
Seed Provision Efficacy of Detached Reproductive Shoots in Restoration Projects for Degraded Eelgrass (Zostera marina L.) Meadows
Abstract
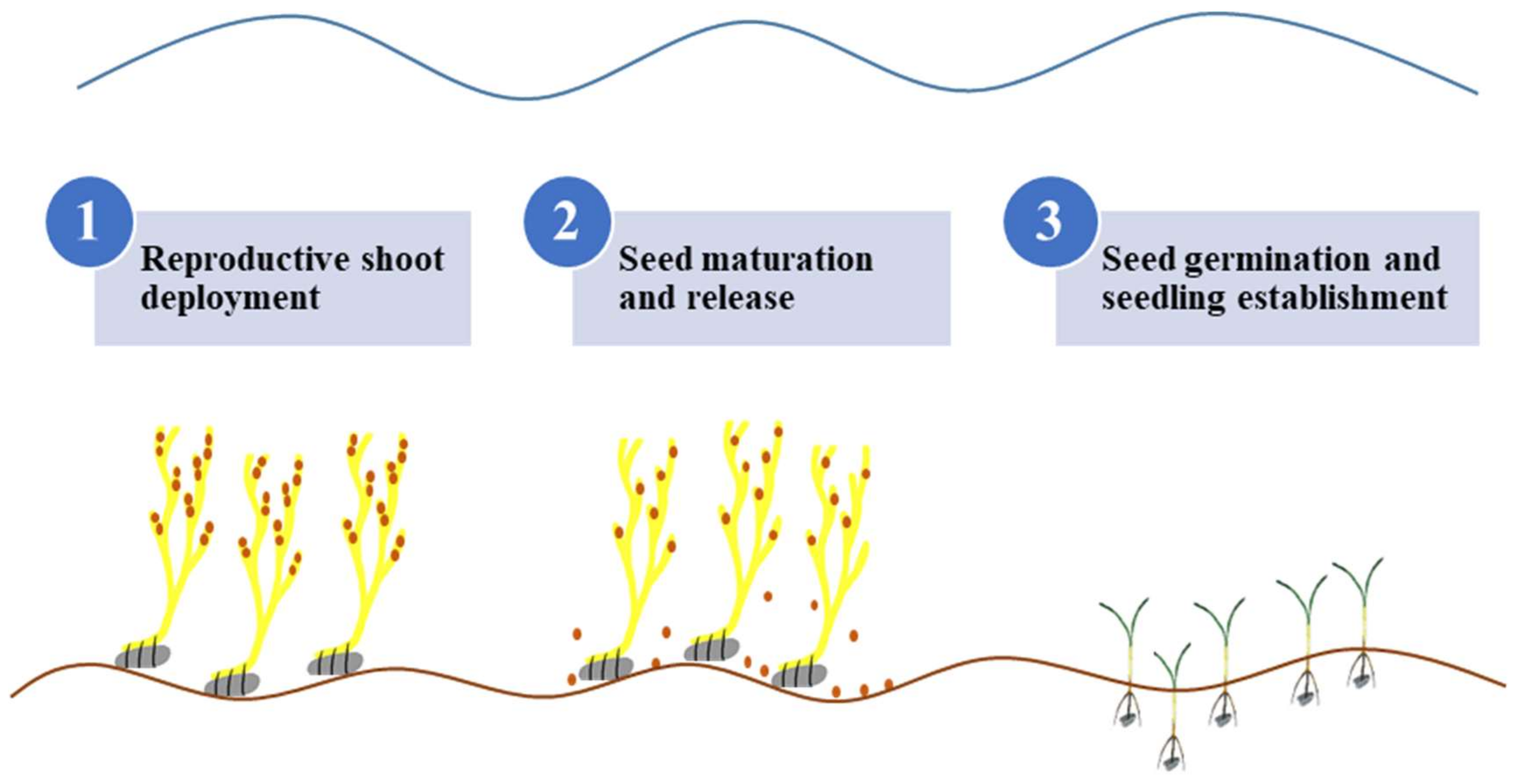
:1. Introduction
2. Materials and Methods
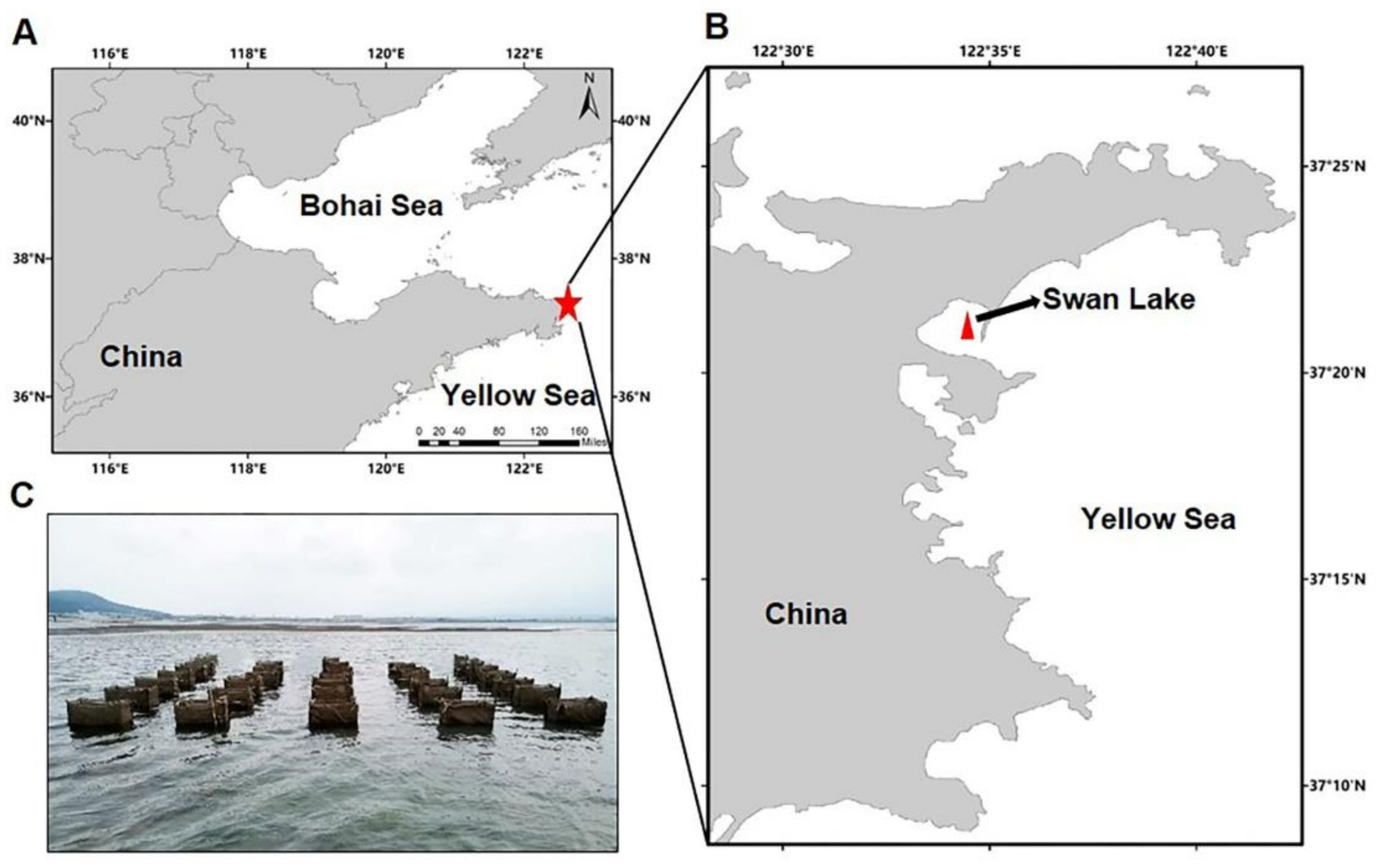
2.1. Study Site
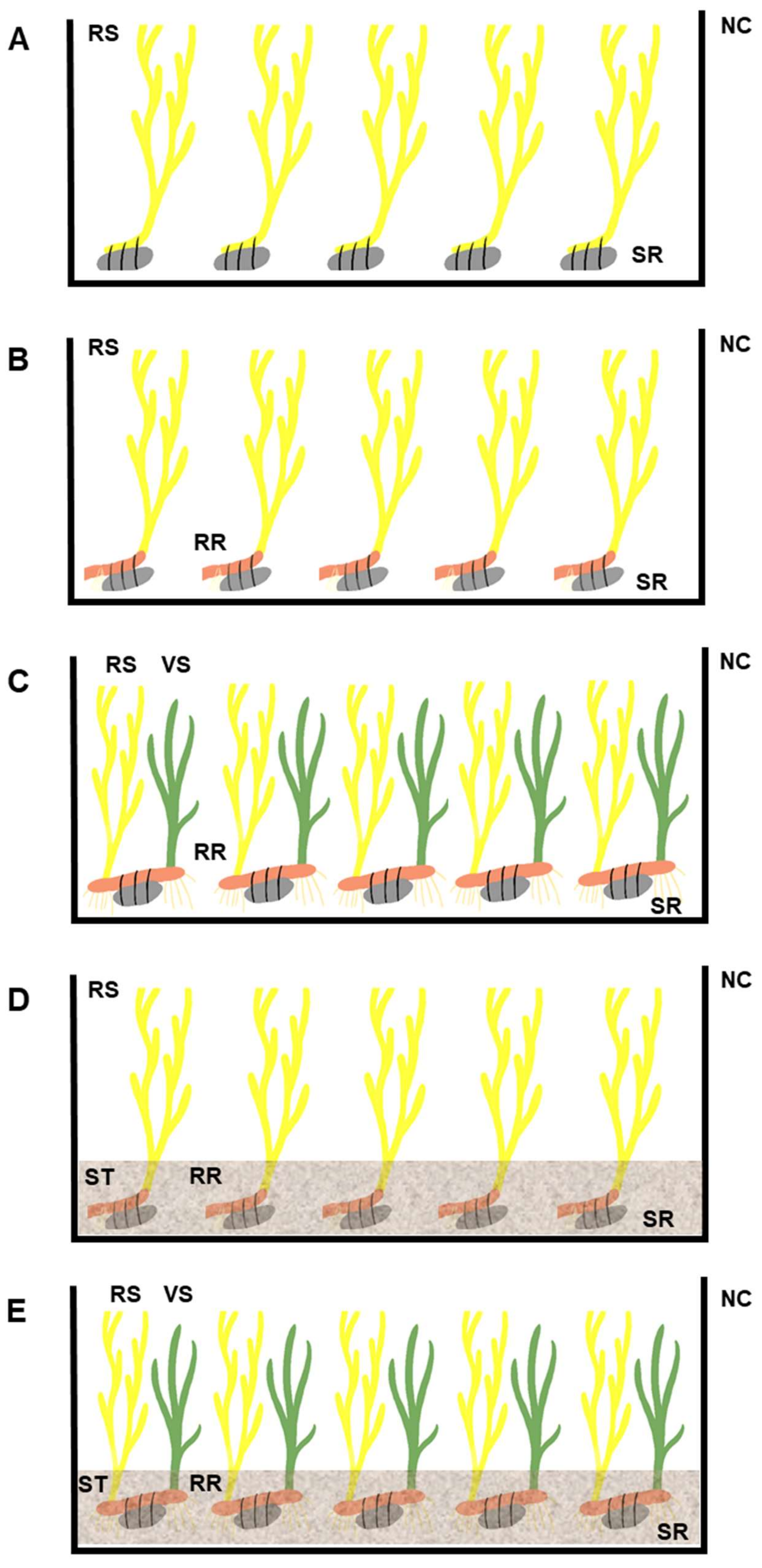
2.2. Experimental Design
2.3. Seed Collection and Evaluation

2.4. Environmental Variables
2.5. Data Analysis
3. Results
3.1. Environment Variables
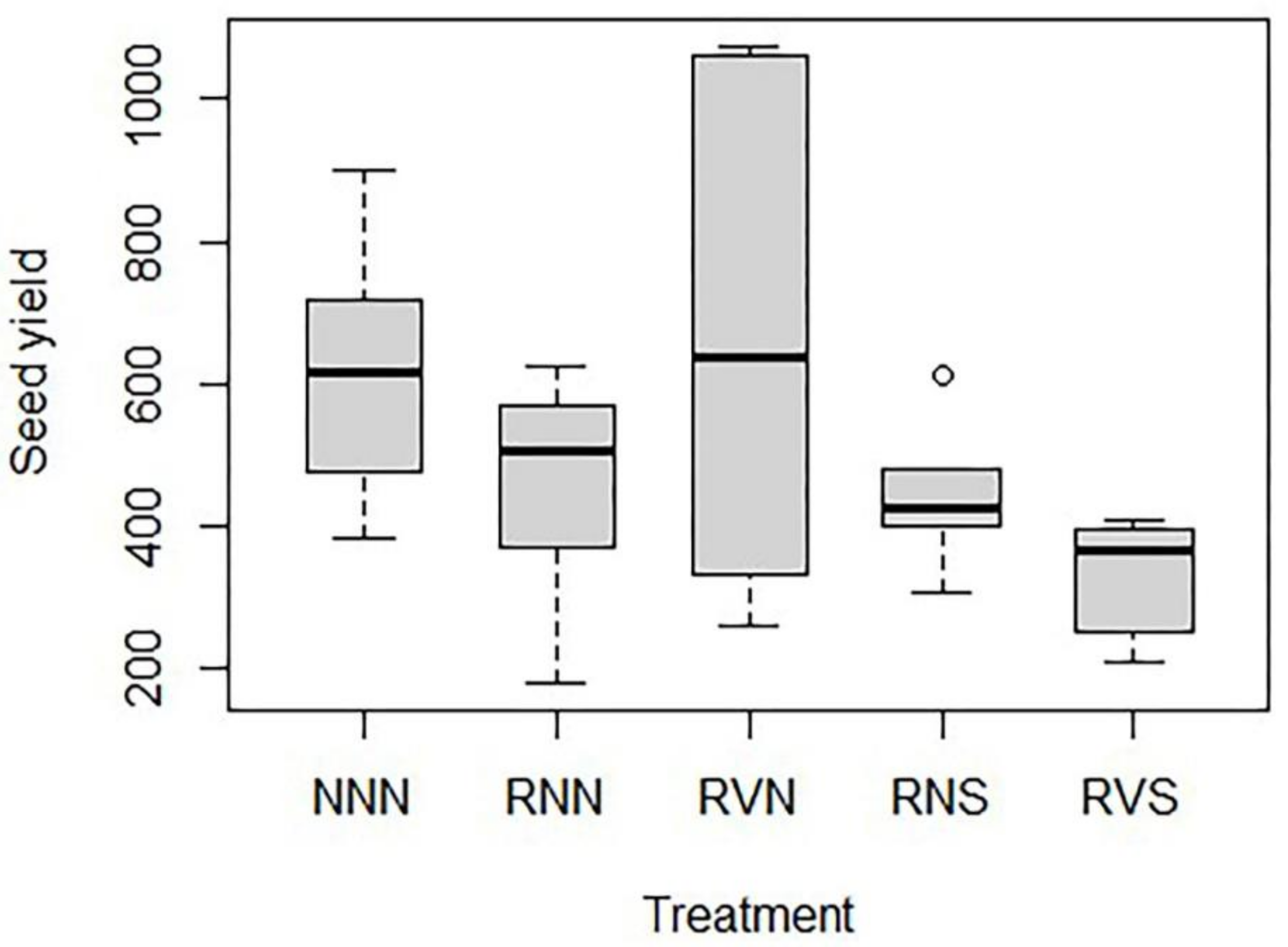
3.2. Seed Yield
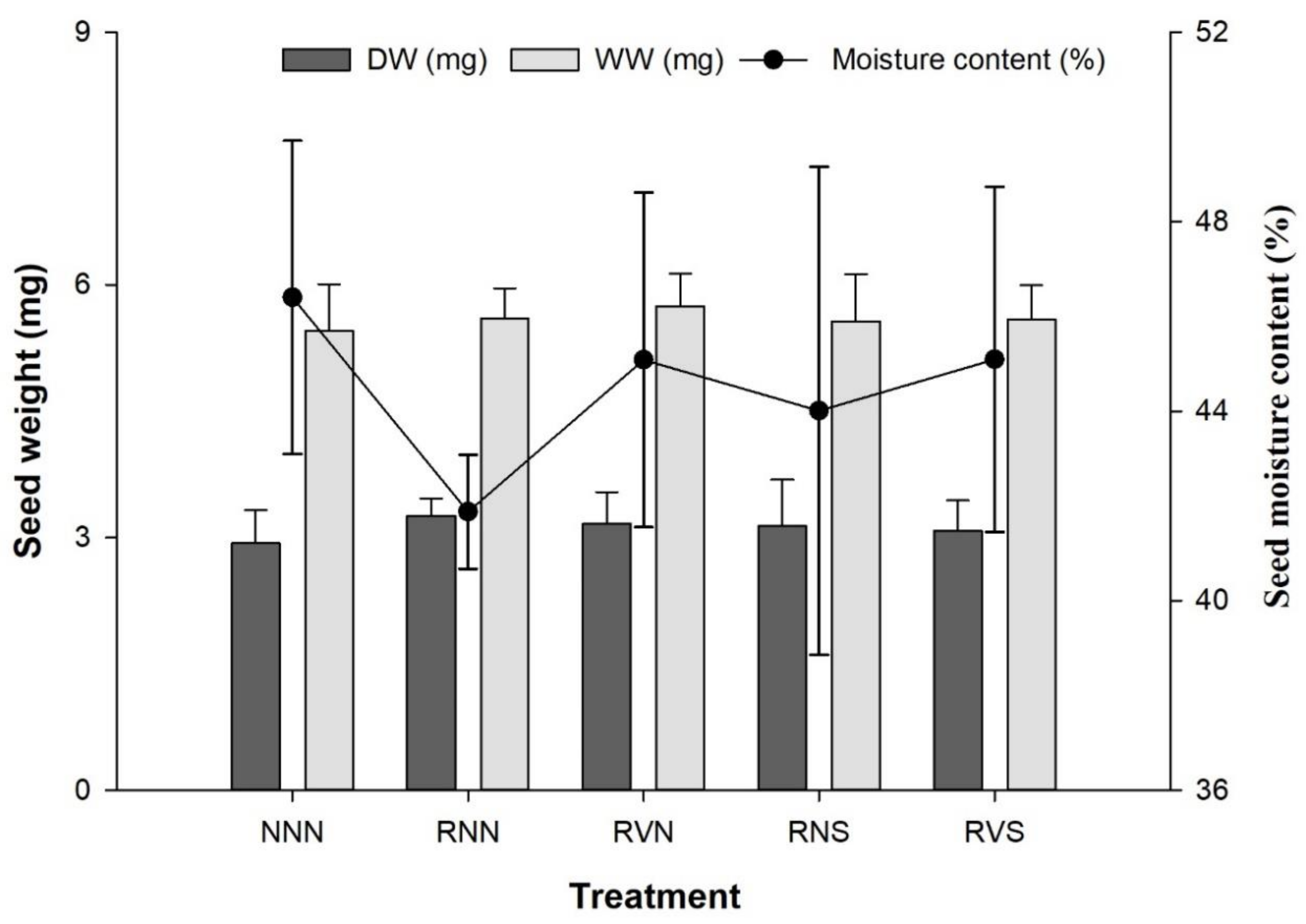
3.3. Seed Weight
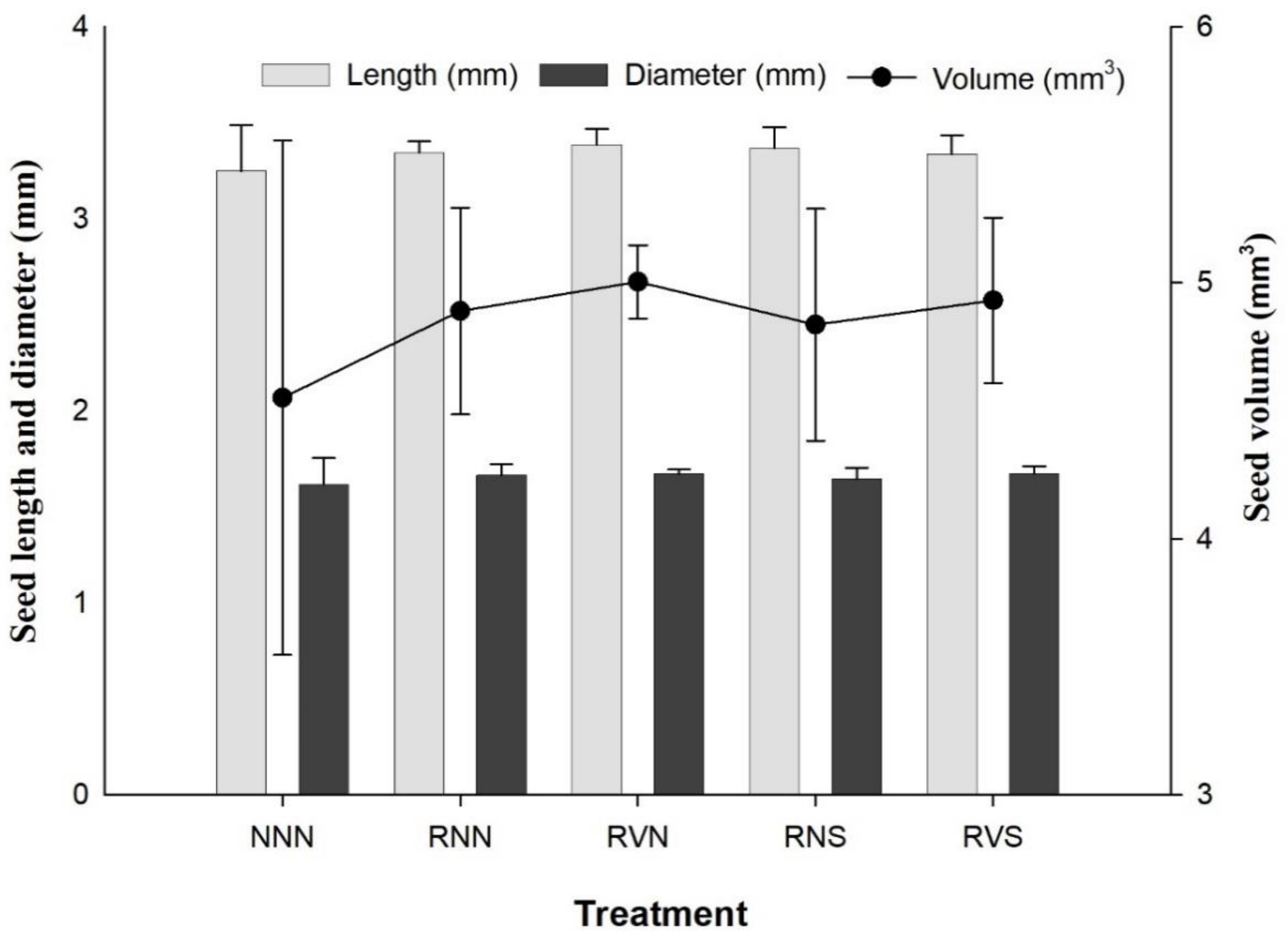
3.4. Seed Size
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Short, F.; Carruthers, T.; Dennison, W.; Waycott, M. Global seagrass distribution and diversity: A bioregional model. J. Exp. Mar. Biol. Ecol. 2007, 350, 3–20. [Google Scholar] [CrossRef]
- Unsworth, R.K.F.; Nordlund, L.M.; Cullen-Unsworth, L.C. Seagrass meadows support global fisheries production. Conserv. Lett. 2019, 12, e12566. [Google Scholar] [CrossRef]
- Lamb, J.B.; van de Water, J.A.; Bourne, D.G.; Altier, C.; Hein, M.Y.; Fiorenza, E.A.; Abu, N.; Jompa, J.; Harvell, C.D. Seagrass ecosystems reduce exposure to bacterial pathogens of humans, fishes, and invertebrates. Science 2017, 355, 731–733. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.M.; Krause-Jensen, D. Export from seagrass meadows contributes to marine carbon sequestration. Front. Mar. Sci. 2017, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Fourqurean, J.W.; Duarte, C.M.; Kennedy, H.; Marba, N.; Holmer, M.; Mateo, M.A.; Apostolaki, E.T.; Kendrick, G.A.; Krause-Jensen, D.; McGlathery, K.J.; et al. Seagrass ecosystems as a globally significant carbon stock. Nat. Geosci. 2012, 5, 505–509. [Google Scholar] [CrossRef]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L., Jr.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [Green Version]
- Short, F.T.; Neckles, H.A. The effects of global climate change on seagrasses. Aquat. Bot. 1999, 63, 169–196. [Google Scholar] [CrossRef]
- Orth, R.J.; Carruthers, T.J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, S.; et al. A global crisis for seagrass ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, E.A.; Sherman, C.D.H.; Statton, J.; Copeland, C.; Matthews, A.; Waycott, M.; van Dijk, K.J.; Vergés, A.; Kajlich, L.; McLeod, I.M.; et al. Advances in approaches to seagrass restoration in australia. Ecol. Manag. Restor. 2021, 22, 10–21. [Google Scholar] [CrossRef]
- Katwijk, M.M.; Thorhaug, A.; Marbà, N.; Orth, R.J.; Duarte, C.M.; Kendrick, G.A.; Althuizen, I.H.J.; Balestri, E.; Bernard, G.; Cambridge, M.L.; et al. Global analysis of seagrass restoration: The importance of large-scale planting. J. Appl. Ecol. 2015, 53, 567–578. [Google Scholar] [CrossRef] [Green Version]
- Thorhaug, A.; Poulos, H.M.; Lopez-Portillo, J.; Ku, T.C.W.; Berlyn, G.P. Seagrass blue carbon dynamics in the gulf of Mexico: Stocks, losses from anthropogenic disturbance, and gains through seagrass restoration. Sci. Total Environ. 2017, 605–606, 626–636. [Google Scholar] [CrossRef]
- Greiner, J.T.; McGlathery, K.J.; Gunnell, J.; McKee, B.A. Seagrass restoration enhances “blue carbon” sequestration in coastal waters. PLoS ONE 2013, 8, e72469. [Google Scholar] [CrossRef] [Green Version]
- Orth, R.J.; Lefcheck, J.S.; McGlathery, K.S.; Aoki, L.; Luckenbach, M.W.; Moore, K.A.; Oreska, M.P.J.; Snyder, R.; Wilcox, D.J.; Lusk, B. Restoration of seagrass habitat leads to rapid recovery of coastal ecosystem services. Sci. Adv. 2020, 6, eabc6434. [Google Scholar] [CrossRef]
- Johnson, A.J.; Orth, R.J.; Moore, K.A. The role of sexual reproduction in the maintenance of established Zostera marina meadows. J. Ecol. 2020, 108, 945–957. [Google Scholar] [CrossRef] [Green Version]
- Setchell, W.A. Morphological and phenological notes on Zostera marina L. Univ. Calif. Publ. Bot. 1929, 14, 389–452. [Google Scholar]
- Xu, S.; Wang, P.; Zhou, Y.; Zhang, X.; Gu, R.; Liu, X.; Liu, B.; Song, X.; Xu, S.; Yue, S. New insights into different reproductive effort and sexual recruitment contribution between two geographic Zostera marina L. populations in temperate China. Front. Plant Sci. 2018, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Furman, B.T.; Jackson, L.J.; Bricker, E.; Peterson, B.J. Sexual recruitment in Zostera marina: A patch to landscape-scale investigation. Limnol. Oceanogr. 2015, 60, 584–599. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, J.; Moore, K.; Kenworthy, W. Characterization and ecological implication of eelgrass life history strategies near the species southern limit in the western North Atlantic. Mar. Ecol. Prog. Ser. 2012, 444, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Stubler, A.D.; Jackson, L.J.; Furman, B.T.; Peterson, B.J. Seed production patterns in Zostera marina: Effects of patch size and landscape configuration. Estuaries Coasts 2017, 40, 564–572. [Google Scholar] [CrossRef]
- Tomlinson, P.B. Vegetative morphology and meristem dependence—The foundation of productivity in seagrasses. Aquaculture 1974, 4, 107–130. [Google Scholar] [CrossRef]
- Plus, M.; Deslous-Paoli, J.-M.; Dagault, F. Seagrass (Zostera marina L.) bed recolonisation after anoxia-induced full mortality. Aquat. Bot. 2003, 77, 121–134. [Google Scholar] [CrossRef]
- Lee, K.-S.; Park, S.R.; Kim, J.-B. Production dynamics of the eelgrass, Zostera marina in two bay systems on the south coast of the Korean peninsula. Mar. Biol. 2005, 147, 1091–1108. [Google Scholar] [CrossRef]
- Davis, R.C.; Short, F.T. Restoring eelgrass, Zostera marina L., habitat using a new transplanting technique: The horizontal rhizome method. Aquat. Bot. 1997, 59, 1–15. [Google Scholar] [CrossRef]
- Paling, E.I.; van Keulen, M.; Wheeler, K.; Phillips, J.; Dyhrberg, R. Mechanical seagrass transplantation in western Australia. Ecol. Eng. 2001, 16, 331–339. [Google Scholar] [CrossRef]
- Paling, E.I.; van Keulen, M.; Wheeler, K.D.; Phillips, J.; Dyhrberg, R.; Lord, D.A. Improving mechanical seagrass transplantation. Ecol. Eng. 2001, 18, 107–113. [Google Scholar] [CrossRef]
- Fishman, J.R.; Orth, R.J.; Marion, S.; Bieri, J. A comparative test of mechanized and manual transplanting of eelgrass, Zostera marina, in Chesapeake Bay. Restor. Ecol. 2004, 12, 214–219. [Google Scholar] [CrossRef]
- Lee, K.S.; Park, J.I. An effective transplanting technique using shells for restoration of Zostera marina habitats. Mar. Pollut. Bull. 2008, 56, 1015–1021. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, P.; Liu, B.; Liu, X.; Zhang, X.; Wang, F.; Yang, H. Restoring eelgrass (Zostera marina L.) habitats using a simple and effective transplanting technique. PLoS ONE 2014, 9, e92982. [Google Scholar] [CrossRef]
- Marion, S.R.; Orth, R.J. Innovative techniques for large-scale seagrass restoration using Zostera marina (eelgrass) seeds. Restor. Ecol. 2008, 18, 514–526. [Google Scholar] [CrossRef]
- Orth, R.; Bieri, J.; Fishman, J.; Harwell, M.; Marion, S.; Moore, K.; Nowak, J.; Montfrans, J. A review of techniques using adult plants and seeds to transplant eelgrass (Zostera marina L.) in Chesapeake Bay and the Virginia coastal bays. In Proceedings of the Conference: Seagrass Restoration: Success, Failure, and the Costs of Both, Sarasota, FL, USA, 11 March 2003; pp. 1–17. [Google Scholar]
- Phillips, R.C.; Stewart Grant, W.; Peter McRoy, C. Reproductive strategies of eelgrass (Zostera marina L.). Aquat. Bot. 1983, 16, 1–20. [Google Scholar] [CrossRef]
- Olesen, B. Reproduction in Danish eelgrass (Zostera marina L.) stands: Size-dependence and biomass partitioning. Aquat. Bot. 1999, 65, 209–219. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, J.; Park, S.R.; Lee, K.-S. Annual and perennial life history strategies of Zostera marina populations under different light regimes. Mar. Ecol. Prog. Ser. 2014, 509, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Orth, R.J.; Marion, S.R.; Granger, S.; Traber, M. Evaluation of a mechanical seed planter for transplanting Zostera marina (eelgrass) seeds. Aquat. Bot. 2009, 90, 204–208. [Google Scholar] [CrossRef]
- Golden, R.R.; Busch, K.E.; Karrh, L.P.; Parham, T.A.; Lewandowski, M.J.; Naylor, M.D. Large-scale Zostera marina (eelgrass) restoration in Chesapeake Bay, Maryland, USA. Part II: A comparison of restoration methods in the Patuxent and Potomac rivers. Restor. Ecol. 2010, 18, 501–513. [Google Scholar] [CrossRef]
- Pickerell, C.H.; Schott, S.; Wyllie-Echeverria, S. Buoy-deployed seeding: Demonstration of a new eelgrass (Zostera marina L.) planting method. Ecol. Eng. 2005, 25, 127–136. [Google Scholar] [CrossRef]
- Govers, L.L.; Heusinkveld, J.H.T.; Grafnings, M.L.E.; Smeele, Q.; van der Heide, T. Adaptive intertidal seed-based seagrass restoration in the Dutch Wadden Sea. PLoS ONE 2022, 17, e0262845. [Google Scholar] [CrossRef]
- Harwell, M.C.; Orth, R.J. Eelgrass (Zostera marina L.) seed protection for field experiments and implications for large-scale restoration. Aquat. Bot. 1999, 64, 51–61. [Google Scholar] [CrossRef]
- Zhang, P.D.; Fang, C.; Liu, J.; Xu, Q.; Li, W.T.; Liu, Y.S. An effective seed protection method for planting Zostera marina (eelgrass) seeds: Implications for their large-scale restoration. Mar. Pollut. Bull. 2015, 95, 89–99. [Google Scholar] [CrossRef]
- Unsworth, R.K.F.; Bertelli, C.M.; Cullen-Unsworth, L.C.; Esteban, N.; Jones, B.L.; Lilley, R.; Lowe, C.; Nuuttila, H.K.; Rees, S.C. Sowing the seeds of seagrass recovery using hessian bags. Front. Ecol. Evol. 2019, 7, 311. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Zhou, Y.; Qiao, Y.; Yue, S.; Zhang, X.; Zhang, Y.; Liu, M.; Zhang, Y.; Zhang, Z. Seagrass restoration using seed ball burial in northern China. Restor. Ecol. 2022, 31, e13691. [Google Scholar] [CrossRef]
- Reynolds, L.K.; Waycott, M.; McGlathery, K.J.; Orth, R.J.; Zieman, J.C. Eelgrass restoration by seed maintains genetic diversity: Case study from a coastal bay system. Mar. Ecol. Prog. Ser. 2012, 448, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Granger, S.; Traber, M.; Nixon, S.; Keyes, R. A practical guide for the use of seeds in eelgrass (Zostera marina L.) restoration. In Part I. Collection, Processing, and Storage; Schwartz, M., Ed.; Rhode Island Sea Grant: Narragansett, RI, USA, 2003; 20p. [Google Scholar]
- Orth, R.J.; Harwell, M.C.; Bailey, E.M.; Bartholomew, A.; Jawad, J.T.; Lombana, A.V.; Moore, K.A.; Rhode, J.M.; Woods, H.E. A review of issues in seagrass seed dormancy and germination: Implications for conservation and restoration. Mar. Ecol. Prog. Ser. 2000, 200, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, H.; Tang, X. Growth characteristics of a restored Zostera marina population in the Shandong Peninsula, China: A case study. J. Sea Res. 2019, 144, 122–132. [Google Scholar] [CrossRef]
- Eriander, L.; Infantes, E.; Olofsson, M.; Olsen, J.L.; Moksnes, P.-O. Assessing methods for restoration of eelgrass (Zostera marina L.) in a cold temperate region. J. Exp. Mar. Biol. Ecol. 2016, 479, 76–88. [Google Scholar] [CrossRef]
- De Cock, A.W.A.M. Flowering, pollination and fruiting in Zostera marina L. Aquat. Bot. 1980, 9, 201–220. [Google Scholar] [CrossRef]
- Orth, R.J.; Moore, K.A.; Marion, S.R.; Wilcox, D.J.; Parrish, D.B. Seed addition facilitates eelgrass recovery in a coastal bay system. Mar. Ecol. Prog. Ser. 2012, 448, 177–195. [Google Scholar] [CrossRef] [Green Version]
- Busch, K.E.; Golden, R.R.; Parham, T.A.; Karrh, L.P.; Lewandowski, M.J.; Naylor, M.D. Large-scale Zostera marina (eelgrass) restoration in Chesapeake Bay, Maryland, USA. Part I: A comparison of techniques and associated costs. Restor. Ecol. 2008, 18, 490–500. [Google Scholar] [CrossRef]
- Jacobs, R.P.W.M.; Pierson, E.S. Phenology of reproductive shoots of eelgrass, Zostera-marina L., at Roscoff (France). Aquat. Bot. 1981, 10, 45–60. [Google Scholar] [CrossRef]
- Keddy, C.J. Reproduction of annual eelgrass: Variation among habitats and comparison with perennial eelgrass (Zostera marina L.). Aquat. Bot. 1987, 27, 243–256. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, X.J.; Liu, B.J.; Liu, P.; Wang, F.; Zhang, X.M.; Yang, H.S. Unusual pattern in characteristics of the eelgrass Zostera marina L. in a shallow lagoon (Swan Lake), north China: Implications on the importance of seagrass conservation. Aquat. Bot. 2015, 120, 178–184. [Google Scholar] [CrossRef]
- Xu, S.; Xu, S.; Zhou, Y.; Zhao, P.; Yue, S.; Song, X.; Zhang, X.; Gu, R.; Wang, P.; Zhang, Y. Single beam sonar reveals the distribution of the eelgrass Zostera marina L. and threats from the green tide algae Chaetomorpha linum L. in Swan-lake lagoon (China). Mar. Pollut. Bull. 2019, 145, 611–623. [Google Scholar] [CrossRef]
- Probert, R.J.; Brenchley, J.L. The effect of environmental factors on field and laboratory germination in a population of Zostera marina L. from southern England. Seed Sci. Res. 2007, 9, 331–339. [Google Scholar] [CrossRef]
- Wyllie-Echeverria, S.; Cox, P.A.; Churchill, A.C.; Brotherson, J.D.; Wyllie-Echeverria, T. Seed size variation within Zostera marina L. (Zosteraceae). Bot. J. Linn. Soc. 2003, 142, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Dooley, F.D.; Wyllie-Echeverria, S.; Van Volkenburgh, E. Long-term seed storage and viability of Zostera marina. Aquat. Bot. 2013, 111, 130–134. [Google Scholar] [CrossRef]
- Delefosse, M.; Povidisa, K.; Poncet, D.; Kristensen, E.; Olesen, B. Variation in size and chemical composition of seeds from the seagrass Zostera marina—Ecological implications. Aquat. Bot. 2016, 131, 7–14. [Google Scholar] [CrossRef]
- Olesen, B.; Marba, N.; Duarte, C.M.; Savela, R.S.; Fortes, M.D. Recolonization dynamics in a mixed seagrass meadow: The role of clonal versus sexual processes. Estuaries 2004, 27, 770–780. [Google Scholar] [CrossRef] [Green Version]
- Greve, T.M.; Krause-Jensen, D.; Rasmussen, M.B.; Christensen, P.B. Means of rapid eelgrass (Zostera marina L.) recolonisation in former dieback areas. Aquat. Bot. 2005, 82, 143–156. [Google Scholar] [CrossRef]
- Lee, K.S.; Park, J.I.; Kim, Y.K.; Sang, R.P.; Kim, J.H. Recolonization of Zostera marina following destruction caused by a red tide algal bloom: The role of new shoot recruitment from seed banks. Mar. Ecol. Prog. 2007, 342, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Infantes, E.; Moksnes, P.-O. Eelgrass seed harvesting: Flowering shoots development and restoration on the Swedish west coast. Aquat. Bot. 2018, 144, 9–19. [Google Scholar] [CrossRef]
- Marba, N. Rhizome elongation and seagrass clonal growth. Mar. Ecol. Prog. Ser. 1998, 174, 269–280. [Google Scholar] [CrossRef]
- Govers, L.L.; Man In ‘t Veld, W.A.; Meffert, J.P.; Bouma, T.J.; van Rijswick, P.C.; Heusinkveld, J.H.; Orth, R.J.; van Katwijk, M.M.; van der Heide, T. Marine phytophthora species can hamper conservation and restoration of vegetated coastal ecosystems. Proc. R. Soc. B Biol. Sci. 2016, 283, 2016081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgensen, M.S.; Labouriau, R.; Olesen, B. Seed size and burial depth influence Zostera marina L. (eelgrass) seed survival, seedling emergence and initial seedling biomass development. PLoS ONE 2019, 14, e0215157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Zhou, Y.; Wang, P.; Wang, F.; Zhang, X.; Gu, R. Salinity and temperature significantly influence seed germination, seedling establishment, and seedling growth of eelgrass Zostera marina L. PeerJ 2016, 4, e2697. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.-Z.; Li, W.-T.; Zhang, X.-M.; Nie, M.; Li, Y. Sexual reproduction and seed dispersal pattern of annual and perennial Zostera marina in a heterogeneous habitat. Wetl. Ecol. Manag. 2014, 22, 671–682. [Google Scholar] [CrossRef]
- Olesen, B.; Krause-Jensen, D.; Christensen, P.B. Depth-related changes in reproductive strategy of a cold-temperate Zostera marina meadow. Estuaries Coasts 2016, 40, 553–563. [Google Scholar] [CrossRef]
- Hays, C.G.; Hanley, T.C.; Graves, R.M.; Schenck, F.R.; Hughes, A.R. Linking spatial patterns of adult and seed diversity across the depth gradient in the seagrass Zostera marina L. Estuaries Coasts 2020, 44, 383–395. [Google Scholar] [CrossRef]
- von Staats, D.A.; Hanley, T.C.; Hays, C.G.; Madden, S.R.; Sotka, E.E.; Hughes, A.R. Intra-meadow variation in seagrass flowering phenology across depths. Estuaries Coasts 2020, 44, 325–338. [Google Scholar] [CrossRef]
- Blok, S.E.; Olesen, B.; Krause-Jensen, D. Life history events of eelgrass Zostera marina L. Populations across gradients of latitude and temperature. Mar. Ecol. Prog. Ser. 2018, 590, 79–93. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reproductive Shoot Height (cm) | No. Spathes per Reproductive Shoot | No. Seeds per Spathe | No. Seeds per Reproductive Shoot | |
|---|---|---|---|---|
| Mean ± SD | 71.70 ± 12.57 | 23.40 ± 7.12 | 8.74 ± 1.57 | 204.73 ± 11.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Xu, S.; Yue, S.; Qiao, Y.; Zhang, Y.; Zhang, X.; Zhou, Y. Seed Provision Efficacy of Detached Reproductive Shoots in Restoration Projects for Degraded Eelgrass (Zostera marina L.) Meadows. Sustainability 2023, 15, 5904. https://doi.org/10.3390/su15075904
Liu M, Xu S, Yue S, Qiao Y, Zhang Y, Zhang X, Zhou Y. Seed Provision Efficacy of Detached Reproductive Shoots in Restoration Projects for Degraded Eelgrass (Zostera marina L.) Meadows. Sustainability. 2023; 15(7):5904. https://doi.org/10.3390/su15075904
Chicago/Turabian StyleLiu, Mingjie, Shaochun Xu, Shidong Yue, Yongliang Qiao, Yu Zhang, Xiaomei Zhang, and Yi Zhou. 2023. "Seed Provision Efficacy of Detached Reproductive Shoots in Restoration Projects for Degraded Eelgrass (Zostera marina L.) Meadows" Sustainability 15, no. 7: 5904. https://doi.org/10.3390/su15075904