
Biogeographic Variability in Kernel Oil and Press Cake Content of Beauty Leaf Tree (Calophyllum inophyllum L.), as Determined by Chemical and Near-Infrared Spectroscopy Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
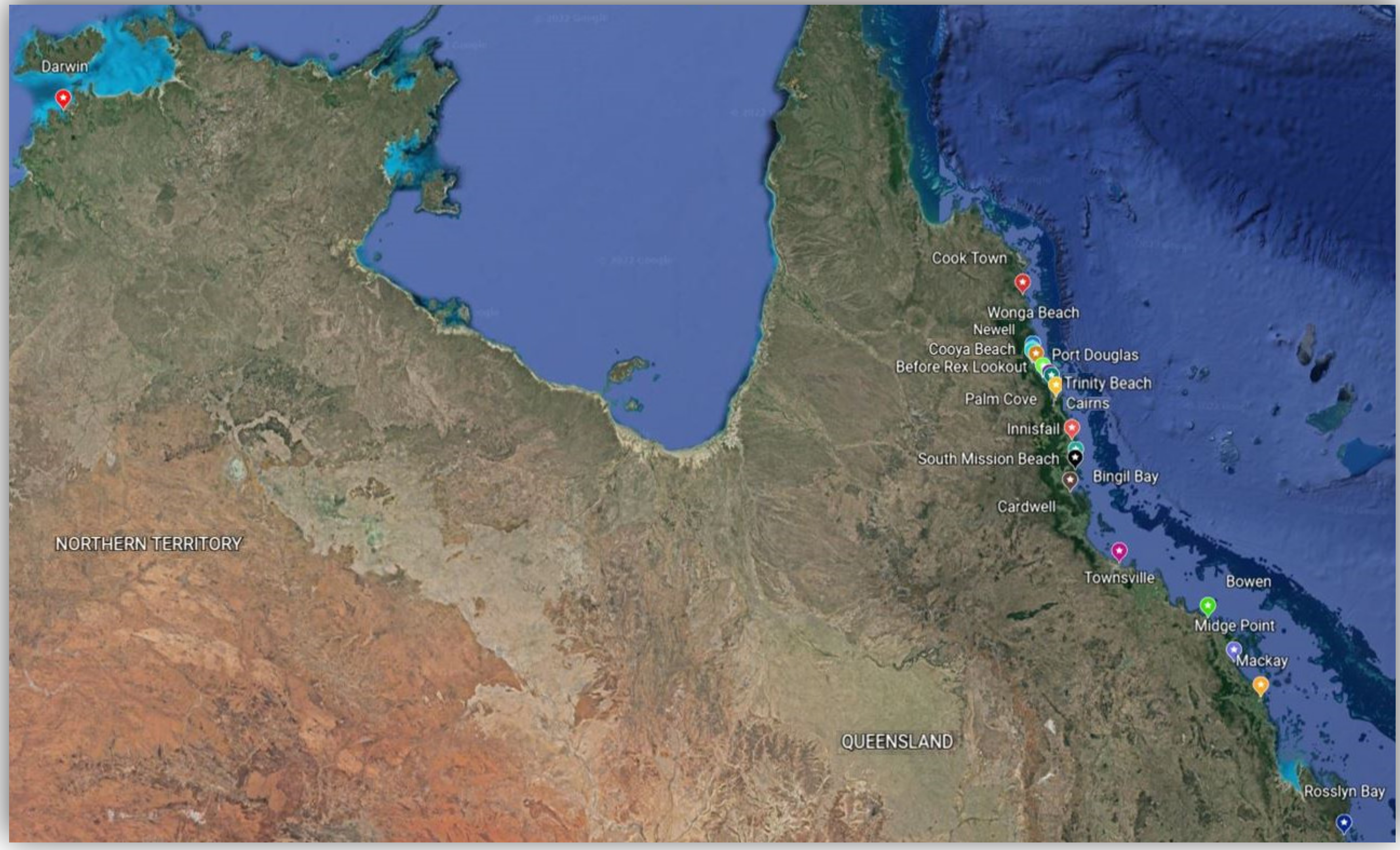
2.1. Study Sites and Seed Collection
2.2. Sample Preparation
2.3. Reference Method for Oil Extraction
2.4. FT–NIR Spectral Acquisition
2.5. Spectral Data Processing and Chemometric Analysis
3. Results and Discussion
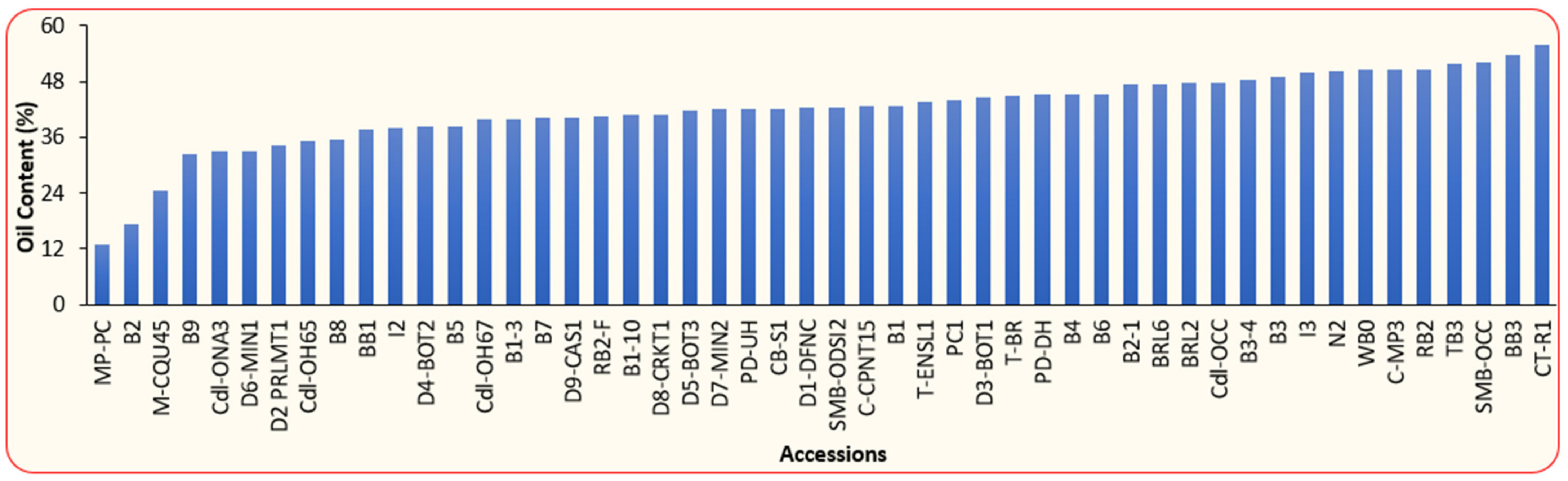
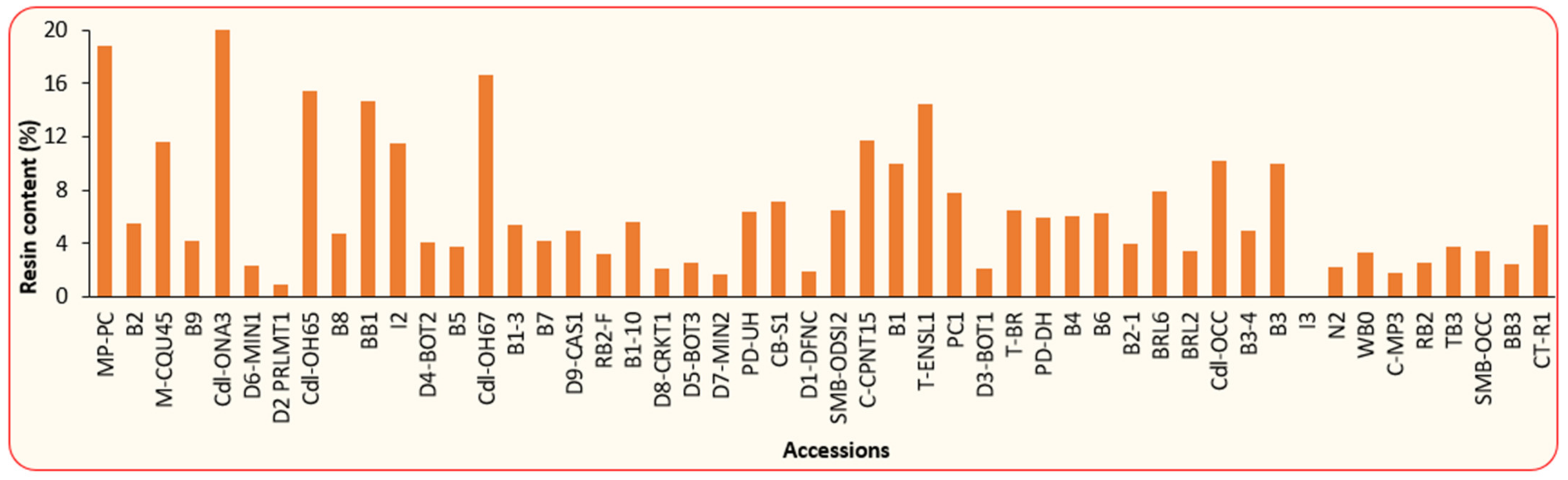
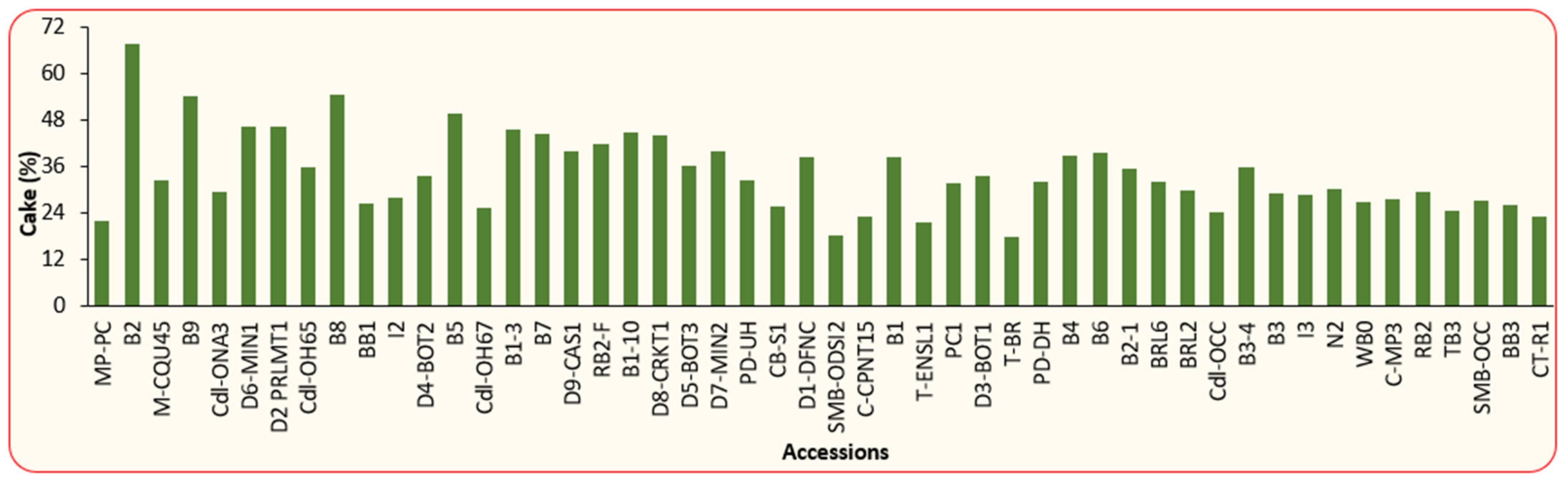
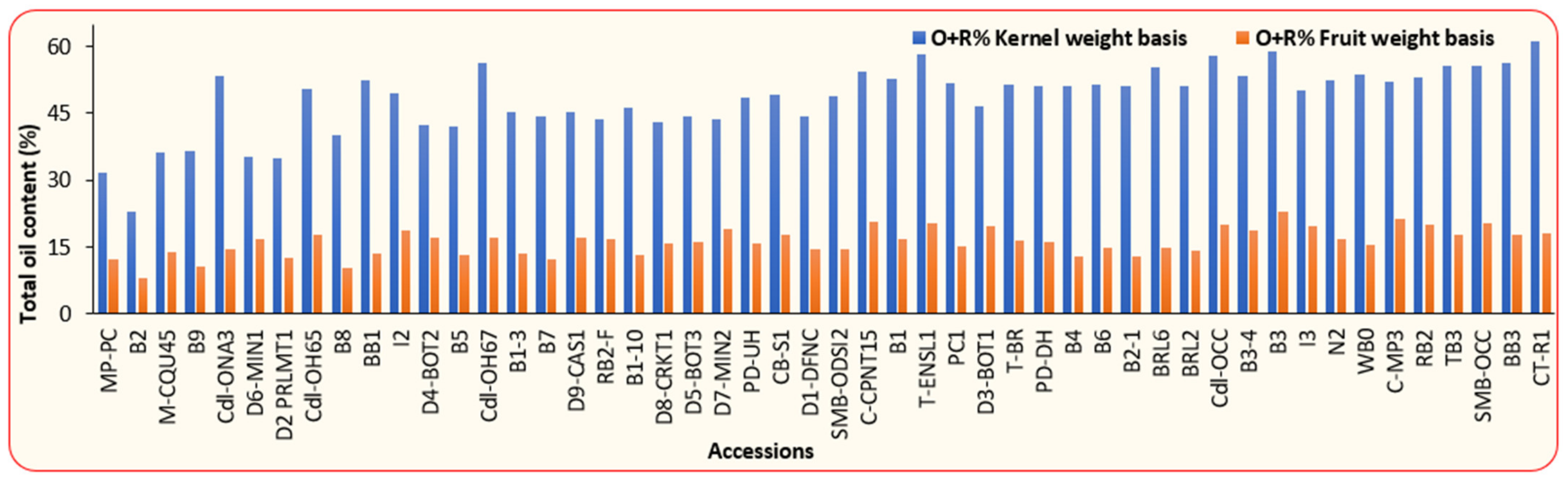
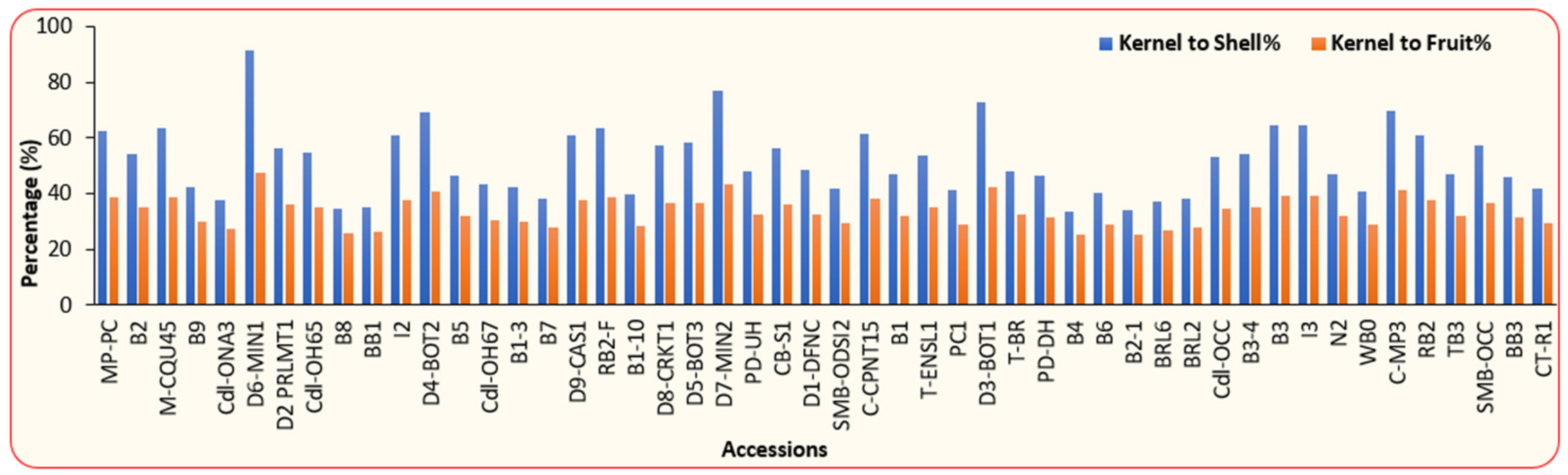
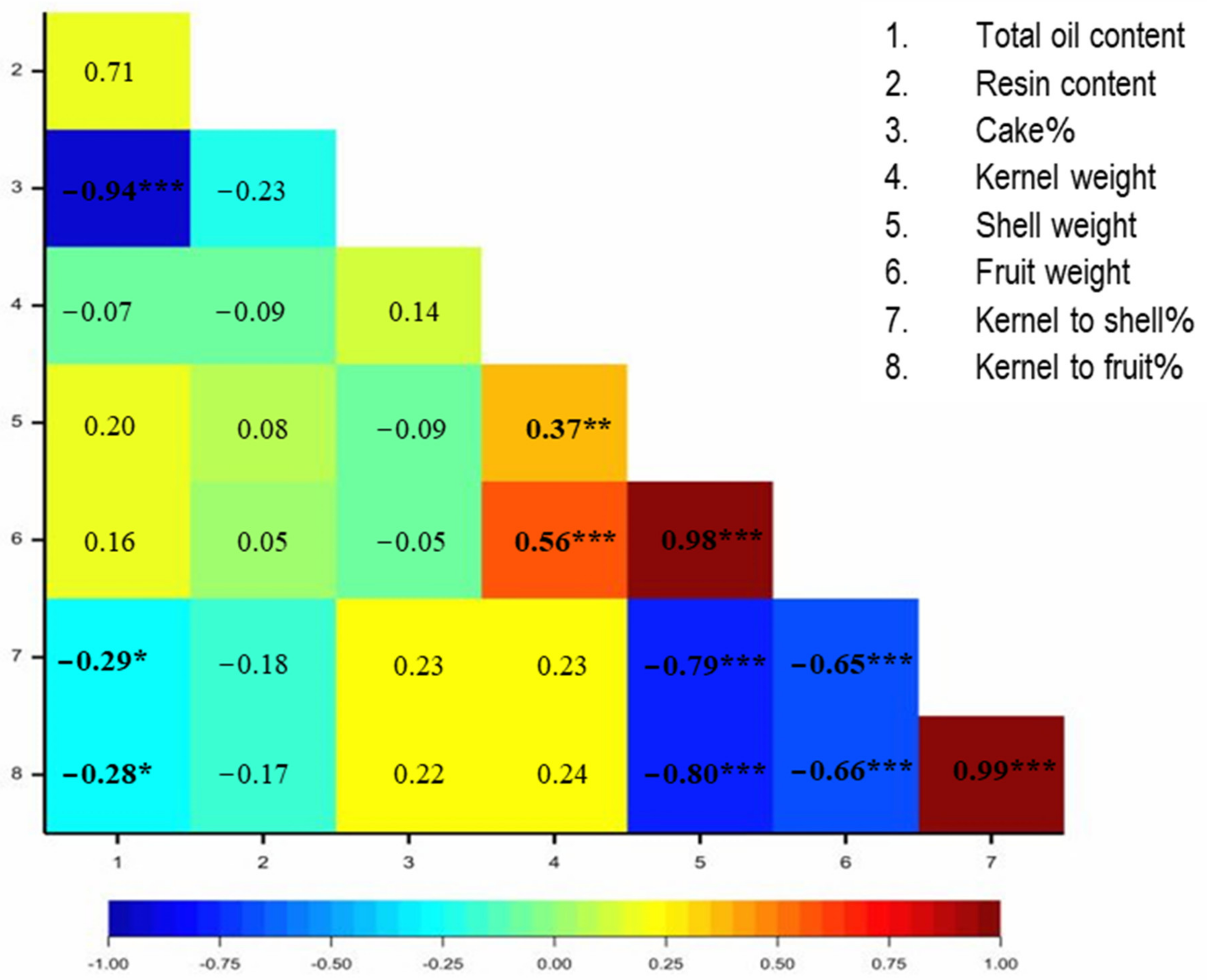
3.1. Physical and Chemical Properties of BLT Kernel Oil
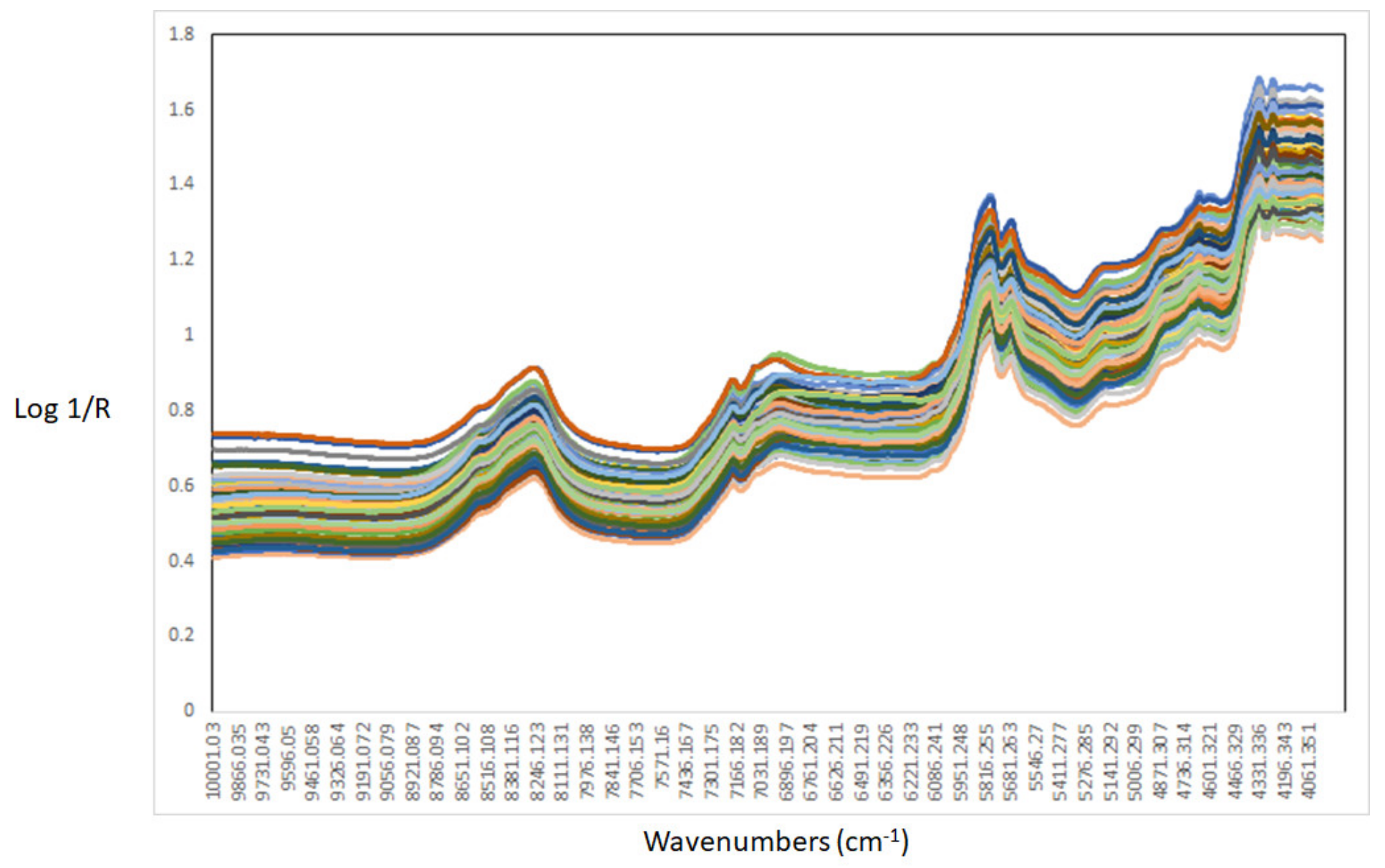
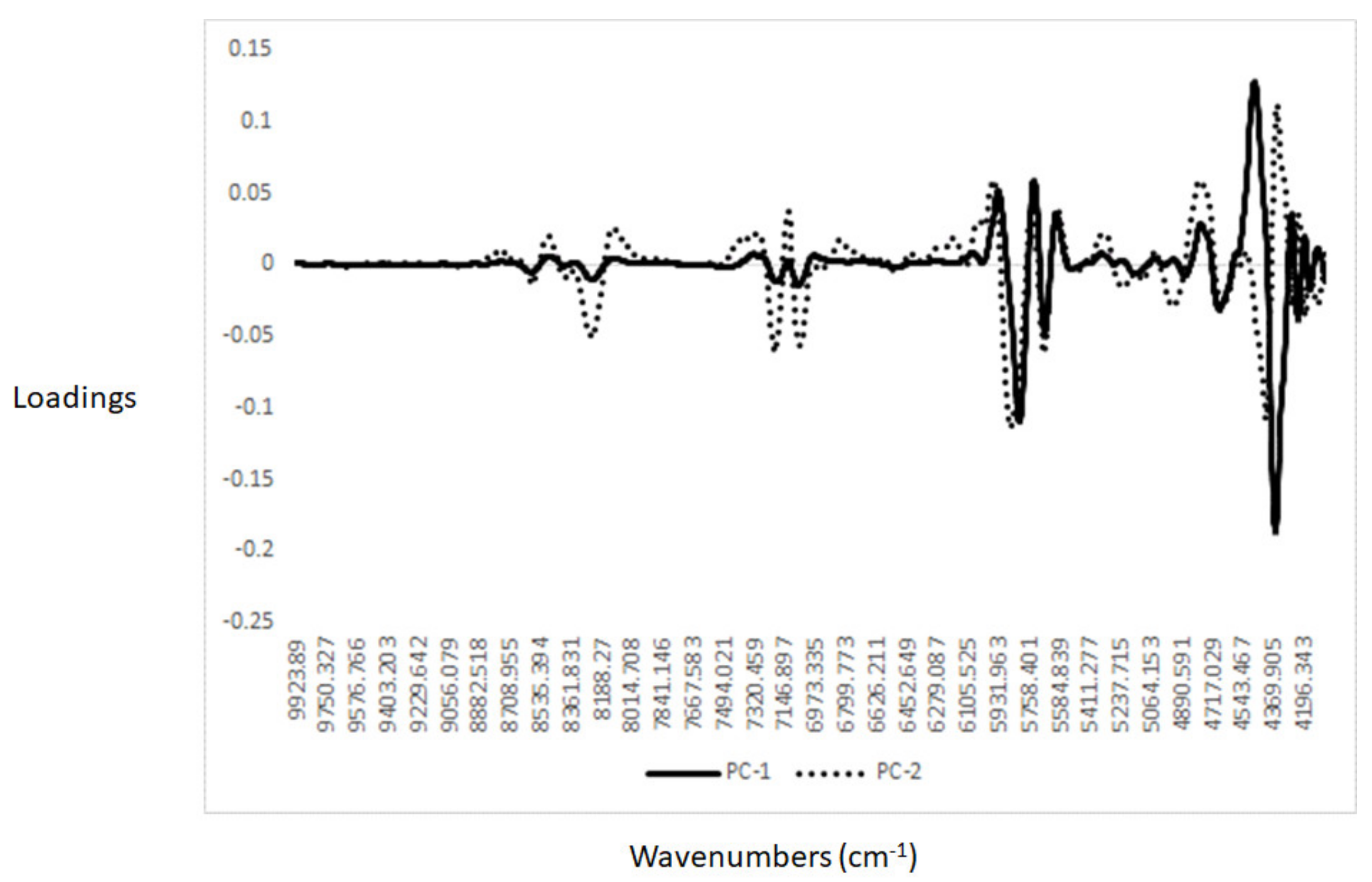
3.2. Near-Infrared Spectra Interpretation
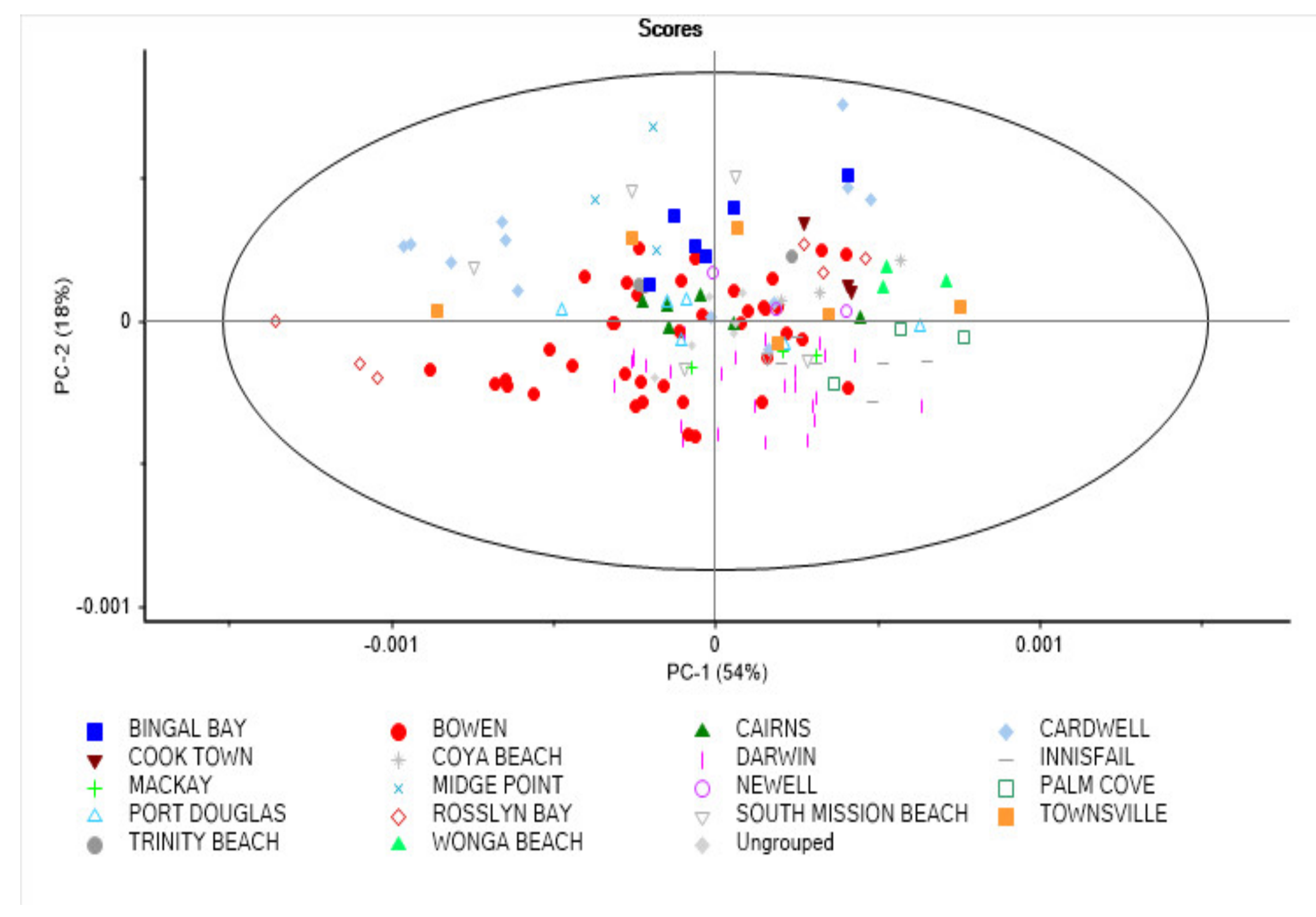
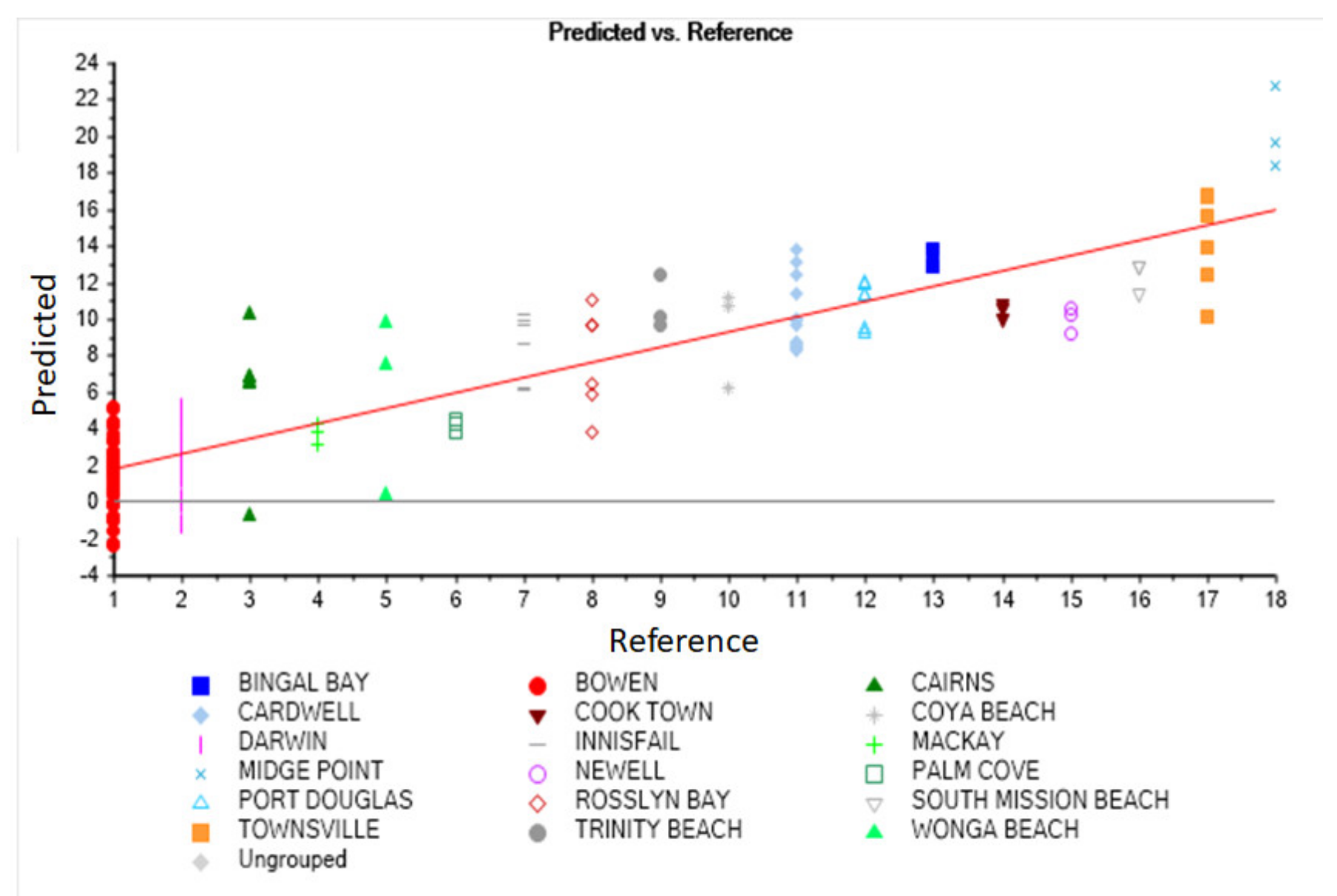
3.3. Principal Component and Discriminant Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Department of Industry, Science, Energy and Resources. Energy Consumption. 2020. Available online: https://www.energy.gov.au/data/energy-consumption (accessed on 10 May 2021).
- Anwar, M.; Rasul, M.G.; Ashwath, N. The synergistic effects of oxygenated additives on papaya biodiesel binary and ternary blends. Fuel 2019, 256, 115980. [Google Scholar] [CrossRef]
- Ginwal, H.; Rawat, P.S.; Srivastava, R.L. Seed source variation in growth performance and oil yield of Jatropha curcas Linn. in Central India. Silvae Genet. 2004, 53, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Ayanoglu, A.; Aksoy, B. Biochar production from various biomass by different technologies: A review. Energy Educ. Sci. Technol. Part C 2015, 7, 29–48. [Google Scholar]
- Bhuiya, M.M.K.; Rasul, M.; Khan, M.; Ashwath, N.; Mofijur, M. Comparison of oil extraction between screw press and solvent (n-hexane) extraction technique from beauty leaf (Calophyllum inophyllum L.) feedstock. Ind. Crops Prod. 2020, 144, 112024. [Google Scholar] [CrossRef]
- Friday, J.; Okano, D. Calophyllum inophyllum (kamani) Species profiles for Pacific Island Agroforestry, Traditional Tree Initiative, Hawaii. 2006. Available online: http//traditionaltree.org (accessed on 1 August 2018).
- Hathurusingha, S.; Ashwath, N.; Subedi, P. Variation in oil content and fatty acid profile of Calophyllum inophyllum L. with fruit maturity and its implications on resultant biodiesel quality. Ind. Crops Prod. 2011, 33, 629–632. [Google Scholar] [CrossRef]
- Agarwal, A.K. Biofuels (alcohols and biodiesel) applications as fuels for internal combustion engines. Prog. Ener. Comb. Sci. 2007, 33, 233–271. [Google Scholar] [CrossRef]
- American Society for Testing Materials, USA. 2023. Available online: https://www.astm.org/products-services/standards-and-publications/standards/bos-standards.html?volume=5&year=2023 (accessed on 1 January 2023).
- Bhuiya, M.M.K.; Rasul, M.G.; Khan, M.M.K.; Ashwath, N.; Azad, A.K.; Hazrat, M.A. Prospects of 2nd generation biodiesel as a sustainable fuel–Part 2: Properties, performance and emission characteristics. Renew. Sustain. Energy Rev. 2016, 55, 1129–1146. [Google Scholar] [CrossRef]
- Chavan, S.B.; Kumbhar, R.R.; Deshmukh, R.B. Calophyllum inophyllum Linn (“honne”) oil, a source for biodiesel production. Res. J. Chem. Sci. 2013, 3, 24–31. [Google Scholar]
- Jahirul, M.I.; Brown, R.J.; Senadeera, W.; Ashwath, N.; Rasul, M.G.; Rahman, M.M.; Hossain, F.M.; Moghaddam, L.; Islam, M.A.; O’Hara, I.M. Physio-chemical assessment of beauty leaf (Calophyllum inophyllum) as second-generation biodiesel feedstock. Energy Rep. 2015, 1, 204–215. [Google Scholar] [CrossRef]
- Leksono, B.; Hendrati, R.L.; Windyarini, E.; Hasnah, T. Variation in biofuel potential of twelve Calophyllum inophyllum populations in Indonesia. Indones. J. For. Res. 2014, 1, 127–138. [Google Scholar]
- Antonovics, J. The effects of a heterogenous environment on the genetics of natural populations. Am. Sci. 1971, 59, 593–595. [Google Scholar]
- Nienstaedt, H. Adaptive Variation—Manifestations in Tree Species and Uses in Forest Management and Tree Improvement. In Proceedings of the 12th Lake States Forest Tree Improvement Conference, Kellogg Biological Station, MI, USA, 18–22 August 1975; pp. 11–23. [Google Scholar]
- Bhat, G.S.; Chauhan, P.S. Provenance variation in seed and seedling traits of Albizzia lebbek Benth. J. Tree Sci. 2002, 21, 52–57. [Google Scholar]
- Usman, M. Provenance Variation for Morphological Traits and Oil Content in Some Jatropha curcas L. Master’s Thesis, Ahmadu Bello University, Zaria, Nigeria, 2014. [Google Scholar]
- Golkar, P.; Arzani, A.; Rezaei, A.M. Genetic analysis of oil content and fatty acid composition in safflower (Carthamus tinctorius L.). J. Am. Oil Chem. Soc. 2011, 88, 975–982. [Google Scholar] [CrossRef]
- Yadav, S.; Suneja, P.; Hussain, Z.; Abraham, Z.; Mishra, S.K. Genetic variability and divergence studies in seed and oil parameters of mahua (Madhuca longifolia Koenig) J.F Macribide accessions. Biol. Bioenergy 2011, 35, 1773–1778. [Google Scholar] [CrossRef]
- Hathurusingha, S.; Ashwath, N.; Midmore, D. Provenance variations in seed-related characters and oil content of Calophyllum inophyllum L. in northern Australia and Sri Lanka. New For. 2011, 41, 89–94. [Google Scholar] [CrossRef]
- Saviztky, A.; Golay, M.J.E. Smoothing and differentiation of data by simplified least squares procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Bureau, S.; Cozzolino, D.; Clark, C.J. Contributions of Fourier-transform mid infrared (FT-MIR) spectroscopy to the study of fruit and vegetables: A review. Post. Biol. Technol. 2019, 148, 1–14. [Google Scholar] [CrossRef]
- Naes, T.; Isaksson, T.; Fearn, T.; Davies, T. A User-Friendly Guide to Multivariate Calibration and Classification, 2nd ed.; NIR: Chichester, UK, 2002; Volume 6. [Google Scholar] [CrossRef]
- Williams, P.; Dardenne, P.; Flinn, P. Tutorial: Items to be include in a report on a near infrared spectroscopy project. J. Near Infrared Spectros. 2017, 25, 85–90. [Google Scholar] [CrossRef]
- Subroto, E.; Manurung, R.; Heeres, H.J.; Broekhuis, A.A. Optimization of mechanical oil extraction from Jatropha curcas L. kernel using response surface method. Ind. Crops Prod. 2015, 63, 294–302. [Google Scholar] [CrossRef]
- Amalia, K.I.; Cerny, M.; Vandenbossche, V.; Rigal, L.; Sablayrolles, C.; Vialle, C.; Suparno, O.; Ariono, D.; Evon, P. Direct Calophyllum oil extraction and resin separation with a binary solvent of n-hexane and methanol mixture. Fuel 2018, 221, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Palinikumaran, B.; Parthiban, K.; Sekar, I.; Umarani, R.; Amirtham, D. Variability studies for seed and seedling traits in Undi (Calophyllum inophyllum L.) from different zones of South India. J. Plant Sci. Res. 2015, 2, 124. [Google Scholar]
- Sreekumar, R. Provenance Variation for Seed Traits, Germination, Seedling Vigour and Oil Content in Calophyllum inophyllum Linn. Master’s Thesis, Navsari Agricultural University, Navsari, India, 2016. [Google Scholar]
- Shamala, K.V.; Ramana, P. Provenance variation in seed oil yield from Calophyllum inophyllum grown in coastal Uttara Kannada and physico chemical characterization of oil. Int. J. Agric. Sci. Res. 2017, 7, 43–48. [Google Scholar]
- Shinde, P.P.; Rane, A.; Bhave, S.; Gunaga, R.; Narkhede, S. Variability and genotype selection in Calophyllum inophyllum for quality fruit yield in the central west coast of India. J. Tree Sci. 2012, 31, 8–14. [Google Scholar]
- Yuniarti, N.; Nurhasybi, N. Viability and Biochemical Content Changes in Seed Storage of Jabon Putih (Anthocephalus cadamba (Roxb) Miq.). J. Trop. For. Manag. 2015, 21, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Lugo, I.; Leopold, A.C. Changes in Soluble Carbohydrates during Seed Storage. Plant Physiol. 1992, 98, 1207–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimić, B.; Popović, R.; Sudarić, A.; Rozman, V.; Kalinovic, I.; Cosic, J. Influence of Storage Condition on Seed Oil Content of Maize, Soybean and Sunflower. Agric. Conspec. Sci. (ACS) 2007, 72, 211–213. [Google Scholar]
- Wu, J.; Gao, S.; Tang, L.; Hou, P.; Gao, J.-H.; Chen, F. The traits, oil content and correlation studies of seed and kernel in Jatropha curcas L. Afr. J. Agric. Res. 2012, 7, 1487–1491. [Google Scholar]
- Reinert, S.; Tassel, D.L.V.; Schlautman, B.; Kane, N.C.; Hulke, B.S. Assessment of the biogeographical variation of seed size and seed oil traits in wild Silphium integrifolium Michx. Genotypes. Plant Genet. Resour. Charact. Util. 2019, 17, 427–436. [Google Scholar] [CrossRef]
- Workman, J.; Weyer, L. Practical Guide to Interpretive Near-Infrared Spectroscopy; CRC Press/Taylor and Francis Group: Boca Raton, FL, USA, 2008. [Google Scholar]
- Sandak, A.; Sandak, J.; Negri, M. Relationship between near-infrared (NIR) spectra and the geographical provenance of timber. Wood Sci. Technol. 2011, 45, 35–48. [Google Scholar] [CrossRef]
- Farhadi, M.; Tigabu, M.; Pietrzykowski, M.; Danusevičius, D.; Odén, P.C. Application of near infrared spectroscopy for authentication of Picea abies seed provenance. New For. 2017, 48, 629–642. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sreekumar, R.; Ashwath, N.; Cozzolino, D. Biogeographic Variability in Kernel Oil and Press Cake Content of Beauty Leaf Tree (Calophyllum inophyllum L.), as Determined by Chemical and Near-Infrared Spectroscopy Analysis. Sustainability 2023, 15, 5529. https://doi.org/10.3390/su15065529
Sreekumar R, Ashwath N, Cozzolino D. Biogeographic Variability in Kernel Oil and Press Cake Content of Beauty Leaf Tree (Calophyllum inophyllum L.), as Determined by Chemical and Near-Infrared Spectroscopy Analysis. Sustainability. 2023; 15(6):5529. https://doi.org/10.3390/su15065529
Chicago/Turabian StyleSreekumar, Rahul, Nanjappa Ashwath, and Daniel Cozzolino. 2023. "Biogeographic Variability in Kernel Oil and Press Cake Content of Beauty Leaf Tree (Calophyllum inophyllum L.), as Determined by Chemical and Near-Infrared Spectroscopy Analysis" Sustainability 15, no. 6: 5529. https://doi.org/10.3390/su15065529