienc. Mar. 1999, 25, 509–524. [Google Scholar] [CrossRef]
- Ramírez-León, M.R. Biomasa y Producción de Fitoplancton en el Norte del Golfo de California. Mater’s Thesis, Centro de Investigación Científica y de Educación Superior de Ensenada, Ensenada, Baja California, Mexico, 2014. [Google Scholar]
- Pérez Arvizu, E.M.; Aragón-Noriega, E.A.; Espinosa-Carreón, T.L. Seasonal variability of chlorophyll a and their response to El Niño and La Niña conditions in the Northern Gulf of California. Rev. Bio. Mar. Oce. 2013, 48, 131–141. [Google Scholar]
- Ramírez-León, M.R.; Álvarez-Borrego, S.; Turrent-Thompson, C.; Gaxiola-Castro, G.; Dziendzielewski, G.H. Nutrient input from the Colorado River to the northern Gulf of California is not required to maintain a productive pelagic ecosystem. Cienc. Mar. 2015, 41, 169–188. [Google Scholar] [CrossRef]
- Brusca, R.C.; Álvarez-Borrego, S.; Hastings, P.A.; Findley, L.T. Colorado River flow and biological productivity in the Northern Gulf of California, Mexico. Earth-Sci. Rev. 2017, 164, 1–30. [Google Scholar] [CrossRef]
- Robles-Tamayo, C.M.; García-Morales, R.; Valdez-Holguín, J.E.; Figueroa-Preciado, G.; Herrera-Cervantes, H.; Ló-pez-Martínez, J.; Enríquez-Ocaña, L.F. Chlorophyll a Concentration Distribution on the Mainland Coast of the Gulf of California. Rem. Sen. 2020, 12, 1335. [Google Scholar] [CrossRef]
- López, M.; Candela, J.; Argote, M.L. Why does the Ballenas Channel have the coldest SST in the Gulf of California? Geophys. Res. Let. 2006, 33, L11603. [Google Scholar] [CrossRef]
- Álvarez-Molina, L.L.; Álvarez-Borrego, S.; Lara-Lara, J.R.; Marinone, S. Annual and semiannual variations of phytoplankton biomass and production in the central Gulf of California estimated from satellite data. Cienc. Mar. 2013, 39, 217–230. [Google Scholar] [CrossRef]
- Lancin, M. Geomorfología y génesis de las fuerzas litorales del Canal del Infiernillo, Estado de Sonora. Rev. Mex. Cie. Geo. 1985, 6, 57–72. [Google Scholar]
- Alonso-Rodríguez, R.; Páez-Osuna, F. Nutrients, phytoplankton and harmful algal blooms in shrimp ponds: A review with special reference to the situation in the Gulf of California. Aquaculture 2003, 219, 317–336. [Google Scholar] [CrossRef]
- Beman, J.M.; Arrigo, K.R.; Matson, P.A. Agricultural runoff fuels large phytoplankton blooms in vulnerable areas of the ocean. Nature 2005, 434, 211–214. [Google Scholar] [CrossRef]
- Valenzuela-Sanchez, C.G.; Pasten-Miranda, N.M.; Enriquez-Ocaña, L.F.; Barraza-Guardado, R.H.; Holguin, J.V.; Martinez-Cordova, L.R. Phytoplankton composition and abundance as indicators of aquaculture effluents impact in coastal environments of mid Gulf of California. Heliyon 2021, 7, e06203. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.; Voltolina, D.; Frías-Espericueta, M.G.; Izaguirre-Fierro, G.; Rivas-Vega, M.E. Budget and discharges of nutrients to the Gulf of California of a semi-intensive shrimp farm (NW Mexico). Hidrobiológica 2009, 19, 43–48. [Google Scholar]
- Martínez-López, A.; Escobedo-Urías, D.; Reyes-Salinas, A.; Hernández-Real, M.T. Phytoplankton response to nutrient runoff in a large lagoon system in the Gulf of California. Hidrobiológica 2007, 17, 101–112. [Google Scholar]
- Martínez-Fuentes, L.M.; Gaxiola-Castro, G.; Gómez-Ocampo, E.; Kahru, M. Effects of interannual events (1997–2012) on the hydrography and phytoplankton biomass of Sebastian Vizcaíno Bay. Cie. Mar. 2016, 42, 81–97. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Ocampo, E.; Gaxiola-Castro, G.; Durazo, R.; Beier, E. Effects of the 2013-2016 warm anomalies on the California Current phytoplankton. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 151, 64–76. [Google Scholar] [CrossRef]
- Ortíz-Ahumada, J.C.; Álvarez-Borrego, S.; Gómez-Valdés, J. Effects of seasonal and interannual events on satellite-derived phytoplankton biomass and production in the southernmost part of the California Current System during 2003–2016. Cie. Mar. 2018, 44, 1–20. [Google Scholar] [CrossRef]
- González-Silvera, A.; Santamaría-Del-Ángel, E.; Camacho-Ibar, V.; López-Calderón, J.; Santander-Cruz, J.; Mercado-Santana, A. The Effect of Cold and Warm Anomalies on Phytoplankton Pigment Composition in Waters off the Northern Baja California Peninsula (México): 2007–2016. J. Mar. Sci. Eng. 2020, 8, 533. [Google Scholar] [CrossRef]
- Herrera-Cervantes, H.; Lluch-Cota, S.E.; Lluch-Cota, D.B.; Gutiérrez-De-Velasco, G. Interannual correlations between sea surface temperature and concentration of chlorophyll pigment off Punta Eugenia, Baja California, during different remote forcing conditions. Ocean Sci. 2014, 10, 345–355. [Google Scholar] [CrossRef]
- Espinosa-Carreón, T.; Gaxiola-Castro, G.; Durazo, R.; De la Cruz-Orozco, M.; Norzagaray-Campos, M.; Solana-Arellano, E. Influence of anomalous subarctic water intrusion on phytoplankton production off Baja California. Cont. Shelf Res. 2015, 92, 108–121. [Google Scholar] [CrossRef]
- Herrera-Cervantes, H.; Lluch-Cota, S.E.; Cortés-Ramos, J.; Farfán, L.; Morales-Aspeitia, R. Interannual variability of surface satellite-derived chlorophyll concentration in the bay of La Paz, Mexico, during 2003–2018 period: The ENSO signature. Cont. Shelf Res. 2020, 209, 104254. [Google Scholar] [CrossRef]
- Valdéz-Holguín, J.; Lara-Lara, J. Primary Productivity in the Gulf of California Effects of El Niño 1982–1983 Event. Cienc. Mar. 1987, 13, 34–50. [Google Scholar] [CrossRef]
- Herrera-Cervantes, H.; Lluch-Cota, S.E.; Lluch-Belda, D.B.; Gutiérrez de Velasco-Sanromán, G.; Lluch-Belda, D. ENSO influence on satellite-derive Chlorophyll trends in the Gulf of California. Atmósfera 2010, 23, 253–262. [Google Scholar]
- Coria Monter, E.; Monreal-Gómez, M.A.; Salas-de León, D.A.; Durán-Campos, E. Impact of the “Godzilla Niño” Event of 2015–2016 on the Sea-Surface Temperature and Chlorophyll-a in the Southern Gulf of California, Mexico, as Evidence by Satellite and In Situ Data. Pac. Sci. 2018, 72, 411–422. [Google Scholar] [CrossRef]
- Espinosa-Carreon, T.L.; Strub, P.T.; Beier, E.; Ocampo-Torres, F.; Gaxiola-Castro, G. Seasonal and interannual variability of satellite-derived chlorophyll pigment, surface height, and temperature off Baja California. J. Geophys. Res. Earth Surf. 2004, 109, C03039. [Google Scholar] [CrossRef]
- López-Calderón, J.; Manzo-Monroy, H.; Santamaría-del-Angel, E.; Castro, R.; González-Silvera, A.; Millán-Núñez, R. Me-soscale variability of the Mexican Tropical Pacific using TOPEX and SeaWiFS data. Cie. Mar. 2006, 32, 539–549. [Google Scholar] [CrossRef]
- Herrera-Cervantes, H. Sea surface temperature, ocean color and wind forcing patterns in the Bay of La Paz, Gulf of California: Seasonal variability. Atmósfera 2019, 32, 25–38. [Google Scholar] [CrossRef]
- Badan-Dangon, A.; Koblinsky, C.J.; Baumgartner, T. Spring and summer in the Gulf of California: Observations of surface thermal patterns. Oce. Act. 1985, 8, 13–22. [Google Scholar]
- Cepeda-Morales, J.; Hernández-Vásquez, F.; Rivera-Caicedo, J.; Romero-Bañuelos, C.; Inda-Díaz, E.; Hernández-Almeida, O. Seasonal variability of satellite derived chlorophyll and sea surface temperature on the continental shelf of Nayarit, Mexico. Rev. Bio. Cie. 2017, 4, 17. [Google Scholar]
- Lenton, T.M.; Held, H.; Kriegler, E.; Hall, J.W.; Lucht, W.; Rahmstorf, S.; Schellnhuber, H.J. Tipping elements in the Earth’s climate system. Proc. Natl. Acad. Sci. USA 2008, 105, 1786–1793. [Google Scholar] [CrossRef]
- Timmermann, A.; Oberhuber, J.M.; Bacher, A.C.; Esch, M.; Latif, M.; Roeckner, E. Increased El Niño frequency in a climate model forced by future greenhouse warming. Nature 1999, 398, 694–697. [Google Scholar] [CrossRef]
- Cane, M.A.; Clement, A.C.; Kaplan, A.; Kushnir, Y.; Pozdnyakov, D.; Seager, R.; Zebiak, S.E.; Murtugudde, R. Twentieth-Century Sea Surface Temperature Trends. Science 1997, 275, 957–960. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Lugo, A.G.; Espinosa-Carreón, T.L.; Seminoff, J.A.; Hart, C.E.; Ley-Quiñonez, C.P.; Alonso-Aguirre, A.; Todd-Jones, T.; Zavala-Norzagaray, A. Movements of loggerhead sea turtles (Caretta caretta) in the Gulf of California: Integrating satellite telemetry and remotely sensed environmental variables. J. Mar. Bio. As. UK 2020, 100, 817–824. [Google Scholar] [CrossRef]
- Silveyra-Bustamante, A.A.; Gómez-Gutiérrez, J.; González-Rodríguez, E.; Sánchez, C.; Schiariti, A.; Mendoza-Becerril, M.A. Seasonal variability of gelatinous zooplankton during an anomalously warm year at Cabo Pulmo National Park, Mexico. Lat. Am. J. Aquat. Res. 2020, 48, 779–793. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Area in Pixels (Km2) | Priority Marine Region | Latitude | Longitude |
|---|---|---|---|---|
| 1 | 47,157 | Ensenada | 32°31′48″ to 29°45′36″N | 117°58′12″ to 115°42′W |
| 2 | 44,049 | Vizcaíno | 28°57′36″ to 26°47′24″N | 116°10′48″ to 113°43′48″W |
| 3 | 9410 | San Ignacio | 27°18′36″ to 26°4′48″N | 114°1′48″ to 112°46′48″W |
| 4 | 25,724 | Magdalena Bay | 25°47′24″ to 23°43′48″N | 112°55′48″ to 111°21′36″W |
| 5 | 30,501 | Barra de Malva-Cabo Falso | 24°21′ to 22°30′36″N | 111°51′ to 109°54′36″W |
| 6 | 3797 | Guadalupe Island | 29°22′12″ to 28°42′ N | 118°36′ to 118°2′24″W |
| 7 | 2829 | Alijos Rock | 25°08′24″ to 24°46′12″N | 115°55′48″ to 115°32′24″W |
| 8 | 262,849 | Revillagigedo Islands | 21°05′24″ to 17°24′00″N | 115°57′36″ to 109°30′00″W |
| 9 | 2171 | Los Cabos | 23°39′ to 22°49′48″N | 109°57′36″ to 109°21′36″W |
| 10 | 37,965 | Baja California Sur Island Complex | 26°31′48″ to 23°41′24″N | 111°28′12″ to 109°47′24″W |
| 11 | 1780 | Concepción Bay | 27°07′12″ to 26°31′48″N | 112°05′24″ to 111°33′W |
| 12 | 1751 | Eastern Vizcaíno Coast | 27°59′24″ to 27°29′24″N | 112°47′24″ to 112°18′36″W |
| 13 | 19,220 | Baja California Island Complex | 29°57′36″ to 28°31′36″N | 114°31′48″ to 112°12′36″W |
| 14 | 5093 | Upper Gulf | 32°10′12″ to 30°55′48″N | 115°31′48″ to 114°11′24″W |
| 15 | 1270 | Infiernillo Channel | 29°22′12″ to 28°43′48″N | 112°28′48″ to 111°43′48″W |
| 16 | 853 | Cajón del Diablo | 28°16′48″ to 27°58′48″W | 111°33′ to 111°09′36″W |
| 17 | 9632 | Southern Sonora Lagoon System | 27°34’12″ to 26°21′36″N | 110°41′24″ to 109°21′36″W |
| 18 | 8460 | Santa María La Reforma Lagoons | 25°26′24″ to 24°22′12″N | 108°51′ to 107°49′48″W |
| 19 | 8460 | Chiricahueto Lagoon | 24°29′24″ to 24°49′48″N | 107°33′ to 107°25′48″W |
| 20 | 2911 | Piaxtla-Urías | 23°48′ to 23°5′24″N | 106°55′48″ to 106°13′48″W |
| 21 | 19,424 | Marismas Nacionales | 22°41′24″ to 21°14′24″N | 106°47′24″ to 105°9′36″W |
| 22 | 7219 | Banderas Bay | 21°27′36″ to 20°23′24″N | 105°54′ to 105°11′24″W |
| 23 | 75,088 | Entrance of the Gulf | 22°51′ to 20°22′48″N | 109°56′24″ to 107°14′24″W |
| 24 | 6597 | Guaymas | 27°49′12″ to 27°17′24″N | 112°09′36″ to 110°54′36″W |
| Number | Region | El Niño Event | La Niña Event | Normal Conditions |
|---|---|---|---|---|
| 1 | Group 1 (Ensenada and Vizcaíno) | 2.66 | 3.79 | 3.52 |
| 2 | Group 2 (Baja California Sur Island Complex and Guaymas) | 2.98 | 3.42 | 3.50 |
| 3 | Group 3 (Isla Guadalupe, Alijos Rocks, Revillagigedo Islands and Gulf Entrance) | 0.44 (a, b) | 0.53 (a) | 0.43 (b) |
| 4 | Group 4 (Magdalena Bay and Barra de Malva-Cabo Falso) | 3.12 | 3.96 | 3.81 |
| 5 | San Ignacio | 6.52 | 8.29 | 7.83 |
| 6 | Los Cabos | 1.55 | 2.03 | 1.60 |
| 7 | Concepción Bay | 5.16 | 6.36 | 6.27 |
| 8 | Eastern Vizcaíno Coast | 7.57 | 7.02 | 6.84 |
| 9 | Baja California Island Complex | 9.77 | 8.57 | 7.90 |
| 10 | Upper Gulf | 9.99 | 8.86 | 8.50 |
| 11 | Infiernillo Channel | 15.94 | 17.10 | 17.68 |
| 12 | Cajón del Diablo | 6.69 | 9.13 | 10.26 |
| 13 | Southern Sonora Lagoon System | 4.98 | 7.09 | 8.75 |
| 14 | Santa María La Reforma Lagoons | 6.04 | 9.04 | 10.76 |
| 15 | Chiricahueto Lagoon | 15.62 | 20.39 | 21.31 |
| 16 | Piaxtla-Urías | 4.00 | 9.50 | 8.80 |
| 17 | Marismas Nacionales | 2.30 | 7.42 | 3.41 |
| 18 | Banderas Bay | 5.21 | 16.20 | 7.26 |
| Number | Region | El Niño Event | La Niña Event | Normal Conditions |
|---|---|---|---|---|
| 1 | Group 1 (Ensenada and Vizcaíno) | 2.38 | 2.34 | 2.39 |
| 2 | Group 2 (Baja California Sur Island Complex and Guaymas) | 1.41 | 1.71 | 1.52 |
| 3 | Group 3 (Isla Guadalupe, Alijos Rocks, Revillagigedo Islands and Gulf Entrance) | 0.42 | 0.40 | 0.39 |
| 4 | Group 4 (Magdalena Bay and Barra de Malva-Cabo Falso) | 3.29 | 3.75 | 3.87 |
| 5 | San Ignacio | 9.25 | 8.85 | 9.24 |
| 6 | Los Cabos | 1.70 | 1.88 | 1.96 |
| 7 | Concepción Bay | 2.30 | 3.11 | 2.56 |
| 8 | Eastern Vizcaíno Coast | 3.56 | 3.54 | 3.78 |
| 9 | Baja California Island Complex | 3.44 | 4.39 | 3.85 |
| 10 | Upper Gulf | 6.98 | 7.54 | 6.68 |
| 11 | Infiernillo Channel | 8.11 | 10.70 | 10.37 |
| 12 | Cajón del Diablo | 1.13 | 2.40 | 1.81 |
| 13 | Southern Sonora Lagoon System | 1.87 | 2.88 | 2.33 |
| 14 | Santa María La Reforma Lagoons | 2.87 | 4.04 | 3.24 |
| 15 | Chiricahueto Lagoon | 10.73 | 12.46 | 11.65 |
| 16 | Piaxtla-Urías | 2.11 (a, b) | 3.52 (a) | 2 (b) |
| 17 | Marismas Nacionales | 2.54 (a, b) | 3.40 (a) | 1.83 (b) |
| 18 | Banderas Bay | 2.86 (a) | 3.49 (a) | 1.84 (b) |
| Number | Region | El Niño Event | La Niña Event | Normal Conditions |
|---|---|---|---|---|
| 1 | Group 1 (Ensenada and Vizcaíno) | 1.27 | 1.52 | 1.23 |
| 2 | Group 2 (Baja California Sur Island Complex and Guaymas) | 1.72 (a, b) | 1.93 (a) | 1.47 (b) |
| 3 | Group 3 (Isla Guadalupe, Alijos Rocks, Revillagigedo Islands and Gulf Entrance) | 0.44 | 0.39 | 0.36 |
| 4 | Group 4 (Magdalena Bay and Barra de Malva-Cabo Falso) | 0.81 | 1.12 | 0.95 |
| 5 | San Ignacio | 1.77 | 2.32 | 1.98 |
| 6 | Los Cabos | 0.59 | 0.80 | 0.71 |
| 7 | Concepción Bay | 3.31 | 4.56 | 4.45 |
| 8 | Eastern Vizcaíno Coast | 5.65 | 5.86 | 5.58 |
| 9 | Baja California Island Complex | 4.55 | 4.66 | 4.32 |
| 10 | Upper Gulf | 7.60 | 8.44 | 7.10 |
| 11 | Infiernillo Channel | 12.33 | 13.04 | 13.42 |
| 12 | Cajón del Diablo | 4.76 | 4.74 | 4.37 |
| 13 | Southern Sonora Lagoon System | 3.29 (a) | 4.60 (b) | 3.51 (a) |
| 14 | Santa María La Reforma Lagoons | 3.82 (a) | 5.54 (b) | 4.48 (a, b) |
| 15 | Chiricahueto Lagoon | 17.59 | 20.07 | 18.00 |
| 16 | Piaxtla-Urías | 3.18 (a) | 4.91 (b) | 3.54 (a, b) |
| 17 | Marismas Nacionales | 2.44 | 3.84 | 2.65 |
| 18 | Banderas Bay | 1.83 | 5.35 | 2.99 |
| Number | Region | El Niño Event | La Niña Event | Normal Conditions |
|---|---|---|---|---|
| 1 | Group 1 (Ensenada and Vizcaíno) | 1.34 (a) | 2.10 (b) | 1.54 (a, b) |
| 2 | Group 2 (Baja California Sur Island Complex and Guaymas) | 3.16 | 3.54 | 3.27 |
| 3 | Group 3 (Isla Guadalupe, Alijos Rocks, Revillagigedo Islands and Gulf Entrance) | 0.50 (a, b) | 0.65 (a) | 0.47 (b) |
| 4 | Group 4 (Magdalena Bay and Barra de Malva-Cabo Falso) | 1.03 (a) | 1.67 (b) | 1.30 (a, b) |
| 5 | San Ignacio | 1.87 (a) | 2.93 (b) | 2 (a) |
| 6 | Los Cabos | 1.59 | 2.64 | 1.87 |
| 7 | Concepción Bay | 6.11 | 7.74 | 8.07 |
| 8 | Eastern Vizcaíno Coast | 3.96 | 4.76 | 5.44 |
| 9 | Baja California Island Complex | 4.03 | 5.15 | 5.18 |
| 10 | Upper Gulf | 6.95 | 8.40 | 8.01 |
| 11 | Infiernillo Channel | 12.94 | 15.83 | 17.45 |
| 12 | Cajón del Diablo | 5.93 | 6.55 | 7.41 |
| 13 | Southern Sonora Lagoon System | 6.11 | 7.30 | 7.00 |
| 14 | Santa María La Reforma Lagoons | 6.30 | 8.56 | 8.23 |
| 15 | Chiricahueto Lagoon | 15.22 | 18.43 | 19.87 |
| 16 | Piaxtla-Urías | 5.22 (a) | 10.59 (b) | 6.31 (a) |
| 17 | Marismas Nacionales | 2.30 (a) | 5.22 (b) | 2.78 (a) |
| 18 | Banderas Bay | 3.07 (a) | 7.98 (b) | 4.36 (a) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robles-Tamayo, C.M.; García-Morales, R.; Romo-León, J.R.; Figueroa-Preciado, G.; Peñalba-Garmendia, M.C.; Enríquez-Ocaña, L.F. Variability of Chl a Concentration of Priority Marine Regions of the Northwest of Mexico. Remote Sens. 2022, 14, 4891. https://doi.org/10.3390/rs14194891
Robles-Tamayo CM, García-Morales R, Romo-León JR, Figueroa-Preciado G, Peñalba-Garmendia MC, Enríquez-Ocaña LF. Variability of Chl a Concentration of Priority Marine Regions of the Northwest of Mexico. Remote Sensing. 2022; 14(19):4891. https://doi.org/10.3390/rs14194891
Chicago/Turabian StyleRobles-Tamayo, Carlos Manuel, Ricardo García-Morales, José Raúl Romo-León, Gudelia Figueroa-Preciado, María Cristina Peñalba-Garmendia, and Luis Fernando Enríquez-Ocaña. 2022. "Variability of Chl a Concentration of Priority Marine Regions of the Northwest of Mexico" Remote Sensing 14, no. 19: 4891. https://doi.org/10.3390/rs14194891