
Microbe-Assisted Rhizoremediation of Hydrocarbons and Growth Promotion of Chickpea Plants in Petroleum Hydrocarbons-Contaminated Soil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Samples Collection and Lab Analysis
2.2. Prescreening and Enrichment of PHCs Degrading Microbial Cultures in Liquid Media
2.3. Isolation of PHCs-Degrading Bacterial Strains from the Enriched Culture by Agar Plate Method
2.4. Characterization of PHCs-Degrading Bacterial Strains
2.5. Identification of Bacterial Strains and Preparation of Bacterial Consortia
2.6. Experimental Setup
2.7. Seedling Emergence and Growth Attributes of Chickpea Plants
2.8. Photosynthetic Pigments and Physiological Attributes of Plants
2.9. Membrane Stability Index and Relative Water Contents of Plants
2.10. Proline and Antioxidant Contents of Plants
2.11. Nitrogen, Phosphorus and Potassium in Plants and Residual PHCs in Soil
2.12. Statistical Analysis
3. Results
3.1. Seedling Emergence and Growth Attributes of Chickpea Plants
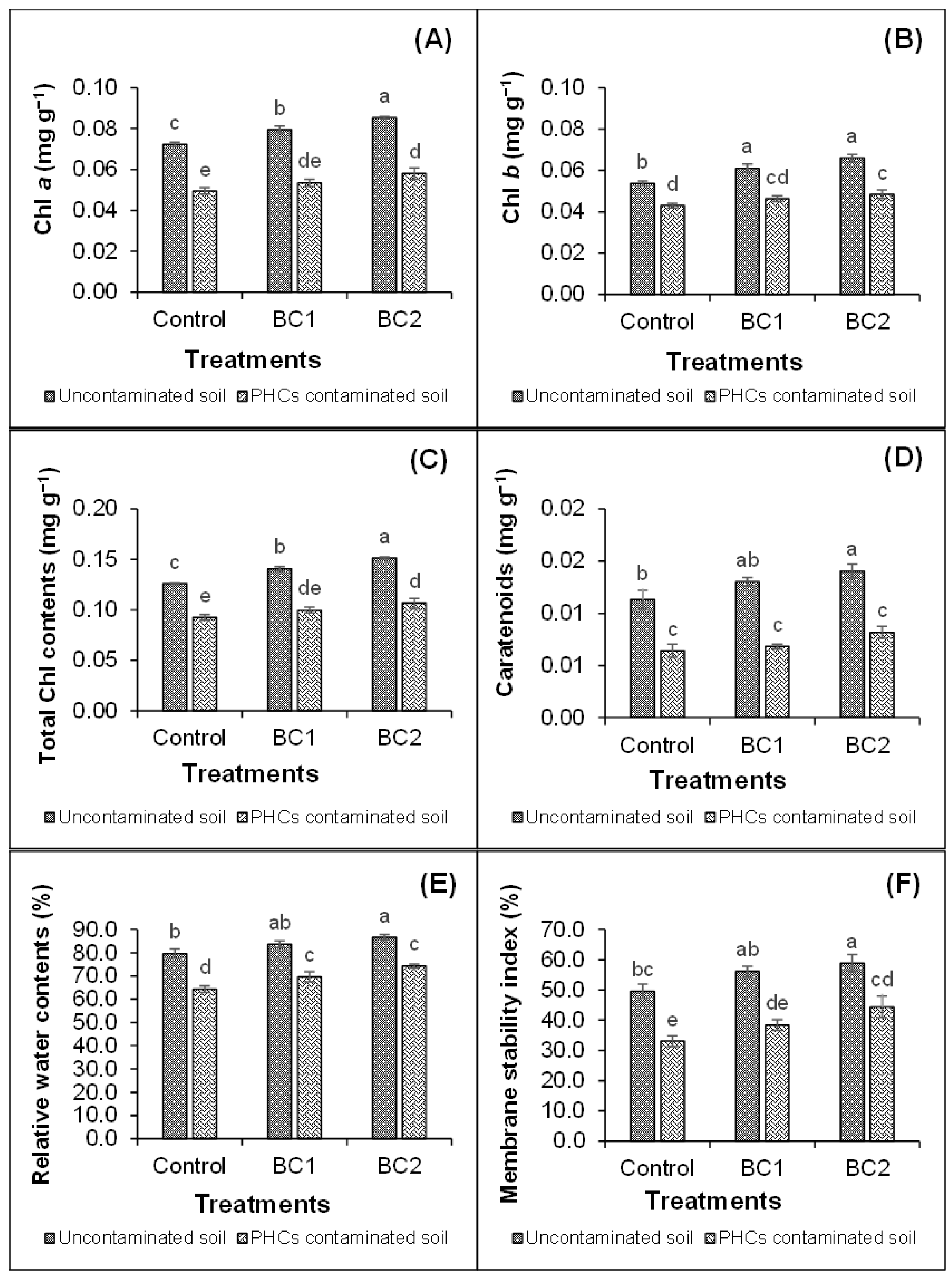
3.2. Photosynthetic Pigments and Physiological Attributes of Plants
3.3. Proline and Antioxidant Contents of Chickpea Plants
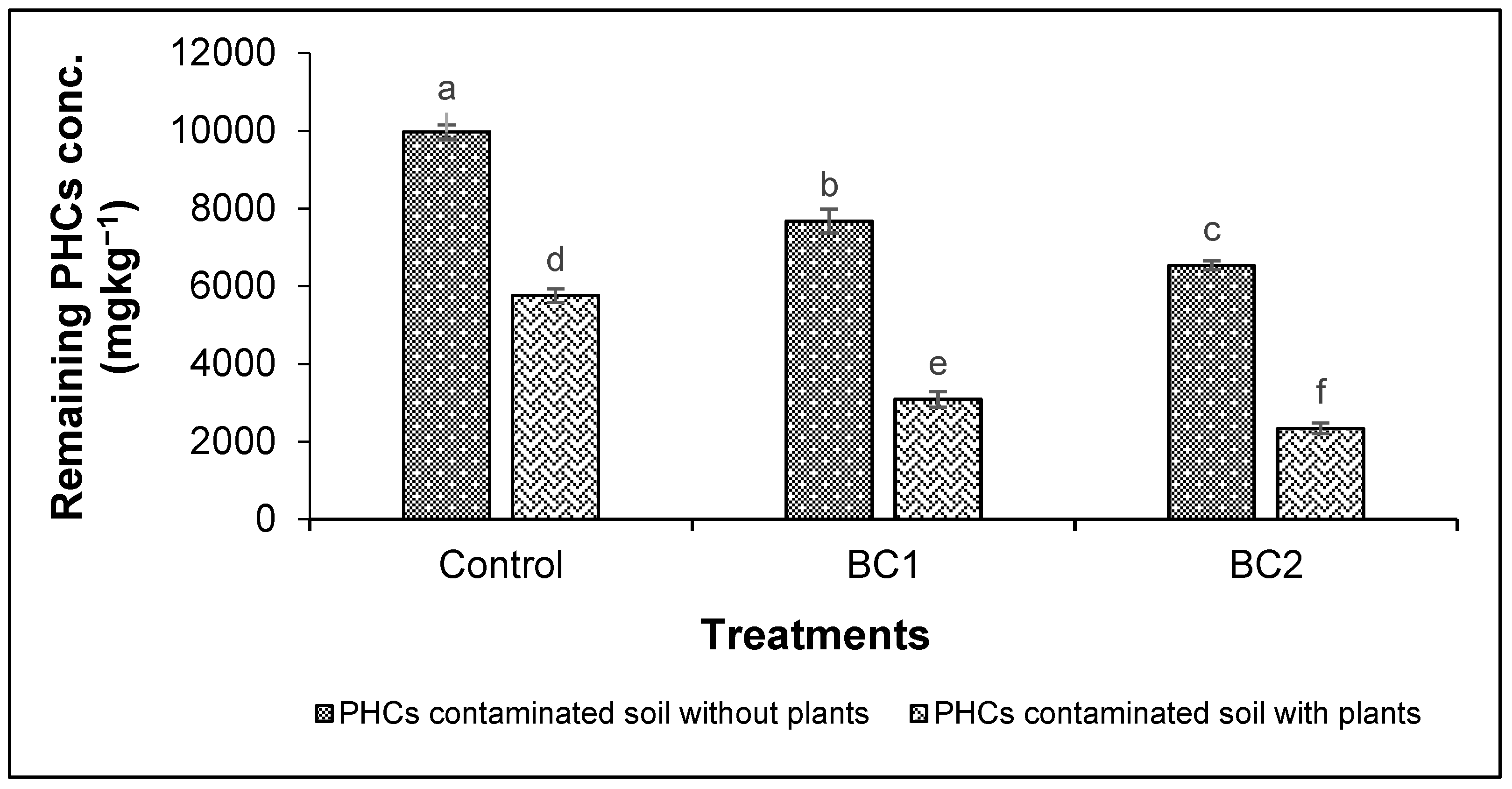
3.4. Nutrient Uptake by Plants and Removal of Phcs from Soil
4. Discussion
4.1. Seedling Emergence and Growth Attributes of Chickpea Plants
4.2. Photosynthetic Pigments and Physiological Attributes of Plants
4.3. Proline and Antioxidant Contents of Chickpea Plants
4.4. Nutrient Uptake by Plants and Removal of PHCs from Soil
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.A.I.; Biswas, B.; Smith, E.; Naidu, R.; Megharaj, M. Toxicity assessment of fresh and weathered petroleum hydrocarbons in contaminated soil—A review. Chemosphere 2018, 212, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, S.R.S.; Al-Baldawi, I.A.; Almansoory, A.F.; Purwanti, I.F.; Al-Sbani, N.H.; Sharuddin, S.S.N. Plant-assisted remediation of hydrocarbons in water and soil: Application, mechanisms, challenges and opportunities. Chemosphere 2020, 247, 125932. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Khan, W.U.; Shah, A.A.; Yasin, N.A.; Ali, A.; Rizwan, M.; Ali, S. Dopamine alleviates hydrocarbon stress in Brassica oleracea through modulation of physio-biochemical attributes and antioxidant defense systems. Chemosphere 2021, 270, 128633. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Cheema, S.A.; Shen, C.; Hassan, I.; Chen, Y. Phytotoxicity assessment of phenanthrene and pyrene in soil using two barley genotypes. Toxicol. Environ. Chem. 2014, 96, 94–105. [Google Scholar] [CrossRef]
- Liu, W.; Hou, J.; Wang, Q.; Ding, L.; Luo, Y. Isolation and characterization of plant growth-promoting rhizobacteria and their effects on phytoremediation of petroleum-contaminated saline-alkali soil. Chemosphere 2014, 117, 303–308. [Google Scholar] [CrossRef]
- Asghar, H.N.; Rafique, H.M.; Zahir, Z.A.; Khan, M.Y.; Akhtar, M.J.; Naveed, M.; Saleem, M. Petroleum hydrocarbons-contaminated soils: Remediation approaches. In Soil Science: Agricultural and Environmental Prospectives; Springer: Berlin/Heidelberg, Germany, 2016; pp. 105–129. [Google Scholar]
- Viesser, J.A.; Sugai-Guerios, M.H.; Malucelli, L.C.; Pincerati, M.R.; Karp, S.G.; Maranho, L.T. Petroleum-tolerant rhizospheric bacteria: Isolation, characterization and bioremediation potential. Sci. Rep. 2020, 10, 2060. [Google Scholar] [CrossRef] [Green Version]
- Varjani, S.J. Microbial degradation of petroleum hydrocarbons. Bioresour. Technol. 2017, 223, 277–286. [Google Scholar] [CrossRef]
- Truskewycz, A.; Gundry, T.D.; Khudur, L.S.; Kolobaric, A.; Taha, M.; Aburto-Medina, A.; Ball, A.S.; Shahsavari, E. Petroleum hydrocarbon contamination in terrestrial ecosystems—Fate and microbial responses. Molecules 2019, 24, 3400. [Google Scholar] [CrossRef] [Green Version]
- Almansoory, A.F.; Hasan, H.A.; Abdullah, S.R.S.; Idris, M.; Anuar, N.; Al-Adiwish, W.M. Biosurfactant produced by the hydrocarbon-degrading bacteria: Characterization, activity and applications in removing TPH from contaminated soil. Environ. Technol. Innov. 2019, 14, 100347. [Google Scholar] [CrossRef]
- Cui, J.; Chen, H.; Sun, M.; Wen, J. Comparison of bacterial community structure and function under different petroleum hydrocarbon degradation conditions. Bioprocess Biosyst. Eng. 2020, 43, 303–313. [Google Scholar] [CrossRef]
- Margesin, R.; Hämmerle, M.; Tscherko, D. Microbial activity and community composition during bioremediation of diesel-oil-contaminated soil: Effects of hydrocarbon concentration, fertilizers, and incubation time. Microb. Ecol. 2007, 53, 259–269. [Google Scholar] [CrossRef]
- Mukherjee, A.; Chettri, B.; Langpoklakpam, J.S.; Basak, P.; Prasad, A.; Mukherjee, A.K.; Bhattacharyya, M.; Singh, A.K.; Chattopadhyay, D. Bioinformatic approaches including predictive metagenomic profiling reveal characteristics of bacterial response to petroleum hydrocarbon contamination in diverse environments. Sci. Rep. 2017, 7, 1108. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Sinkko, H.; Penttinen, P.; Lindström, K. Characterization of successional changes in bacterial community composition during bioremediation of used motor oil-contaminated soil in a boreal climate. Sci. Total Environ. 2016, 542, 817–825. [Google Scholar] [CrossRef] [Green Version]
- Patowary, K.; Patowary, R.; Kalita, M.C.; Deka, S. Characterization of biosurfactant produced during degradation of hydrocarbons using crude oil as sole source of carbon. Front. Microbiol. 2017, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, C.; Rappazzo, A.C.; Michaud, L.; De Domenico, E.; Rochera, C.; Camacho, A.; Giudice, A.L. Efficiency in hydrocarbon degradation and biosurfactant production by Joostella sp. A8 when grown in pure culture and consortia. J. Environ. Sci. 2018, 67, 115–126. [Google Scholar] [CrossRef]
- Kshirsagar, S.D.; Mattam, A.J.; Jose, S.; Ramachandrarao, B.; Velankar, H.R. Heavy hydrocarbons as selective substrates for isolation of asphaltene degraders: A substrate-based bacterial isolation strategy for petroleum hydrocarbon biodegradation. Environ. Technol. Innov. 2020, 19, 100832. [Google Scholar] [CrossRef]
- Xu, X.; Liu, W.; Tian, S.; Wang, W.; Qi, Q.; Jiang, P.; Gao, X.; Li, F.; Li, H.; Yu, H. Petroleum hydrocarbon-degrading bacteria for the remediation of oil pollution under aerobic conditions: A perspective analysis. Front. Microbiol. 2018, 9, 2885. [Google Scholar] [CrossRef]
- Aziz, A.; Agamuthu, P.; Alaribe, F.O.; Fauziah, S.H. Biodegradation of benzo[a]pyrene by bacterial consortium isolated from mangrove sediment. Environm. Technol. 2018, 39, 527–535. [Google Scholar] [CrossRef]
- Brzeszcz, J.; Kapusta, P.; Steliga, T.; Turkiewicz, A. Hydrocarbon removal by two differently developed microbial inoculants and comparing their actions with biostimulation treatment. Molecules 2020, 25, 661. [Google Scholar] [CrossRef] [Green Version]
- Rostami, S.; Azhdarpoor, A. The application of plant growth regulators to improve phytoremediation of contaminated soils: A review. Chemosphere 2019, 220, 818–827. [Google Scholar] [CrossRef]
- Zhang, X.; Su, C.; Liu, X.; Liu, Z.; Liang, X.; Zhang, Y.; Feng, Y. Effect of plant-growth-promoting rhizobacteria on phytoremediation efficiency of Scirpus triqueter in pyrene-Ni co-contaminated soils. Chemosphere 2020, 241, 125027. [Google Scholar] [CrossRef] [PubMed]
- Baoune, H.; El Hadj-Khelil, A.O.; Pucci, G.; Sineli, P.; Loucif, L.; Polti, M.A. Petroleum degradation by endophytic Streptomyces spp. Isolated from plants grown in contaminated soil of southern algeria. Ecotoxicol. Environ. Saf. 2018, 147, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, A.; Arshad, M.; Karthikeyan, R.; Gentry, T.J.; Rashid, J.; Ahmed, I.; Schwab, A.P. Diesel degrading bacterial endophytes with plant growth promoting potential isolated from a petroleum storage facility. 3 Biotech 2019, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Rafique, H.; Asghar, H.; Zahir, Z.; Shahbaz, M. Evaluation of plant growth promoting bacteria for inducing stress tolerance in plants against petroleum hydrocarbons. Pak. J. Agric. Sci. 2015, 52, 905–914. [Google Scholar]
- Guo, M.; Gong, Z.; Miao, R.; Rookes, J.; Cahill, D.; Zhuang, J. Microbial mechanisms controlling the rhizosphere effect of ryegrass on degradation of polycyclic aromatic hydrocarbons in an aged-contaminated agricultural soil. Soil Biol. Biochem. 2017, 113, 130–142. [Google Scholar] [CrossRef]
- García-Cruz, N.; Valdivia-Rivera, S.; Narciso-Ortiz, L.; García-Maldonado, J.; Uribe-Flores, M.; Aguirre-Macedo, M.; Lizardi-Jiménez, M. Diesel uptake by an indigenous microbial consortium isolated from sediments of the Southern Gulf of Mexico: Emulsion characterisation. Environ. Pollut. 2019, 250, 849–855. [Google Scholar] [CrossRef]
- Shahsavari, E.; Adetutu, E.M.; Anderson, P.A.; Ball, A.S. Tolerance of selected plant species to petrogenic hydrocarbons and effect of plant rhizosphere on the microbial removal of hydrocarbons in contaminated soil. Water Air Soil Pollut. 2013, 224, 1495. [Google Scholar] [CrossRef]
- Jukanti, A.K.; Gaur, P.M.; Gowda, C.; Chibbar, R.N. Nutritional quality and health benefits of chickpea (Cicer arietinum L.): A review. Br. J. Nutr. 2012, 108, S11–S26. [Google Scholar] [CrossRef] [Green Version]
- Shcherbakova, E.; Shcherbakov, A.; Andronov, E.; Gonchar, L.; Kalenskaya, S.; Chebotar, V. Combined pre-seed treatment with microbial inoculants and mo nanoparticles changes composition of root exudates and rhizosphere microbiome structure of chickpea (Cicer arietinum L.) plants. Symbiosis 2017, 73, 57–69. [Google Scholar] [CrossRef]
- FAOSTAT. Agriculture Database; FAOSTAT: Rome, Italy, 2015. [Google Scholar]
- Sani, S.G.A.S.; Chang, P.L.; Zubair, A.; Carrasquilla-Garcia, N.; Cordeiro, M.; Penmetsa, R.V.; Munis, M.F.H.; Nuzhdin, S.V.; Cook, D.R.; von Wettberg, E.J. Genetic diversity, population structure, and genetic correlation with climatic variation in chickpea (Cicer arietinum) landraces from Pakistan. Plant Genome 2018, 11, 170067. [Google Scholar] [CrossRef]
- Gartler, J.; Wimmer, B.; Soja, G.; Reichenauer, T.G. Effects of rapeseed oil on the rhizodegradation of polyaromatic hydrocarbons in contaminated soil. Int. J. Phytoremediat. 2014, 16, 671–683. [Google Scholar] [CrossRef]
- Hall, J.; Soole, K.; Bentham, R. Hydrocarbon phytoremediation in the family fabacea—A review. Int. J. Phytoremediat. 2011, 13, 317–332. [Google Scholar] [CrossRef]
- Gao, Y.; Zhu, L. Plant uptake, accumulation and translocation of phenanthrene and pyrene in soils. Chemosphere 2004, 55, 1169–1178. [Google Scholar] [CrossRef]
- Yang, H.; Su, Y.H.; Zhu, Y.G.; Chen, M.M.; Chen, B.D.; Liu, Y.X. Influences of polycyclic aromatic hydrocarbons (PAHs) on soil microbial community composition with or without vegetation. J. Environ. Sci. Health A 2007, 42, 65–72. [Google Scholar] [CrossRef]
- Xu, S.; Chen, Y.; Lin, K.; Chen, X.; Lin, Q.; Li, F.; Wang, Z. Removal of pyrene from contaminated soils by white clover. Pedosphere 2009, 19, 265–272. [Google Scholar] [CrossRef]
- Phillips, L.A.; Greer, C.W.; Germida, J.J. Culture-based and culture-independent assessment of the impact of mixed and single plant treatments on rhizosphere microbial communities in hydrocarbon contaminated flare-pit soil. Soil Biol. Biochem. 2006, 38, 2823–2833. [Google Scholar] [CrossRef]
- Thorsen, W.A.; Cope, W.G.; Shea, D. Bioavailability of PAHs: Effects of soot carbon and PAH source. Environ. Sci. Technol. 2004, 38, 2029–2037. [Google Scholar] [CrossRef]
- Estefan, G. Methods of Soil, Plant, and Water Analysis: A Manual for the West Asia and North Africa Region; ICARDA: Beirut, Lebanon, 2013. [Google Scholar]
- Ali, M.H.; Sattar, M.T.; Khan, M.I.; Naveed, M.; Rafique, M.; Alamri, S.; Siddiqui, M.H. Enhanced growth of mungbean and remediation of petroleum hydrocarbons by Enterobacter sp. MN17 and biochar addition in diesel contaminated soil. Appl. Sci. 2020, 10, 8548. [Google Scholar] [CrossRef]
- Afzal, M.J.; Khan, M.I.; Cheema, S.A.; Hussain, S.; Anwar-ul-Haq, M.; Ali, M.H.; Naveed, M. Combined application of Bacillus sp. MN-54 and phosphorus improved growth and reduced lead uptake by maize in the lead-contaminated soil. Environ. Sci. Pollut. Res. 2020, 27, 44528–44539. [Google Scholar] [CrossRef]
- Suja, F.; Rahim, F.; Taha, M.R.; Hambali, N.; Razali, M.R.; Khalid, A.; Hamzah, A. Effects of local microbial bioaugmentation and biostimulation on the bioremediation of total petroleum hydrocarbons (TPH) in crude oil contaminated soil based on laboratory and field observations. Int. Biodeterior. Biodegr. 2014, 90, 115–122. [Google Scholar] [CrossRef]
- Davis, K.E.; Joseph, S.J.; Janssen, P.H. Effects of growth medium, inoculum size, and incubation time on culturability and isolation of soil bacteria. Appl. Environ. Microbiol. 2005, 71, 826–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdani, S.A.F.; Aon, M.; Ali, L.; Aslam, Z.; Khalid, M.; Naveed, M. Application of Dalbergia sissoo biochar enhanced wheat growth, yield and nutrient recovery under reduced fertilizer doses in calcareous soil. Pak. J. Agric. Sci. 2017, 54, 5102. [Google Scholar]
- Ahamd, M.; Hussain, A.; Akhtar, M.F.-U.-Z.; Zafar-Ul-Hye, M.; Iqbal, Z.; Naz, T.; Iqbal, M.M. Effectiveness of multi-strain biofertilizer in combination with organic sources for improving the productivity of chickpea in drought ecology. Asian J. Agric. Biol. 2017, 5, 228–237. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [Green Version]
- Sairam, R.K.; Rao, K.V.; Srivastava, G. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Angelini, R.; Manes, F.; Federico, R. Spatial and functional correlation between diamine-oxidase and peroxidase activities and their dependence upon de-etiolation and wounding in chick-pea stems. Planta 1990, 182, 89–96. [Google Scholar] [CrossRef]
- Snedecor, G.W. Cochran, WG: Statistical Methods; Iowa State College Press: Ames, IA, USA, 1980. [Google Scholar]
- Ikram, M.; Ali, N.; Jan, G.; Jan, F.G.; Rahman, I.U.; Iqbal, A.; Hamayun, M. IAA producing fungal endophyte Penicillium roqueforti Thom., enhances stress tolerance and nutrients uptake in wheat plants grown on heavy metal contaminated soils. PLoS ONE 2018, 13, e0208150. [Google Scholar] [CrossRef] [Green Version]
- Barati, M.; Bakhtiari, F.; Mowla, D.; Safarzadeh, S. Total petroleum hydrocarbon degradation in contaminated soil as affected by plants growth and biochar. Environ. Earth Sci. 2017, 76, 688. [Google Scholar] [CrossRef]
- Haq, I.; Kalamdhad, A.S. Phytotoxicity and cyto-genotoxicity evaluation of organic and inorganic pollutants containing petroleum refinery wastewater using plant bioassay. Environ. Technol Innov. 2021, 23, 101651. [Google Scholar] [CrossRef]
- Graj, W.; Lisiecki, P.; Szulc, A.; Chrzanowski, Ł.; Wojtera-Kwiczor, J. Bioaugmentation with petroleum-degrading consortia has a selective growth-promoting impact on crop plants germinated in diesel oil-contaminated soil. Water Air Soil Pollut. 2013, 224, 1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singha, L.P.; Sinha, N.; Pandey, P. Rhizoremediation prospects of polyaromatic hydrocarbon degrading rhizobacteria, that facilitate glutathione and glutathione-s-transferase mediated stress response, and enhance growth of rice plants in pyrene contaminated soil. Ecotoxicol. Environ. Saf. 2018, 164, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Silambarasan, S.; Logeswari, P.; Ruiz, A.; Cornejo, P.; Kannan, V.R. Influence of plant beneficial Stenotrophomonas rhizophila strain CASB3 on the degradation of diuron-contaminated saline soil and improvement of Lactuca sativa growth. Environ. Sci. Pollut. Res. 2020, 27, 35195–35207. [Google Scholar] [CrossRef] [PubMed]
- Chicca, I.; Becarelli, S.; Bernabei, G.; Siracusa, G.; Di Gregorio, S. Innovative culturomic approaches and predictive functional metagenomic analysis: The isolation of hydrocarbonoclastic bacteria with plant growth promoting capacity. Water 2022, 14, 142. [Google Scholar] [CrossRef]
- Marques, A.P.; Pires, C.; Moreira, H.; Rangel, A.O.; Castro, P.M. Assessment of the plant growth promotion abilities of six bacterial isolates using Zea mays as indicator plant. Soil Biol. Biochem. 2010, 42, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Hussain, I.; Puschenreiter, M.; Gerhard, S.; Sani, S.G.A.S.; Reichenauer, T.G. Differentiation between physical and chemical effects of oil presence in freshly spiked soil during rhizoremediation trial. Environ. Sci. Pollut. Res. 2019, 26, 18451–18464. [Google Scholar] [CrossRef] [Green Version]
- Liang, B.; Gao, T.; Zhao, Q.; Ma, C.; Chen, Q.; Wei, Z.; Li, C.; Li, C.; Ma, F. Effects of exogenous dopamine on the uptake, transport, and resorption of apple ionome under moderate drought. Front. Plant Sci. 2018, 9, 755. [Google Scholar] [CrossRef]
- Tariq, M.; Shah, A.A.; Yasin, N.A.; Ahmad, A.; Rizwan, M. Enhanced performance of Bacillus megaterium OSR-3 in combination with putrescine ammeliorated hydrocarbon stress in Nicotiana tabacum. Int. J. Phytoremediat. 2021, 23, 119–129. [Google Scholar] [CrossRef]
- Kaya, C.; Okant, M.; Ugurlar, F.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Melatonin-mediated nitric oxide improves tolerance to cadmium toxicity by reducing oxidative stress in wheat plants. Chemosphere 2019, 225, 627–638. [Google Scholar] [CrossRef]
- Jin, X.; Liu, T.; Xu, J.; Gao, Z.; Hu, X. Exogenous GABA enhances muskmelon tolerance to salinity-alkalinity stress by regulating redox balance and chlorophyll biosynthesis. BMC Plant Biol. 2019, 19, 48. [Google Scholar] [CrossRef]
- Jabeen, M.; Akram, N.A.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Thiamin stimulates growth and secondary metabolites in turnip (Brassica rapa L.) leaf and root under drought stress. Physiol. Plant. 2021, 172, 1399–1411. [Google Scholar] [CrossRef]
- Arellano, P.; Tansey, K.; Balzter, H.; Boyd, D.S. Field spectroscopy and radiative transfer modelling to assess impacts of petroleum pollution on biophysical and biochemical parameters of the amazon rainforest. Environ. Earth Sci. 2017, 76, 217. [Google Scholar] [CrossRef] [Green Version]
- Cevher-Keskin, B.; Selçukcan-Erol, Ç.; Yüksel, B.; Ertekin, Ö.; Yıldızhan, Y.; Onarıcı, S.; Kulen, O.; Memon, A.R. Comparative transcriptome analysis of Zea mays in response to petroleum hydrocarbon stress. Environ. Sci. Pollut. Res. 2018, 25, 32660–32674. [Google Scholar] [CrossRef]
- Singh, R.; Jha, A.B.; Misra, A.N.; Sharma, P. Differential responses of growth, photosynthesis, oxidative stress, metals accumulation and NRAMP genes in contrasting Ricinus communis genotypes under arsenic stress. Environ. Sci. Pollut. Res. 2019, 26, 31166–31177. [Google Scholar] [CrossRef]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Mondal, M.H.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2018, 169, 20–32. [Google Scholar] [CrossRef]
- Sapre, S.; Gontia-Mishra, I.; Tiwari, S. ACC deaminase-producing bacteria: A key player in alleviating abiotic stresses in plants. In Plant Growth Promoting Rhizobacteria for Agricultural Sustainability; Springer: Berlin/Heidelberg, Germany, 2019; pp. 267–291. [Google Scholar]
- Zahir, Z.A.; Zafar-ul-Hye, M.; Sajjad, S.; Naveed, M. Comparative effectiveness of Pseudomonas and Serratia sp. Containing ACC-deaminase for coinoculation with Rhizobium leguminosarum to improve growth, nodulation, and yield of lentil. Biol. Fertil. Soils 2011, 47, 457–465. [Google Scholar] [CrossRef]
- Ali, M.H.; Khan, M.I.; Bashir, S.; Azam, M.; Naveed, M.; Qadri, R.; Bashir, S.; Mehmood, F.; Shoukat, M.A.; Li, Y. Biochar and Bacillus sp. MN54 assisted phytoremediation of diesel and plant growth promotion of maize in hydrocarbons contaminated soil. Agronomy 2021, 11, 1795. [Google Scholar] [CrossRef]
- Xun, F.; Xie, B.; Liu, S.; Guo, C. Effect of plant growth-promoting bacteria (PGPR) and arbuscular mycorrhizal fungi (AMF) inoculation on oats in saline-alkali soil contaminated by petroleum to enhance phytoremediation. Environ. Sci. Pollut. Res. 2015, 22, 598–608. [Google Scholar] [CrossRef]
- Rusin, M.; Gospodarek, J.; Barczyk, G.; Nadgórska-Socha, A. Antioxidant responses of Triticum aestivum plants to petroleum-derived substances. Ecotoxicology 2018, 27, 1353–1367. [Google Scholar] [CrossRef] [Green Version]
- Molina, L.; Segura, A. Biochemical and metabolic plant responses toward polycyclic aromatic hydrocarbons and heavy metals present in atmospheric pollution. Plants 2021, 10, 2305. [Google Scholar] [CrossRef] [PubMed]
- Desoky, E.-S.M.; Saad, A.M.; El-Saadony, M.T.; Merwad, A.-R.M.; Rady, M.M. Plant growth-promoting rhizobacteria: Potential improvement in antioxidant defense system and suppression of oxidative stress for alleviating salinity stress in Triticum aestivum (L.) plants. Biocatal. Agric. Biotechnol. 2020, 30, 101878. [Google Scholar] [CrossRef]
- Zhang, L.; Becker, D. Connecting proline metabolism and signaling pathways in plant senescence. Front. Plant Sci. 2015, 6, 552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Li, J.; Gu, R.; Yue, L.; Wang, H.; Zhan, X.; Xing, B. Carotenoid and superoxide dismutase are the most effective antioxidants participating in ROS scavenging in phenanthrene accumulated wheat leaf. Chemosphere 2018, 197, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, N.; Shoukat, U.; Saeed, M.; Akhtar, N.; Yasmin, H.; Khan, W.; Iqbal, S. Comparison of plant growth and remediation potential of pyrochar and thermal desorption for crude oil-contaminated soils. Sci. Rep. 2021, 11, 2817. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Egamberdieva, D.; Alam, P.; Alyemeni, M.N.; Ashraf, M. Modification of osmolytes and antioxidant enzymes by 24-epibrassinolide in chickpea seedlings under mercury (Hg) toxicity. J. Plant Growth Regul. 2018, 37, 309–322. [Google Scholar] [CrossRef]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hasanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide dismutase—Mentor of abiotic stress tolerance in crop plants. Environ. Sci. Pollut. Res. 2015, 22, 10375–10394. [Google Scholar] [CrossRef]
- Shafi, A.; Chauhan, R.; Gill, T.; Swarnkar, M.K.; Sreenivasulu, Y.; Kumar, S.; Kumar, N.; Shankar, R.; Ahuja, P.S.; Singh, A.K. Expression of SOD and APX genes positively regulates secondary cell wall biosynthesis and promotes plant growth and yield in Arabidopsis under salt stress. Plant Mol. Biol. 2015, 87, 615–631. [Google Scholar] [CrossRef]
- Hernández-Hernández, H.; Juárez-Maldonado, A.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Sánchez-Aspeytia, D.; González-Morales, S. Chitosan-pva and copper nanoparticles improve growth and overexpress the SOD and JA genes in tomato plants under salt stress. Agronomy 2018, 8, 175. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Xu, Y.; Zhao, J.; Li, F.; Gao, D.; Xing, B. Remediation of petroleum contaminated soils through composting and rhizosphere degradation. J. Hazard. Mater. 2011, 190, 677–685. [Google Scholar] [CrossRef]
- Ambreen, S.; Javed, M.; Hina, M.; Rasul, S.; Zafar, Z.U.; Manzoor, H.; Ogbaga, C.C.; Afzal, M.; Al-Qurainy, F.; Ashraf, M. Influence of sub-lethal crude oil concentration on growth, water relations and photosynthetic capacity of maize (Zea mays L.) plants. Environ. Sci. Pollut. Res. 2016, 23, 18320–18331. [Google Scholar]
- Heidari, M.; Karami, V. Effects of different mycorrhiza species on grain yield, nutrient uptake and oil content of sunflower under water stress. J. Saudi Soc. Agric. Sci. 2014, 13, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Quan, X.; Zeng, J.; Ye, L.; Chen, G.; Han, Z.; Shah, J.M.; Zhang, G. Transcriptome profiling analysis for two tibetan wild barley genotypes in responses to low nitrogen. BMC Plant Biol. 2016, 16, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, F.U.; Ejaz, M.; Cheema, S.A.; Khan, M.I.; Zhao, B.; Liqun, C.; Salim, M.A.; Naveed, M.; Khan, N.; Nunez-Delgado, A. Phytotoxicity of petroleum hydrocarbons: Sources, impacts and remediation strategies. Environ. Res. 2021, 197, 111031. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Zhou, Q.; Cai, Z.; Zhang, Z. Phytoremediation of petroleum contaminated soils by Mirabilis jalapa L. In a greenhouse plot experiment. J. Hazard. Mater. 2009, 168, 1490–1496. [Google Scholar] [CrossRef]
- Kakar, K.U.; Nawaz, Z.; Cui, Z.; Almoneafy, A.A.; Ullah, R.; Shu, Q.Y. Rhizosphere-associated Alcaligenes and Bacillus strains that induce resistance against blast and sheath blight diseases, enhance plant growth and improve mineral content in rice. J. Appl. Microbiol. 2018, 124, 779–796. [Google Scholar] [CrossRef]
- Sałek, K.; Zgoła-Grześkowiak, A.; Kaczorek, E. Modification of surface and enzymatic properties of Achromobacter denitrificans and Stenotrophomonas maltophilia in association with diesel oil biodegradation enhanced with alkyl polyglucosides. Colloids Surf. B Biointerfaces 2013, 111, 36–42. [Google Scholar] [CrossRef]
- Xiao, Y.; Jiang, R.; Wu, X.; Zhong, Q.; Li, Y.; Wang, H. Comparative genomic analysis of Stenotrophomonas maltophilia strain W18 reveals its adaptative genomic features for fegrading polycyclic aromatic hydrocarbons. Microbiol Spectr. 2021, 9, e0142021. [Google Scholar] [CrossRef]
- Sreenivas, A.; Sathyanarayana Reddy, G.; Shivaji, S. Draft genome sequence of a psychrophilic bacterium, Sphingomonas antarcticum, isolated from the soils of Schirmacher oasis, Antarctica. Genome Announc. 2014, 2, e00696-14. [Google Scholar] [CrossRef] [Green Version]
- Satti, S.M.; Castro-Aguirre, E.; Shah, A.A.; Marsh, T.L.; Auras, R. Genome annotation of poly(lactic acid) degrading Pseudomonas aeruginosa, Sphingobacterium sp. and Geobacillus sp. Int. J. Mol. Sci. 2021, 22, 7385. [Google Scholar] [CrossRef]
- Karray, F.; Aloui, F.; Jemli, M.; Mhiri, N.; Loukil, S.; Bouhdida, R.; Sayadi, S. Pilot-scale petroleum refinery wastewaters treatment systems: Performance and microbial communities’ analysis. Process Saf. Environ. Prot. 2020, 141, 73–82. [Google Scholar] [CrossRef]
- Cerqueira, V.S.; Hollenbach, E.B.; Maboni, F.; Vainstein, M.H.; Camargo, F.A.; Maria do Carmo, R.P.; Bento, F.M. Biodegradation potential of oily sludge by pure and mixed bacterial cultures. Bioresour. Technol. 2011, 102, 11003–11010. [Google Scholar] [CrossRef] [Green Version]
- Gargouri, B.; Karray, F.; Mhiri, N.; Aloui, F.; Sayadi, S. Bioremediation of petroleum hydrocarbons-contaminated soil by bacterial consortium isolated from an industrial wastewater treatment plant. J. Chem. Technol. Biotechnol. 2014, 89, 978–987. [Google Scholar] [CrossRef]
- Cerqueira, A.; Souza, P.; Marques, M. Effects of direct and alternating current on the treatment of oily water in an electroflocculation process. Braz. J. Chem. Eng. 2014, 31, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Shankar, S.; Kansrajh, C.; Dinesh, M.; Satyan, R.; Kiruthika, S.; Tharanipriya, A. Application of indigenous microbial consortia in bioremediation of oil-contaminated soils. Int. J. Environ. Sci. Technol. 2014, 11, 367–376. [Google Scholar] [CrossRef]
- Helmy, Q.; Laksmono, R.; Kardena, E. Bioremediation of aged petroleum oil contaminated soil: From laboratory scale to full scale application. Procedia Chem. 2015, 14, 326–333. [Google Scholar] [CrossRef] [Green Version]
- Shen, T.; Pi, Y.; Bao, M.; Xu, N.; Li, Y.; Lu, J. Biodegradation of different petroleum hydrocarbons by free and immobilized microbial consortia. Environ. Sci. Process. Impacts 2015, 17, 2022–2033. [Google Scholar] [CrossRef]
- Maddela, N.R.; Scalvenzi, L.; Venkateswarlu, K. Microbial degradation of total petroleum hydrocarbons in crude oil: A field-scale study at the low-land rainforest of ecuador. Environ. Technol. 2017, 38, 2543–2550. [Google Scholar] [CrossRef]
- Guarino, C.; Spada, V.; Sciarrillo, R. Assessment of three approaches of bioremediation (natural attenuation, landfarming and bioagumentation–assistited landfarming) for a petroleum hydrocarbons contaminated soil. Chemosphere 2017, 170, 10–16. [Google Scholar] [CrossRef]
- Zafra, G.; Absalón, Á.E.; Anducho-Reyes, M.Á.; Fernandez, F.J.; Cortés-Espinosa, D.V. Construction of PAH-degrading mixed microbial consortia by induced selection in soil. Chemosphere 2017, 172, 120–126. [Google Scholar] [CrossRef]
- Tao, K.; Liu, X.; Chen, X.; Hu, X.; Cao, L.; Yuan, X. Biodegradation of crude oil by a defined co-culture of indigenous bacterial consortium and exogenous Bacillus subtilis. Bioresour. Technol. 2017, 224, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; AlMomin, S.; Al-Aqeel, H.; Al-Salameen, F.; Nair, S.; Shajan, A. Metagenomic analysis of rhizosphere microflora of oil-contaminated soil planted with barley and alfalfa. PLoS ONE 2018, 13, e0202127. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Regar, R.K.; Manickam, N. Improved polycyclic aromatic hydrocarbon degradation in a crude oil by individual and a consortium of bacteria. Bioresour. Technol. 2018, 254, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Djahnit, N.; Chernai, S.; Catania, V.; Hamdi, B.; China, B.; Cappello, S.; Quatrini, P. Isolation, characterization and determination of biotechnological potential of oil-degrading bacteria from algerian centre coast. J. Appl. Microbiol. 2019, 126, 780–795. [Google Scholar] [CrossRef]
- Hussain, F.; Hussain, I.; Khan, A.H.A.; Muhammad, Y.S.; Iqbal, M.; Soja, G.; Reichenauer, T.G.; Yousaf, S. Combined application of biochar, compost, and bacterial consortia with Italian ryegrass enhanced phytoremediation of petroleum hydrocarbon contaminated soil. Environ. Exp. Bot. 2018, 153, 80–88. [Google Scholar] [CrossRef]
- Omrani, R.; Spini, G.; Puglisi, E.; Saidane, D. Modulation of microbial consortia enriched from different polluted environments during petroleum biodegradation. Biodegradation 2018, 29, 187–209. [Google Scholar] [CrossRef]
- Piubeli, F.A.; Dos Santos, L.G.; Fernandez, E.N.; Da Silva, F.H.; Durrant, L.R.; Grossman, M.J. The emergence of different functionally equivalent PAH degrading microbial communities from a single soil in liquid PAH enrichment cultures and soil microcosms receiving PAHs with and without bioaugmentation. Pol. J. Microbiol. 2018, 67, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Suganthi, S.H.; Murshid, S.; Sriram, S.; Ramani, K. Enhanced biodegradation of hydrocarbons in petroleum tank bottom oil sludge and characterization of biocatalysts and biosurfactants. J. Environ. Manage. 2018, 220, 87–95. [Google Scholar] [CrossRef]
- Tiralerdpanich, P.; Sonthiphand, P.; Luepromchai, E.; Pinyakong, O.; Pokethitiyook, P. Potential microbial consortium involved in the biodegradation of diesel, hexadecane and phenanthrene in mangrove sediment explored by metagenomics analysis. Mar. Pollut. Bull. 2018, 133, 595–605. [Google Scholar] [CrossRef]
- Lu, C.; Hong, Y.; Liu, J.; Gao, Y.; Ma, Z.; Yang, B.; Ling, W.; Waigi, M.G. A PAH-degrading bacterial community enriched with contaminated agricultural soil and its utility for microbial bioremediation. Environ. Pollut. 2019, 251, 773–782. [Google Scholar] [CrossRef]
- Woźniak-Karczewska, M.; Lisiecki, P.; Białas, W.; Owsianiak, M.; Piotrowska-Cyplik, A.; Wolko, Ł.; Ławniczak, Ł.; Heipieper, H.J.; Gutierrez, T.; Chrzanowski, Ł. Effect of bioaugmentation on long-term biodegradation of diesel/biodiesel blends in soil microcosms. Sci. Total Environ. 2019, 671, 948–958. [Google Scholar] [CrossRef]
- Barbato, M.; Mapelli, F.; Crotti, E.; Daffonchio, D.; Borin, S. Cultivable hydrocarbon degrading bacteria have low phylogenetic diversity but highly versatile functional potential. Int. Biodeterior. Biodegrad. 2019, 142, 43–51. [Google Scholar] [CrossRef]
- Cavalcanti, T.G.; de Souza, A.F.; Ferreira, G.F.; Dias, D.S.B.; Severino, L.S.; Morais, J.P.S.; de Sousa, K.A.; Vasconcelos, U. Use of agro-industrial waste in the removal of phenanthrene and pyrene by microbial consortia in soil. Waste Biomass Valorization 2019, 10, 205–214. [Google Scholar] [CrossRef]
- Xia, M.; Fu, D.; Chakraborty, R.; Singh, R.P.; Terry, N. Enhanced crude oil depletion by constructed bacterial consortium comprising bioemulsifier producer and petroleum hydrocarbon degraders. Bioresour. Technol. 2019, 282, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Liu, G.; Chen, T.; Li, S.; An, L.; Zhang, G.; Li, G.; Chang, S.; Zhang, W.; Chen, X. Characterization of the genome of a nocardia strain isolated from soils in the qinghai-tibetan plateau that specifically degrades crude oil and of this biodegradation. Genomics 2019, 111, 356–366. [Google Scholar] [CrossRef]
- Diallo, M.; Vural, C.; Cay, H.; Ozdemir, G. Enhanced biodegradation of crude oil in soil by a developed bacterial consortium and indigenous plant growth promoting bacteria. J. Appl. Microbiol. 2021, 130, 1192–1207. [Google Scholar] [CrossRef]
- Milić, J.; Avdalović, J.; Šolević Knudsen, T.; Gojgić-Cvijović, G.; Jednak, T.; Vrvić, M. Initial microbial degradation of polycyclic aromatic hydrocarbons. Chem. Ind. Chem. Eng. Q. 2016, 22, 293–299. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Parameters | Uncontaminated Soil | Contaminated Soil |
|---|---|---|
| Texture | Sandy loam (sand 53%, silt 25% and clay 22%) | Sandy loam (sand 73%, silt 08% and clay 19%) |
| pH | 7.95 ± 0.17 | 8.2 ± 0.24 |
| EC (dS m−1) | 1.07 ± 0.28 | 1.45 ± 0.21 |
| CEC (mmolc L−1) | 16.3 ± 1.43 | 14.3 ± 1.73 |
| Organic matter (%) | 0.73 ± 0.05 | 0.47 ± 0.03 |
| Total N (mg kg−1) | 179.8 ± 3.20 | 91.8 ± 8.42 |
| Available P (mg kg−1) | 86.7 ± 1.09 | 32.7 ± 3.49 |
| Available K (mg kg−1) | 58.4 ± 2.35 | 39.5 ± 2.05 |
| PHCs (%) | ND | 1.2 ± 0.04 |
| Treatments | Detail |
|---|---|
| Control | Uncontaminated soil with chickpea plants but without bacterial consortium |
| PHCs | Hydrocarbons contaminated soil without chickpea plants and microbes |
| PHCs + CP | Hydrocarbons contaminated soil with chickpea plants but without bacterial consortium |
| CP + BC1 | Uncontaminated soil with chickpea plants and bacterial consortium 1 |
| PHCs + BC1 | Hydrocarbons contaminated soil with bacterial consortium 1 but without chickpea plants |
| PHCs + CP + BC1 | Hydrocarbons contaminated soil with chickpea plants and bacterial consortium 1 |
| CP + BC2 | Uncontaminated soil with chickpea plants and bacterial consortium 2 |
| PHCs + BC2 | Hydrocarbons contaminated soil with bacterial consortium 2 but without chickpea plants |
| PHCs + CP + BC2 | Hydrocarbons contaminated soil with chickpea plants and bacterial consortium 2 |
| Treatments | Seedling Emergence (%) | ||||
|---|---|---|---|---|---|
| 3 DAS | 4 DAS | 5 DAS | 6 DAS | 7 DAS | |
| Control | 44.4 ± 5.57 ab | 66.7 ± 9.62 ab | 77.8 ± 5.54 ab | 83.3 ± 9.62 ab | 83.3 ± 9.62 ab |
| PHCs + CP | 11.1 ± 5.57 c | 38.9 ± 5.57 c | 55.6 ± 5.57 c | 55.6 ± 5.57 c | 55.6 ± 5.57 c |
| CP + BC1 | 50.1 ± 9.59 a | 72.2 ± 5.53 a | 83.3 ± 9.62 a | 88.9 ± 5.57 ab | 88.9 ± 5.57 ab |
| PHCs + CP + BC1 | 22.2 ± 5.53 c | 44.4 ± 5.57 c | 61.1 ± 5.57 bc | 72.2 ± 5.54 bc | 72.2 ± 5.54 bc |
| CP + BC2 | 55.6 ± 5.57 a | 72.2 ± 5.54 a | 88.9 ± 5.57 a | 94.4 ± 5.57 a | 94.4 ± 5.57 a |
| PHCs + CP + BC2 | 27.8 ± 5.53 bc | 50.0 ± 9.65 bc | 72.2 ± 5.54 abc | 77.8 ± 5.54 ab | 77.8 ± 5.54 ab |
| Treatments | SL (cm) | RL (cm) | NN | NP | NS | NCL |
|---|---|---|---|---|---|---|
| Control | 36.3 ± 0.88 b | 21.3 ± 0.88 bc | 10.7 ± 0.73 c | 20.33 ± 0.88 b | 3.33 ± 0.33 ab | 71.7 ± 4.92 c |
| PHCs + CP | 28.7 ± 1.20 c | 16.0 ± 0.58 d | 7.67 ± 0.60 e | 15.0 ± 0.58 c | 2.67 ± 0.33 b | 51.0 ± 4.17 d |
| CP + BC1 | 47.3 ± 2.03 a | 26.3 ± 2.61 ab | 12.3 ± 0.33 b | 23.0 ± 1.16 ab | 3.67 ± 0.33 ab | 87.0 ± 5.29 b |
| PHCs + CP + BC1 | 32.3 ± 0.88 bc | 17.3 ± 0.33 cd | 8.67 ± 0.29 de | 16.3 ± 0.68 bc | 3.00 ± 0.58 b | 57.3 ± 2.97 d |
| CP + BC2 | 51.3 ± 3.93 a | 31.0 ± 2.31 a | 14.3 ± 0.33 a | 25.7 ± 0.88 a | 4.33 ± 0.33 a | 101 ± 3.85 a |
| PHCs + CP + BC2 | 34.7 ± 0.88 bc | 18.7 ± 1.20 cd | 9.33 ± 0.33 cd | 17.3 ± 0.33 c | 3.33 ± 0.33 ab | 62.3 ± 1.77 cd |
| Treatments | CC (SPAD Value) | Ft (µmolm−2 s−1) | PAR (µmolm−2 s−1) | YII (µmolm−2 s−1) | ETR (µmolm−2 s−1) |
|---|---|---|---|---|---|
| Control | 41.3 ± 0.89 b | 232.7 ± 23.1 cd | 746 ± 20.1 a | 0.74 ± 0.04 bc | 50.1 ± 2.98 c |
| PHCs + CP | 31.7 ± 1.41 c | 173.0 ± 22.3 d | 550 ± 22.4 c | 0.59 ± 0.03 d | 43.7 ± 2.02 c |
| CP + BC1 | 46.4 ± 0.40 ba | 320.3 ± 22.3 b | 777 ± 18.5 a | 0.80 ± 0.01 ab | 61.3 ± 4.87 ab |
| PHCs + CP + BC1 | 38.5 ± 1.43 b | 230.3 ± 10.4 cd | 618 ± 13.9 b | 0.68 ± 0.02 cd | 48.1 ± 1.95 c |
| CP + BC2 | 49.7 ± 1.02 a | 392 ± 15.9 a | 783 ± 21.2 a | 0.88 ± 0.02 a | 70.0 ± 2.66 a |
| PHCs + CP + BC2 | 40.9 ± 0.84 b | 268 ± 8.97 bc | 644 ± 11.2 b | 0.72 ± 0.01 bc | 51.3 ± 2.43 bc |
| Treatments | PC (µmole g−1 FW) | CAT (U mg−1 Protein FW) | SOD (U mg−1 Protein FW) | POD (U mg−1 Protein FW) |
|---|---|---|---|---|
| Control | 23.7 ± 1.34 d | 4.29 ± 0.32 b | 7.44 ± 0.27 c | 69.3 ± 2.60 c |
| PHCs + CP | 35.0 ± 1.88 a | 5.89 ± 0.56 a | 11.3 ± 0.56 a | 88.1 ± 3.52 a |
| CP + BC1 | 25.7 ± 1.59 cd | 4.85 ± 0.21 b | 7.78 ± 0.58 c | 76.7 ± 1.70 bc |
| PHCs + CP + BC1 | 30.2 ± 1.71 abc | 5.21 ± 0.32 ab | 10.6 ± 1.12 ab | 78.7 ± 3.33 b |
| CP + BC2 | 26.5 ± 2.11 bcd | 5.09 ± 0.08 ab | 8.58 ± 0.73 bc | 77.8 ± 2.32 bc |
| PHCs + CP + BC2 | 30.9 ± 1.02 ab | 5.29 ± 0.15 ab | 9.81 ± 0.20 ab | 74.4 ± 1.19 b |
| Treatments | Nutrient Uptake (%) by Plants | ||
|---|---|---|---|
| Nitrogen | Phosphorus | Potassium | |
| Control | 0.025 ± 0.001 b | 0.015 ± 0.001 ab | 0.015 ± 0.001 b |
| PHCs + CP | 0.017 ± 0.001 c | 0.012 ± 0.001 c | 0.012 ± 0.001 c |
| CP + BC1 | 0.031 ± 0.001 a | 0.017 ± 0.001 ab | 0.020 ± 0.001 a |
| PHCs + CP + BC1 | 0.021 ± 0.001 c | 0.014 ± 0.001 bc | 0.015 ± 0.001 b |
| CP + BC2 | 0.034 ± 0.001 a | 0.017 ± 0.001 a | 0.021 ± 0.001 a |
| PHCs + CP + BC2 | 0.022 ± 0.001 bc | 0.014 ± 0.001 bc | 0.016 ± 0.001 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.H.; Khan, M.I.; Naveed, M.; Tanvir, M.A. Microbe-Assisted Rhizoremediation of Hydrocarbons and Growth Promotion of Chickpea Plants in Petroleum Hydrocarbons-Contaminated Soil. Sustainability 2023, 15, 6081. https://doi.org/10.3390/su15076081
Ali MH, Khan MI, Naveed M, Tanvir MA. Microbe-Assisted Rhizoremediation of Hydrocarbons and Growth Promotion of Chickpea Plants in Petroleum Hydrocarbons-Contaminated Soil. Sustainability. 2023; 15(7):6081. https://doi.org/10.3390/su15076081
Chicago/Turabian StyleAli, Muhammad Hayder, Muhammad Imran Khan, Muhammad Naveed, and Muhammad Ayyoub Tanvir. 2023. "Microbe-Assisted Rhizoremediation of Hydrocarbons and Growth Promotion of Chickpea Plants in Petroleum Hydrocarbons-Contaminated Soil" Sustainability 15, no. 7: 6081. https://doi.org/10.3390/su15076081