
Introducing the Power of Plant Growth Promoting Microorganisms in Soilless Systems: A Promising Alternative for Sustainable Agriculture





Abstract
:
1. Introduction
2. Categorization of Studies
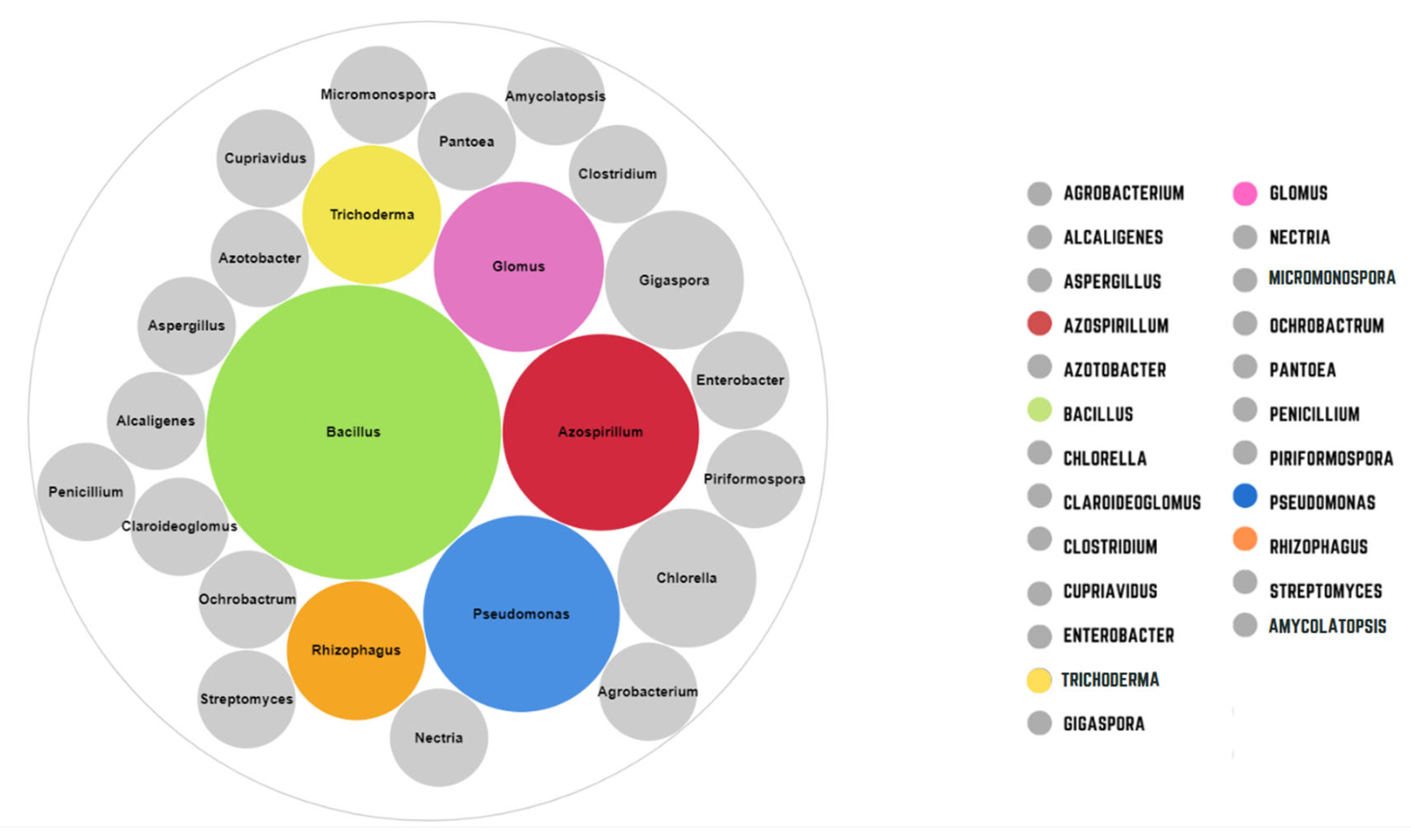
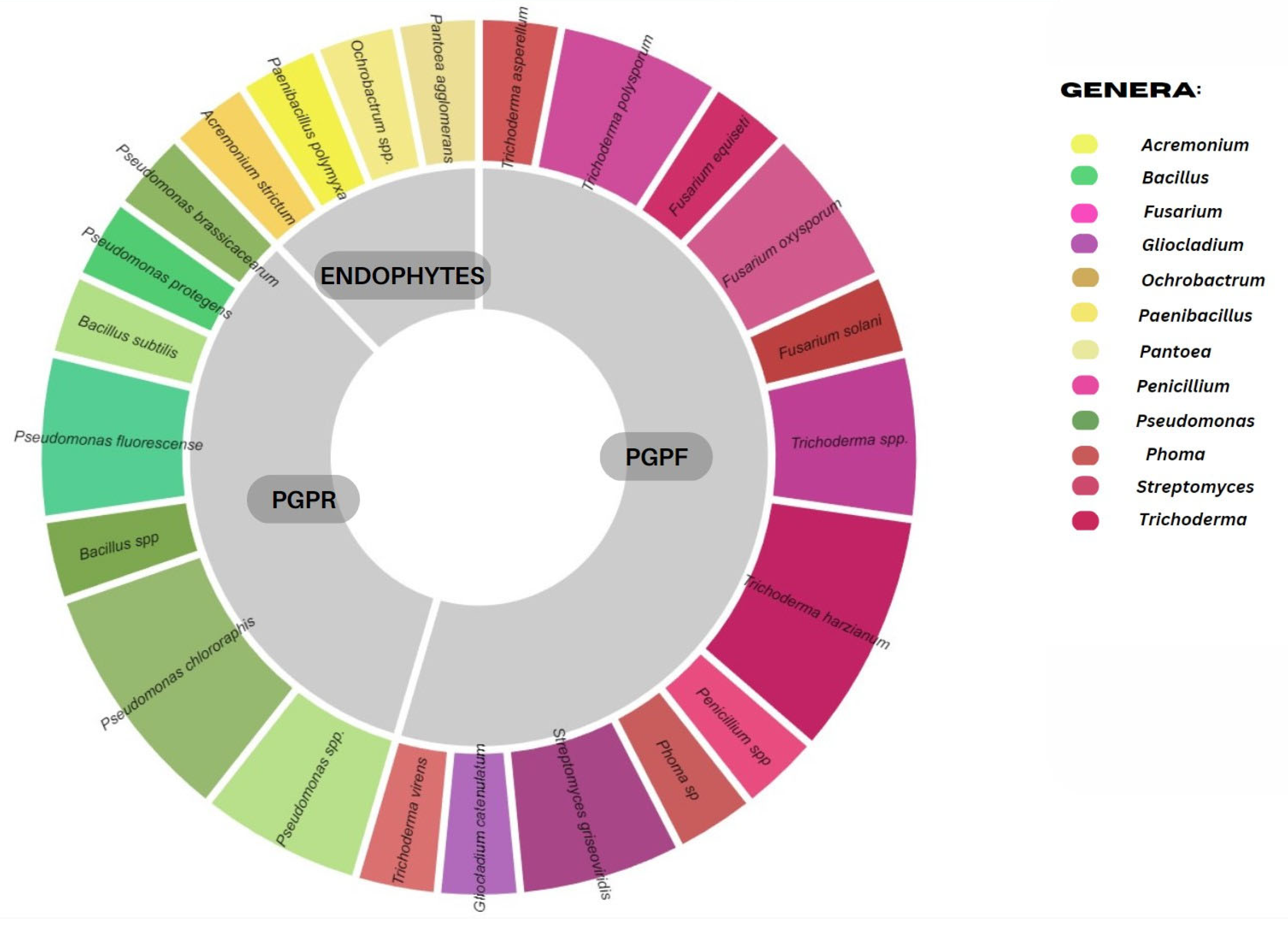
3. PGPM as Plant-Promoting Growth Treatment and Facilitation of Nutrient Uptake

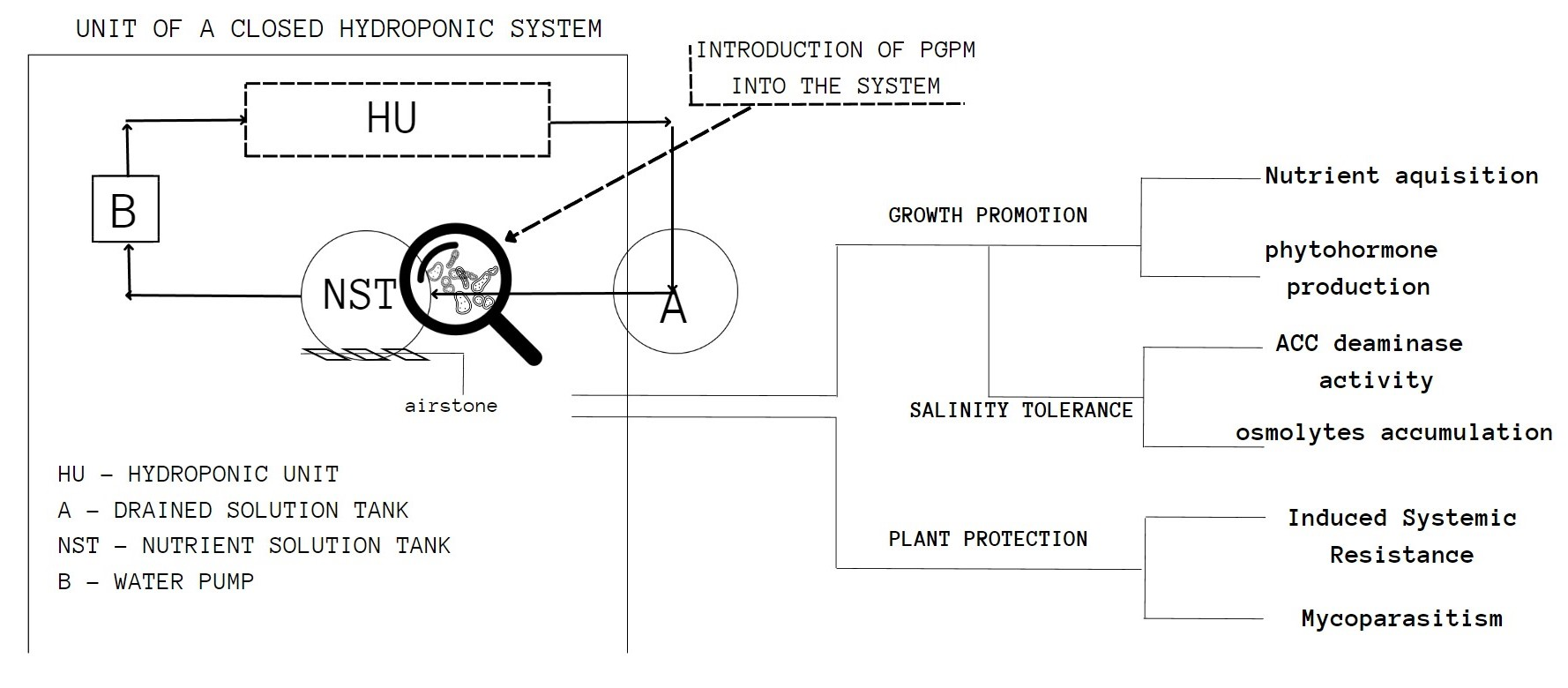{kind=link}
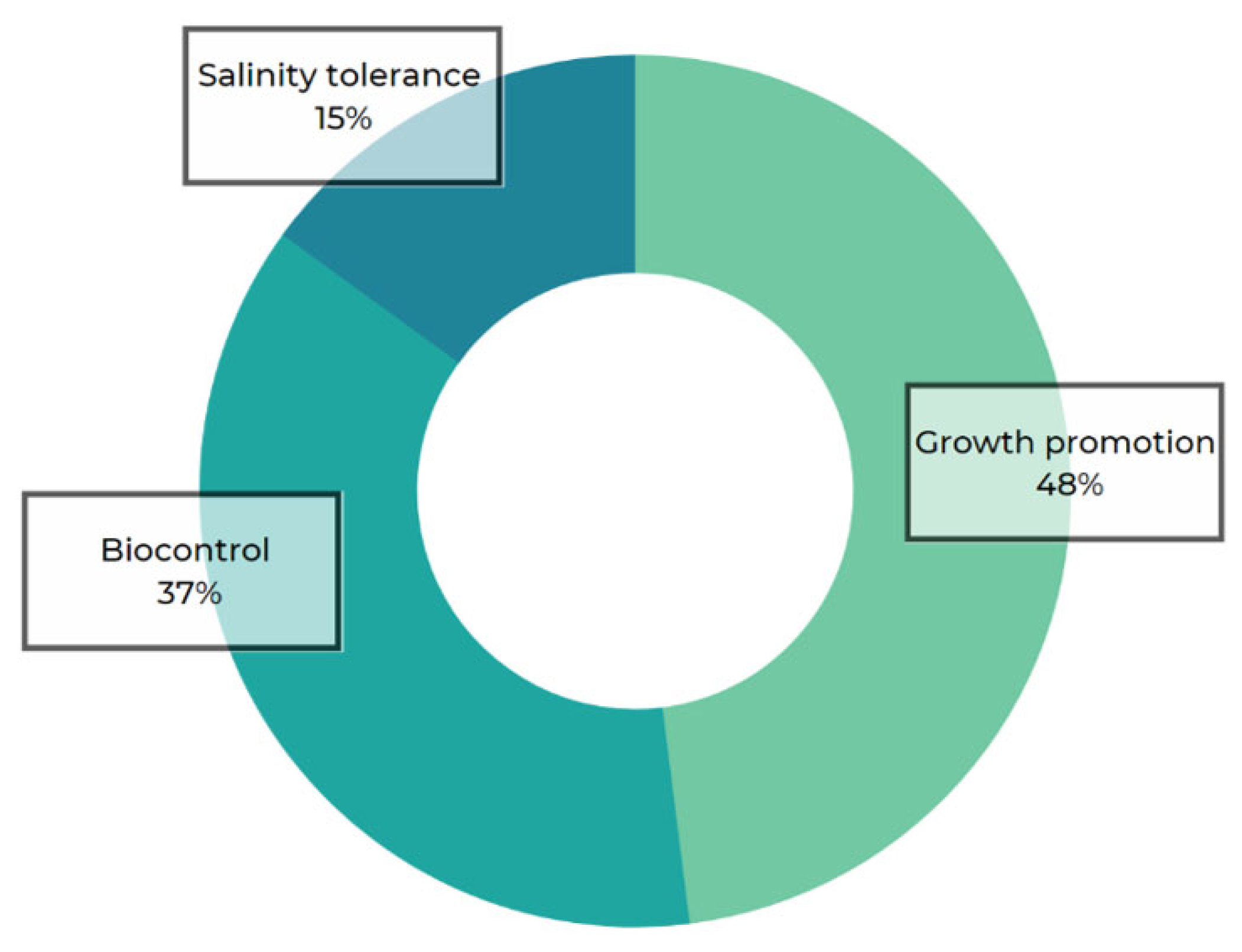{kind=link}
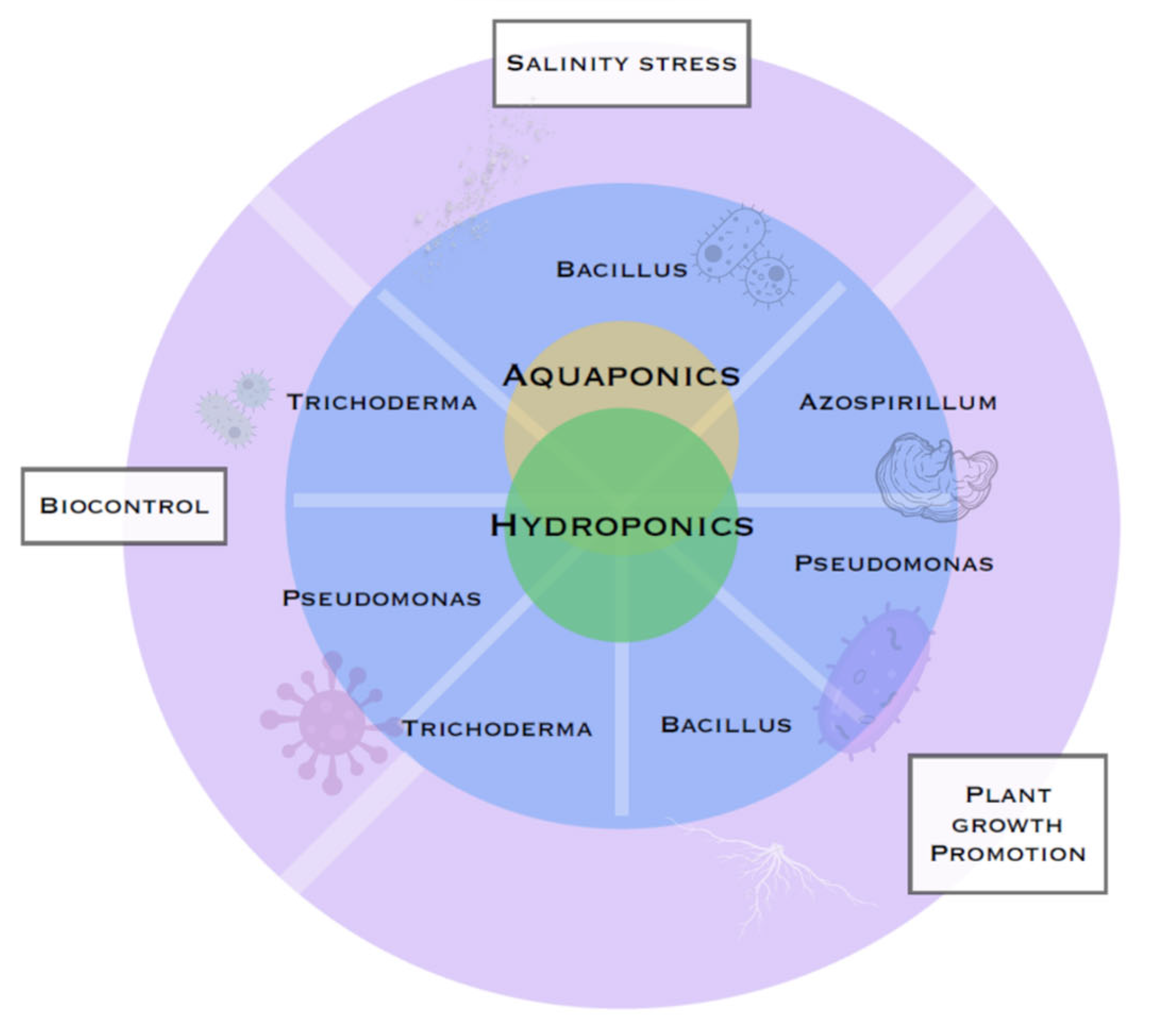{kind=link}
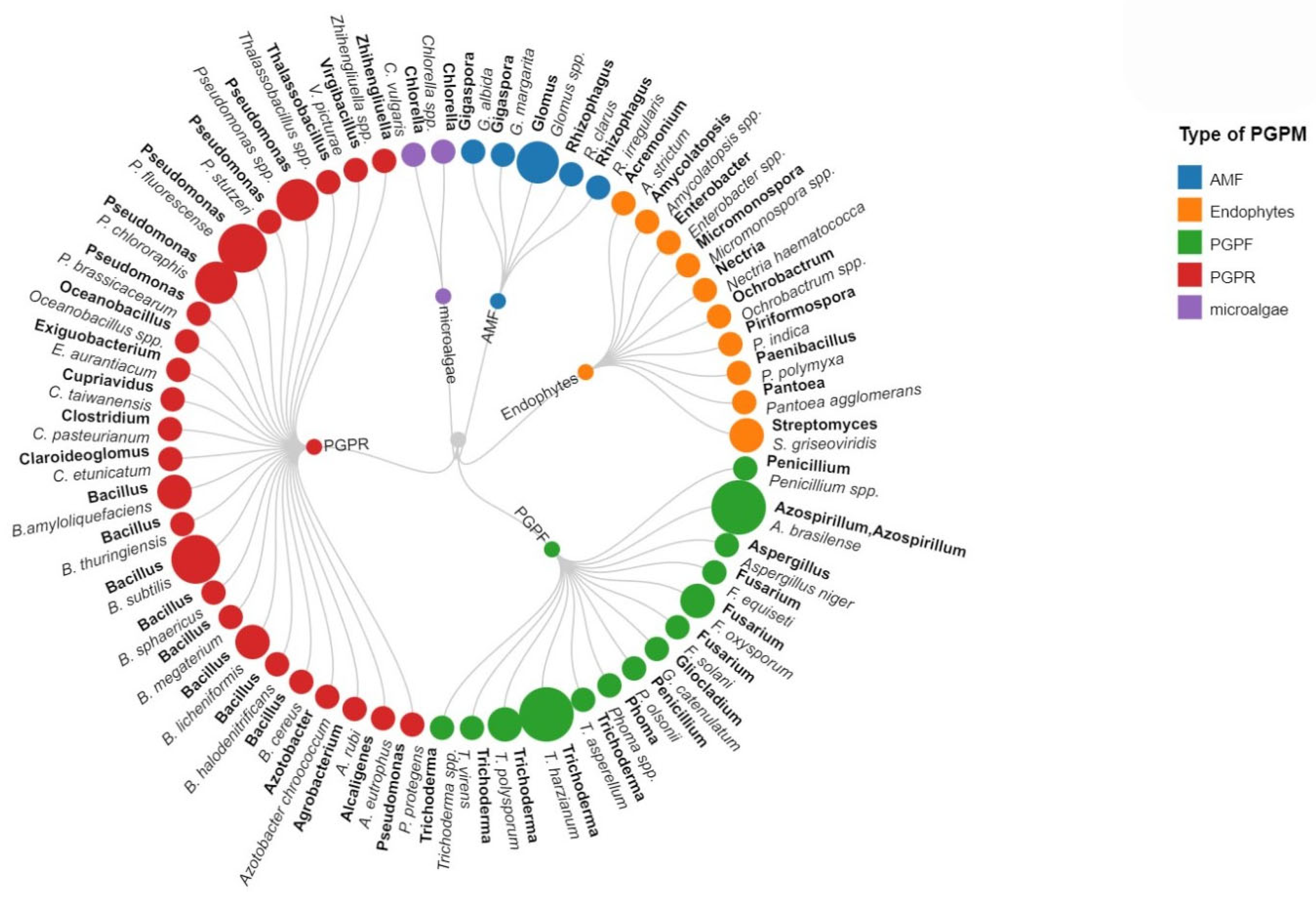{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of PGPM | Species | Mechanism | Crop | Application | System | Reference |
|---|---|---|---|---|---|---|
| PGPF | Azospirillum brasilense | auxin-IAA -> alter root morphology | basil | seedling | aquaponics, hydroponic gullies | [24] |
| PGPF | Azospirillum brasilense | elicited terpenoids and phenylpropanoids + synthesizing phytohormones | basil | NS | liquid medium | [25] |
| PGPF | Pseudomonas spp. | siderophore and IAA-like compounds production | apple * | NS | liquid medium | [29] |
| PGPF | Trichoderma harzianum, Azospirillum brasilense | increased leaf chlorophyll content, shoot and root growth, N acquisition | lettuce | foliar | NFT | [27] |
| Endophyte | Ochrobactrum spp., Pantoea agglomerans | secondary metabolite production, increasing iron uptake | cucumber ** | seed coating | drip irrigation system, peat | [30] |
| Endophyte | Nectria haematococca | production of growth hormones, enhanced P content | mung bean | seeds | vermiculite | [31] |
| Endophyte | Enterobacter spp. | enhanced biomass, root growth, superoxide dismutase (SOD), catalase (CAT) content | pak choi | roots | liquid medium | [32] |
| Endophyte | Streptomyces spp., Amycolatopsis spp., Micromonospora spp. | IAA, NH3, Siderophore production, P solubilization | wheat | seedlings | liquid medium | [33] |
| AMF | Glomus spp., Gigaspora margarita EndoRoots Soluble® | improved photosynthesis, better supply of water and nutrients to the fruits | melon * | NS | cocopeat and perlite | [34] |
| AMF | Rhizophagus irregularis MycoPlant® | increased N, P content, seedling nutrient uptake, higher photosynthetic ability | tomato | NS | floating system | [35] |
| PGPR + AMF | Azotobacter chroococcum, Azospirillum brasilense, Pseudomonas flourescens, Bacillus subtilis, Aspergillus niger, Glomus spp. | increased efficiency of water translocation process, mineralization and solubilization of nutrients, production of phytohormones | tomato | roots | drip irrigation system, mix of rice straw charcoal, sand planting medium | [36] |
| PGPR, endophyte | Bacillus cereus, Bacillus thuringiensis, Buttiauxella agrestis | improved N content, extraction of nutrients from NS | banana | NS | floating system | [37] |
| endophyte + AMF | Piriformospora indica, Rhizophagus clarus, Claroideoglomus etunicatum, Gigaspora albida | affected mechanisms of absorption, translocation, and redistribution of nutrient | tomato *** | substrate | drip irrigation system, coconut fiber | [38] |
| AMF, Microalgae, PGPR | Glomus spp. EndoRoots Soluble®, Chlorella spp. Allgrow®, Clostridium pasteurianum Bio-one® | inoculation reduced effect of low nutrients supply | squash * | NS | perlite and cocopeat | [39] |
| PGPR(F) | Bacillus subtilis, Trichoderma harzianum | improved nutrient uptake | tomato | NS | perlite (open and close HS) | [40] |
| PGPR | Bacillus megaterium, Agrobacterium rubi, Alcaligenes eutrophus | mitigating effects on alkaline conditions | grapevine *** | NS | peat and perlite | [41] |
| PGPR | Pseudomonas stutzeri, Cupriavidus taiwanensis | production of antioxidant enzymes, higher root sulfide content, production of metal-binding peptides | rice *** | seedlings | liquid medium | [42] |
| PGPR | Bacillus sphaericus, Azospirillum spp. | increased chlorophyll content, N concentration, secretion of auxins, gibberellins and cytokinins | banana * | seedlings | liquid medium | [26] |
| PGPR | Bacillus subtilis, Bacillus licheniformis | solubilization of minerals, nitrogen fixation, enhanced concentrations of K, Na, P, and Zn | lettuce | NS | aquaponics, deep water culture | [23] |
| PGPR | Pseudomonas chlororaphis | upregulation of genes related to plant growth | lettuce ** | roots | window farm, clay pellets | [43] |
| PGPR | Bacillus subtilis Sanolife® Pro-W | fish had higher activity of the digestive enzymes; production of endospores | mint | sump tank | recirculating aquaponics (Nile tilapia), HS-deep water culture | [44] |
| microalgae | Chlorella vulgaris | production of phytohormones-polyamines, betaines, brassinosteroids, and secretion of chelates; increased soluble solids (brix) and vitamin C | lettuce * | NS | liquid medium | [45] |
4. PGPM for Alleviation of Salinity Stress
| Species | Mechanism | Crop | Growing Media | Reference |
|---|---|---|---|---|
| Bacillus subtillis | reduced synthesis of ethylene hormone; biophysics of the photosynthetic electron transport system | lettuce * | perlite | [53] |
| Bacillus subtillis FZB24® WG | resistance-inducing metabolites, production of lipopolysaccharides, enzymes, siderophores | tomato ** | coco peat | [56] |
| Bacillus spp., Zhihengliuella spp., Exiguobacterium aurantiacum, Virgibacillus picturae, Oceanobacillus spp., Thalassobacillus spp. | IAA production, N fixation, NH3 production, P solubilization, ACC deaminase activity, accumulation of osmolytes | wheat | liquid medium | [55] |
| Pseudomonas fluorescens | upregulated protein expression levels, ACC deaminase | canola | liquid medium | [57] |
| Bacillus amyloliquefaciens | enhanced nutrient uptake, stress alleviation, enhanced proline accumulation | rice | liquid medium | [12] |
| Bacillus amyloliquefaciens, B. brevis, B. circulans, B. coagulans, B. firmus, B. halodenitrificans B. laterosporus, B. licheniformis, B. megaterium, B. mycoides, B. pasteurii, B. subtilis TNC BactorrS13® | changes in phytohormone content, antioxidant defense, osmolyte production, ACC | lettuce | liquid medium | [54] |
| Penicillium olsonii | IAA production induced the catalase (CAT) and the superoxide dismutase (SOD) activities, enhanced chlorophyll, proline content | tobacco plants | liquid medium | [58] |
5. PGPM as Biocontrol Agents
| BCA Species | Mechanism | Plant Disease | Crop | Application | System | Ref. |
|---|---|---|---|---|---|---|
| Trichoderma asperellum | mycoparasitism | Cercospora hypha (Leaf spot) | lettuce | foliar | NFT | [60] |
| Fusarium oxysporum | antagonistic activity, ISR, production of enzymes(chitinase) | F. oxysporum f. sp. Cucumerinum (wilt) | cucumber | roots | liquid medium | [61] |
| Trichoderma polysporum, Trichoderma harzianum Binab T® | competition for niche and nutrients, ISR | Ralstonia solanacearum (bacterial wilt) | tomato | seedlings | liquid medium | [62] |
| Fusarium equiseti, Trichoderma spp., Penicillium spp., Phoma sp. | inhibited the spore germination, ISR, direct antagonism | Fusarium oxysporum (crown and root rot) | tomato | seeds | rockwool, sub-irrigation system | [63] |
| Trichoderma polysporum, Trichoderma harzianum, Streptomyces griseoviridis | antagonistic activity | Pythium aphanidermatum, Phytophthora cryptogea | tomato | seedlings | pumice, peat; ebb and flow | [64] |
| Fusarium oxysporum | direct antagonism, ISR | Curvularia lunata, Rhizoctonia solani, F. oxysporum f. sp. lactucae | lettuce | roots | Deep Flow Technique | [65] |
| Pseudomonas sp., Fusarium solani. Trichoderma sp. | mycoparasitism, nutrient competition, ISR | Phytophthora capsici (crown rot) | zucchini | NS | peat | [13] |
| Pseudomonas chlororaphis, Trichoderma harzianum RootShield® Drench, Streptomyces griseoviridis Mycostop®, Gliocladium catenulatum Prestop® WP, Trichoderma virens SoilGard® | outcompeting the pathogen for niche, nutrients; reduced spore germination | Fusarium oxysporum (root and stem rot) | cucumber | seeds | peat, rockwool | [66] |
| Pseudomonas spp., Pseudomonas protegens, Pseudomonas brassicacearum | production of antibiotics, phenazines, pyrrolnitrin, and/or pyoluteorin; ISR | Agrobacterium rhizogenes (hairy root disease) | tomato | roots | rockwool | [67] |
| Bacillus spp. | production of siderophores, protease, glucanase; ISR, induced SOD activity | Pyricularia oryzae (blast disease) | rice | NS | liquid medium | [68] |
| Pseudomonas chlororaphis, Pseudomonas fluorescens | antibiosis, ISR | Fusarium oxysporum (root rot) | tomato | seeds | liquid medium | [69] |
| Pseudomonas fluorescens | antagonistic activity-> reduction in severity of disease | Pythium ultimum (rot) | tomato | NS | NFT | [70] |
| Bacillus subtilis FZB24® WG | resistance-inducing metabolites, production of lipopolysaccharides, enzymes, siderophores | salinity, Pythium, B. tabaci | tomato | NS | coco peat | [56] |
| Pseudomonas chlororaphis | ISR, upregulation of protein involved in the jasmonic acid biosynthesis pathway | Pythium ultimum | lettuce | roots | window farm, clay pellets | [42] |
| Paenibacillus polymyxa | antagonistic activity, ISR | Cercospora sp. (leaf spot) | lettuce | NS | NFT | [71] |
| Acremonium strictum | antagonistic activity-> reduction in severity of disease | Phytophthora cactorum | strawberry | substrate | peat | [72] |
| Ochrobactrum spp., Pantoea agglomerans | colonization in all tissues provided sustainable antagonistic effects, ISR | angular leaf spot disease | cucumber | seed coating | drip irrigation system, peat | [30] |
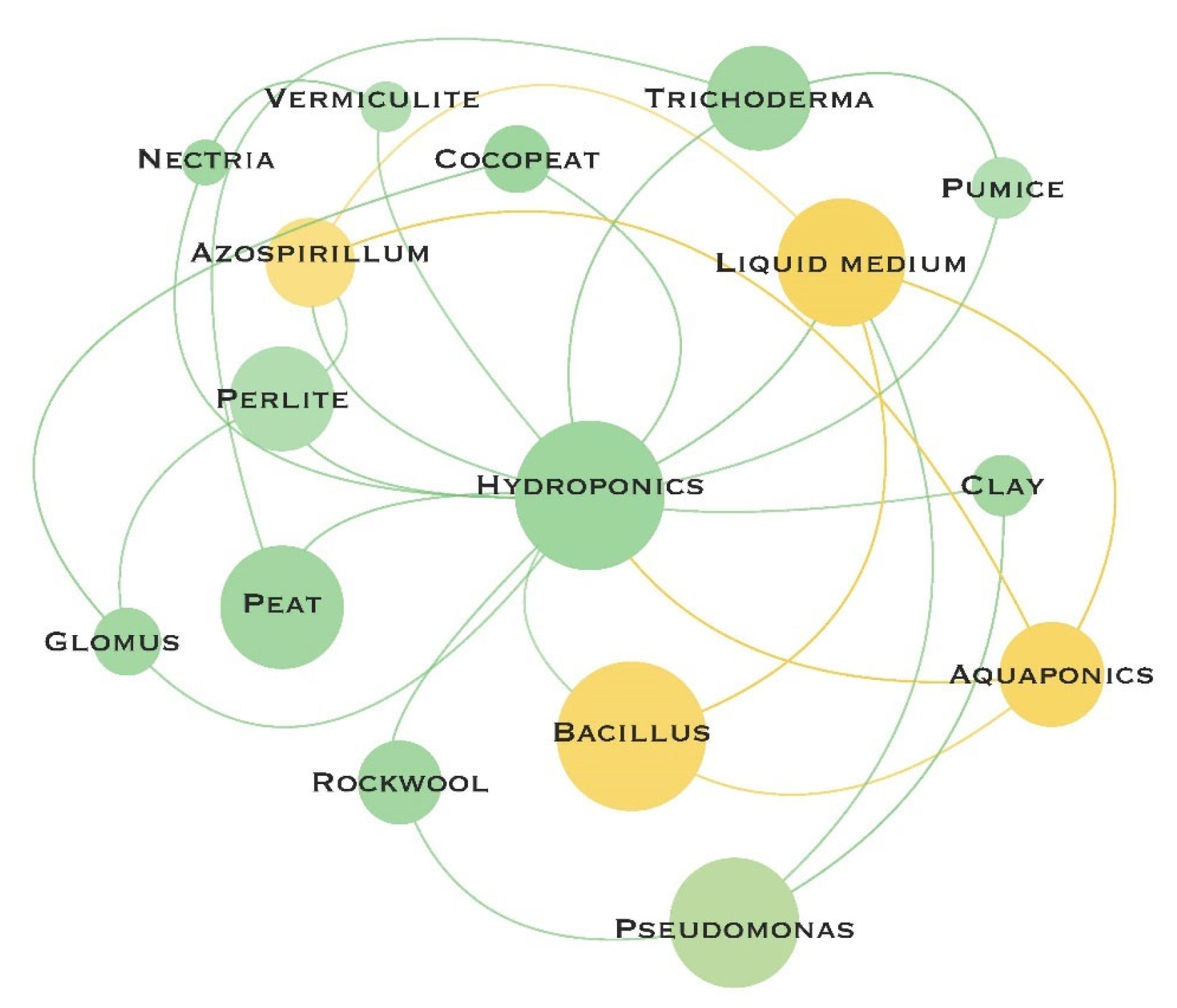
6. Substrate-PGPM Specificity
7. Concluding Remarks and Future Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. The Future of Food and Agriculture: Alternative Pathways to 2050; FAO: Rome, Italy, 2020. [Google Scholar]
- Magwaza, S.T.; Magwaza, L.S.; Odindo, A.O.; Mditshwa, A. Hydroponic Technology as Decentralised System for Do-mestic Wastewater Treatment and Vegetable Production in Urban Agriculture: A Review. Sci. Total Environ. 2020, 698, 134154. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Cáceres, G.P.; Pérez-Urrestarazu, L.; Avilés, M.; Borrero, C.; Eguíbar, J.R.L.; Fernández-Cabanás, V.M. Susceptibility to water-borne plant diseases of hydroponic vs. aquaponics systems. Aquaculture 2021, 544, 737093. [Google Scholar] [CrossRef]
- Sambo, P.; Nicoletto, C.; Giro, A.; Pii, Y.; Valentinuzzi, F.; Mimmo, T.; Lugli, P.; Orzes, G.; Mazzetto, F.; Astolfi, S.; et al. Hydroponic Solutions for Soilless Production Systems: Issues and Opportunities in a Smart Agriculture Perspective. Front. Plant Sci. 2019, 10, 923. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Asensio, J.S.; Parra, M.; Intrigliolo, D.S. Chapter 30—Open Field Hydroponics in Fruit Crops: Developments and Challenges. In Fruit Crops; Srivastava, A.K., Hu, C., Eds.; Elsevier: Amsterdam, Netherlands, 2020; pp. 419–430. [Google Scholar]
- Prakash, O.; Kumar, V.; Kumar, A.; Singh, R. Hydroponic System for Vegetable Production: A Review. Int. J. Environ. Sci. Technol. 2020, 17, 364–371. [Google Scholar]
- Sathyanarayan, S.; Rajkumar, M.; Anandham, R.; Sa, T.M. Plant Growth-Promoting Rhizobacteria (PGPR) and Their Modes of Action in Hydroponic Systems. Microorganisms 2022, 10, 147–203. [Google Scholar]
- Pinstrup-Andersen, P. Is it time to take vertical indoor farming seriously? Glob. Food Secur. 2018, 17, 233–235. [Google Scholar] [CrossRef]
- Bartelme, R.P.; Oyserman, B.O.; Blom, J.E.; Sepulveda-Villet, O.J.; Newton, R.J. Stripping Away the Soil: Plant Growth Promoting Microbiology Opportunities in Aquaponics. Front. Microbiol. 2018, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Rajalakshmi, M.; Manoj, V.R.; Manoj, H. Comprehensive Review of Aquaponic, Hydroponic, and Recirculating Aquaculture Systems. J. Exp. Biol. Agric. Sci. 2022, 10, 1266–1289. [Google Scholar] [CrossRef]
- Pattillo, D.; Cline, D.; Hager, J.; Roy, L.; Hanson, T. Challenges Experienced by Aquaponic Hobbyists, Producers, and Educators. J. Ext. 2022, 60. [Google Scholar] [CrossRef]
- Nautiyal, C.S.; Chauhan, P.S.; DasGupta, S. Plant Growth-Promoting Bacteria: Potential Tools for Salinity Stress Tolerance in Plants. In Microorganisms for Green Revolution 2013; Springer: New Delhi, India, 2013; pp. 45–60. [Google Scholar]
- Gilardi, G.; Pugliese, M.; Gullino, M.L.; Garibaldi, A. Occurrence and Efficacy of Fungicides against Soil-Borne Pathogens in Hydroponic Systems: A Review. Appl. Sci. 2020, 10, 8677. [Google Scholar]
- Ray, P.; Lakshmanan, V.; Labbé, J.L.; Craven, K.D. Microbe to Microbiome: A Paradigm Shift in the Application of Microorganisms for Sustainable Agriculture. Front. Microbiol. 2020, 11, 622926. [Google Scholar] [CrossRef] [PubMed]
- MarketsandMarkets. Compound Biofertilizers & Customized Fertilizers Market by Type (Nitrogen-Fixing, Phosphorus Solubilizing & Mobilizing, Potassium Solubilizing & Mobilizing), Application (Seed Treatment, Soil Treatment), Form (Solid, Liquid), Crop Type, and Region—Global Forecast to 2027; MarketsandMarkets: Maharashtra, India, 2022. [Google Scholar]
- Fadiji, A.E.; Babalola, O.O.; Santoyo, G.; Perazzolli, M. The Potential Role of Microbial Biostimulants in the Amelioration of Climate Change-Associated Abiotic Stresses on Crops. Front. Microbiol. 2022, 12, 4392. [Google Scholar] [CrossRef]
- Woitke, M.; Schitzler, w. Biotic stress relief on plants in hydroponic systems. Acta Hortic. 2005, 697, 557–565. [Google Scholar] [CrossRef]
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddique, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere Engineering with Plant Growth-Promoting Microorganisms for Agriculture and Ecological Sustainability. Front. Sustain. Food Syst. 2021, 5, 617157. [Google Scholar] [CrossRef]
- Chiaranunt, P.; White, J.F. Plant Beneficial Bacteria and Their Potential Applications in Vertical Farming Systems. Plants 2023, 12, 400. [Google Scholar] [CrossRef] [PubMed]
- da Silva Cerozi, B.; Fitzsimmons, K. Use of Bacillus spp. to enhance phosphorus availability and serve as a plant growth promoter in aquaponics systems. Sci. Hortic. 2016, 211, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [Green Version]
- Riaz, U.; Murtaza, G.; Anum, W.; Samreen, T.; Sarfraz, M.; Nazir, M.Z. Plant Growth-Promoting Rhizobacteria (PGPR) as Biofertilizers and Biopesticides. In Microbiota and Biofertilizers: A Sustainable Continuum for Plant and Soil Health; Springer International Publishing: Cham, Switzerland, 2021; pp. 181–196. [Google Scholar] [CrossRef]
- Kasozi, N.; Abraham, B.; Kaiser, H.; Wilhelmi, B. The complex microbiome in aquaponics: Significance of the bacterial ecosystem. Ann. Microbiol. 2021, 71, 1–13. [Google Scholar] [CrossRef]
- Mangmang, J.S.; Deaker, R.; Rogers, G. Response of Cucumber Seedlings Fertilized with Fish Effluent to Azospirillum brasilense. Int. J. Veg. Sci. 2016, 22, 129–140. [Google Scholar] [CrossRef]
- Kolega, S.; Miras-Moreno, B.; Buffagni, V.; Lucini, L.; Valentinuzzi, F.; Maver, M.; Mimmo, T.; Trevisan, M.; Pii, Y.; Cesco, S. Nutraceutical Profiles of Two Hydroponically Grown Sweet Basil Cultivars as Affected by the Composition of the Nutrient Solution and the Inoculation with Azospirillum brasilense. Front. Plant Sci. 2020, 11, 596000. [Google Scholar] [CrossRef]
- Mia, M.A.; Shamsuddin, Z.H.; Wahab, Z.; Marziah, M. Effect of plant growth promoting rhizobacterial (PGPR) inoculation on growth and nitrogen incorporation of tissue-cultured Musa plantlets under nitrogen-free hydroponics condition. Aust. J. Crop Sci. 2010, 4, 85–90. [Google Scholar]
- Moreira, V.D.A.; Oliveira, C.E.D.S.; Jalal, A.; Gato, I.M.B.; Oliveira, T.J.S.S.; Boleta, G.H.M.; Giolo, V.M.; Vitória, L.S.; Tamburi, K.V.; Filho, M.C.M.T. Inoculation with Trichoderma harzianum and Azospirillum brasilense increases nutrition and yield of hydroponic lettuce. Arch. Microbiol. 2022, 204, 440. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.P.; Gomes, E.A.; de Sousa, S.M.; Lana, U.G.D.P.; Coelho, A.M.; Marriel, I.E.; de Oliveira-Paiva, C.A. Co-inoculation with tropical strains of Azospirillum and Bacillus is more efficient than single inoculation for improving plant growth and nutrient uptake in maize. Arch. Microbiol. 2022, 204, 143. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Chai, X.; Huang, Y.; Wang, X.; Han, Z.; Xu, X.; Wu, T.; Zhang, X.; Wang, Y. Siderophore production in pseudomonas SP. strain SP3 enhances iron acquisition in apple rootstock. J. Appl. Microbiol. 2022, 133, 720–732. [Google Scholar] [CrossRef] [PubMed]
- Akköprü, A.; Akat, Ş.; Özaktan, H.; Gül, A.; Akbaba, M. The long-term colonization dynamics of endophytic bacteria in cucumber plants, and their effects on yield, fruit quality and Angular Leaf Spot Disease. Sci. Hortic. 2021, 282, 110005. [Google Scholar] [CrossRef]
- Muthukumar, T.; Sulaiman, M.R. Root Endophytic Nectria haematococca Influences Growth, Root Architecture and Phosphorus Content of Green Gram in Different Substrates. Proc. Natl. Acad. Sci. India Sect. B Boil. Sci. 2021, 91, 131–138. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, D.; Huang, X.; Wang, S.; Qiu, R.; Zhang, Z.; Ming, J. Effect of Enterobacter sp. EG16 on Selenium biofortification and speciation in pak choi (Brassica rapa ssp. chinensis). Sci. Hortic. 2023, 310, 111723. [Google Scholar] [CrossRef]
- Dede, A.; Güven, K. Plant growth-promoting of olive and walnut actinobacteria: Isolation, screening PGP traits, antifungal activities, identification, and hydroponic production of wheat. Arch. Agron. Soil Sci. 2022, 1–16. [Google Scholar] [CrossRef]
- Dere, S.; Coban, A.; Akhoundnejad, Y.; Ozsoy, S.; Dasgan, H.Y. Use of Mycorrhiza to Reduce Mineral Fertilizers in Soilless Melon (Cucumis melo L.) Cultivation. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 1331–1336. [Google Scholar] [CrossRef] [Green Version]
- Roussis, I.; Beslemes, D.; Kosma, C.; Triantafyllidis, V.; Zotos, A.; Tigka, E.; Mavroeidis, A.; Karydogianni, S.; Kouneli, V.; Travlos, I.; et al. The Influence of Arbuscular Mycorrhizal Fungus Rhizophagus irregularis on the Growth and Quality of Processing Tomato (Lycopersicon esculentum Mill.) Seedlings. Sustainability 2022, 14, 9001. [Google Scholar] [CrossRef]
- Aini, N.; Yamika, W.S.D.; Pahlevi, R.W. The effect of nutrient concentration and inoculation of PGPR and AMF on the yield and fruit quality of hydroponic cherry tomatoes (Lycopersicum esculentum Mill. var. cerasiforme). J. Appl. Hortic. 2019, 21, 116–122. [Google Scholar] [CrossRef]
- Araújo, R.C.; Ribeiro, M.S.; Rodrigues, F.A.; Silva, B.S.; Dória, J.; Pasqual, M. Association of growth-promoting bacteria and hydroponic system aiming at reducing the time of production of banana seedlings. Arch. Agron. Soil Sci. 2022, 1–14. [Google Scholar] [CrossRef]
- Cardoso, D.S.C.P.; Martinez, H.E.P.; de Abreu, J.A.A.; Kasuya, M.C.M.; Sediyama, M.A.N. Growth-promoting fungi and potassium doses affects productivity and nutrition of cherry type tomatoes. J. Plant Nutr. 2022, 46, 835–851. [Google Scholar] [CrossRef]
- Dasgan, H.; Aydoner, G.; Akyol, M. Use of Some Microorganisms as Bio-Fertilizers in Soilless Grown Squash for Saving Chemical Nutrients. J. Plant Nutr. 2012, 46, 835–885. [Google Scholar] [CrossRef]
- Gül, A.; Kidoglu, F.; Tüzel, Y.; Tuzel, I. Different Treatments for Increasing Sustainability in Soilless Culture. Acta Hortic. 2007, 747, 595–602. [Google Scholar] [CrossRef]
- Sabir, A.; Karaca, U.; Yazar, K.; Sabir, F.; Yazici, M.A.; Dogan, O.; Kara, Z. Vine growth and yield response of alphonse lavallée (V. vinifera L.) Grapevines to plant growth promoting rhizobacteria under alkaline condition in soilless culture. Acta Sci. Pol. Hortorum Cultus 2017, 16, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Thongnok, S.; Siripornadulsil, W.; Siripornadulsil, S. Responses to arsenic stress of rice varieties coinoculated with the heavy metal-resistant and rice growth-promoting bacteria Pseudomonas stutzeri and Cupriavidus taiwanensis. Plant Physiol. Biochem. 2022, 191, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; An, R.; Grewal, P.; Yu, Z.; Borherova, Z.; Lee, J. High-Performing Windowfarm Hydroponic System: Transcriptomes of Fresh Produce and Microbial Communities in Response to Beneficial Bacterial Treatment. Mol. Plant-Microbe Interact. 2016, 29, 965–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al, M.M.S.E. Effect of the Probiotic (Bacillus spp.) on Water Quality, Production Performance, Microbial Profile, and Food Safety of the Nile Tilapia and Mint in Recirculating Aquaponic System. Egypt. J. Aquat. Biol. Fish. 2022, 26, 351–372. [Google Scholar] [CrossRef]
- Ergun, O.; Dasgan, H.; Isık, O. Effects of microalgae Chlorella vulgaris on hydroponically grown lettuce. Acta Hortic. 2020, 169–176. [Google Scholar] [CrossRef]
- Stegelmeier, A.A.; Rose, D.M.; Joris, B.R.; Glick, B.R. The Use of PGPB to Promote Plant Hydroponic Growth. Plants 2022, 11, 2783. [Google Scholar] [CrossRef] [PubMed]
- Li, R.-X.; Cai, F.; Pang, G.; Shen, Q.-R.; Li, R.; Chen, W. Solubilisation of Phosphate and Micronutrients by Trichoderma harzianum and Its Relationship with the Promotion of Tomato Plant Growth. PLoS ONE 2015, 10, e0130081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kıdoğlu, F.; Gül, A.; Tüzel, Y.; Özaktan, H.; Kidoglu, F. Yield Enhancement of Hydroponically Grown Tomatoes by Rhizobacteria. Acta Hortic. 2009, 475–480. [Google Scholar] [CrossRef]
- O’Khastini, R.; Idaryanto, F.R.; Alimuddin; Wahyuni, I.; Sari, I.J.; Puspita, N.; Wahyuni, A.F. Microbial consortia effects on the yields of water spinach in milkfish aquaponics system. IOP Conf. Ser. Earth Environ. Sci. 2019, 383, 012040. [Google Scholar] [CrossRef]
- Sakamoto, M.; Komatsu, Y.; Suzuki, T. Nutrient Deficiency Affects the Growth and Nitrate Concentration of Hydroponic Radish. Horticulturae 2021, 7, 525. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Ortega, W.M.; Martínez, V.; Nieves, M.; Simón, I.; Lidón, V.; Fernandez-Zapata, J.C.; Martinez-Nico-las, J.J.; Cáma-ra-Zapata, J.M.; García-Sánchez, F. Agricultural and physiological Responses of tomato plants Grown in Different Soilless Culturesystems with saline Water under Greenhouse Conditions. Sci. Rep. 2019, 9, 6733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifi Kalhor, M.; Aliniaeifard, S.; Seif, M.; Javadi, E.; Bernard, F.; Li, T.; Lastochkina, O. Rhizobacterium Bacillus subtilis reduces toxic effects of high electrical conductivity in soilless culture of lettuce. Acta Hortic. 2018, 1227, 471–478. [Google Scholar] [CrossRef]
- Moncada, A.; Vetrano, F.; Miceli, A. Alleviation of Salt Stress by Plant Growth-Promoting Bacteria in Hydroponic Leaf Lettuce. Agronomy 2020, 10, 1523. [Google Scholar] [CrossRef]
- Orhan, F. Alleviation of salt stress by halotolerant and halophilic plant growth-promoting bacteria in wheat (Triticum aestivum). Braz. J. Microbiol. 2016, 47, 621–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanafi, A.; Traoré, M.; Schnitzler, W.; Woitke, M. Induced resistance of tomato to whiteflies and pythium with the pgpr bacillus subtilis in a soilless crop grown under greenhouse conditions. Acta Hortic. 2007, 747, 315–323. [Google Scholar] [CrossRef]
- Gharelo, R.S.; Bandehag, A.; Toorchi, M.; Farajzadeh, D. Canola 2-dimensional proteome profiles under osmotic stress and inoculation with Pseudomonas fluorescens FY32. Plant Cell Biotech. Mol. Biol 2016, 17, 257–266. [Google Scholar]
- Tarroum, M.; Ben Romdhane, W.; Al-Qurainy, F.; Ali, A.A.M.; Al-Doss, A.; Fki, L.; Hassairi, A. A novel PGPF Penicillium olsonii isolated from the rhizosphere of Aeluropus littoralis promotes plant growth, enhances salt stress tolerance, and reduces chemical fertilizers inputs in hydroponic system. Front. Microbiol. 2022, 13, 996054. [Google Scholar] [CrossRef] [PubMed]
- Postma, J.; Van Os, E.; Bonants, P.J. Pathogen Detection and Management Strategies in Soilless Plant Growing Systems. In Soilless Culture: Theory and Practice, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2008; pp. 425–457. [Google Scholar] [CrossRef]
- Promwee, A.; Intana, M. Trichoderma asperellum (NST-009): A potential native antagonistic fungus to control Cercospora leaf spot and promote the growth of 'Green Oak' lettuce (Lactuca sativa L.) cultivated in the commercial NFT hydroponic system. Plant Prot. Sci. 2022, 58, 139–149. [Google Scholar] [CrossRef]
- Pu, X.; Xie, B.; Li, P.; Mao, Z.; Ling, J.; Shen, H.; Zhang, J.; Huang, N.; Lin, B. Analysis of the defence-related mechanism in cucumber seedlings in relation to root colonization by nonpathogenic Fusarium oxysporum CS-20. FEMS Microbiol. Lett. 2014, 355, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Xiao, R.; Chen, M.; Wu, H.; Gao, X.; Wang, J. An avirulent Ralstonia solanacearum strain FJAT1458 outcompetes with virulent strain and induces tomato plant resistance against bacterial wilt. Pest Manag. Sci. 2022, 78, 5002–5013. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, H.; Muslim, A.; Suzuki, T.; Hyakumachi, M. Fusarium equiseti GF191 as an effective biocontrol agent against Fusarium crown and root rot of tomato in rock wool systems. Crop. Prot. 2007, 26, 1514–1523. [Google Scholar] [CrossRef]
- Khalil, S. Influence of electrical conductivity on biological activity of Pythium ultimum and Binab T in a closed soilless system. J. Plant Dis. Prot. 2011, 118, 102–108. [Google Scholar] [CrossRef]
- Thongkamngam, T.; Jaenaksorn, T. Fusarium oxysporum (F221-B) as biocontrol agent against plant pathogenic fungi in vitro and in hydroponics. Plant Prot. Sci. 2017, 53, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Rose, S.; Parker, M.; Punja, Z.K. Efficacy of Biological and Chemical Treatments for Control of Fusarium Root and Stem Rot on Greenhouse Cucumber. Plant Dis. 2003, 87, 1462–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Freitas, C.C.; Taylor, C.G. Biological control of hairy root disease using beneficial Pseudomonas strains. Biol. Control. 2023, 177, 105098. [Google Scholar] [CrossRef]
- Rais, A.; Jabeen, Z.; Shair, F.; Hafeez, F.Y.; Hassan, M.N. Bacillus spp., a bio-control agent enhances the activity of antioxidant defense enzymes in rice against Pyricularia oryzae. PLoS ONE 2017, 12, e0187412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamilova, F.; Validov, S.; Lugtenberg, B. Biological control of tomato foot and root rot caused by Fusarium oxysporum f. sp. Radicis-lycopersici by pseudomonas bacteria. Acta Hortic. 2009, 808, 317–320. [Google Scholar] [CrossRef]
- Alsanius, B.; Lundqvist, S.; Persson, E.; Gustafsson, K.-A.; Olsson, M.; Khalil, S. Yield and fruit quality of tomato grown in a closed hydroponic greenhouse system as affected by ythium ultimum attack and biological control agents. Acta Hortic. 2004, 644, 575–582. [Google Scholar] [CrossRef]
- Adhikari, R.; Monkhung, S.; Mongkol, R.; Kanjanamaneesathian, M. Paenibacillus polymyxa reduces leaf spot disease caused by Cersocpora sp. and has potential to promote growth in green oak lettuce (Lactuca sativa var. crispa) in the commercial hydroponic farm. Acta Hortic. 2022, 1339, 463–470. [Google Scholar] [CrossRef]
- Evenhuis, B.; Nijhuis, E.; Lamers, J.; Verhoeven, J.; Postma, J. Alternative Methods to Control Phytophthora Cactorum in Strawberry Cultivated in Soilless Growing Media. In Proceedings of the VIII International Symposium on Chemical and Non-Chemical Soil and Substrate Disinfestation, Torino, Italy, 13 July 2014; pp. 337–342. [Google Scholar] [CrossRef]
- De Jonghe, K.; De Dobbelaere, I.; Sarrazyn, R.; Hofte, M. Control of Phytophthora cryptogea in the hydroponic forcing of witloof chicory with the rhamnolipid-based biosurfactant formulation PRO1. Plant Pathol. 2005, 54, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Van Os, E. Disease Management in Soilless Culture Systems. Acta Hortic. 2010, 883, 385–393. [Google Scholar] [CrossRef]
- Vallance, J.; Déniel, F.; Le Floch, G.; Guérin-Dubrana, L.; Blancard, D.; Rey, P. Pathogenic and beneficial microorganisms in soilless cultures. Agron. Sustain. Dev. 2011, 31, 191–203. [Google Scholar] [CrossRef]
- Schnitzler, W. Pest and Disease Management of Soilless Culture. Acta Hortic. 2004, 648, 191–203. [Google Scholar] [CrossRef]
- Alsanius, B.W.; Wohanka, W. Root Zone Microbiology of Soilless Cropping Systems. In Soilless Culture: Theory and Practice Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2019; pp. 149–194. [Google Scholar] [CrossRef]
- Van Gerrewey, T.; Boon, N.; Geelen, D. Vertical Farming: The Only Way Is Up? Agronomy 2021, 12, 2. [Google Scholar] [CrossRef]
- Maboko, M.M.; Bertling, I.; Du Plooy, C.P. Arbuscular mycorrhiza has limited effects on yield and quality of tomatoes grown under soilless cultivation. Acta Agric. Scand. Sect. B -Soil Plant Sci. 2013, 63, 261–270. [Google Scholar] [CrossRef]
- Kowalska, I.; Gąstoł, M.; Konieczny, A. Effect of mycorrhiza and the phosphorus content in a nutrient solution on the yield and nutritional status of lettuce grown on various substrates. J. Elem. 2012, 20, 631–642. [Google Scholar] [CrossRef] [Green Version]
- Cela, F.; Avio, L.; Giordani, T.; Vangelisti, A.; Cavallini, A.; Turrini, A.; Sbrana, C.; Pardossi, A.; Incrocci, L. Arbuscular Mycorrhizal Fungi Increase Nutritional Quality of Soilless Grown Lettuce while Overcoming Low Phosphorus Supply. Foods 2022, 11, 3612. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Corá, J.E. Bulk density and relationship air/water of horticultural substrate. Sci. Agric. 2004, 61, 446–450. [Google Scholar] [CrossRef] [Green Version]
- Gaur, A.; Adholeya, A. Effects of the particle size of soil-less substrates upon AM fungus inoculum production. Mycorrhiza 2000, 10, 43–48. [Google Scholar] [CrossRef]
- Azizoglu, U.; Yilmaz, N.; Simsek, O.; Ibal, J.C.; Tagele, S.B.; Shin, J.-H. The fate of plant growth-promoting rhizobacteria in soilless agriculture: Future perspectives. 3 Biotech 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Ngosong, C.; Tatah, B.N.; Olougou, M.N.E.; Suh, C.; Nkongho, R.N.; Ngone, M.A.; Achiri, D.T.; Tchakounté, G.V.T.; Ruppel, S. Inoculating plant growth-promoting bacteria and arbuscular mycorrhiza fungi modulates rhizosphere acid phosphatase and nodulation activities and enhance the productivity of soybean (Glycine max). Front. Plant Sci. 2022, 13, 934339. [Google Scholar] [CrossRef]
- Rios-Ruiz, W.F.; Tuanama-Reátegui, C.; Huamán-Córdova, G.; Valdez-Nuñez, R.A. Co-Inoculation of Endophytes Bacillus siamensis TUR07-02b and Priestia megaterium SMBH14-02 Promotes Growth in Rice with Low Doses of Nitrogen Fertilizer. Plants 2023, 12, 524. [Google Scholar] [CrossRef]
- Rivas-García, T.; González-Estrada, R.R.; Chiquito-Contreras, R.G.; Reyes-Pérez, J.J.; González-Salas, U.; Hernández-Montiel, L.G.; Murillo-Amador, B. Biocontrol of Phytopathogens under Aquaponics Systems. Water 2020, 12, 2061. [Google Scholar] [CrossRef]
- Qiu, Z.; Egidi, E.; Liu, H.; Kaur, S.; Singh, B.K. New frontiers in agriculture productivity: Optimised microbial inoculants and in situ microbiome engineering. Biotechnol. Adv. 2019, 37, 107371. [Google Scholar] [CrossRef]
- Koohakan, P.; Ikeda, H.; Jeanaksorn, T.; Tojo, M.; Kusakari, S.-I.; Okada, K.; Sato, S. Evaluation of the indigenous microorganisms in soilless culture: Occurrence and quantitative characteristics in the different growing systems. Sci. Hortic. 2004, 101, 179–188. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mourouzidou, S.; Ntinas, G.K.; Tsaballa, A.; Monokrousos, N. Introducing the Power of Plant Growth Promoting Microorganisms in Soilless Systems: A Promising Alternative for Sustainable Agriculture. Sustainability 2023, 15, 5959. https://doi.org/10.3390/su15075959
Mourouzidou S, Ntinas GK, Tsaballa A, Monokrousos N. Introducing the Power of Plant Growth Promoting Microorganisms in Soilless Systems: A Promising Alternative for Sustainable Agriculture. Sustainability. 2023; 15(7):5959. https://doi.org/10.3390/su15075959
Chicago/Turabian StyleMourouzidou, Snezhana, Georgios K. Ntinas, Aphrodite Tsaballa, and Nikolaos Monokrousos. 2023. "Introducing the Power of Plant Growth Promoting Microorganisms in Soilless Systems: A Promising Alternative for Sustainable Agriculture" Sustainability 15, no. 7: 5959. https://doi.org/10.3390/su15075959