
Effect of Water Level Reduction on the Littoral Zone in Terms of Its Efficiency in Lake Protection

Abstract
:1. Introduction
2. Methods
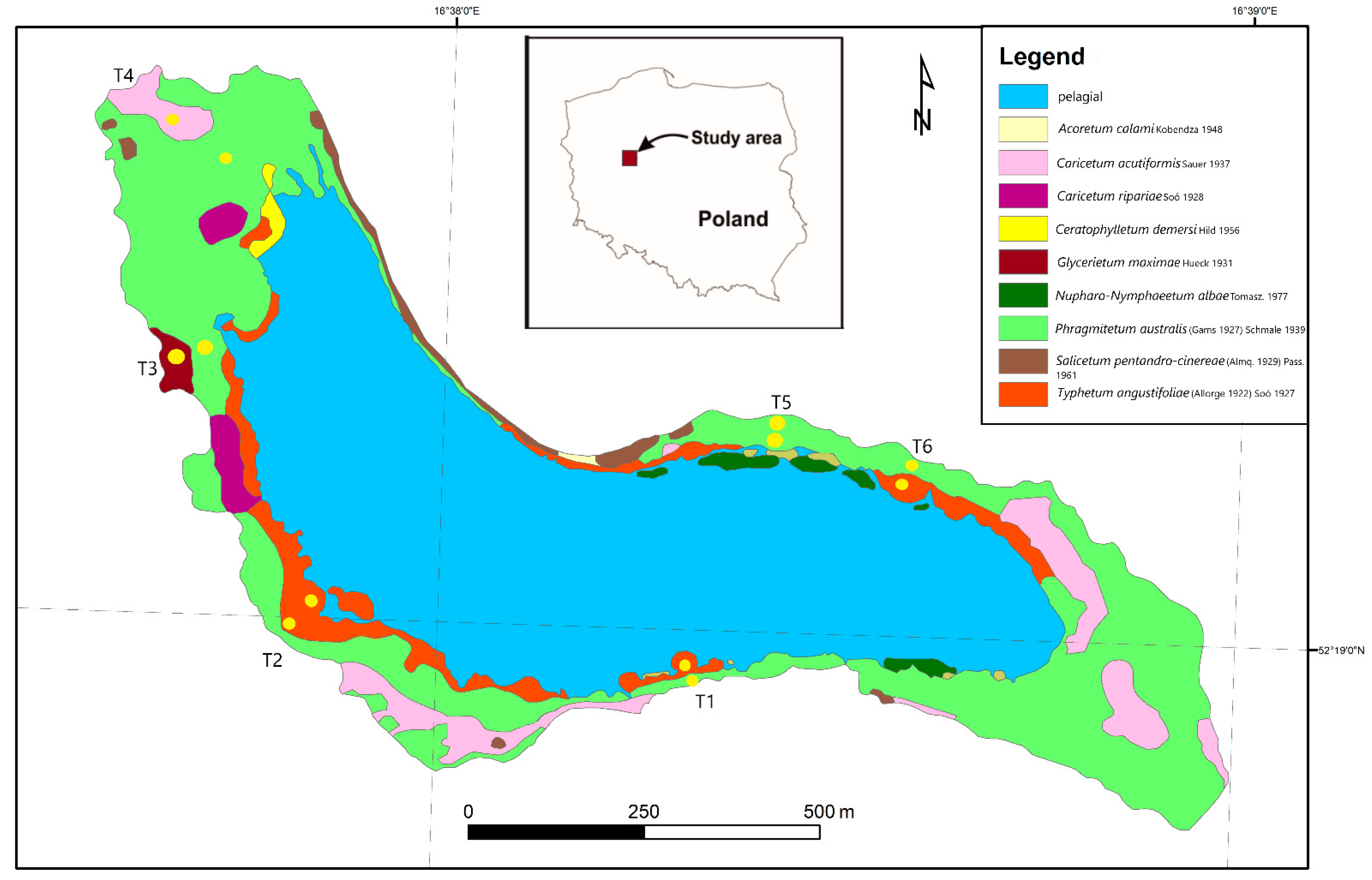
2.1. Site Description
2.2. Field and Laboratory Procedures
2.3. Statistical Analyses
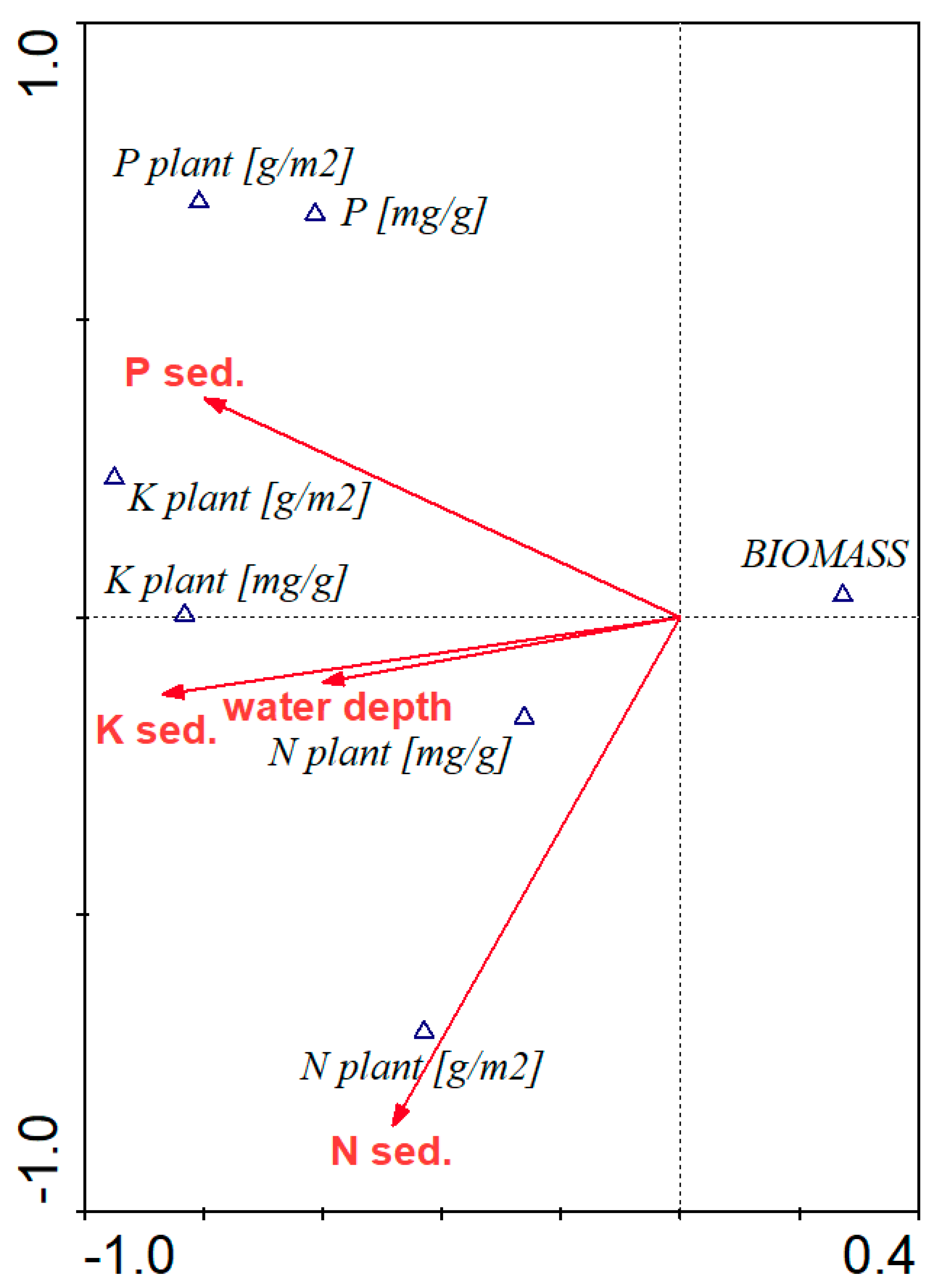
3. Results
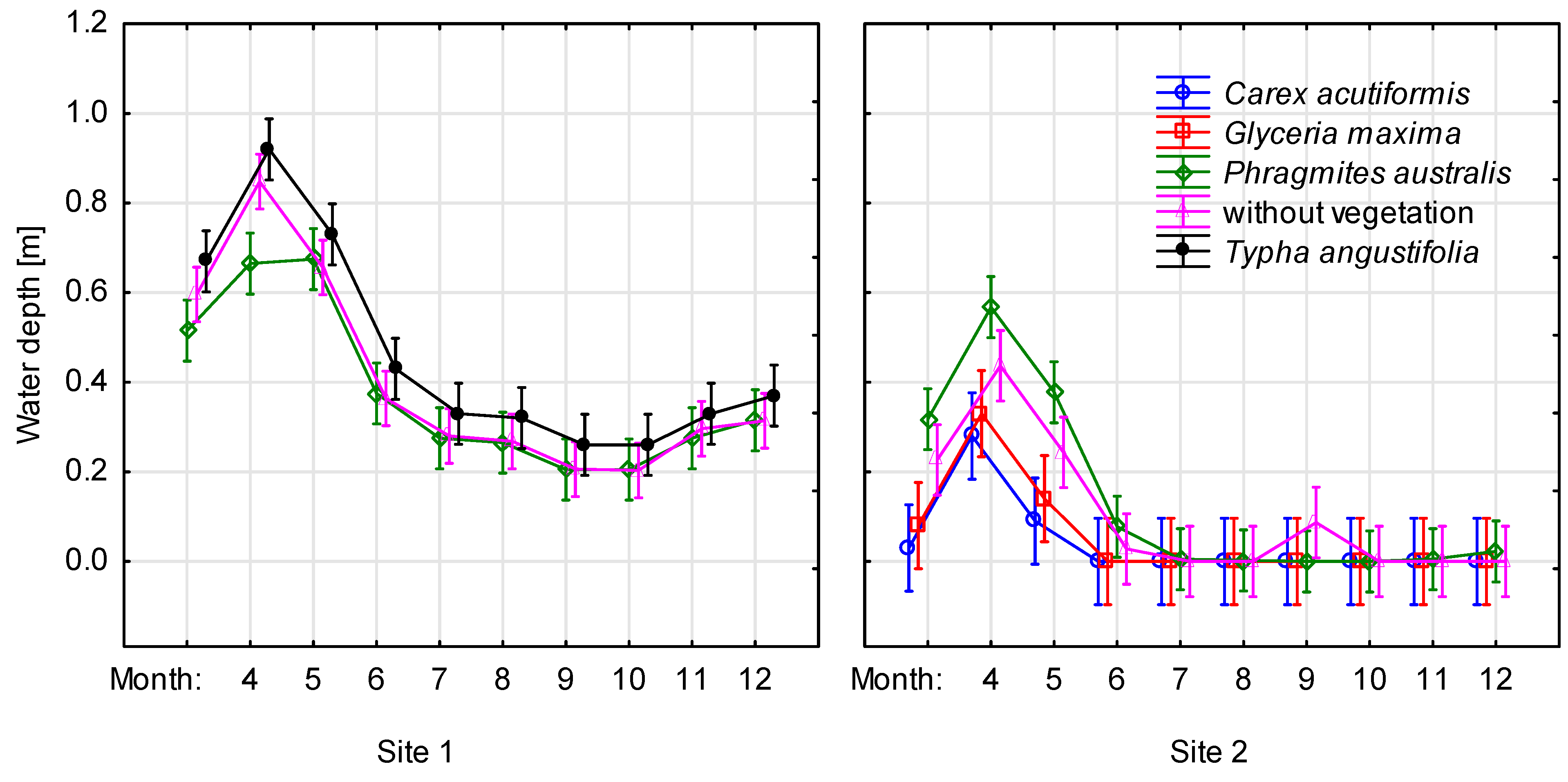
3.1. Changes of Water Level in the: Littoral Zone
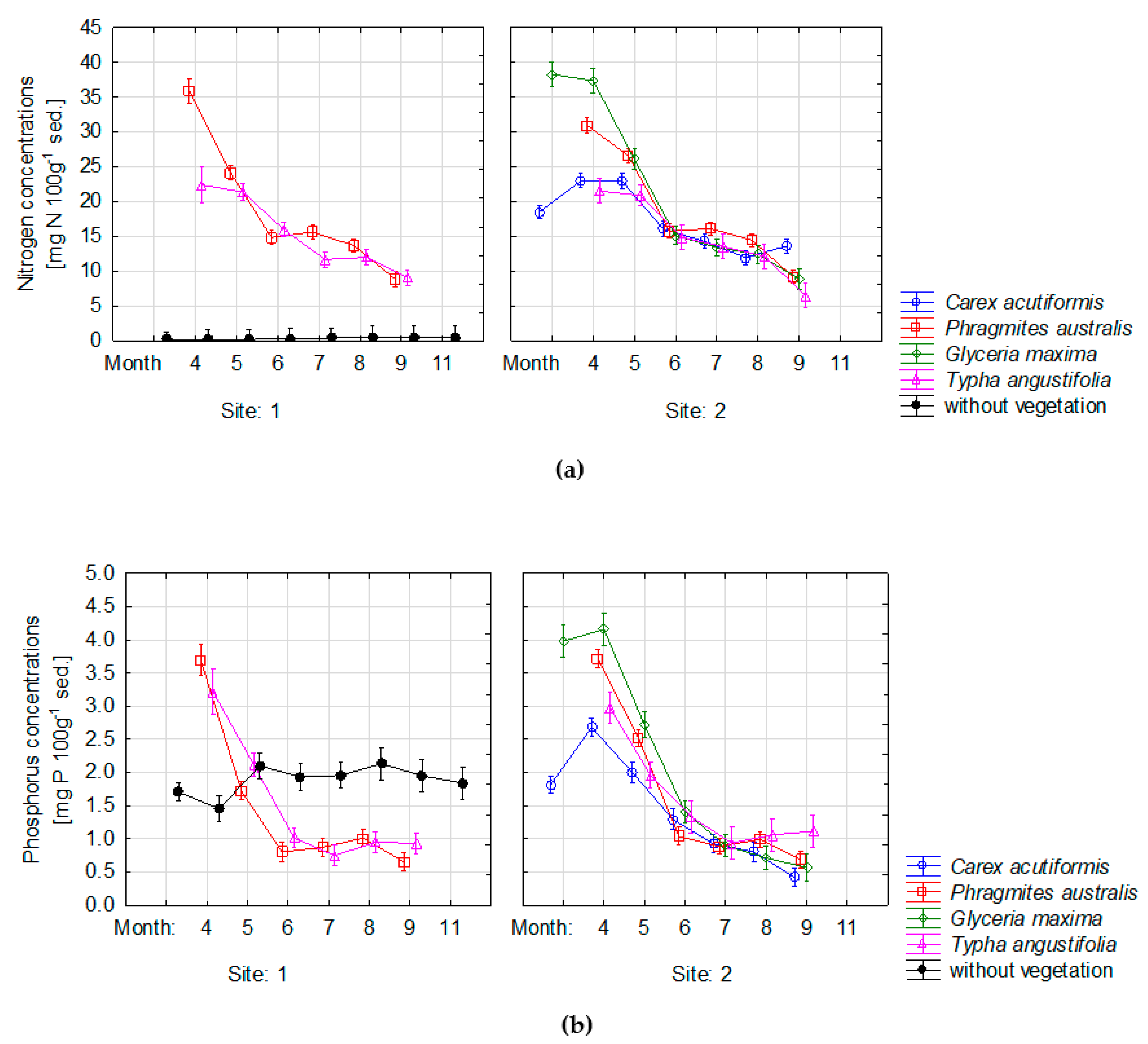
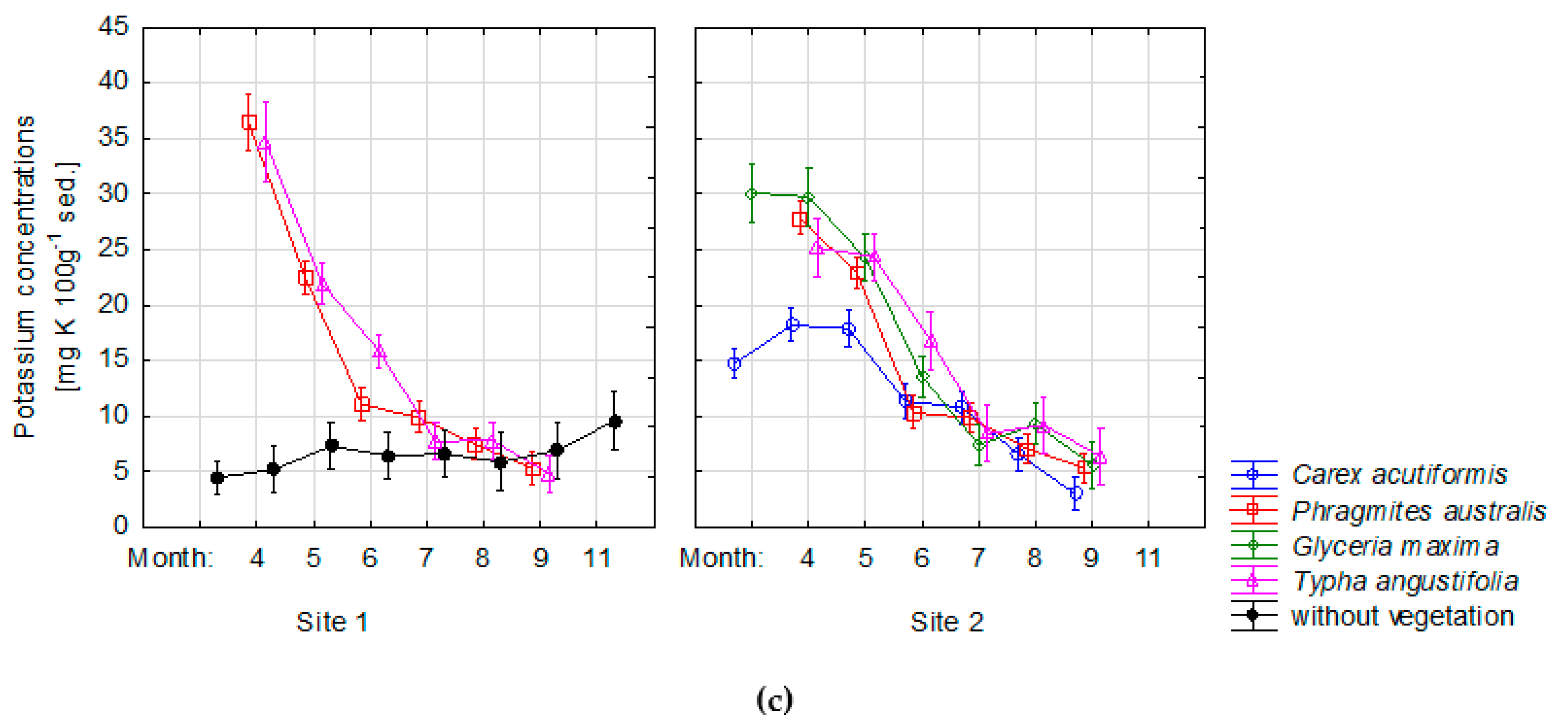
3.2. Changes of Nutrient Concentrations in Sediments
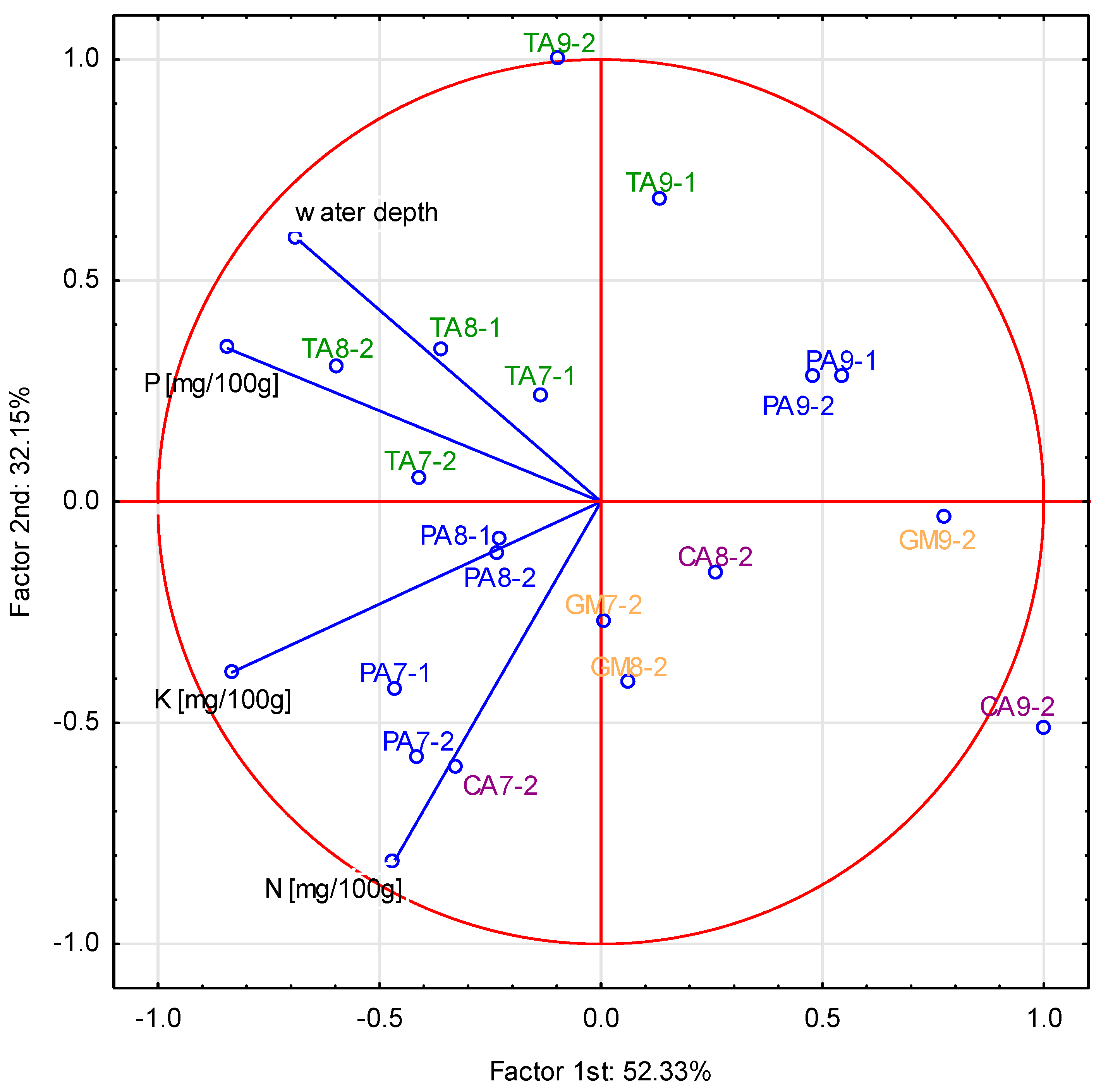
3.3. Variations of Nutrient Concentration in Sediment and Water Depth between Analyzed Sites
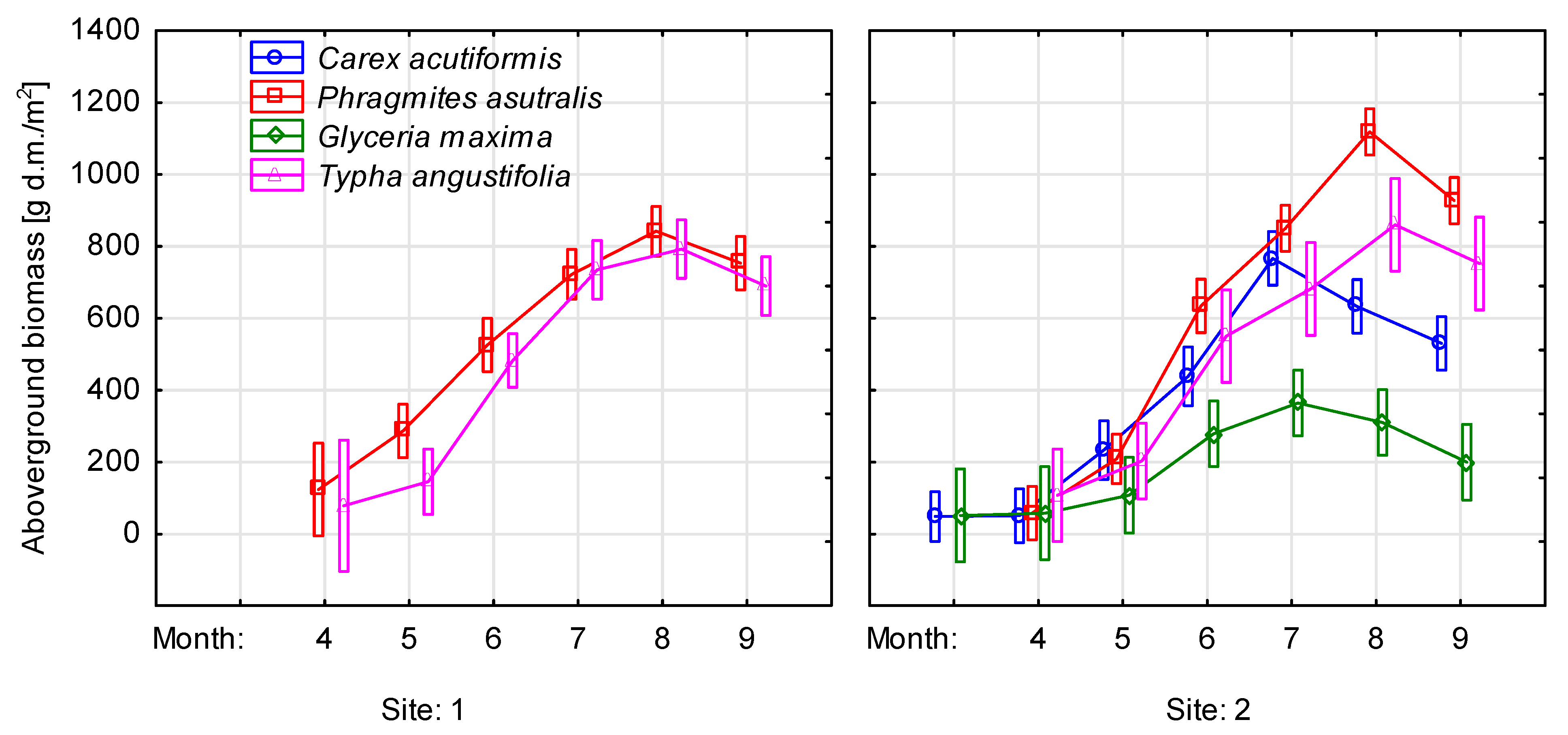
3.4. Plant Productivity
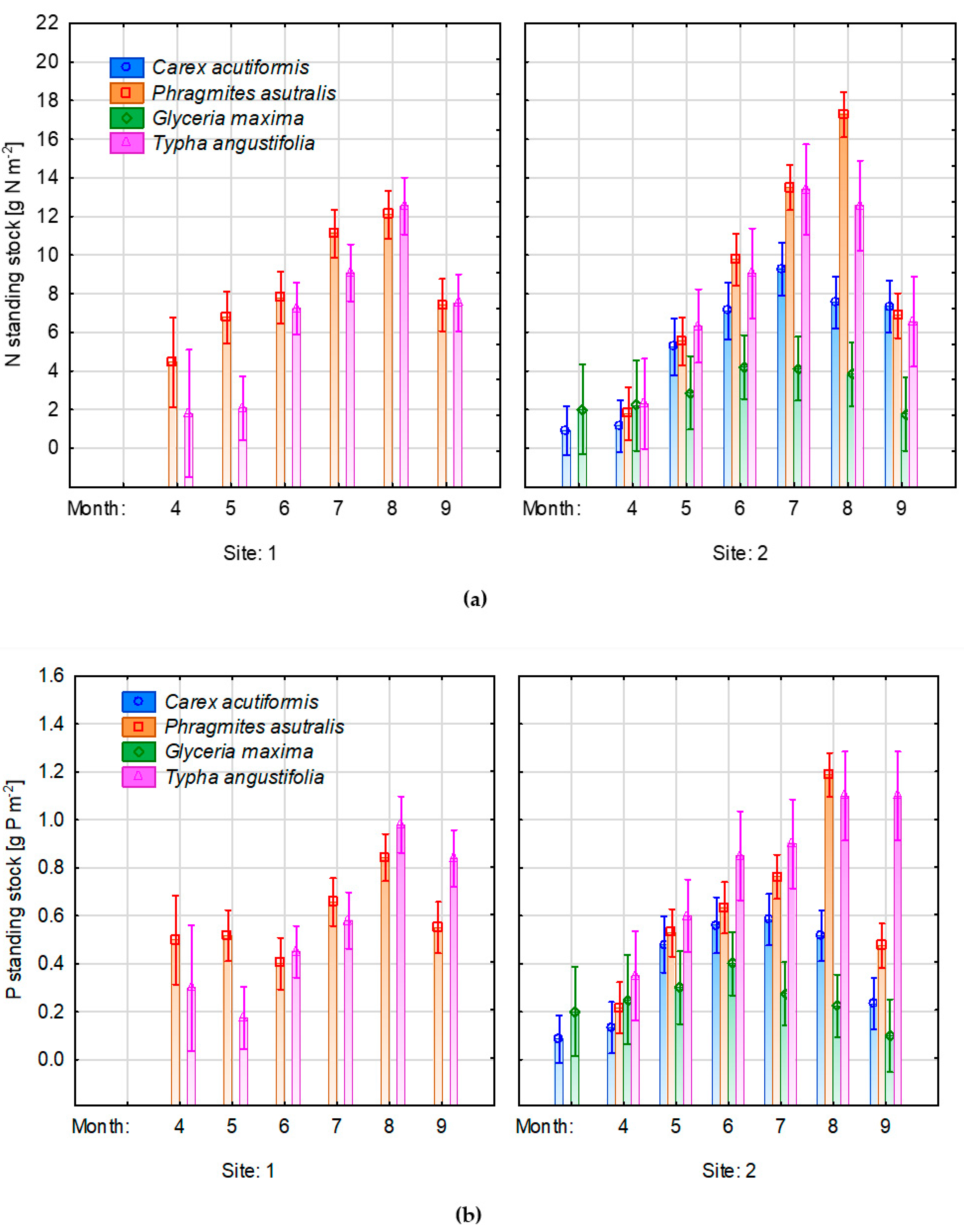
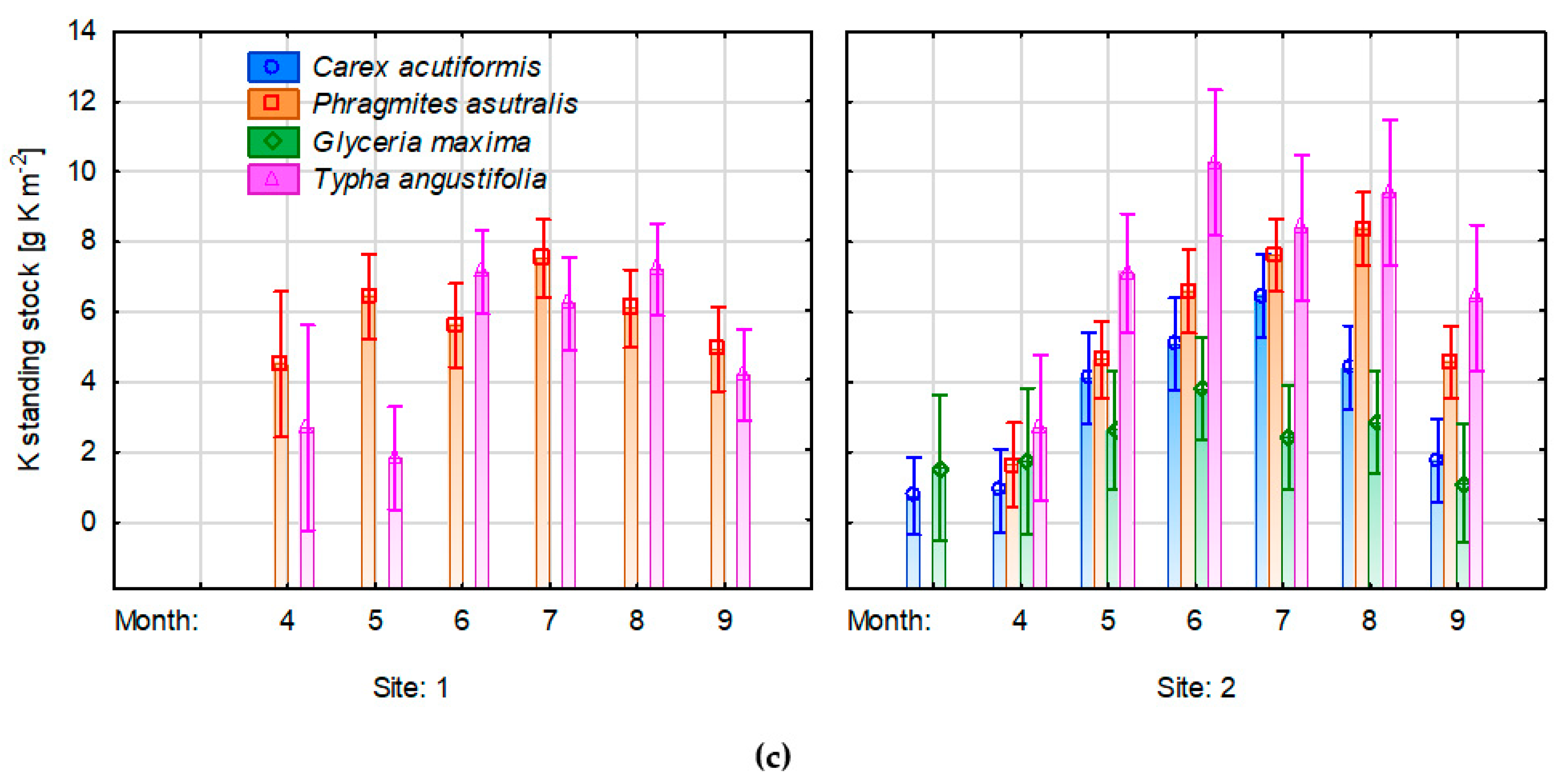
3.5. Changes of Nutrient Concentrations in Plant Biomass
3.6. Impact of Water Level Changes and Nutrient Availability on Plant Biomass
4. Discussion
4.1. Effect of Reduction of the Water Level on Nutrient Availability in Sediments in the Littoral Zone
4.2. Changes of Water Level on Nutrient Concentrations in the Aboveground Biomass of Emergent Species
4.3. Assessment of the Efficiency of Littoral Zone in Reduction of Nutrient Pollutions
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Site | Month | N [mg/g] | P [mg/g] | K [mg/g] | |||
|---|---|---|---|---|---|---|---|---|
| Mean | Stand. Dev. | Mean | Stand. Dev. | Mean | Stand. Dev. | |||
| Carex acutiformis L. | 2 | 3 | 0.91 | 0.35 | 0.09 | 0.05 | 0.74 | 0.33 |
| 2 | 4 | 1.16 | 0.10 | 0.14 | 0.03 | 0.91 | 0.19 | |
| 2 | 5 | 5.22 | 1.97 | 0.48 | 0.23 | 4.08 | 1.81 | |
| 2 | 6 | 7.11 | 2.86 | 0.58 | 0.25 | 5.08 | 2.01 | |
| 2 | 7 | 9.24 | 2.91 | 0.61 | 0.29 | 6.45 | 2.35 | |
| 2 | 8 | 7.54 | 1.51 | 0.51 | 0.19 | 4.38 | 2.78 | |
| 2 | 9 | 7.35 | 2.95 | 0.23 | 0.10 | 1.76 | 1.48 | |
| Phragmites australis (Cav.) Trin. ex Steud. | 1 | 4 | 4.47 | 1.03 | 0.46 | 0.14 | 4.52 | 0.27 |
| 1 | 5 | 6.75 | 3.87 | 0.51 | 0.32 | 6.44 | 4.01 | |
| 1 | 6 | 7.78 | 3.74 | 0.41 | 0.21 | 5.65 | 2.53 | |
| 1 | 7 | 11.14 | 4.91 | 0.60 | 0.36 | 7.52 | 4.97 | |
| 1 | 8 | 11.25 | 5.94 | 0.80 | 0.29 | 5.68 | 2.26 | |
| 1 | 9 | 7.43 | 4.60 | 0.56 | 0.38 | 4.93 | 3.42 | |
| 2 | 4 | 1.80 | 0.36 | 0.22 | 0.06 | 1.61 | 0.48 | |
| 2 | 5 | 5.54 | 1.85 | 0.52 | 0.17 | 4.65 | 1.86 | |
| 2 | 6 | 9.74 | 5.11 | 0.61 | 0.29 | 6.60 | 4.43 | |
| 2 | 7 | 13.50 | 3.44 | 0.66 | 0.34 | 7.63 | 2.97 | |
| 2 | 8 | 14.96 | 5.58 | 1.06 | 0.50 | 7.02 | 3.64 | |
| 2 | 9 | 6.86 | 5.11 | 0.48 | 0.40 | 4.57 | 5.39 | |
| Glyceria maxima (C. Hartm.) Holmb. | 2 | 3 | 1.98 | 0.28 | 0.21 | 0.05 | 1.55 | 0.09 |
| 2 | 4 | 2.19 | 0.89 | 0.25 | 0.12 | 1.73 | 0.62 | |
| 2 | 5 | 2.84 | 0.80 | 0.29 | 0.01 | 2.60 | 0.77 | |
| 2 | 6 | 4.22 | 1.15 | 0.40 | 0.07 | 3.79 | 1.22 | |
| 2 | 7 | 4.11 | 0.41 | 0.29 | 0.12 | 2.41 | 1.55 | |
| 2 | 8 | 3.82 | 0.76 | 0.22 | 0.07 | 2.86 | 1.35 | |
| 2 | 9 | 1.79 | 0.45 | 0.11 | 0.01 | 1.12 | 0.02 | |
| Typha angustifolia L. | 1 | 4 | 1.76 | 0.74 | 0.25 | 0.14 | 2.73 | 1.21 |
| 1 | 5 | 2.08 | 1.73 | 0.19 | 0.15 | 1.83 | 1.45 | |
| 1 | 6 | 8.93 | 4.52 | 0.57 | 0.30 | 8.74 | 4.24 | |
| 1 | 7 | 6.33 | 3.98 | 0.38 | 0.23 | 4.47 | 3.48 | |
| 1 | 8 | 11.77 | 4.74 | 0.89 | 0.25 | 6.75 | 3.04 | |
| 1 | 9 | 7.78 | 1.47 | 0.70 | 0.52 | 4.35 | 2.88 | |
| 2 | 4 | 2.31 | 0.38 | 0.33 | 0.11 | 2.70 | 0.40 | |
| 2 | 5 | 6.32 | 4.11 | 0.58 | 0.35 | 3.80 | 2.37 | |
| 2 | 6 | 4.17 | 0.55 | 0.34 | 0.11 | 4.75 | 0.72 | |
| 2 | 7 | 9.34 | 3.37 | 0.64 | 0.01 | 5.85 | 2.13 | |
| 2 | 8 | 10.54 | 0.23 | 0.91 | 0.18 | 8.17 | 2.19 | |
| 2 | 9 | 12.30 | 8.78 | 0.98 | 0.69 | 4.45 | 4.50 | |
References
- Zhang, G.; Wu, Y.; Li, H.; Zhao, W.; Wang, F.; Chen, J.; Sivakumar, B.; Liu, S.; Qiu, L.; Wang, W. Assessment of water retention variation and risk warning under climate change in an inner headwater basin in the 21st century. J. Hydrol. 2022, 615, 128717. [Google Scholar] [CrossRef]
- Jeppesen, E.; Meerhoff, M.; Davidson, T.A.; Trolle, D.; Søndergaard, M.; Lauridsen, T.L.; Beklioǧlu, M.; Brucet, S.; Volta, P.; González-Bergonzoni, I.; et al. Climate change impacts on lakes: An integrated ecological perspective based on a multi-faceted approach, with special focus on shallow lakes. J. Limnol. 2014, 73, 88–111. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2022: Impacts, Adaptation and Vulnerability; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022. [Google Scholar]
- Gizińska, J.; Sojka, M. How Climate Change Affects River and Lake Water Temperature in Central- How Climate Change Affects River and Lake Water Temperature in Central-West Poland—A Case Study of the Warta River Catchment. Atmosphere 2023, 14, 330. [Google Scholar] [CrossRef]
- Nowak, B.M.; Ptak, M. Natural and anthropogenic conditions of water level fluctuations in lakes-Lake Powidzkie case study (Central-Western Poland). J. Water Land Dev. 2019, 40, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Choiński, A.; Ptak, M.; Ławniczak, A.E. Changes in water resources of Polish lakes as influenced by natural and anthropogenic factors. Polish J. Environ. Stud. 2016, 25, 1883–1890. [Google Scholar] [CrossRef] [PubMed]
- Freeman, E.C.; Creed, I.F.; Jones, B.; Bergström, A.-K. Global changes may be promoting a rise in select cyanobacteria in nutrient-poor northern lakes. Glob. Chang. Biol. 2020, 26, 4966–4987. [Google Scholar] [CrossRef]
- Kakouei, K.; Kraemer, B.M.; Anneville, O.; Carvalho, L.; Feuchtmayr, H.; Graham, J.L.; Higgins, S.; Pomati, F.; Rudstam, L.G.; Stockwell, J.D.; et al. Phytoplankton and cyanobacteria abundances in mid-21st century lakes depend strongly on future land use and climate projections. Glob. Chang. Biol. 2021, 27, 6409–6422. [Google Scholar] [CrossRef]
- Bullerjahn, G.S.; McKay, R.M.L.; Bernát, G.; Prášil, O.; Vörös, L.; Pálffy, K.; Tugyi, N.; Somogyi, B. Community dynamics and function of algae and bacteria during winter in central European great lakes. J. Great Lakes Res. 2020, 46, 732–740. [Google Scholar] [CrossRef]
- Gámez, T.E.; Benton, L.; Manning, S.R. Observations of two reservoirs during a drought in central Texas, USA: Strategies for detecting harmful algal blooms. Ecol. Indic. 2019, 104, 588–593. [Google Scholar] [CrossRef]
- Dunalska, J.A.; Grochowska, J.; Wiśniewski, G.; Napiórkowska-Krzebietke, A. Can we restore badly degraded urban lakes? Ecol. Eng. 2015, 82, 432–441. [Google Scholar] [CrossRef]
- Baastrup-Spohr, L.; Sand-Jensen, K.; Olesen, S.C.H.; Bruun, H.H. Recovery of lake vegetation following reduced eutrophication and acidification. Freshw. Biol. 2017, 62, 1847–1857. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, H.; Du, C.; Zhang, W.; Shen, J.; Yang, S.; Yang, L. Spatiotemporal differences in phosphorus release potential of bloom-forming cyanobacteria in Lake Taihu. Environ. Pollut. 2021, 271, 116294. [Google Scholar] [CrossRef]
- A Rahman, A.R.A.; Sinang, S.C.; Nayan, N. Response of algal biomass and macrophyte communities to internal or external nutrient loading. Environ. Monit Assess. 2022, 194, 491. [Google Scholar] [CrossRef]
- Ławniczak, A. Variability of nutrient concentrations in sediments and wetland plants during the vegetation season under different sediment moisture conditions. Limnol. Rev. 2011, 11, 47–58. [Google Scholar] [CrossRef]
- Räty, M.; Uusi-Kämppä, J.; Yli-Halla, M.; Rasa, K.; Pietola, L. Phosphorus and nitrogen cycles in the vegetation of differently managed buffer zones. Nutr. Cycl. Agroecosyst. 2010, 86, 121–132. [Google Scholar] [CrossRef]
- Liira, J.; Feldmann, T.; Mäemets, H.; Peterson, U. Two decades of macrophyte expansion on the shores of a large shallow northern temperate lake-A retrospective series of satellite images. Aquat. Bot. 2010, 93, 207–215. [Google Scholar] [CrossRef]
- Bachmann, R.W.; Horsburgh, C.A.; Hoyer, M.V.; Mataraza, L.K.; Canfield, D.E. Relations between trophic state indicators and plant biomass in Florida lakes. Hydrobiologia 2002, 470, 219–234. [Google Scholar] [CrossRef]
- Ma, D.; Chen, S.; Lu, J.; Liao, H. Study of the effect of periphyton nutrient removal on eutrophic lake water quality. Ecol. Eng. 2019, 130, 122–130. [Google Scholar] [CrossRef]
- Seitz, C.; Scordo, F.; Vitale, A.J.; Science, C. The effects of extreme drought events on the morphometry of shallow lakes: Implications for sediment resuspension and littoral and pelagic zone distribution. J. S. Am. Earth Sci. 2020, 103, 102743. [Google Scholar] [CrossRef]
- Peršić, V.; Horvatić, J.; Has-Schön, E.; Bogut, I. Changes in N and P limitation induced by water level fluctuations in Nature Park Kopački Rit (Croatia): Nutrient enrichment bioassay. Aquat. Ecol. 2009, 43, 27–36. [Google Scholar] [CrossRef]
- Sfriso, A.; Buosi, A.; Juhmani, A.S.; Tomio, Y.; Mistri, M.; Munari, C.; Sfriso, A.A. Sedimentation Rates: Anthropogenic Impacts and Environmental Changes in Transitional Water Systems. Water 2022, 14, 3843. [Google Scholar] [CrossRef]
- Palmer, M.A.; Covich, A.P.; Lake, S.; Biro, P.; Brooks, J.J.; Cole, J.; Dahm, C.; Gibert, J.; Goedkoop, W.; Martens, K.; et al. Linkages between aquatic sediment biota and life above sediments as potential drivers of biodiversity and ecological processes. Bioscience 2000, 50, 1062–1075. [Google Scholar] [CrossRef]
- Chang, J.; Liu, D.; Cao, H.; Chang, S.X.; Wang, X.; Huang, C.; Ge, Y. NO3-/NH4+ratios affect the growth and N removal ability of Acorus calamus and Iris pseudacorus in a hydroponic system. Aquat. Bot. 2010, 93, 216–220. [Google Scholar] [CrossRef]
- Small, G.E.; Finlay, J.C.; McKay, R.M.L.; Rozmarynowycz, M.J.; Brovold, S.; Bullerjahn, G.S.; Spokas, K.; Sterner, R.W. Large differences in potential denitrification and sediment microbial communities across the Laurentian great lakes. Biogeochemistry 2016, 128, 353–368. [Google Scholar] [CrossRef]
- Ławniczak, A.E. The role of emergent macrophytes in nutrient cycling in Lake Niepruszewskie (western Poland). Oceanol. Hydrobiol. Stud. 2010, 39, 75–83. [Google Scholar] [CrossRef]
- Tylová, E.; Steinbachová, L.; Votrubová, O.; Lorenzen, B.; Brix, H. Different sensitivity of Phragmites australis and Glyceria maxima to high availability of ammonium-N. Aquat. Bot. 2008, 88, 93–98. [Google Scholar] [CrossRef]
- Alahuhta, J.; Kosten, S.; Akasaka, M.; Auderset, D.; Azzella, M.M.; Bolpagni, R.; Bove, C.P.; Chambers, P.A.; Chappuis, E.; Clayton, J.; et al. Global variation in the beta diversity of lake macrophytes is driven by environmental heterogeneity rather than latitude. J. Biogeogr. 2017, 44, 1758–1769. [Google Scholar] [CrossRef] [Green Version]
- Lawniczak-Malińska, A.; Ptak, M.; Celewicz, S.; Choiński, A. Impact of lake morphology and shallowing on the rate of overgrowth in hard-water eutrophic lakes. Water 2018, 10, 1827. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Guo, X.; Yu, X.; Liu, L.; Wang, N.; Eller, F.; Guo, W. Is there evidence of local adaptation of Phragmites australis to water level gradients and fluctuation frequencies? Sci. Total Environ. 2021, 756, 144065. [Google Scholar] [CrossRef]
- Kolada, A. The use of helophytes in assessing eutrophication of temperate lowland lakes: Added value? Aquat. Bot. 2016, 129, 44–54. [Google Scholar] [CrossRef]
- Lawniczak, A.E.; Zbierska, J.; Choiński, A.; Szczepaniak, W. Response of emergent macrophytes to hydrological changes in a shallow lake, with special reference to nutrient cycling. Hydrobiologia 2010, 656, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Lawniczak, A.E. Nitrogen, Phosphorus, and potassium resorption efficiency and proficiency of four emergent macrophytes from Nutrient-Rich Wetlands. Polish J. Environ. Stud. 2011, 20, 1227–1234. [Google Scholar]
- Alahuhta, J.; Kanninen, A.; Hellsten, S.; Vuori, K.M.; Kuoppala, M.; Hämäläinen, H. Variable response of functional macrophyte groups to lake characteristics, land use, and space: Implications for bioassessment. Hydrobiologia 2014, 737, 201–214. [Google Scholar] [CrossRef]
- Lawniczak, A.E.; Pełechata, A.; Czerniawska-Kusza, I.; Golski, J. Wody powierzchniowe Wielkopolskiego Parku Narodowego i jego Otuliny. Tom I. JEZIORA. (Freshwaters of Wielkopolska National Park and its Buffer zone Vol. I Lakes); Bogucki Wyd. Naukowe: Poznań, Poland, 2016; Volume I, ISBN 9789896540821. [Google Scholar]
- Ławniczak, A.E. Long-term changes in water quality and ecological status of lakes in the Wielkopolska National Park and its buffer zone with special reference to effectiveness of the lakes’ protection. Oceanol. Hydrobiol. Stud. 2016, 45, 524–538. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Valjarević, A.; Milanović, M.; Gultepe, I.; Filipović, D.; Lukić, T. Updated Trewartha climate classification with four climate change scenarios. Geogr. J. 2022, 188, 506–517. [Google Scholar] [CrossRef]
- Kundzewicz, Z.W.; Matczak, P. Climate change regional review: Poland. WIREs Clim. Chang. 2012, 3, 297–311. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen—Total. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1983; pp. 595–624. ISBN 9780891189770. [Google Scholar]
- Tanner, C.C. Plants for constructed wetland treatment systems—A comparison of the growth and nutrient uptake of eight emergent species. Ecol. Eng. 1996, 7, 59–83. [Google Scholar] [CrossRef]
- Brändle, R.; Pokorný, J.; Květ, J.; Čížková, H. Wetland plants as a subject of interdisciplinary research. Folia Geobot. 1996, 31, 1–6. [Google Scholar] [CrossRef]
- Lawniczak, A. Response of two wetland graminoids to N:K supply ratios in a two-year growth experiment. J. Elem. 2011, 16, 421–436. [Google Scholar] [CrossRef]
- Güsewell, S.; Koerselman, W. Variation in nitrogen and phosphorus concentrations of wetland plants. Perspect. Plant Ecol. Evol. Syst. 2002, 5, 37–61. [Google Scholar] [CrossRef]
- Verhoeven, J.T.A.; Arts, H.H.M. Nutrient dynamics in small mesotrophic fens surrounded by cultivated land: II. N and P accumulation in plant biomass in relation to the release of inorganic N and P in the peat soil. Oecologia 1987, 72, 557–561. [Google Scholar] [CrossRef]
- Shaver, G.R.; Chapin, F.S. Long–term responses to factorial, NPK fertilizer treatment by Alaskan wet and moist tundra sedge species. Ecography 1995, 18, 259–275. [Google Scholar] [CrossRef]
- Lawniczak, A.E.; Drapikowska, M.; Celka, Z.; Szkudlarz, P.; Jackowiak, B. Response of Anthoxanthum Odoratum and a. Aristatum To Different Habitat Types and Nutrient Concentrations in Soil. Fresenius Environ. Bull. 2011, 20, 2465–2474. [Google Scholar]
- Sánchez Pérez, J.M.; Antiguedad, I.; Arrate, I.; García-Linares, C.; Morell, I. The influence of nitrate leaching through unsaturated soil on groundwater pollution in an agricultural area of the Basque country: A case study. Sci. Total Environ. 2003, 317, 173–187. [Google Scholar] [CrossRef] [Green Version]
- Wei, A.; Chow-Fraser, P. Synergistic impact of water level fluctuation and invasion of Glyceria on Typha in a freshwater marsh of Lake Ontario. Aquat. Bot. 2006, 84, 63–69. [Google Scholar] [CrossRef]
- Weiher, E.; Wisheu, I.C.; Keddy, P.A.; Moore, D.R.J. Establishment, persistence, and management implications of experimental wetland plant communities. Wetlands 1996, 16, 208–218. [Google Scholar] [CrossRef]
- Steinbachová-Vojtıšková, L.; Tylová, E.; Soukup, A.; Novická, H.; Votrubová, O.; Lipavská, H.; Čıžková, H. Influence of nutrient supply on growth, carbohydrate, and nitrogen metabolic relations in Typha angustifolia. Environ. Exp. Bot. 2006, 57, 246–257. [Google Scholar] [CrossRef]
- Meeker, J.E.; Wilcox, D.A.; Johnson, S.E.; Tillison, N. Tracking vegetation transitions due to invasion of cattail (Typha) in lake superior coastal peatlands. Wetlands 2023, 43, 18. [Google Scholar] [CrossRef]
- Munzarova, E.; Lorenzen, B.; Brix, H.; Vojtiskova, L.; Votrubova, O. Effect of NH4+/NO3- availability on nitrate reductase activity and nitrogen accumulation in wetland helophytes Phragmites australis and Glyceria maxima. Environ. Exp. Bot. 2006, 55, 49–60. [Google Scholar] [CrossRef]
- Ciecierska, H. Macrophyte based indices of the ecological state of lakes. In Dissertations and Monographs, 139; University of Warmia and Mazury in Olsztyn: Olsztyn, Poland, 2008. [Google Scholar]
- Available online: http://www.iucngisd.org/gisd/species.php?sc=891 (accessed on 20 November 2022).
- Lawniczak-Malińska, A.E.; Achtenberg, K. Indicator values of emergent vegetation in overgrowing lakes in relation towater and sediment chemistry. Water 2018, 10, 498. [Google Scholar] [CrossRef] [Green Version]
- Saltonstall, K. Cryptic invasion by a non-native genotype of the common reed, Phragmites australis, into North America. Proc. Natl. Acad. Sci. USA 2002, 99, 2445–2449. [Google Scholar] [CrossRef] [Green Version]
- Kirk, H.; Paul, J.; Straka, J.; Freeland, J.R. Long-distance dispersal and high genetic diversity are implicated in the invasive spread of the common reed, Phragmites australis (Poaceae), in northeastern North America. Am. J. Bot. 2011, 98, 1180–1190. [Google Scholar] [CrossRef]
- Eller, F.; Skálová, H.; Caplan, J.S.; Bhattarai, G.P.; Burger, M.K.; Cronin, J.T.; Guo, W.-Y.; Guo, X.; Hazelton, E.L.G.; Kettenring, K.M.; et al. Cosmopolitan Species As Models for Ecophysiological Responses to Global Change: The Common Reed Phragmites australis. Front. Plant Sci. 2017, 8, 1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banaszuk, P.; Kamocki, A.K.; Wysocka-Czubaszek, A.; Czubaszek, R.; Roj-Rojewski, S. Closing the loop-Recovery of nutrients and energy from wetland biomass. Ecol. Eng. 2020, 143, 105643. [Google Scholar] [CrossRef]
- Gonzalez Mateu, M.; Yarwood, S.A.; Baldwin, A.H. Positive interactions occur between Phragmites australis lineages across short term experimental nutrient regimes. Aquat. Bot. 2021, 172, 103382. [Google Scholar] [CrossRef]
- Portielje, R.; Van der Molen, D.T. Relationships between eutrophication variables: From nutrient loading to transparency. Hydrobiologia 1999, 409, 375–387. [Google Scholar] [CrossRef]
- Vukov, D.; Ili, M.; Mirjana, Ć.; Igi, R. Environmental Drivers of Functional Structure and Diversity of Vascular Macrophyte Assemblages in Altered Waterbodies in Serbia. Diversity 2023, 15, 231. [Google Scholar] [CrossRef]
- Ławniczak, A. Overgrowing of two polymictic lakes in Central-Western Poland. Limnol. Rev. 2010, 10, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Ławniczak, A.E.; Choiński, A.; Kurzyca, I. Dynamics of lake morphometry and bathymetry in various hydrological conditions. Polish J. Environ. Stud. 2011, 20, 931–940. [Google Scholar]
- Zhao, J.; Liu, C.; Li, H.; Liu, J.; Jiang, T.; Yan, D.; Tong, J.; Dong, L. Review on Ecological Response of Aquatic Plants to Balanced Harvesting. Sustainability 2022, 14, 2451. [Google Scholar] [CrossRef]









| Source | N | P | K | Biomass | |||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | Sig. | F | Sig. | F | Sig. | F | Sig. | |
| Month | 6 | 31.103 | *** | 13.449 | *** | 16.621 | *** | 41.429 | *** |
| Species | 3 | 10.444 | *** | 10.345 | *** | 10.901 | *** | 4.535 | * |
| Site | 1 | 9.909 | ** | 6.214 | * | 14.821 | ** | 4.535 | * |
| Site * Species | 5 | 6.317 | *** | 6.225 | *** | 6.875 | *** | 7.480 | *** |
| Month * Site | 6.317 | *** | 6.225 | *** | 6.875 | *** | 20.175 | *** | |
| Month * Species | 12 | 16.550 | *** | 8.014 | *** | 10.246 | *** | 7.480 | *** |
| Month * Site * Species | 37 | 9.641 | *** | 4.668 | *** | 4.689 | *** | 14.241 | *** |
| Axes | 1 | 2 | 3 | 4 | Total Variance |
|---|---|---|---|---|---|
| Eigenvalues: | 0.447 | 0.005 | 0.002 | 0.000 | 1.000 |
| Water quality–environment correlations: | 0.676 | 0.628 | 0.816 | 0.286 | |
| Cumulative percentage variance | |||||
| of water quality data: | 44.7 | 45.2 | 45.4 | 45.5 | |
| of water quality–environment relation: | 98.3 | 99.4 | 99.9 | 100.0 | |
| Sum of all eigenvalues | 1 | ||||
| Sum of all canonical eigenvalues | 0.455 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ławniczak-Malińska, A. Effect of Water Level Reduction on the Littoral Zone in Terms of Its Efficiency in Lake Protection. Sustainability 2023, 15, 5563. https://doi.org/10.3390/su15065563
Ławniczak-Malińska A. Effect of Water Level Reduction on the Littoral Zone in Terms of Its Efficiency in Lake Protection. Sustainability. 2023; 15(6):5563. https://doi.org/10.3390/su15065563
Chicago/Turabian StyleŁawniczak-Malińska, Agnieszka. 2023. "Effect of Water Level Reduction on the Littoral Zone in Terms of Its Efficiency in Lake Protection" Sustainability 15, no. 6: 5563. https://doi.org/10.3390/su15065563