
Prediction of Potential Distribution Area of Two Parapatric Species in Triosteum under Climate Change

Abstract
:1. Introduction
2. Materials and Methods
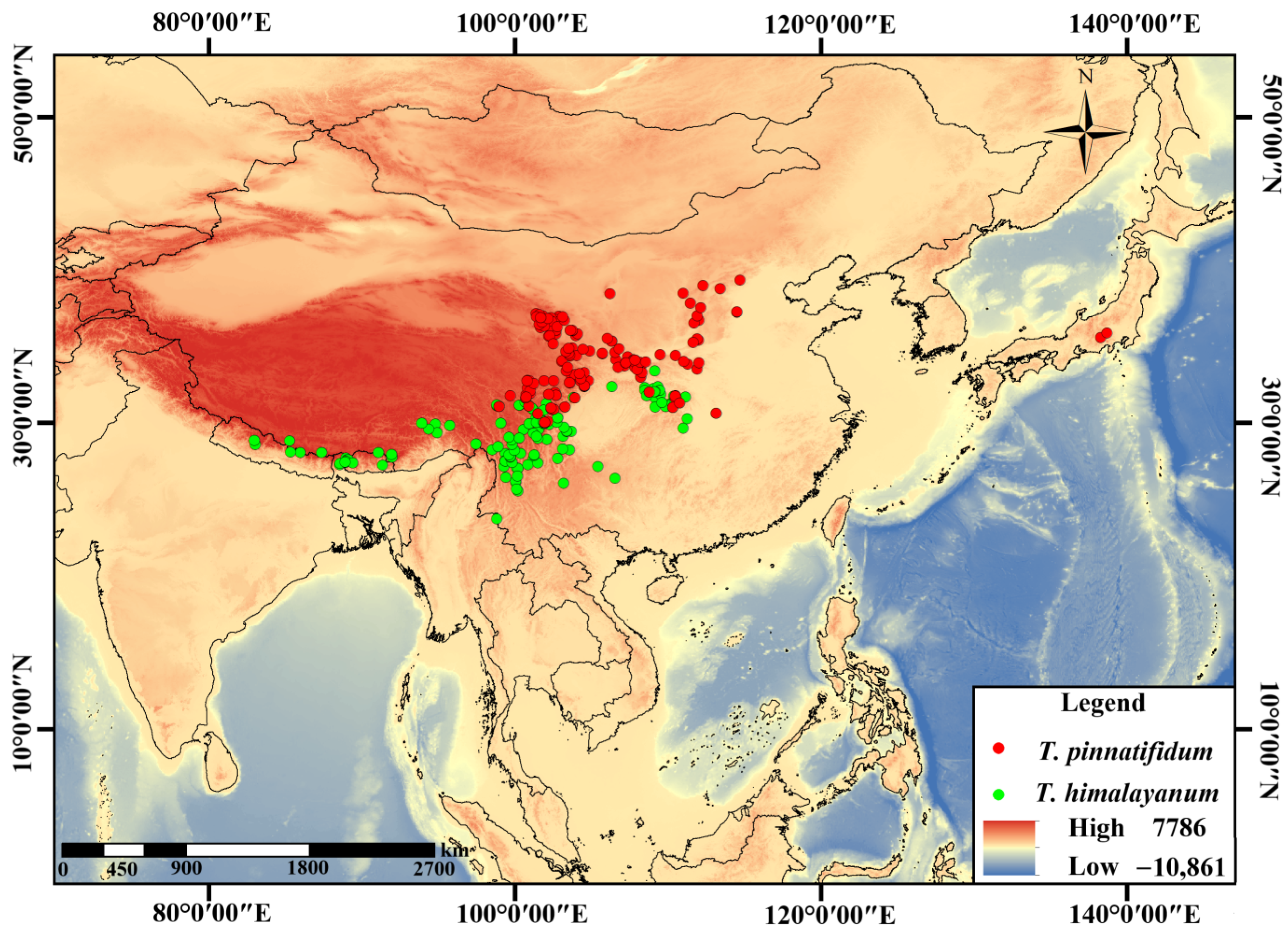
2.1. Species Occurrence Data Acquisition and Screening
2.2. Environmental Data Acquisition and Screening
2.3. Model Establishment, Optimization and Evaluation
2.4. Suitable Area Classification
2.5. Spatial Pattern Change of Suitable Areas
2.6. Geographic Distribution Centroid Calculation
2.7. Niche Overlap, Niche Breadth and Range Overlap Calculation
3. Results
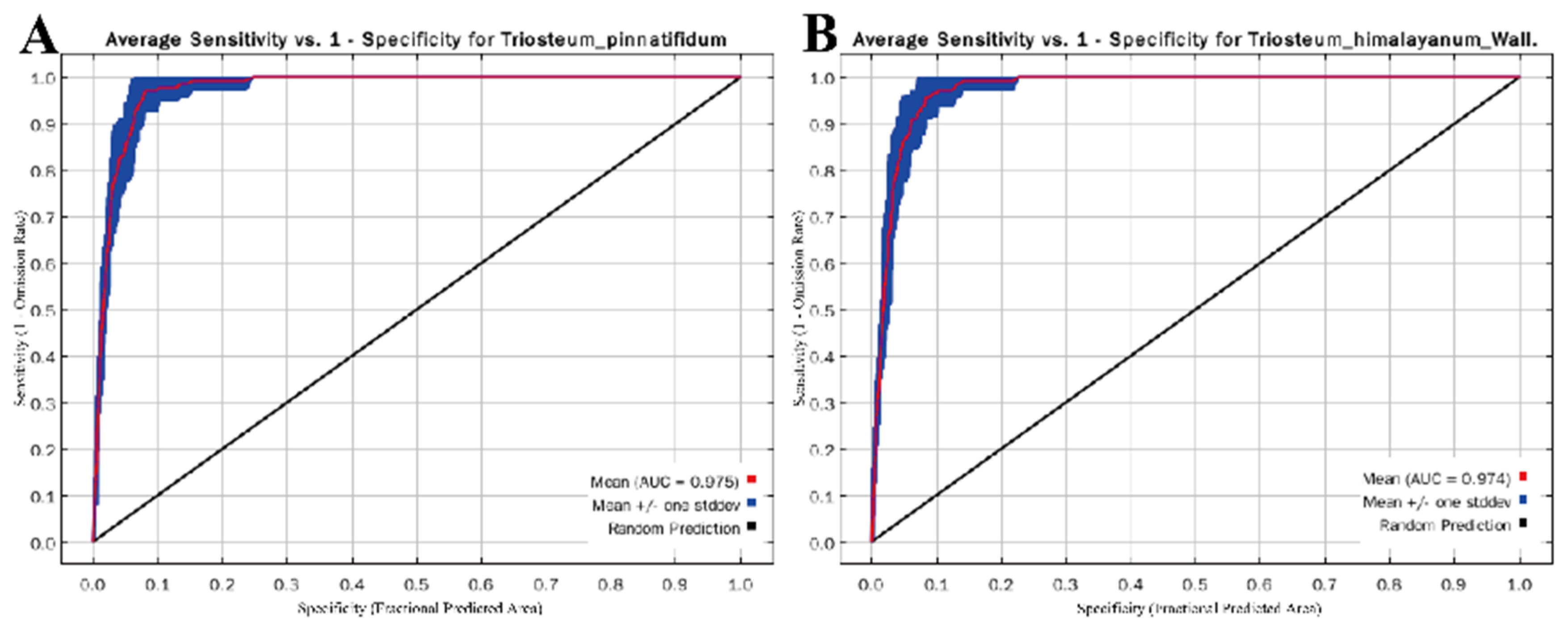
3.1. Model Optimization and Accuracy Evaluation
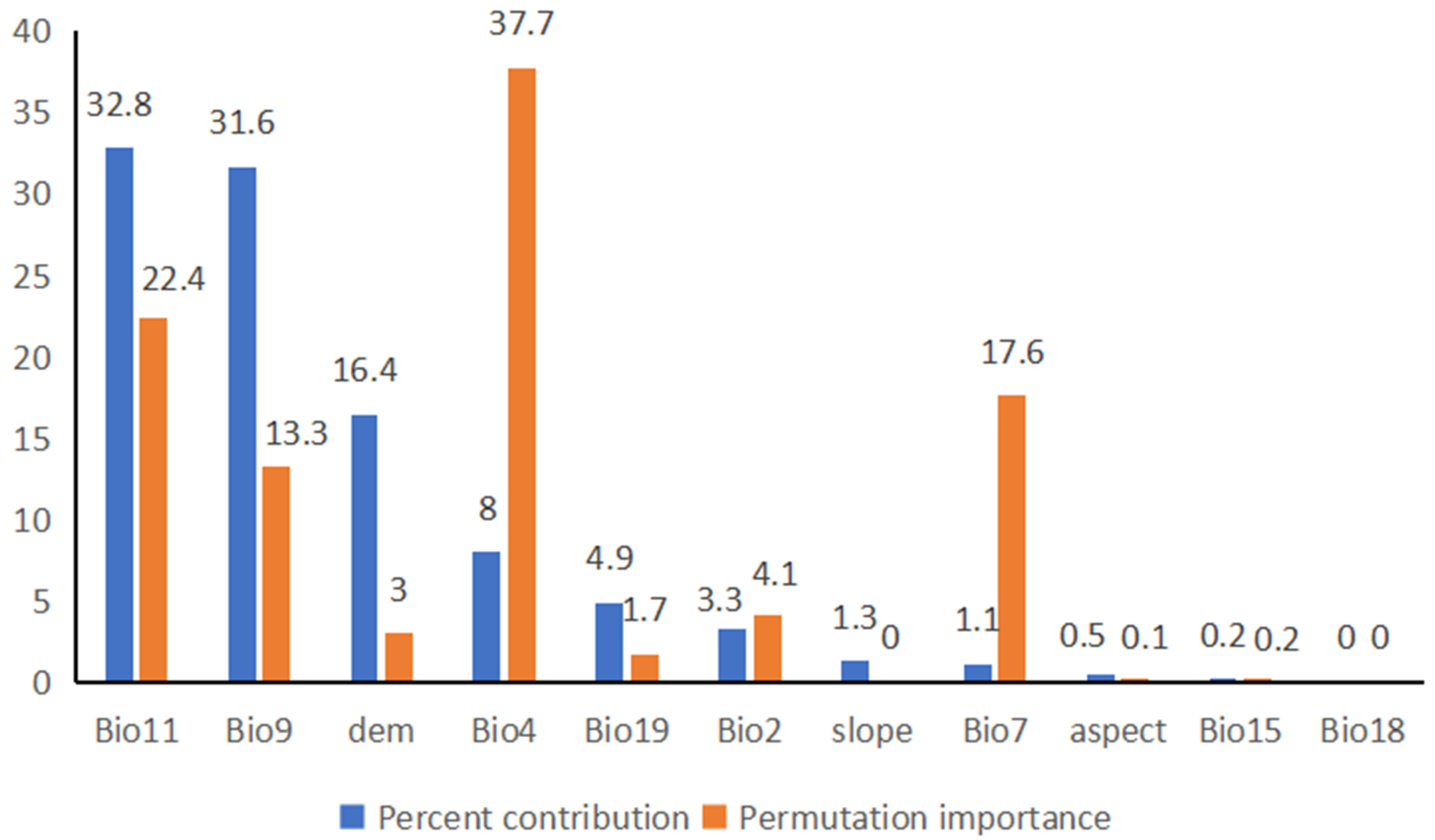
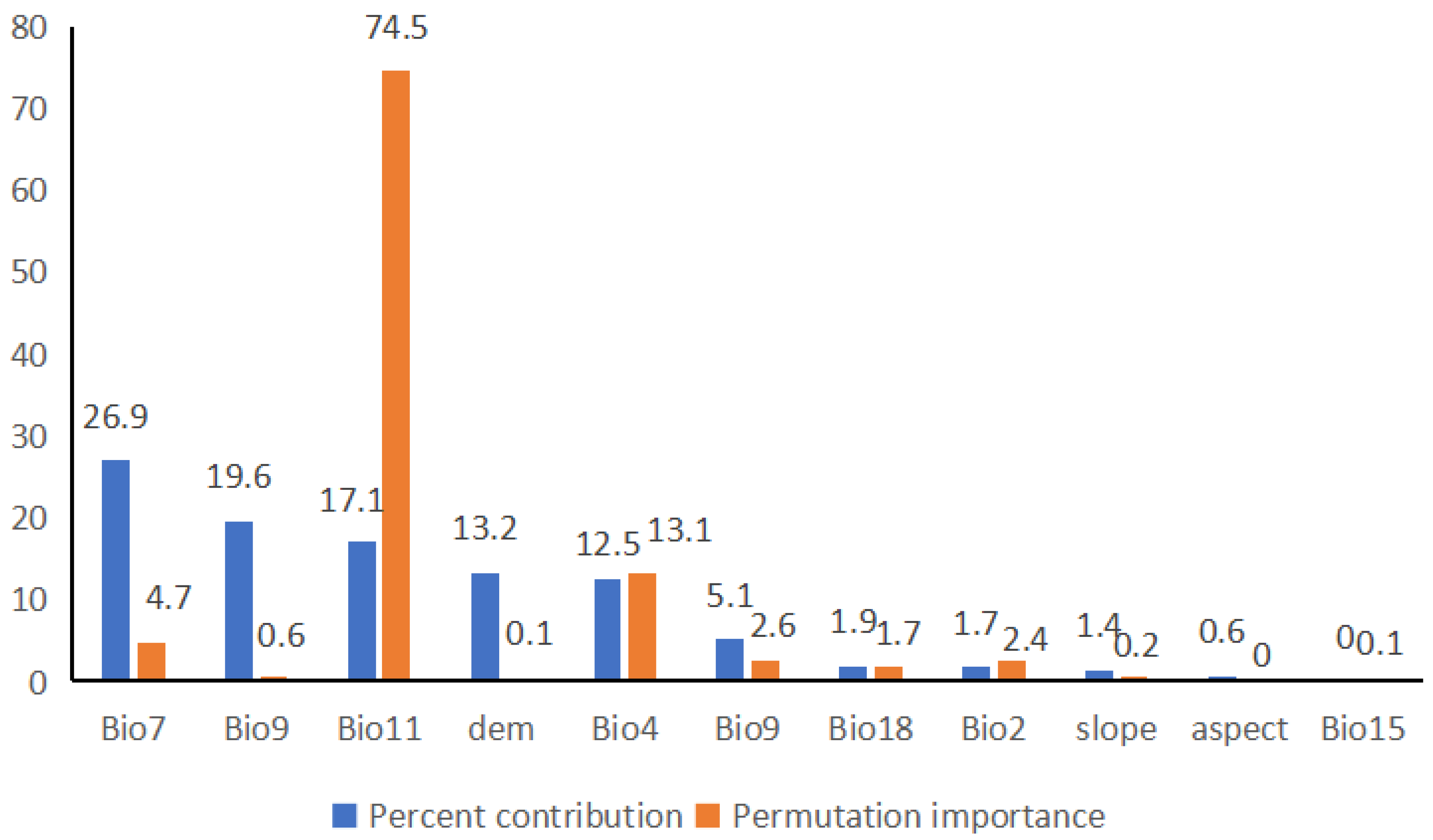
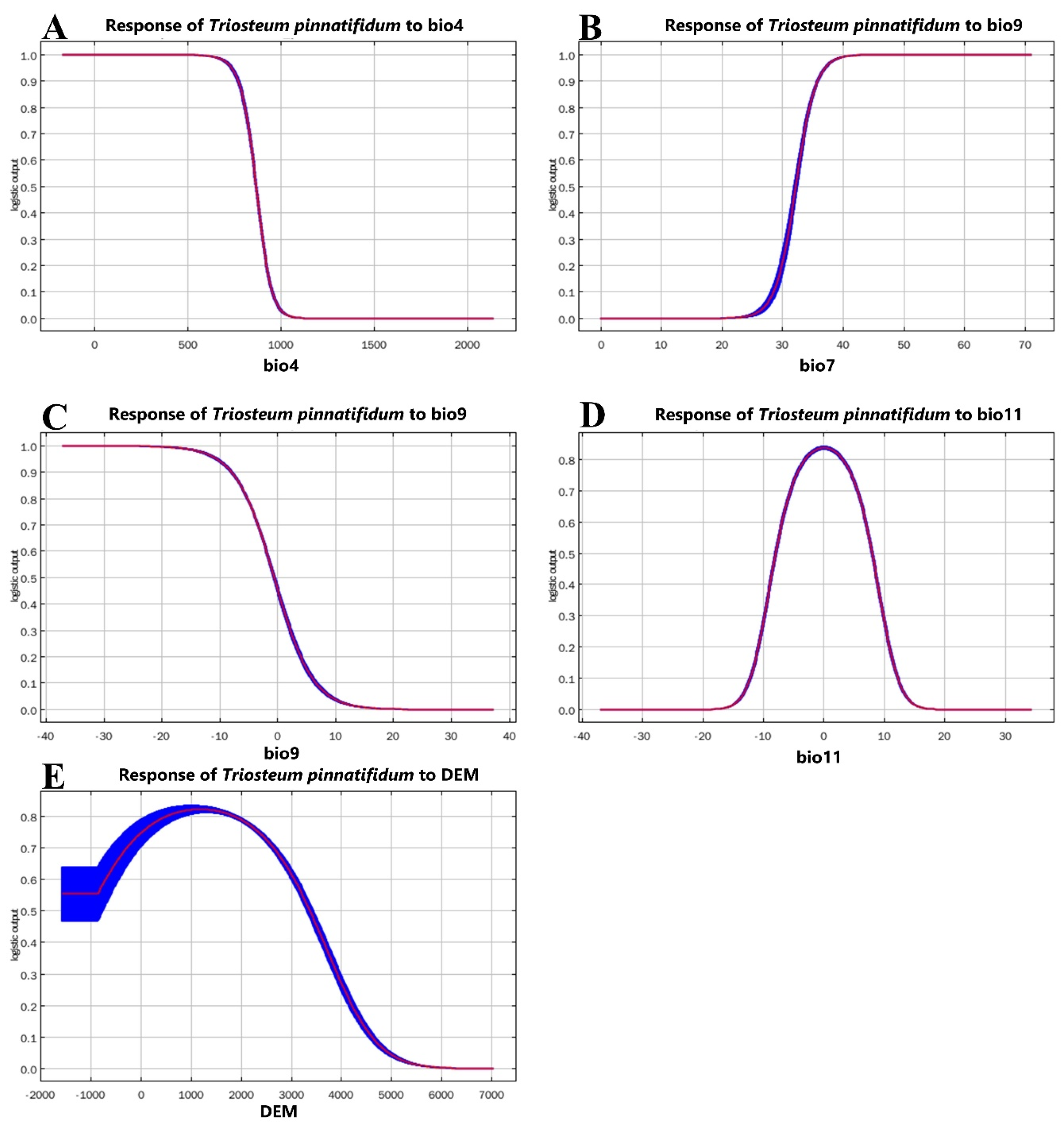
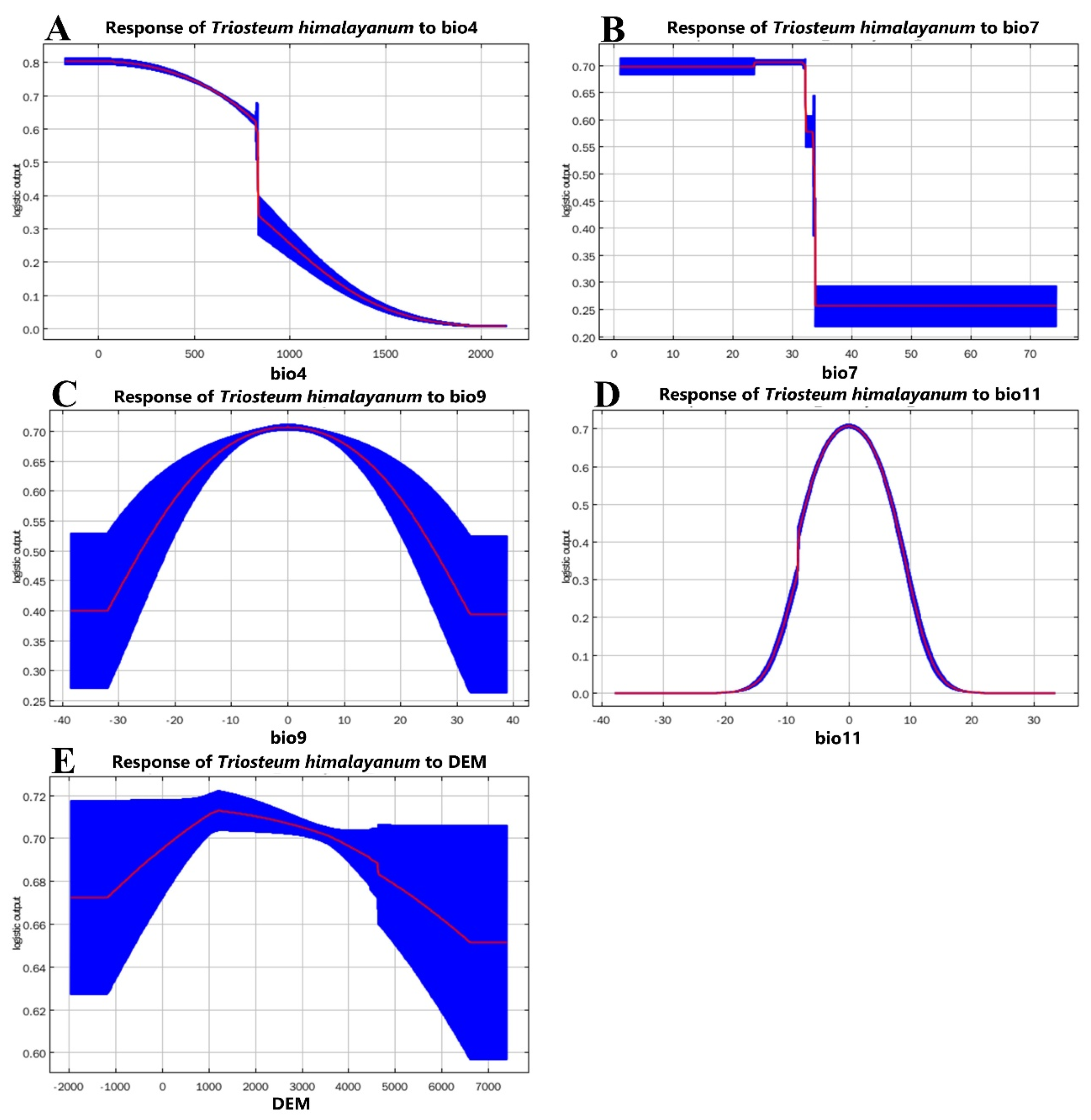
3.2. Importance of Environmental Variables
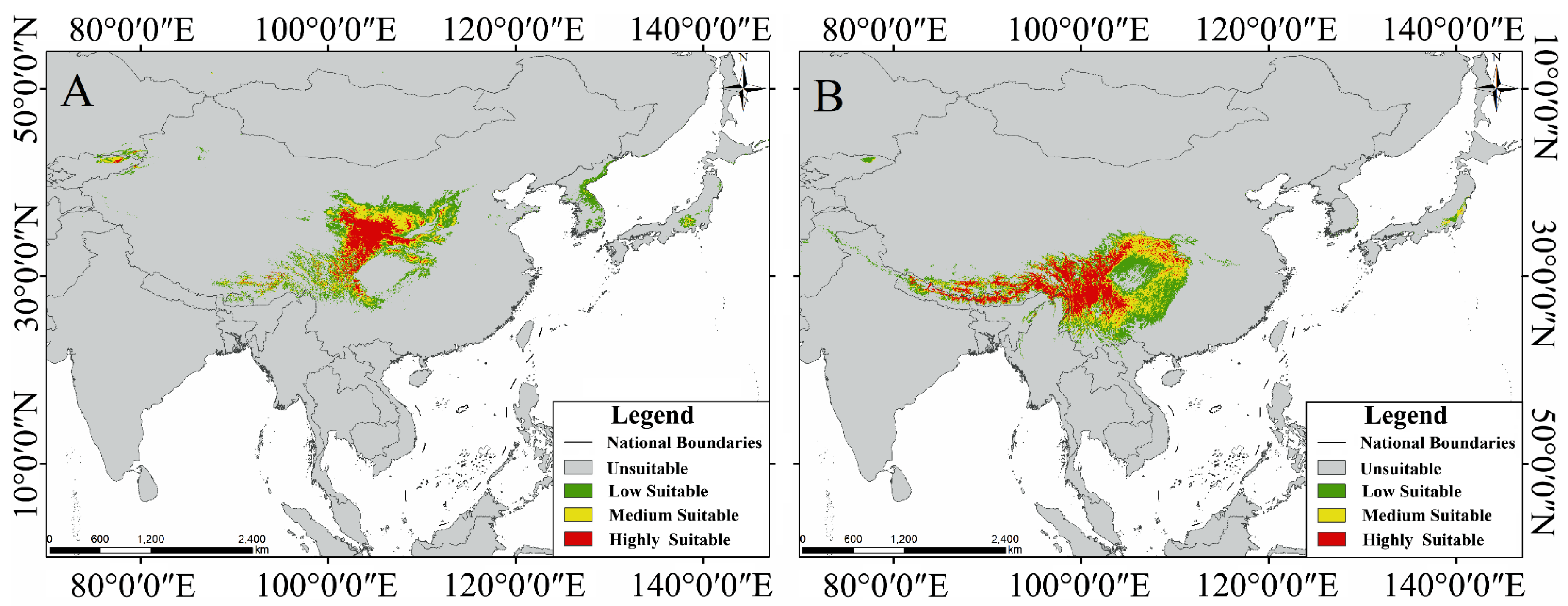
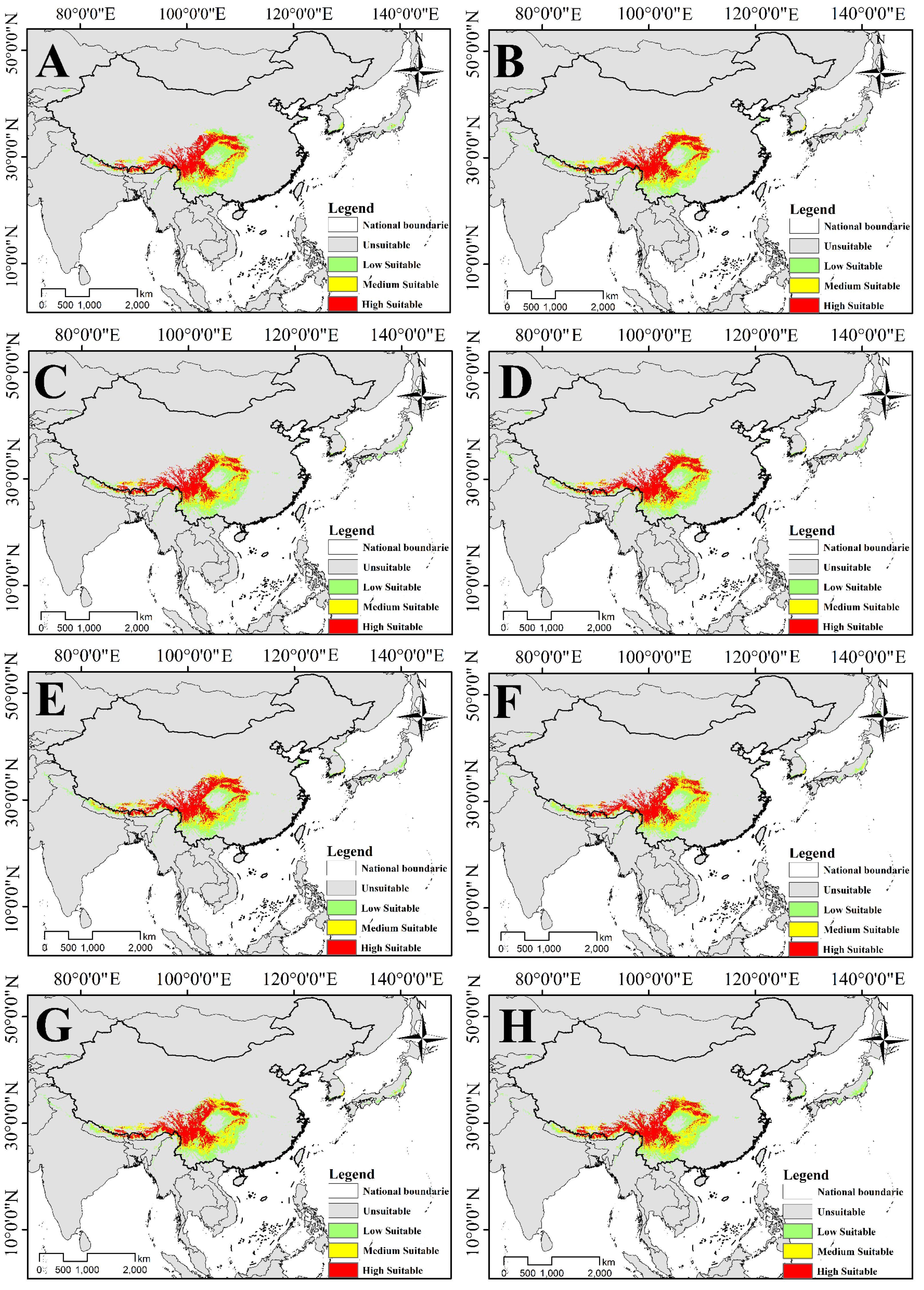
3.3. Potential Suitable Habitat for Two Species of Triosteum under Current Climate
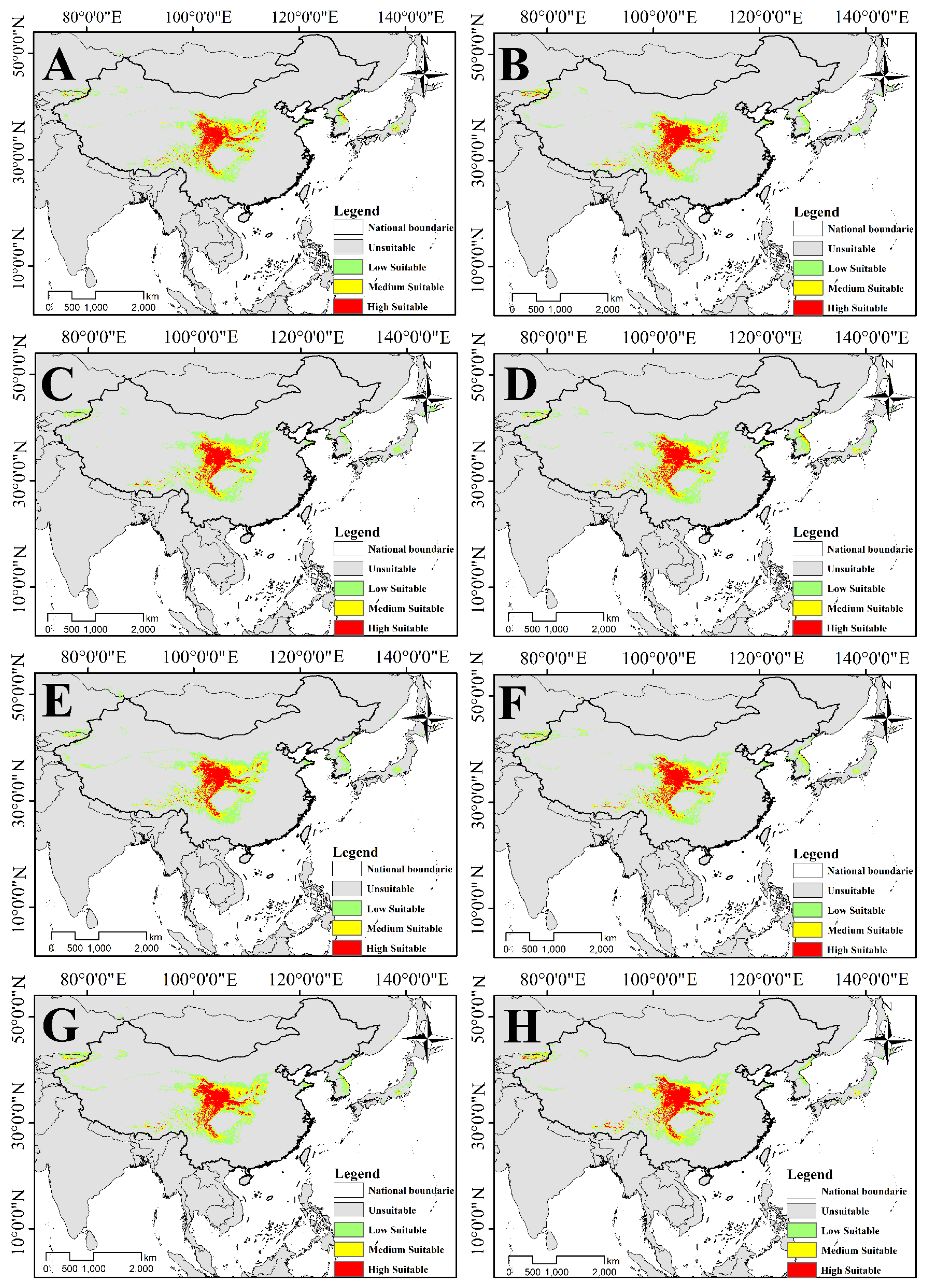
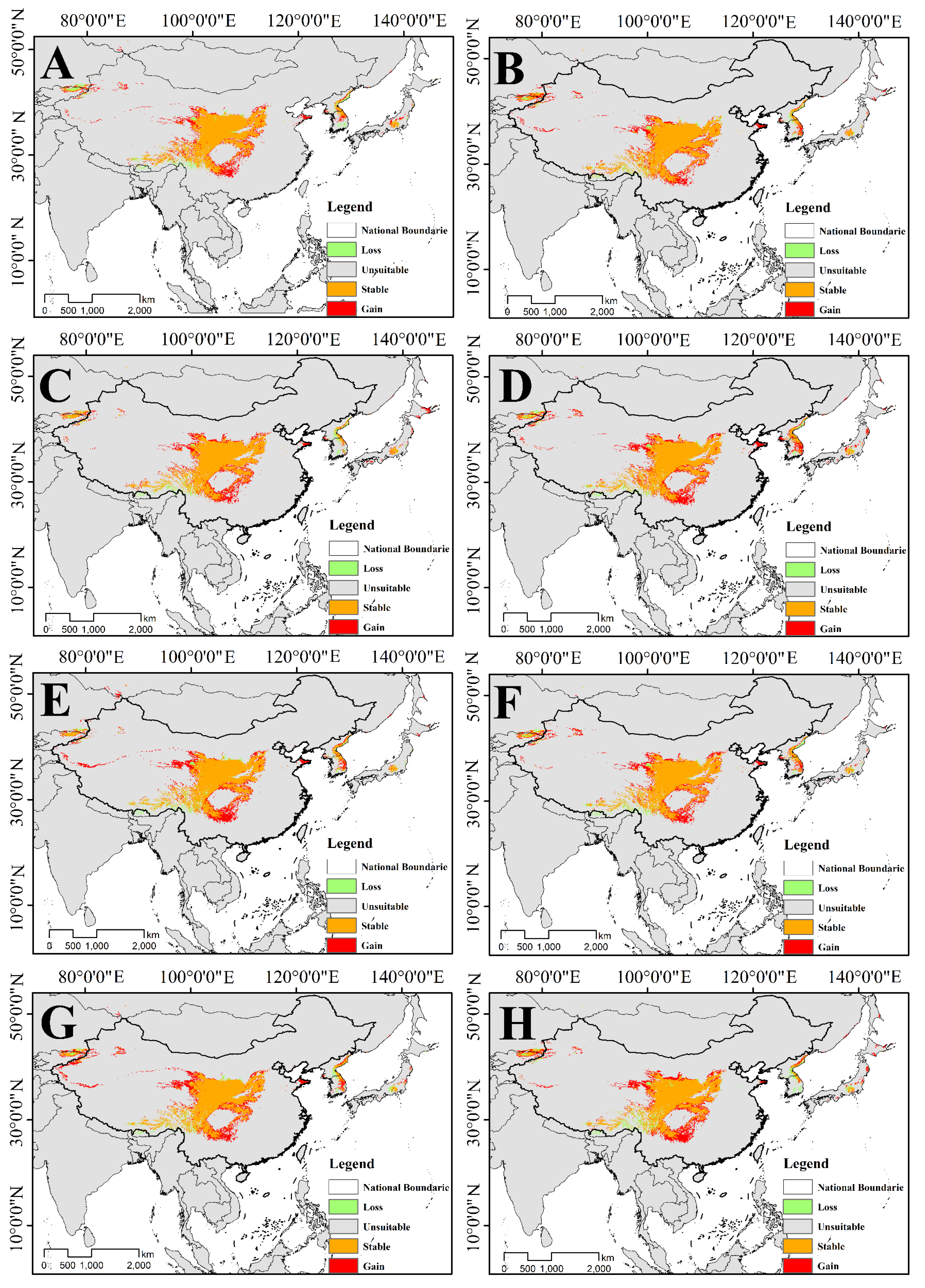
3.4. Distribution and Change of Future Potential Suitable Habitat
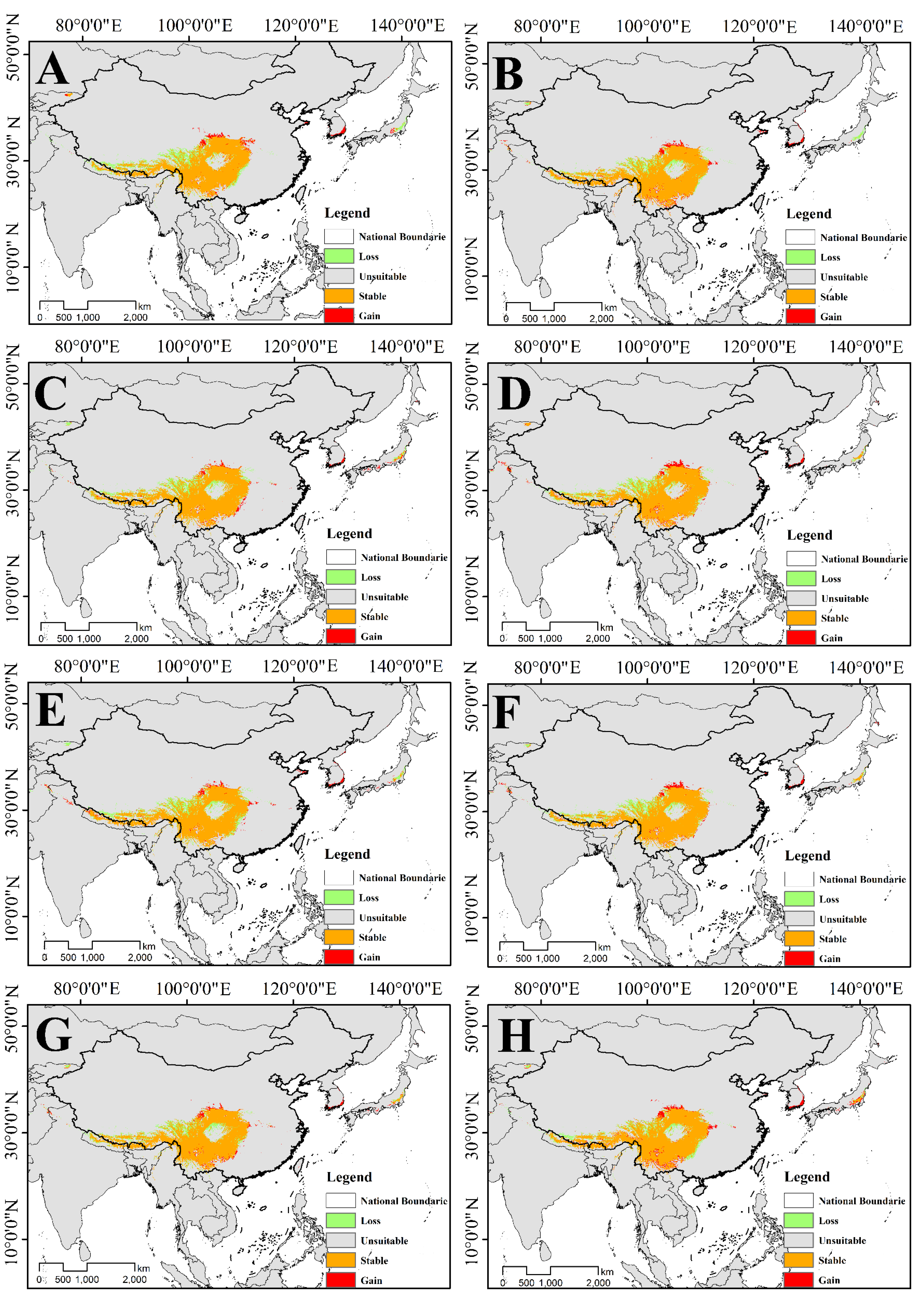
3.4.1. Changes in the Suitable Habitat of T. Pinnatifidum
3.4.2. Changes in the Suitable Habitat of T. himalayanum
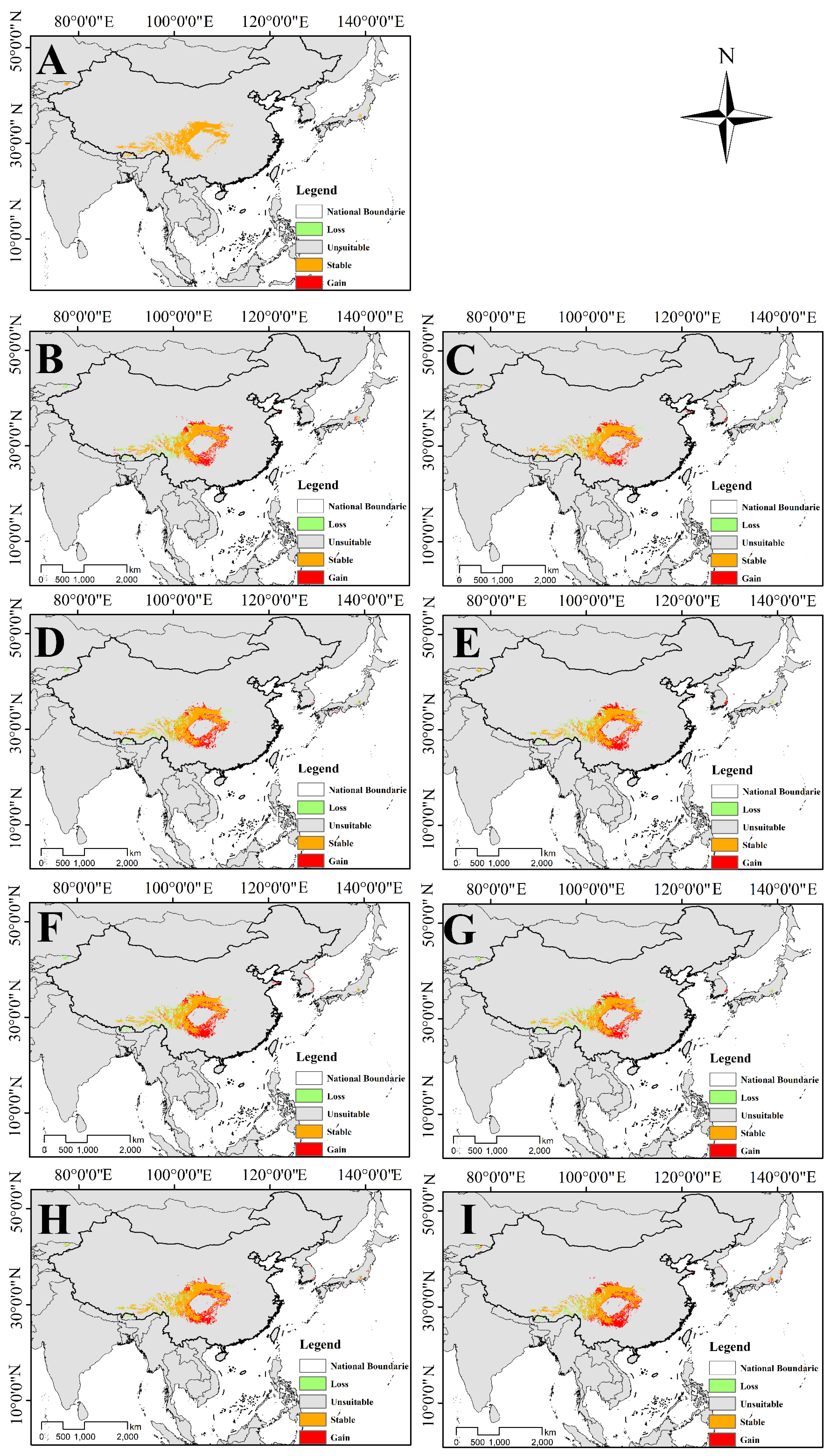
3.4.3. Changes in Overlapping Suitable Habitat of Two Species of Triosteum
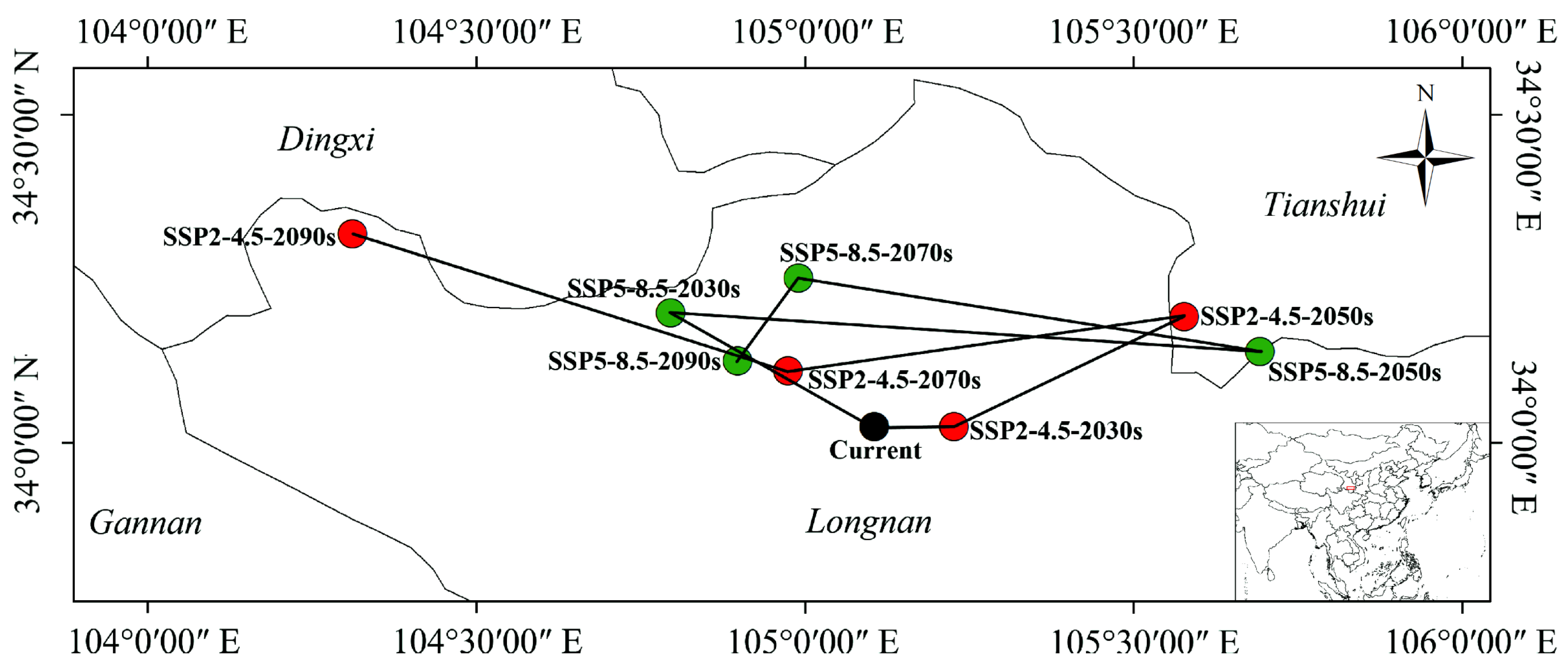
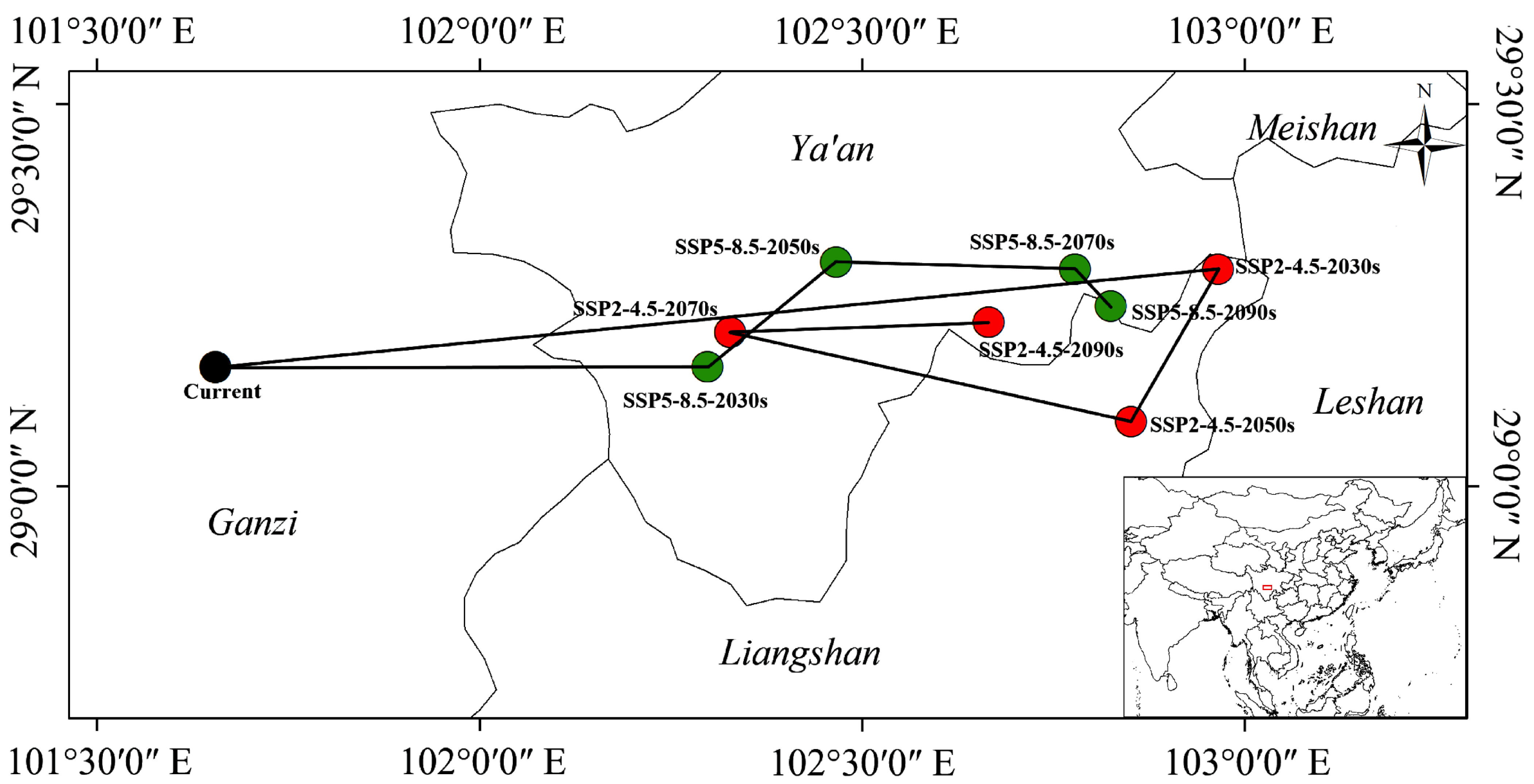
3.5. Changes in Suitable Habitat Centroid under Different Climatic Scenarios
3.6. Niche Analysis of T. Pinnatifidum and T. himalayanum
4. Discussion
4.1. Accuracy of Model Prediction
4.2. Effects of Environmental Variables on Species Distribution
4.3. Potential Suitability Analysis, Distribution Pattern and Centroid Changes of T. Pinnatifidum and T. himalayanum
4.4. Change of Potential Overlapping Distribution Area and Ecological Niche Analysis of Two Species of Triosteum
4.5. The Benefits and Limitations of the Modeling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, Q.; Mi, Z.; Lu, C.; Zhang, X.; Chen, L.; Wang, S.; Niu, J.; Wang, Z. Predicting potential distribution of Ziziphus spinosa (Bunge) H.H. Hu ex F.H. Chen in China under climate change scenarios. Ecol. Evol. 2022, 12, e8629. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, H.; Pan, H.; Shi, W.; Zhao, Y.; Li, S.; Liu, J.; Tao, J. Shifts in potential geographical distribution of Pterocarya stenoptera under climate change scenarios in China. Ecol. Evol. 2020, 10, 4828–4837. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Wang, S.; Duan, Y.; Han, J.; Huang, D.; Zhou, J. Current and future distribution of the deciduous shrub Hydrangea macrophylla in China estimated by MaxEnt. Ecol. Evol. 2021, 11, 16099–16112. [Google Scholar] [CrossRef] [PubMed]
- Arshad, F.; Waheed, M.; Fatima, K.; Harun, N.; Iqbal, M.; Fatima, K.; Umbreen, S. Predicting the Suitable Current and Future Potential Distribution of the Native Endangered Tree Tecomella undulata (Sm.) Seem. in Pakistan. Sustainability 2022, 14, 7215. [Google Scholar] [CrossRef]
- Grimm, N.B.; Chapin, F.S., III; Bierwagen, B.; Gonzalez, P.; Groffman, P.M.; Luo, Y.; Melton, F.; Nadelhoffer, K.; Pairis, A.; Raymond, P.A. The impacts of climate change on ecosystem structure and function. Front. Ecol. Environ. 2013, 11, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Guan, L.; Zhao, H.; Huang, Y.; Mou, Q.; Liu, K.; Chen, T.; Wang, X.; Zhang, Y.; Wei, B.; et al. Modeling habitat suitability of Houttuynia cordata Thunb (Ceercao) using MaxEnt under climate change in China. Ecol. Inform. 2021, 63, 101324. [Google Scholar] [CrossRef]
- Soilhia, Z.; Sayaria, N.; Benalouacheb, N.; Mekkia, M. Predicting current and future distributions of Mentha pulegium L. in Tunisia under climate change conditions, using the MaxEnt model. Ecol. Inform. 2022, 68, 101533. [Google Scholar] [CrossRef]
- Koo, K.A.; Park, S.U.; Kong, W.S.; Hong, S.; Jang, I.; Seo, C. Potential climate change effects on tree distributions in the Korean Peninsula: Understanding model & climate uncertainties. Ecol. Model. 2017, 353, 17–27. [Google Scholar] [CrossRef]
- Zhao, Y.; Deng, X.; Xiang, W.; Chen, L.; Ouyang, S. Predicting potential suitable habitats of Chinese fir under current and future climatic scenarios based on Maxent model. Ecol. Inform. 2021, 64, 101393. [Google Scholar] [CrossRef]
- Zhao, R.; He, Q.; Chu, X.; Lu, Z.; Zhu, Z. Prediction of potential distribution of Carpinus cordata in China under climate change. Chin. J. Appl. Ecol. 2019, 30, 3833–3843. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change onthe potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Boyce, M.; McDonald, L. Relating populations to habitats using resource selection functions. Trends Ecol Evol. 1999, 14, 268–272. [Google Scholar] [CrossRef] [PubMed]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models; Chapman & Hall: London, UK, 1989. [Google Scholar]
- Ripley, B.D. Pattern Recognition and Neural Networks; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Breiman, L.; Friedman, J.; Stone, C.J.; Olshen, R.A. Classification and Regression Trees; Chapman & Hall: London, UK, 1984. [Google Scholar]
- Phillips, S.J.; Dudı’k, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; p. 83. [Google Scholar]
- Wang, G.; Wang, C.; Guo, Z.; Dai, L.; Wu, Y.; Liu, H.; Li, Y.; Chen, H.; Zhang, Y.; Zhao, Y.; et al. Integrating Maxent model and landscape ecology theory for studying spatiotemporal dynamics of habitat: Suggestions for conservation of endangered Red-crowned crane. Ecol. Indic. 2020, 116, 106427. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Ye, X.; Zhang, M.; Lai, W.; Yang, M.; Fan, H.; Chen, S.; Liu, B. Prediction of potential suitable distribution of Phoebe bournei based on MaxEnt optimization model. Acta Ecol. Sin. 2021, 41, 8135–8144. [Google Scholar] [CrossRef]
- Qi, S.; Luo, W.; Chen, K.L.; Li, X.; Luo, H.L.; Yang, Z.Q.; Yin, D.M. The Prediction of the Potentially Suitable Distribution Area of Cinnamomum mairei H. Lév in China Based on the MaxEnt Model. Sustainability 2022, 14, 7682. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Y.; Zeng, H. Application of the MaxEnt model in improving the accuracy of ecological red line identification: A case study of Zhanjiang, China. Ecol. Indic. 2022, 137, 108767. [Google Scholar] [CrossRef]
- Xu, B.S.; Hu, J.Q.; Wang, J.H. (Eds.) Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1988; pp. 72–105. [Google Scholar]
- Cao, D.; LI, T.; WU, L.; Ge, L.; Lei, Y. Study on the morphology and histology of Triosteum pinnatifidum. West China J. Pharm. Sci. 2014, 29, 056–058. [Google Scholar] [CrossRef]
- Liu, H.R.; Khan, G.; Gao, Q.B.; Zhang, F.Q.; Liu, W.H.; Wang, Y.F.; Fang, J.; Chen, S.L.; Afridi, S.G. Dispersal into the Qinghai–Tibet plateau: Evidence from the genetic structure and demography of the alpine plant Triosteum pinnatifidum. PeerJ 2022, 10, e12754. [Google Scholar] [CrossRef]
- Liu, H.R.; Gao, Q.B.; Zhang, F.Q.; Khan, G.; Chen, S.L. Westwards and northwards dispersal of Triosteum himalayanum (Caprifoliaceae) from the Hengduan Mountains region based on chloroplast DNA phylogeography. PeerJ 2018, 6, e4748. [Google Scholar] [CrossRef] [Green Version]
- Braunisch, V.; Suchant, R. Predicting species distributions based on incomplete survey data: The trade-off between precision andscale. Ecography 2010, 33, 826–840. [Google Scholar] [CrossRef]
- Hefley, T.J.; Baasch, D.M.; Tyre, A.J.; Blankenship, E.E. Correction of location errors for presence-only species distribution models. Methods Ecol. Evol. 2014, 5, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.F.; Momohara, A.; Herrando-Moraira, S.; Qian, S.; Yang, Y.; Ohsawa, M.; Luu, H.T.; et al. Identifying long-term stable refugia for relict plant species in East Asia. Nat. Commun. 2018, 9, 4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fick, S.E.; Hijmans, R. WorldClim 2: New 1 km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Wu, T.; Lu, Y.; Fang, Y.; Xin, X.; Li, L.; Jie, W.; Zhang, J.; Liu, Y.; Zhang, L.; Zhang, F.; et al. The Beijing Climate Center Climate System Model (BCC-CSM): The main progress from CMIP5 to CMIP6. Geosci. Model Dev. 2019, 12, 1573–1600. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, B.C.; Tebaldi, C.; van Vuuren, D.P.; Eyring, V.; Friedlingstein, P.; Hurtt, G.; Knutti, R.; Kriegler, E.; Lamarque, J.F.; Lowe, J.; et al. The Scenario Model Intercomparison Project (ScenarioMIP) for CMIP6. Geosci. Model Dev. 2016, 9, 3461–3482. [Google Scholar] [CrossRef] [Green Version]
- IBM Corp. IBM SPSS Statistics for Windows, Version 22.0; IBM Corp: Armonk, NY, USA, 2013. [Google Scholar]
- Cao, Z.; Zhang, L.; Zhang, X.; Guo, Z. Predicting the Potential Distribution of Hylomecon japonica in China under Current and Future Climate Change Based on Maxent Model. Sustainability 2021, 13, 11253. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better MAXENT models of species distributions: Complexity, overfitting and evaluation. J. Evol. Biol. 2013, 41, 629–943. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. Kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Li, Z.; Liu, Z.; Yang, Y.; Khoso, A.G.; Wang, L.; Liu, D. Climate change simulations revealed potentially drastic shifts in insect community structure and crop yields in China’s farmland. J. Pest Sci. 2023, 96, 55–69. [Google Scholar] [CrossRef]
- Phillips, S.J. Transferability, sample selection bias and background data in presence-only modeling: A response to Peterson et al. and (2007). Ecography 2008, 31, 272–278. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the measurement of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Swets, J. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, Z.; Jian, S. Simulation of potential suitable distribution area of Pinus tabuliformis in Yiluo River basin on MaxEnt Model. J. Zhejiang For. Sci. Technol. 2023, 43, 1–8. [Google Scholar]
- Jiang, J.M.; Jin, L.; Huang, L.; Wang, W.T. The Future Climate under Different CO2 Emission Scenarios Significantly Influences the Potential Distribution of Achnatherum inebrians in China. Sustainability 2022, 14, 4806. [Google Scholar] [CrossRef]
- Silvertown, J.W. The distribution of plants in limestone pavement: Tests of species interaction and niche separation against null hypotheses. J. Ecol. 1983, 71, 819–828. [Google Scholar] [CrossRef]
- Dong, H.; Zhang, N.; Shen, S.; Zhu, S.; Fan, S.; Lu, Y. Effects of Climate Change on the Spatial Distribution of the Threatened Species Rhododendron purdomii in Qinling-Daba Mountains of Central China: Implications for Conservation. Sustainability 2023, 15, 3181. [Google Scholar] [CrossRef]
- Song, D.; Li, Z.; Wang, T.; Qi, Y.; Han, H.; Chen, Z. Prediction of Changes to the Suitable Distribution Area of Fritillaria przewalskii Maxim. in the Qinghai-Tibet Plateau under Shared Socioeconomic Pathways (SSPs). Sustainability 2023, 15, 2833. [Google Scholar] [CrossRef]
- Peterson, A.T. Ecological niche conservatism: A time-structured review of evidence. J. Evol. Biol. 2011, 38, 817–827. [Google Scholar] [CrossRef]
- Araújo, M.B.; Guisan, A. Five (or so) challenges for species distribution modelling. J. Biogeogr. 2006, 33, 1677–1688. [Google Scholar] [CrossRef]
- Anderson, R.P.; Gonzalez, I. Species-specific tuning increases robustness to sampling bias in models of species distributions: An implementation with Maxent. Ecol. Model. 2011, 222, 2796–2811. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, L.; Wang, J.; Wang, W.; Niyati, N.; Guo, Y.; Wang, X. Chinese caterpillar fungus (Ophiocordyceps sinensis) in China: Current distribution, trading, and futures under climate change and overexploitation. Sci. Total Environ. 2021, 755, 142548. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Bai, Y.; Alatalo, J.M.; Huang, Z.; Yang, F.; Pu, X.; Wang, R.; Yang, W.; Guo, X. Spatio- temporal variation in potential habitats for rare and endangered plants and habitat conservation based on the maximum entropy model. Sci. Total Environ. 2021, 784, 147080. [Google Scholar] [CrossRef]
- Anand, V.; Oinam, B.; Singh, I.H. Predicting the current and future potential spatial distribution of endangered Rucervus eldii eldii (Sangai) using MaxEnt model. Environ. Monit. Assess. 2021, 193, 147. [Google Scholar] [CrossRef]
- Mafuwe, K.; Broadley, S.; Moyo, S. Use of maximum entropy (Maxent) niche modelling to predict the occurrence of threatened freshwater species in a biodiversity hotspot of Zimbabwe. Afr. J. Ecol. 2021, 60, 557–565. [Google Scholar] [CrossRef]
- Abeli, T.; Ghitti, M.; Socchi, R. Does ecological marginality reflect physiological marginality in plants? Plant Biosyst. 2020, 154, 149–157. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, Y.; Huang, D.; Wang, H.; Cao, Q.; Fan, P.; Yang, N.; Zheng, P.; Wang, R. The effect of climate change on the richness distribution pattern of oaks (Quercus L.) in China. Sci. Total Environ. 2020, 744, 140786. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Chen, M.; Zheng, N.; Xu, Z.; Zhang, J.; Hu, X.; Li, L.; Gu, R.; Du, X.; Wang, J. Transcriptome Analysis Identifies Novel Genes Associated with Low-Temperature Seed Germination in Sweet Corn. Plants 2023, 12, 159. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, W.; Jiang, M.; Yang, L.; Zhou, X. QTL mapping for low temperature germination in rapeseed. Sci. Rep. 2021, 11, 23382. [Google Scholar] [CrossRef]
- Fang, Z.Z.; Wang, L.K.; Dai, H.; Zhou, R.D.; Jiang, C.C.; Espley, V.R.; Deng, C.; Lin, Y.J.; Pan, S.L.; Ye, X.F. The genome of low-chill Chinese plum “Sanyueli” (Prunus salicina Lindl.) provides insights into the regulation of the chilling requirement of flower buds. Mol. Ecol. Resour. 2021, 22, 1919–1938. [Google Scholar] [CrossRef] [PubMed]
- Kazakis, G.; Ghosn, D.; Vogiatzakis, I.N.; Papanastasis, V.P. Vascular plant diversity and climate change in the alpine zone of the Lefka Ori, Crete. Biodivers. Conserv. 2007, 16, 1603–1615. [Google Scholar] [CrossRef]
- Pickering, C.; Hill, W.; Green, K. Vascular plant diversity and climate change in the alpine zone of the Snowy Mountains, Australia. Biodivers. Conserv. 2008, 17, 1627–1644. [Google Scholar] [CrossRef] [Green Version]
- Klanderud, K.; Totland, Ø. Simulated Climate Change Altered Dominance Hierarchies and Diversity of an Alpine Biodiversity Hotspot. Ecology 2005, 86, 2047–2054. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, U.; Ali, H.; Chaudry, M.N.; Ashraf, I.; Batool, A.; Saqib, Z. Predicting the potential distribution of Olea ferruginea in Pakistan incorporating climate change by using Maxent model. Sustainability 2016, 8, 722. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Shen, X.; Tong, L.; Lei, F.; Mu, X.; Zhang, Z. Impact of past and future climate change on the potential distribution of an endangered Montane Shrub Lonicera oblata and its conservation implications. Forests 2021, 12, 125. [Google Scholar] [CrossRef]
- Wang, R.; Li, Q.; He, S.; Liu, Y. Potential distribution of Actinidia chinensis in China and its predicted response to climate change. Chin. J. Eco- Agric. 2018, 26, 27–37. [Google Scholar] [CrossRef]
- Wang, L.; Wu, X.; Li, Y.; Xu, X. Prediction of suitable cultivation area for Halesia carolina L. in China. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2018, 42, 10–16. [Google Scholar] [CrossRef]
- Gould, K.R.; Donoghue, M.J. Phylogeny and biogeography of Triosteum. Harv. Pap. Botany. 2000, 5, 157–166. [Google Scholar]
- Jing, G.H.; Cheng, J.M.; Su, J.S.; Wei, L.; Shi, X.X.; Jin, J.W. Response of dominant population niche breadths and niche overlaps to various disturbance factors in typical steppe fenced grassland of China’s Loess Plateau region. Acta Pratacult. Sin. 2015, 24, 43–52. [Google Scholar] [CrossRef]
- Li, M.; Jiang, D.M.; Toshio, O.; Zhou, Q.L.; Luo, Y.M. Niche characteristic of herbages in artificial sand-fixing communities in Horqin sandy land. Pratacult. Sci. 2009, 26, 10–16. [Google Scholar]
- Jiao, S.W.; Qing, Z.; Sun, G.Q.; Lei, G.C. Improving conservation of cranes by modeling potential wintering distributions in China. J. Resour. Ecol. 2016, 7, 44–50. [Google Scholar] [CrossRef]
- Deb, J.C.; Phinn, S.; Butt, N.; McAlpin, C.A. Climatic-induced shifts in the distribution of teak (Tectona grandis) in tropical Asia: Implications for forest management and planning. Environ. Manag. 2017, 60, 422–435. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description | Units |
|---|---|---|
| Bio1 | Annual mean temperature | °C |
| Bio2 | Mean diurnal range | °C |
| Bio3 | Isothermality | / |
| Bio4 | Temperature seasonality | / |
| Bio5 | Max Temperature of Warmest Month | °C |
| Bio6 | Min temperature for coldest month | °C |
| Bio7 | Temperature annual range | °C |
| Bio8 | Mean temperature of wettest quarter | °C |
| Bio9 | Mean temperature of driest quarter | °C |
| Bio10 | Mean temperature of warmest quarter | °C |
| Bio11 | Mean temperature of coldest quarter | °C |
| Bio12 | Annual precipitation | mm |
| Bio13 | Precipitation of wettest month | mm |
| Bio14 | Precipitation of driest month | mm |
| Bio15 | Precipitation seasonality | / |
| Bio16 | Precipitation of wettest quarter | mm |
| Bio17 | Precipitation of driest quarter | mm |
| Bio18 | Precipitation of warmest quarter | mm |
| Bio19 | Precipitation of coldest quarter | mm |
| dem | Elevation | m |
| slope | Slope | ° |
| aspect | Aspect | ° |
| Species | Period | Predicted Area (×104 km2) and % of the Corresponding Current Area | |||
|---|---|---|---|---|---|
| Low Suitable Habitat | Medium Suitable Habitat | Highly Suitable Habitat | Total Suitable Habitat | ||
| T. Pinnatifidum | Present | 56.11 | 30.48 | 24.44 | 111.03 |
| 2030S(SSP2-4.5) | 69.03 (123.02%) | 38.34 (125.79%) | 29.34 (120.05%) | 136.71 (123.13%) | |
| 2030S(SSP5-8.5) | 74.36 (132.53%) | 38.79 (127.26%) | 32.80 (134.21%) | 145.95 (131.45%) | |
| 2050S(SSP2-4.5) | 77.79 (138.64%) | 36.77 (120.64%) | 26.97 (110.35%) | 141.53 (127.47%) | |
| 2050S(SSP5-8.5) | 79.59 (141.85%) | 40.68 (133.46%) | 31.18 (127.58%) | 151.45 (136.40%) | |
| 2070S(SSP2-4.5) | 76.66 (136.62%) | 38.75 (127.13%) | 30.25 (123.77%) | 145.66 (131.19%) | |
| 2070S(SSP5-8.5) | 74.39 (132.58%) | 38.52 (126.38%) | 30.08 (123.08%) | 142.99 (128.79%) | |
| 2090S(SSP2-4.5) | 77.11 (137.43%) | 40.59 (133.17%) | 31.40 (128.48%) | 149.10 (134.29%) | |
| 2090S(SSP5-8.5) | 70.19 (125.09%) | 39.96 (131.10%) | 36.01 (147.34%) | 146.16 (131.64%) | |
| T. himalayanum | Present | 58.41 | 45.74 | 41.53 | 145.68 |
| 2030S(SSP2-4.5) | 48.98 (83.86%) | 31.96 (69.87%) | 50.24 (120.97%) | 131.18 (90.05%) | |
| 2030S(SSP5-8.5) | 48.05 (82.26%) | 37.39 (81.74%) | 53.77 (129.47%) | 139.21 (95.56%) | |
| 2050S(SSP2-4.5) | 51.07 (87.43%) | 34.24 (74.86%) | 50.00 (120.39%) | 135.31 (92.88%) | |
| 2050S(SSP5-8.5) | 50.04 (85.67%) | 33.31 (72.82%) | 53.80 (129.54%) | 137.15 (94.14%) | |
| 2070S(SSP2-4.5) | 48.43 (82.91%) | 31.78 (69.48%) | 53.04 (127.71%) | 133.25 (91.47%) | |
| 2070S(SSP5-8.5) | 50.76 (86.90%) | 34.96 (76.43%) | 48.56 (116.93%) | 134.28 (92.17%) | |
| 2090S(SSP2-4.5) | 51.29 (87.81%) | 36.39 (79.56%) | 49.98 (120.35%) | 137.66 (94.49%) | |
| 2090S(SSP5-8.5) | 56.27 (96.34%) | 35.69 (78.03%) | 54.37 (130.92%) | 146.33 (100.45%) | |
| Niche Overlap | Range Overlap | Niche Breadth | |
|---|---|---|---|
| D | I | ||
| 0.439597 | 0.714068 | 0.455649 | 0.851083/0.858738 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Yao, Z.; Yuan, Q.; Xing, R.; Guo, Y.; Zhang, D.; Ahmad, I.; Liu, W.; Liu, H. Prediction of Potential Distribution Area of Two Parapatric Species in Triosteum under Climate Change. Sustainability 2023, 15, 5604. https://doi.org/10.3390/su15065604
Li X, Yao Z, Yuan Q, Xing R, Guo Y, Zhang D, Ahmad I, Liu W, Liu H. Prediction of Potential Distribution Area of Two Parapatric Species in Triosteum under Climate Change. Sustainability. 2023; 15(6):5604. https://doi.org/10.3390/su15065604
Chicago/Turabian StyleLi, Xumin, Zhiwen Yao, Qing Yuan, Rui Xing, Yuqin Guo, Dejun Zhang, Israr Ahmad, Wenhui Liu, and Hairui Liu. 2023. "Prediction of Potential Distribution Area of Two Parapatric Species in Triosteum under Climate Change" Sustainability 15, no. 6: 5604. https://doi.org/10.3390/su15065604