

Effects of Urea Application on Soil Organic Nitrogen Mineralization and Nitrogen Fertilizer Availability in a Rice–Broad Bean Rotation System

Abstract
:1. Introduction
2. Materials and Methods
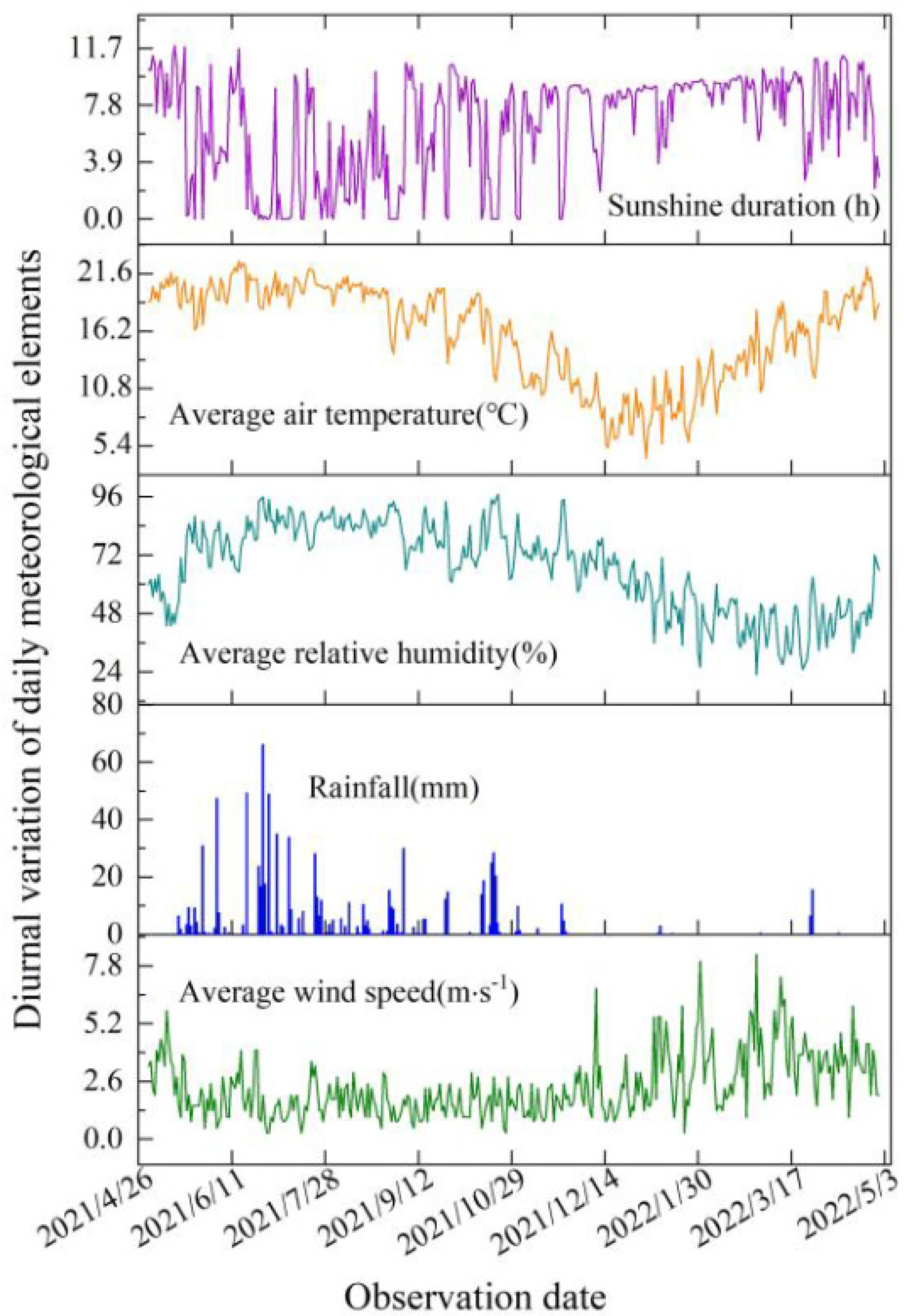
2.1. Test Area Overview
2.2. Experimental Design
2.3. Observation Contents and Methods
2.4. Study Methodology and Data Analysis
3. Results
3.1. Succession Patterns of Soil Microhabitat Environment in the RBRS
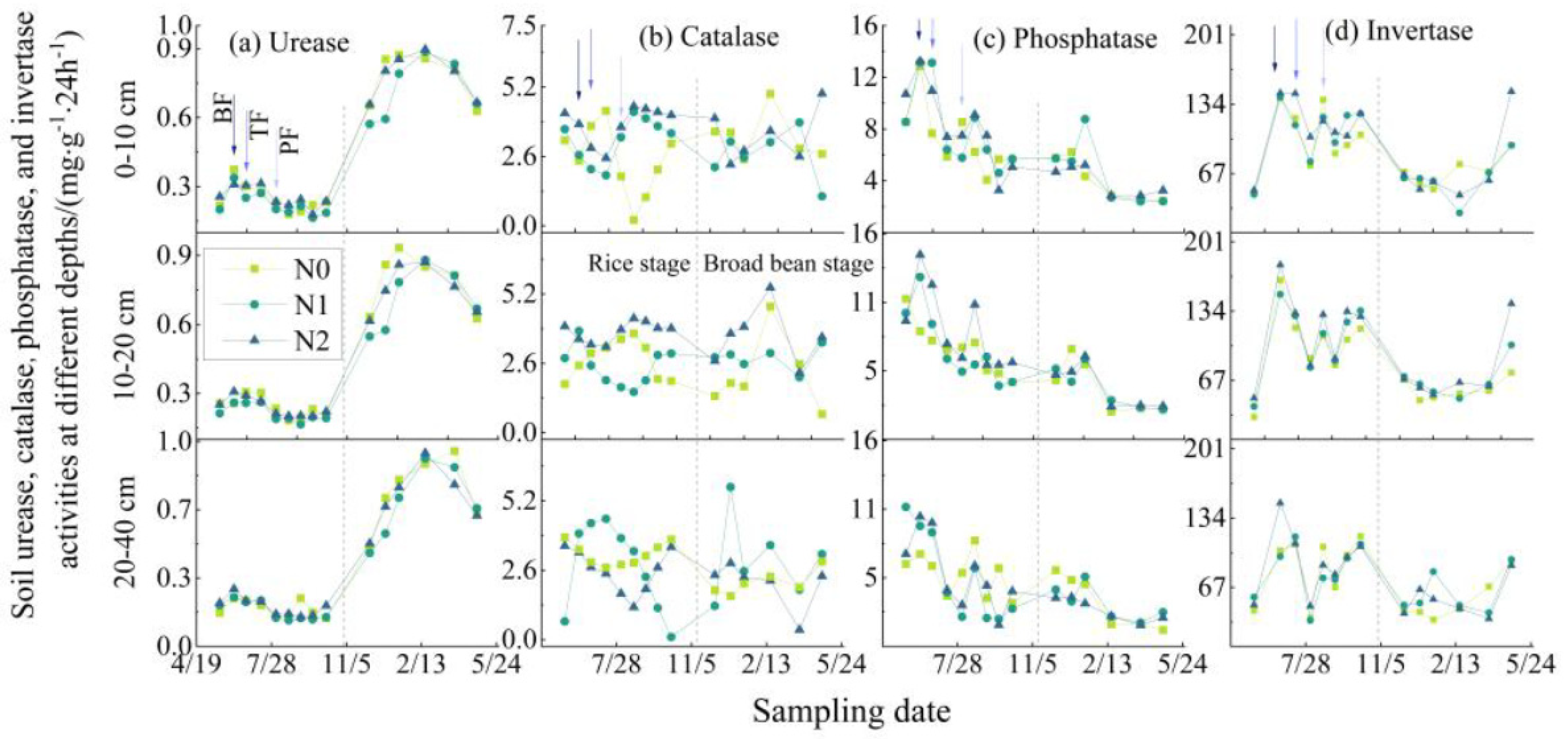
3.1.1. Soil Enzyme Activity
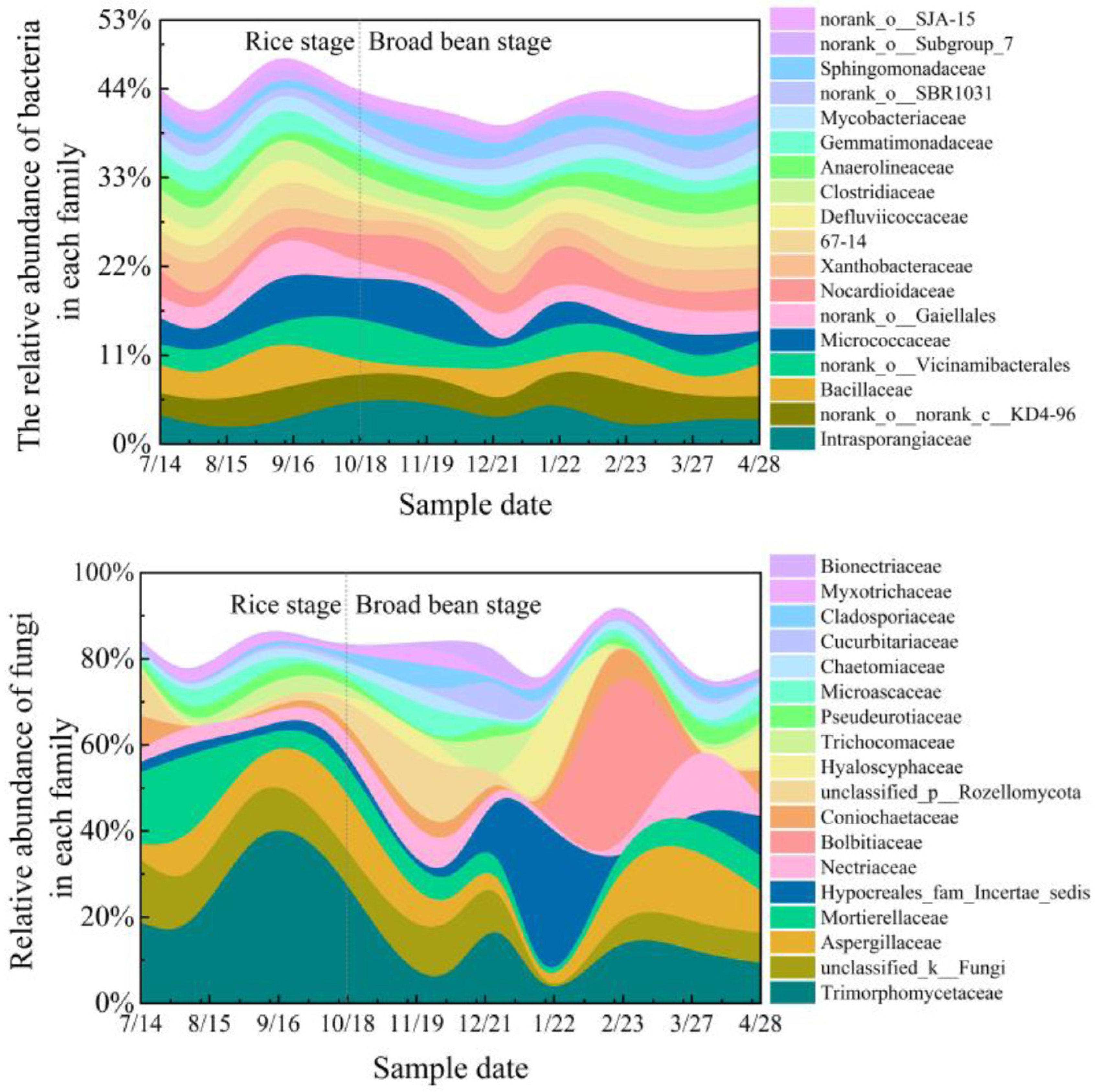
3.1.2. Soil Microbial Diversity and Abundance
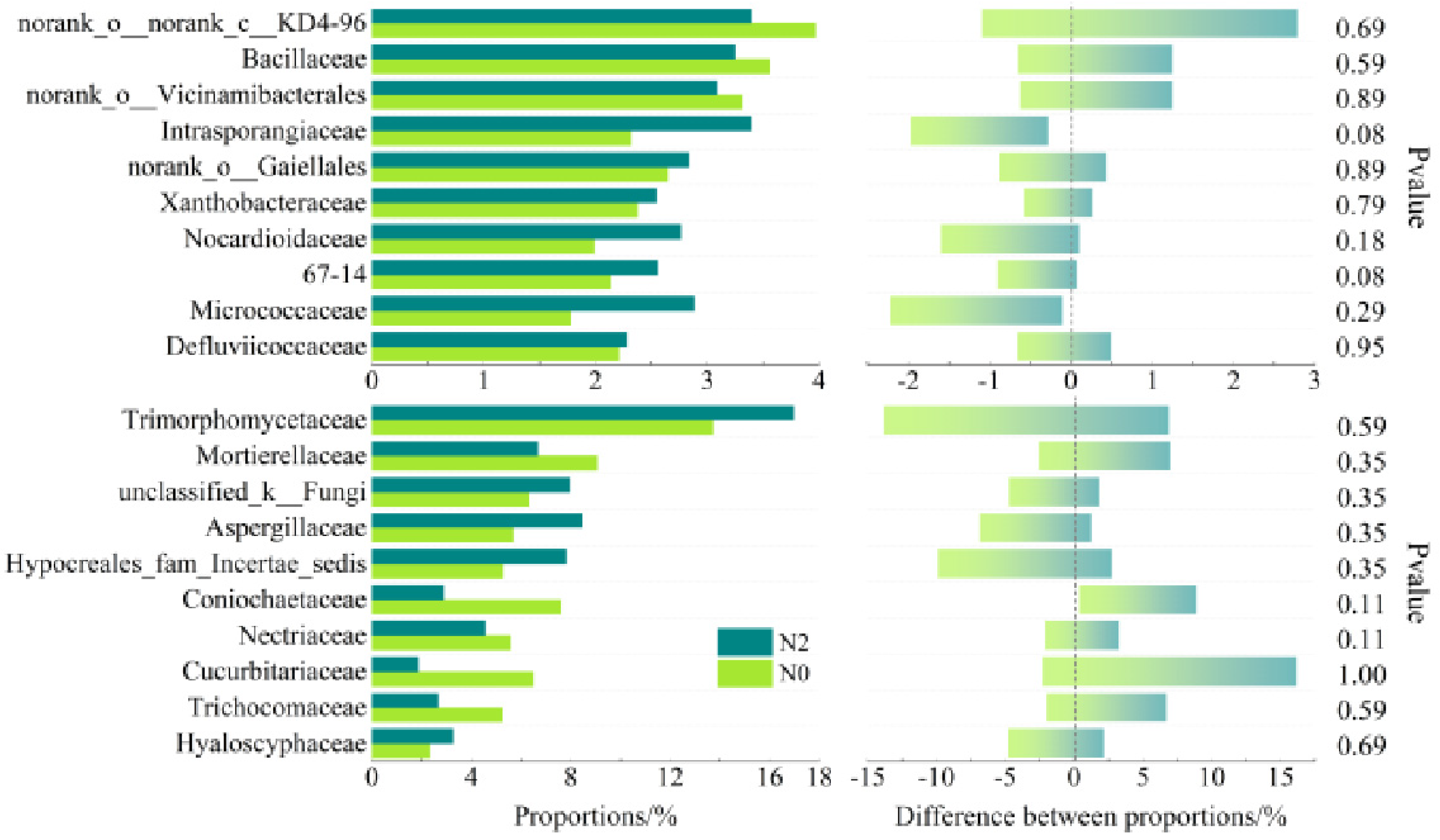
- Bacteria families’ diversity and abundance
- Fungi families’ diversity and abundance
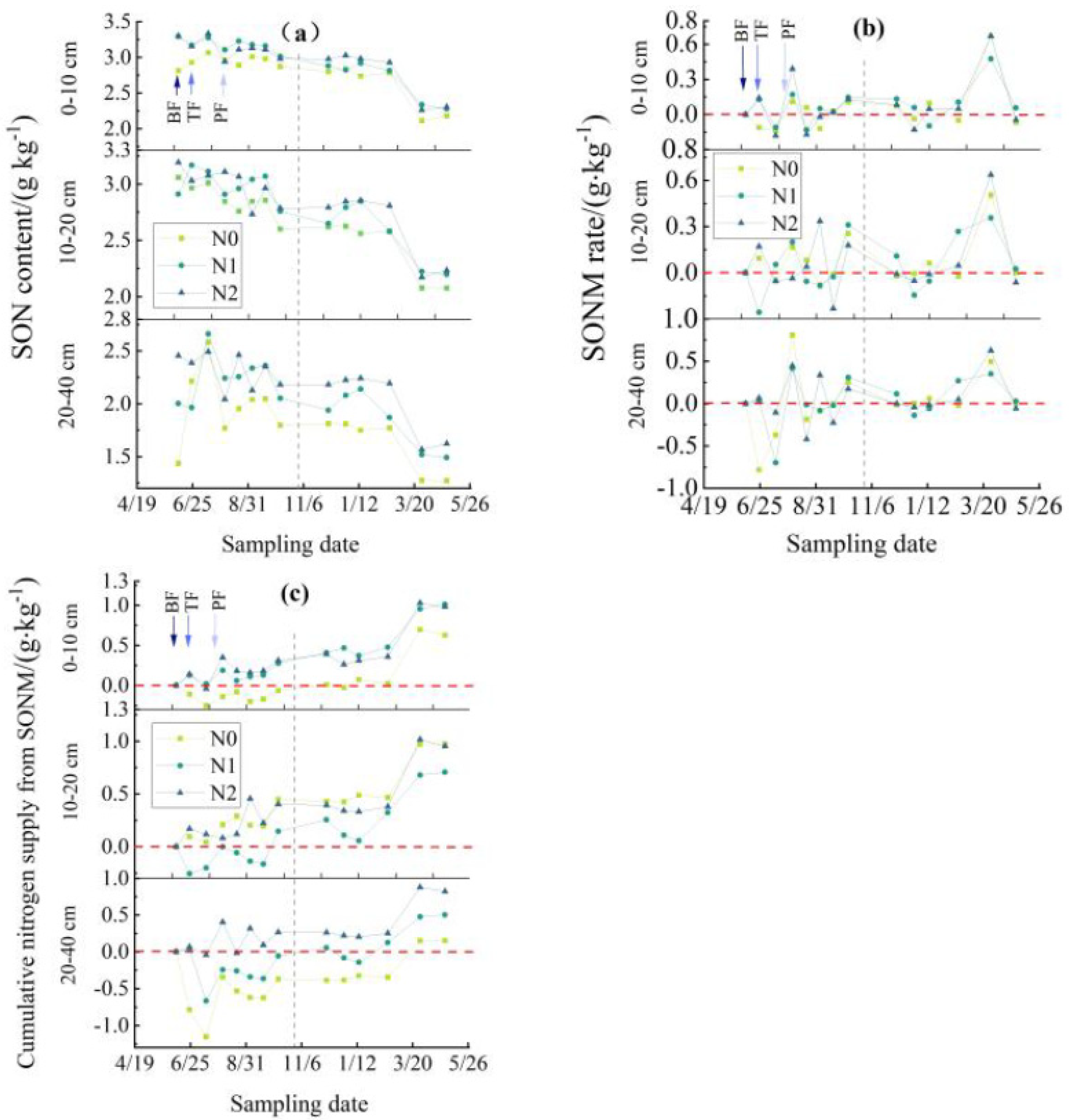
3.2. Dynamic Changes of SON Content and SONM in the RBRS
3.2.1. SON Content
3.2.2. SONM
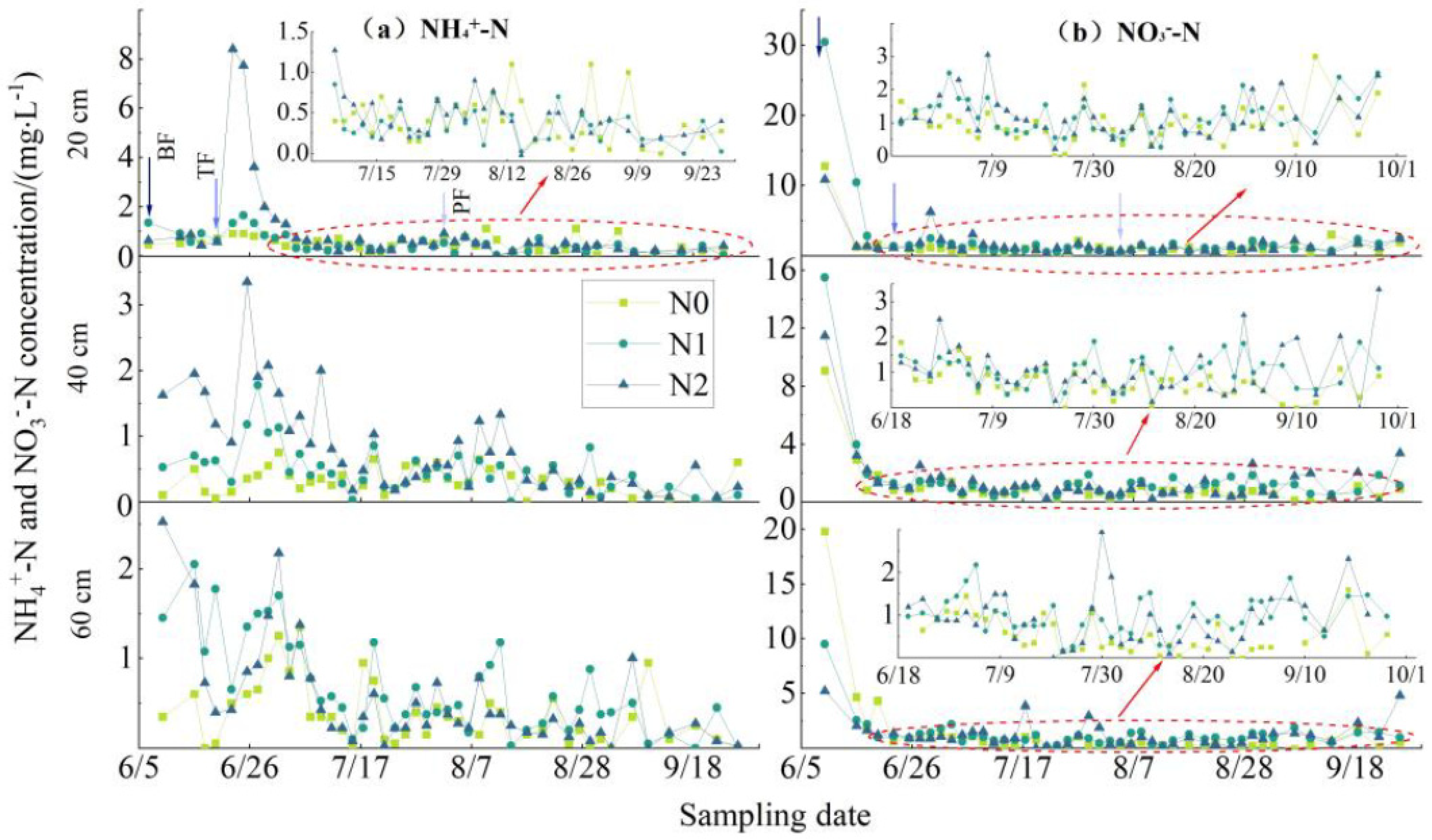
3.3. NF Utilization in the RBRS
4. Discussion
4.1. Effects of Urea Application on SONM
4.2. Effects of Urea Application on NF Availability
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cai, S.; Zhao, X.; Pittelkow, C.M.; Fan, M.; Zhang, X.; Yan, X. Optimal nitrogen rate strategy for sustainable rice production in China. Nature 2023, 615, 73–79. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). FAOSTAT: FAO Statistical Databases. Available online: http://faostat.fao.org/default.aspx (accessed on 4 December 2022).
- Mkhinini, M.; Boughattas, I.; Hattab, S.; Amamou, C.; Banni, M. Effect of treated wastewater irrigation on physiological and agronomic properties of beans Vicia faba. Int. J. Environ. Agric. Biotechnol. 2018, 3, 1414–1420. [Google Scholar] [CrossRef]
- Zhang, H.; Jing, W.J.; Zhao, B.H.; Wang, W.L.; Xu, Y.J.; Zhang, W.Y.; Gu, J.F.; Liu, L.J.; Wang, Z.Q.; Yang, J.C. Alternative fertilizer and irrigation practices improve rice yield and resource use efficiency by regulating source-sink relationships. Field Crops Res. 2021, 265, 108124. [Google Scholar] [CrossRef]
- Yao, H.; Eva, L.; Wulf, A.; Reiner, W.; Ma, C.; Georg, V.; Jan, S. Drainage and leaching losses of nitrogen and dissolved organic carbon after introducing maize into a continuous paddy-rice crop rotation. Agric. Ecosyst. Environ. 2017, 249, 91–100. [Google Scholar]
- Han, H.; Gao, R.; Cui, Y.; Gu, S. Transport and transformation of water and nitrogen under different irrigation modes and urea application regimes in paddy fields. Agric. Water Manag. 2021, 255, 107024. [Google Scholar] [CrossRef]
- Zhang, M.; Yao, Y.; Zhao, M.; Zhang, B.; Tian, Y.; Yin, B.; Zhu, Z. Integration of urea deep placement and organic addition for improving yield and soil properties and decreasing N loss in paddy field. Agric. Ecosyst. Environ. 2017, 247, 236–245. [Google Scholar] [CrossRef]
- Han, H.; Gao, R.; Cui, Y.; Gu, S. A semi-empirical semi-process model of ammonia volatilization from paddy fields under different irrigation modes and urea application regimes. Agric. Water Manag. 2022, 272, 107841. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, A.; Liu, J.; Liu, H.; Lei, B.; Zhai, L.; Zhang, D.; Wang, H. Cropping systems affect paddy soil organic carbon and total nitrogen stocks (in rice-garlic and rice-fava systems) in temperate region of southern China. Sci. Total Environ. 2017, 609, 1640–1649. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Wang, Y.; Zhu, H.; Wei, X.; Shao, M. Drying-wetting cycles consistently increase net nitrogen mineralization in 25 agricultural soils across the intensity and number of drying-wetting cycles. Sci. Total Environ. 2020, 710, 135574. [Google Scholar] [CrossRef]
- Ma, Y.; Shen, S.; Wan, C.; Wang, S.; Yang, F.; Zhang, K.; Gao, W. Organic fertilizer substitution over six years improves the productivity of garlic, bacterial diversity, and microbial communities network complexity. Appl. Soil Ecol. 2023, 182, 104718. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Y.; Chang, S.X.; Xu, Q.; Li, Y.; Ma, Z.; Qin, H.; Cai, Y. Linking enhanced soil nitrogen mineralization to increased fungal decomposition capacity with Moso bamboo invasion of broadleaf fore. Sci. Total Environ. 2021, 771, 144779. [Google Scholar] [CrossRef] [PubMed]
- Yemadje, P.L.; Chevallier, T.; Guibert, H.; Bertrand, I.; Bernoux, M. Wetting-drying cycles do not increase organic carbon and nitrogen mineralization in soils with straw amendment. Geoderma 2016, 304, 68–75. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Q.; Gao, J.; Hu, Y.; Song, M.; Yue, Y. Effects of rainfall amount and frequency on soil nitrogen mineralization in Zoige alpine wetland. Eur. J. Soil Biol. 2020, 97, 103170. [Google Scholar] [CrossRef]
- Xu, P.; Liu, Y.; Zhu, J.; Shi, L.; Fu, Q.; Chen, J.; Hu, H.; Huang, Q. Influence mechanisms of long-term fertilizations on the mineralization of organic matter in Ultisol. Soil Tillage Res. 2020, 201, 104594. [Google Scholar] [CrossRef]
- Shen, H.; Zhang, Q.; Zhang, X.; Jiang, X.; Zhu, S.; Chen, A.; Wu, Z.; Xiong, Z. In situ effects of biochar field-aged for six years on net N mineralization in paddy soil. Soil Tillage Res. 2021, 205, 104766. [Google Scholar] [CrossRef]
- Gao, J.; Feng, J.; Zhang, X.; Yu, F.; Xu, X.; Kuzyakov, Y. Drying-rewetting cycles alter carbon and nitrogen mineralization in litter-amended alpine wetland soil. CATENA 2016, 145, 285–290. [Google Scholar] [CrossRef]
- Liu, N.; Li, Y.; Cong, P.; Wang, J.; Guo, W.; Pang, H.; Li, Z. Depth of straw incorporation significantly alters crop yield, soil organic carbon and total nitrogen in the north china plain. Soil Tillage Res. 2021, 205, 104772. [Google Scholar] [CrossRef]
- Rodríguez, A.; Durán, J.; Rey, A.; Boudouris, I.; Valladares, F.; Gallardo, A.; Yuste, J.C. Interactive effects of forest die-off and drying-rewetting cycles on C and N mineralization. Geoderma 2019, 333, 81–89. [Google Scholar] [CrossRef]
- Dai, X.; Wang, H.; Fu, X. Soil microbial community composition and its role in carbon mineralization in long-term fertilization paddy soils. Sci. Total Environ. 2017, 580, 556–563. [Google Scholar] [CrossRef]
- Tian, J.; Lou, Y.; Gao, Y.; Fang, H.; Liu, S.; Xu, M.; Blagodatskaya, E.; Kuzyakov, Y. Response of soil organic matter fractions and composition of microbial community to long-term organic and mineral fertilization. Biol. Fertil. Soils 2017, 53, 523–532. [Google Scholar] [CrossRef]
- Clivot, H.; Mary, B.; Valé, M.; Cohan, J.P.; Champolivier, L.; Piraux, F.; Laurent, F.; Justes, E. Quantifying in situ and modeling net nitrogen mineralization from soil organic matter in arable cropping systems. Soil Biol. Biochem. 2017, 111, 44–59. [Google Scholar] [CrossRef]
- Scherger, L.E.; Valdes-Abellan, J.; Zanello, V.; Lexow, C. Projecting climate change effect on soil water fluxes and urea fertilizer fate in the semiarid Pampas of Argentina. Earth Syst. Environ. 2022, 6, 745–758. [Google Scholar] [CrossRef]
- Chen, D.; Yuan, L.; Liu, Y.; Ji, J.; Hou, H. Long-term application of manures plus chemical fertilizers sustained high rice yield and improved soil chemical and bacterial properties. Eur. J. Agron. 2017, 90, 34–42. [Google Scholar] [CrossRef]
- Hu, R.; Wang, X.; Xu, J.; Zhang, Y.; Pan, Y.; Su, X. Themechanism of soil nitrogen transformation under different biocrusts to warming and reduced precipitation: From microbial functional genes to enzyme activity. Sci. Total Environ. 2020, 722, 137849. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Tian, H.; Niu, H.; Wang, Z.; Dai, Y.; Megharaj, M.; He, W. Soil phosphatase assay to evaluate arsenic toxicity should be performed at the soil’s actual pH. Sci. Total Environ. 2023, 859, 160184. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, H.; Li, J.; Wang, Z.; Jiang, Y. Adsorption of soil invertase to goethite, gibbsite and their organic complexes and the effects on enzyme catalytic performance. Colloids Surf. B Biointerfaces 2023, 222, 113073. [Google Scholar] [CrossRef]
- Chabot, M.; Morales, E.; Cummings, J.; Rios, N.; Giatpaiboon, S.; Mogul, R. Simple kinetics, assay, and trends for soil microbial catalases. Anal. Biochem. 2020, 610, 113901. [Google Scholar] [CrossRef] [PubMed]
- Smets, W.; Leff, J.W.; Bradford, M.A.; McCulley, R.L.; Lebeer, S.; Fierer, N. A method for simultaneous measurement of soil bacterial abundances and community composition via 16S rRNA gene sequencing. Soil Biol. Biochem. 2016, 96, 145–151. [Google Scholar] [CrossRef]
- Liu, F.; Wang, S.; Huang, K.; Yeager, K.M.; Li, Y.; Lv, L.; Jia, G.; Ma, B. Great and fast increase in soil CH4 uptake after reforestation in karst cropland area is linked to the environmental and microbial factors. Agric. Ecosyst. Environ. 2023, 347, 108367. [Google Scholar] [CrossRef]
- Du, S.; Zhang, Z.; Chen, P.; Li, T.; Han, Y.; Song, J. Fate of each period fertilizer N in Mollisols under water and N management: A 15N tracer study. Agric. Water Manag. 2022, 272, 107872. [Google Scholar] [CrossRef]
- Li, G.; Lin, J.; Xue, L.; Ding, Y.; Wang, S.; Yang, L. Fate of basal N under split fertilization in rice with 15N isotope tracer. Pedosphere 2018, 28, 135–143. [Google Scholar] [CrossRef]
- Quan, Z.; Zhang, X.; Fang, E.; Davidson, A. Different quantification approaches for nitrogen use efficiency lead to divergent estimates with varying advantages. Nat. Food 2021, 2, 241–245. [Google Scholar] [CrossRef]
- Shahbaz, M.; Kumarb, A.; Kuzyakov, Y.; Börjessona, G.; Blagodatskay, E. Priming effects induced by glucose and decaying plant residues on SOM decomposition: A three-source 13C/14C partitioning study. Soil Biol. Biochem. 2018, 121, 138–146. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, H.; Lu, X.; Wang, M.; Chu, Q.; Wen, X. Lowering carbon footprint of winter wheat by improving management practices in north china plain. J. Clean. Prod. 2016, 112, 149–157. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, W.; Duan, P.; Cong, Y.; An, T.; Yu, N.; Zou, H.; Dang, X.; An, J.; Fan, Q.; et al. Evaluation and simulation of nitrogen mineralization of paddy soils in Mollisols area of Northeast China under waterlogged incubation. PLoS ONE 2017, 12, e0171022. [Google Scholar] [CrossRef]
- Hegab, A.; Fayed, M.; Hamada, M.; Abdrabbo, M. Productivity and irrigation requirements of faba-bean in North Delta of Egypt in relation to planting dates. Ann. Agric. Sci. 2014, 59, 185–193. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, S.; Xing, G. Maintaining rice yield and reducing N pollution by substituting winter legume for wheat in a heavily-fertilized rice-based cropping system of southeast China. Agric. Ecosyst. Environ. 2015, 202, 79–89. [Google Scholar] [CrossRef]
- Zhang, L.; Ruiz-Menjivar, J.; Tong, Q.; Zhang, J.; Yue, M. Examining the carbon footprint of rice production and consumption in Hubei, China: A life cycle assessment and uncertainty analysis approach. J. Environ. Manag. 2021, 300, 113698. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Zhang, M.; Tian, Y.; Zhao, M.; Zhang, B.; Zhao, M.; Zeng, K.; Yin, B. Duckweed (Spirodela polyrhiza) as green manure for increasing yield and reducing nitrogen loss in rice production. Field Crops Res. 2017, 214, 273–282. [Google Scholar] [CrossRef]
- Dong, W.; Zhang, X.; Dai, X.; Fu, X.; Yang, F.; Liu, X.; Sun, X.; Wen, X.; Schaeffer, S. Changes in soil microbial community composition in response to fertilization of paddy soils in subtropical China. Appl. Soil Ecol. 2014, 84, 140–147. [Google Scholar] [CrossRef]
- Lin, Y.; Ye, G.; Kuzyakov, Y.; Liu, D.; Fan, J.; Ding, W. Long-term manure application increases soil organic matter and aggregation, and alters microbial community structure and keystone taxa. Soil Biol. Biochem. 2019, 134, 187–196. [Google Scholar] [CrossRef]
- Wang, J.; Wu, L.; Xiao, Q.; Huang, Y.; Liu, K.; Wu, Y.; Li, D.; Duan, Y.; Zhang, W. Long-term manuring enhances soil gross nitrogen mineralization and ammonium immobilization in subtropical area. Agric. Ecosyst. Environ. 2023, 348, 108439. [Google Scholar] [CrossRef]
- Liu, W.; Qiao, C.; Yang, S.; Bai, W.; Liu, L. Microbial carbon use efficiency and priming effect regulate soil carbon storage under nitrogen deposition by slowing soil organic matter decomposition. Geoderma 2018, 332, 37–44. [Google Scholar] [CrossRef]
- Klink, S.; Keller, A.B.; Wild, A.J.; Baumert, V.L.; Gube, M.; Lehndorff, E.; Meyer, N.; Mueller, C.W.; Phillips, R.P.; Pausch, J. Stable isotopes reveal that fungal residues contribute more to mineral-associated organic matter pools than plant residues. Soil Biol. Biochem. 2022, 168, 108634. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Gao, S.; Wang, P.; Qiu, J.; Shang, S. Impacts of simulated nitrogen deposition on soil enzyme activity in a northern temperate forest ecosystem depend on the form and level of added nitrogen. Eur. J. Soil Biol. 2021, 103, 103287. [Google Scholar] [CrossRef]
- Honeker, L.K.; Gullo, C.F.; Neilson, J.W.; Chorover, J.; Maier, R.M. Effect of Re-acidification on Buffalo Grass Rhizosphere and Bulk Microbial Communities During Phytostabilization of Metalliferous Mine Tailings. Front. Microbiol. 2019, 10, 1209. [Google Scholar] [CrossRef]
- Jemaneh, Z.H.; Bobbi, L.H.; Sandra, F.Y.; Henry, H.J.; Benjamin, H.E.; Edward, G.G. Warming effects on the structure of bacterial and fungal communities in diverse soils. Appl. Soil Ecol. 2021, 163, 103973. [Google Scholar]
- Yufita, D.C.; Yoshitaka, U.; Hajime, A. Roles of soil bacteria and fungi in controlling the availability of nitrogen from cover crop residues during the microbial hot moments. Appl. Soil Ecol. 2021, 168, 104135. [Google Scholar]
- Deng, S.; Zheng, X.; Chen, X.; Zheng, S.; He, X.; Ge, T.; Kuzyakov, Y.; Wu, J.; Su, Y.; Hu, Y. Divergent mineralization of hydrophilic and hydrophobic organic substrates and their priming effect in soils depending on their preferential utilization by bacteria and fungi. Biol. Fertil. Soils 2021, 57, 65–76. [Google Scholar] [CrossRef]
- Kong, Y.; Kuzyakov, Y.; Ruan, Y.; Zhang, J.; Wang, T.; Wang, M.; Guo, S.; Shen, Q.; Ling, N. DNA stable-isotope probing delineates carbon flows from rice residues into soil microbial communities depending on fertilization. Appl. Environ. Microbiol. 2020, 86, e02151-19. [Google Scholar] [CrossRef]
- Rashad, M.; Hafez, M.; Popov, A.I. Humic substances composition and properties as an environmentally sustainable system: A review and way forward to soil conservation. J. Plant Nutr. 2022, 45, 7. [Google Scholar] [CrossRef]
- Smith, C.J.; Chalk, P.M. The residual value of fertilizer N in crop sequences: An appraisal of 60 years of research using 15N tracer. Field Crop Res. 2018, 217, 66–74. [Google Scholar] [CrossRef]
- Compton, J.E.; Pearlstein, S.L.; Erban, L.; Coulombe, R.A.; Hatteberg, B.; Henning, A.; Brooks, J.R.; Selker, J.E. Nitrogen inputs best predict farm field nitrate leaching in the Willamette Valley, Oregon. Nutr. Cycl. Agroecosyst. 2021, 120, 223–242. [Google Scholar] [CrossRef] [PubMed]
- Hafez, M.; Popov, A.I.; Rashad, M. Evaluation of the effects of new environmental additives compared to mineral fertilizers on the leaching characteristics of some anions and cations under greenhouse plant growth of saline-sodic soils. Open Agric. J. 2020, 14, 246–256. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Rice Season (g kg−1) | Broad Bean Season (g kg−1) | ||||
|---|---|---|---|---|---|---|
| 0–10 cm | 10–20 cm | 20–40 cm | 0–10 cm | 10–20 cm | 20–40 cm | |
| N0 | 3.32c | 3.14b | 2.56b | 3.22c | 2.47c | 2.32c |
| N1 | 3.56a | 3.387a | 2.48c | 3.37b | 2.59b | 2.49a |
| N2 | 3.51b | 3.39a | 2.69a | 3.41a | 2.62a | 2.44b |
| Plants | Treatments | Dry Matter (t ha−1) | Nitrogen Content (%) | NA (kg ha−1) | PAPC (%) | U (kg ha−1) | RNE (%) | Yield (kg ha−1) |
|---|---|---|---|---|---|---|---|---|
| Rice | N0 | 11.3b | 0.994b | 112.2c | 0.366 | 0 | 5.01b | |
| N1 | 18.7a | 1.105a | 206.6b | 1.685 | 51.4a | 38.1a | 11.8a | |
| N2 | 20.9a | 1.157a | 241.6a | 1.763 | 63.7a | 33.0a | 11.7a | |
| Broad bean | N0 in rice season | 4.0a | 1.765a | 70.9b | 0.366 | 0 | 1.96b | |
| N1 in rice season | 5.2a | 1.854a | 95.7a | 0.665 | 5.4a | 4.0a | 2.69a | |
| N2 in rice season | 5.6a | 1.851a | 103.5a | 0.724 | 7.0a | 3.6a | 2.73a |
| Urease | Catalase | Phosphatase | Invertase | SONM Rate | |
|---|---|---|---|---|---|
| Urease | 1 | ||||
| Catalase | −0.138 | 1 | |||
| Phosphatase | −0.509 ** | 0.188 | 1 | ||
| Invertase | −0.651 ** | 0.142 | 0.608 ** | 1 | |
| SONM rate | 0.272 * | −0.181 | −0.276 * | −0.342 ** | 1 |
| Soil Layer | Fungi | Bacteria | |||||
|---|---|---|---|---|---|---|---|
| Aspergillaceae | Nectriaceae | Pseudeurotiaceae | Chaetomiaceae | Unclassified_o __Helotiales | Unclassified_c__ Tremellomycetes | Norank_o __C0119 | |
| 0–10 cm | 0.717 * | 0.793 ** | 0.530 | 0.594 | 0.744 * | 0.519 | −0.406 |
| 10–20 cm | 0.707 * | 0.753 * | 0.703 * | 0.750 * | 0.709 * | 0.059 | −0.826 ** |
| 20–40 cm | 0.707 * | 0.657 * | 0.706 * | 0.535 | 0.698 * | 0.638 * | −0.491 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Han, H.; Gu, S.; Gao, R. Effects of Urea Application on Soil Organic Nitrogen Mineralization and Nitrogen Fertilizer Availability in a Rice–Broad Bean Rotation System. Sustainability 2023, 15, 6091. https://doi.org/10.3390/su15076091
Liu X, Han H, Gu S, Gao R. Effects of Urea Application on Soil Organic Nitrogen Mineralization and Nitrogen Fertilizer Availability in a Rice–Broad Bean Rotation System. Sustainability. 2023; 15(7):6091. https://doi.org/10.3390/su15076091
Chicago/Turabian StyleLiu, Xinyan, Huanhao Han, Shixiang Gu, and Rong Gao. 2023. "Effects of Urea Application on Soil Organic Nitrogen Mineralization and Nitrogen Fertilizer Availability in a Rice–Broad Bean Rotation System" Sustainability 15, no. 7: 6091. https://doi.org/10.3390/su15076091