


Effects of Different Levels of Nitrogen Supply on Key Enzyme Activities of Nitrogen Metabolism and Growth Stimulation of Endive (Cichorium endivia L.)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Experimental Design
2.2. Determination of Related Indicators
2.2.1. Determination of the Key Enzyme Activities of Nitrogen Metabolism and Protein Content
2.2.2. Determination of Dry Matter and Nitrogen Accumulation in the Shoots of Plants
2.3. The Model Construction Method of Dry Matter and the Nitrogen Accumulation of Endive
2.4. Acquisition of Environmental Data
2.5. Statistical Analysis
3. Results
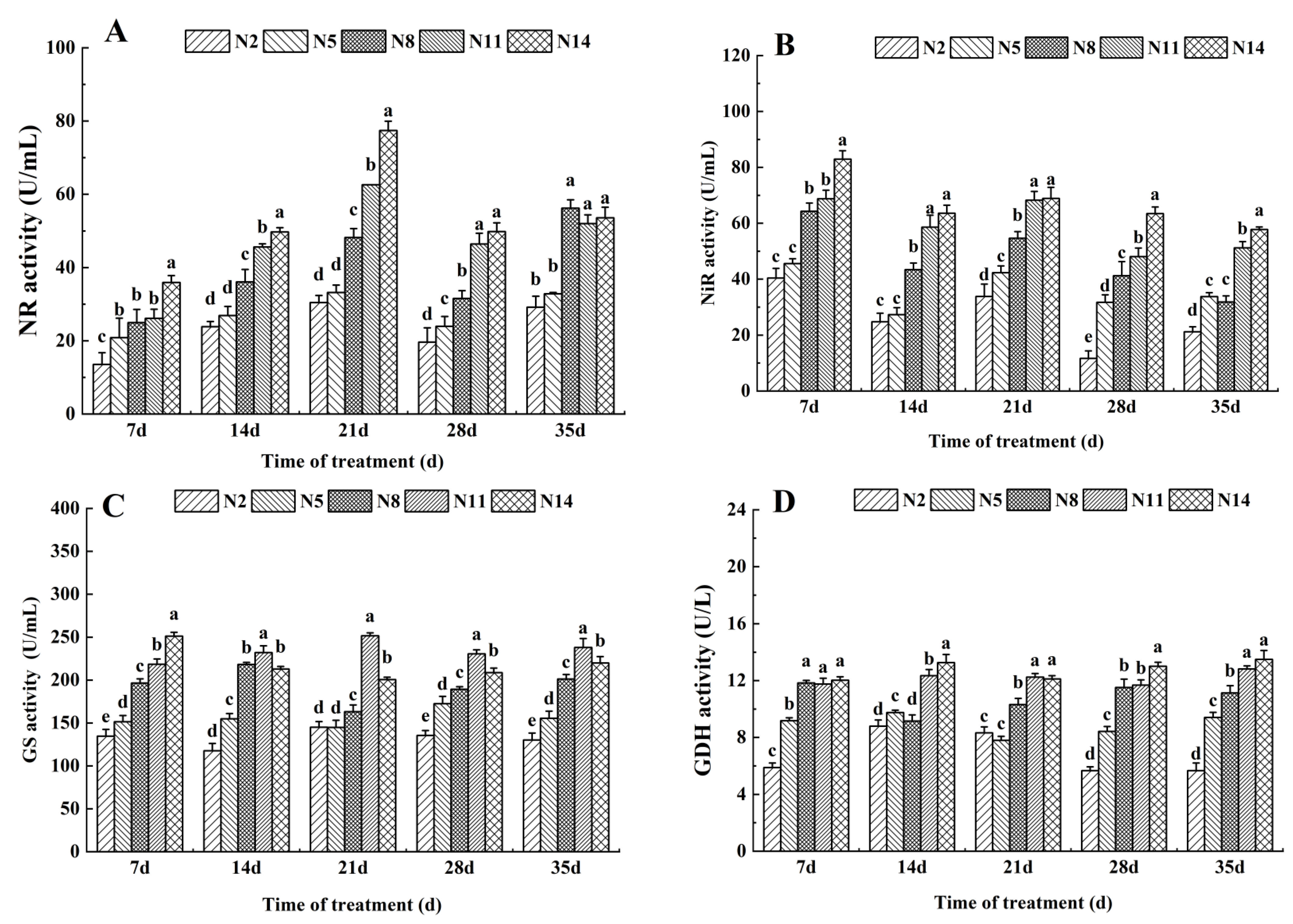
3.1. Effects of Nitrogen Levels on the Activities of Key Enzymes in Nitrogen Metabolism of Endive Leaves
3.2. Effect of Nitrogen Level on the Crude and Total Protein of Endive Plant
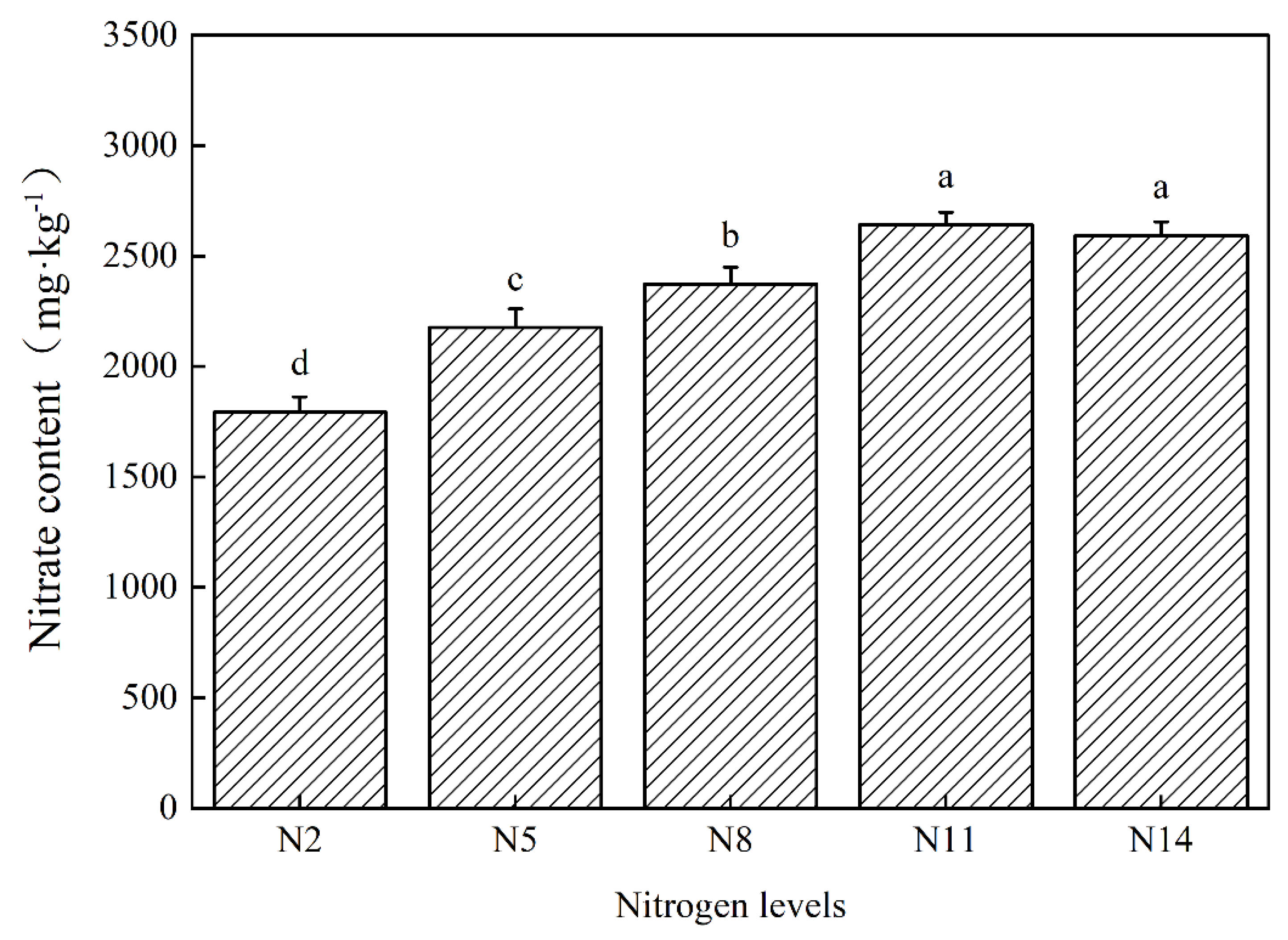
3.3. Effects of Nitrogen Levels on the Nitrate Content in the Leaves of Endive during Harvest
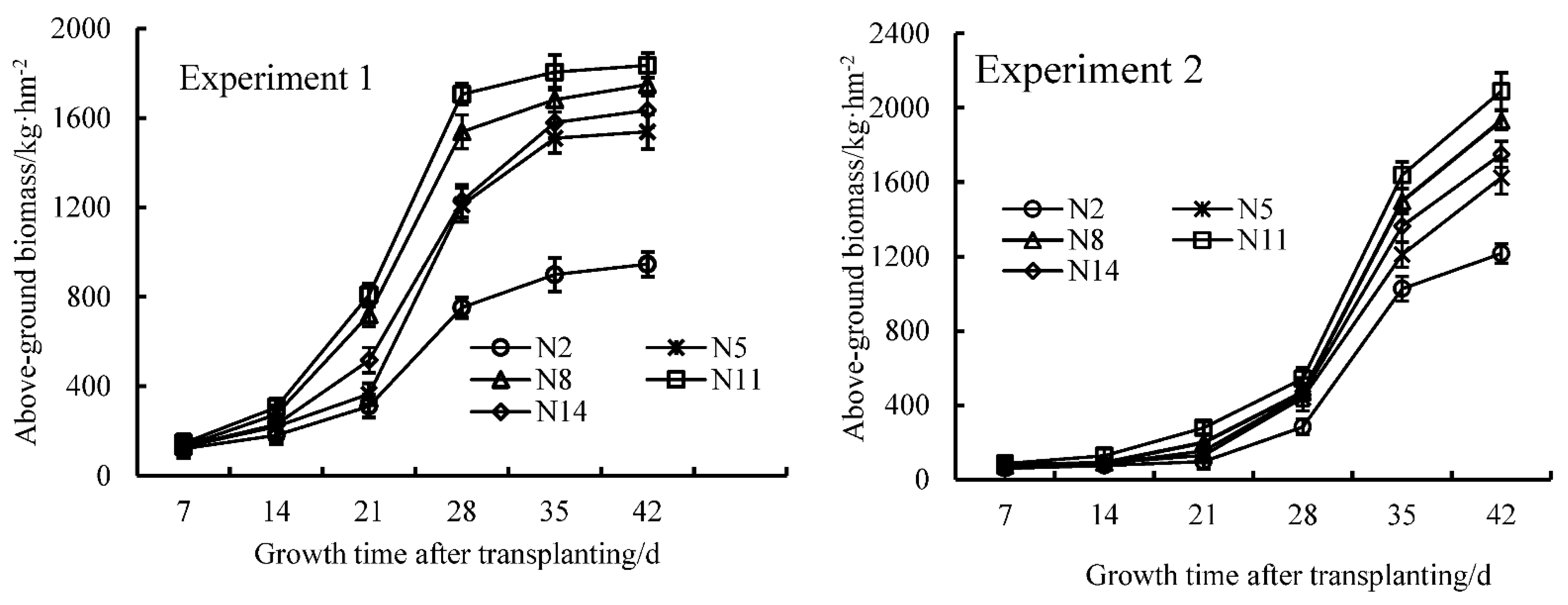
3.4. Effect of the Level of Nitrogen Supplied on Dry Matter Accumulation and the Dynamic Simulation of Endive
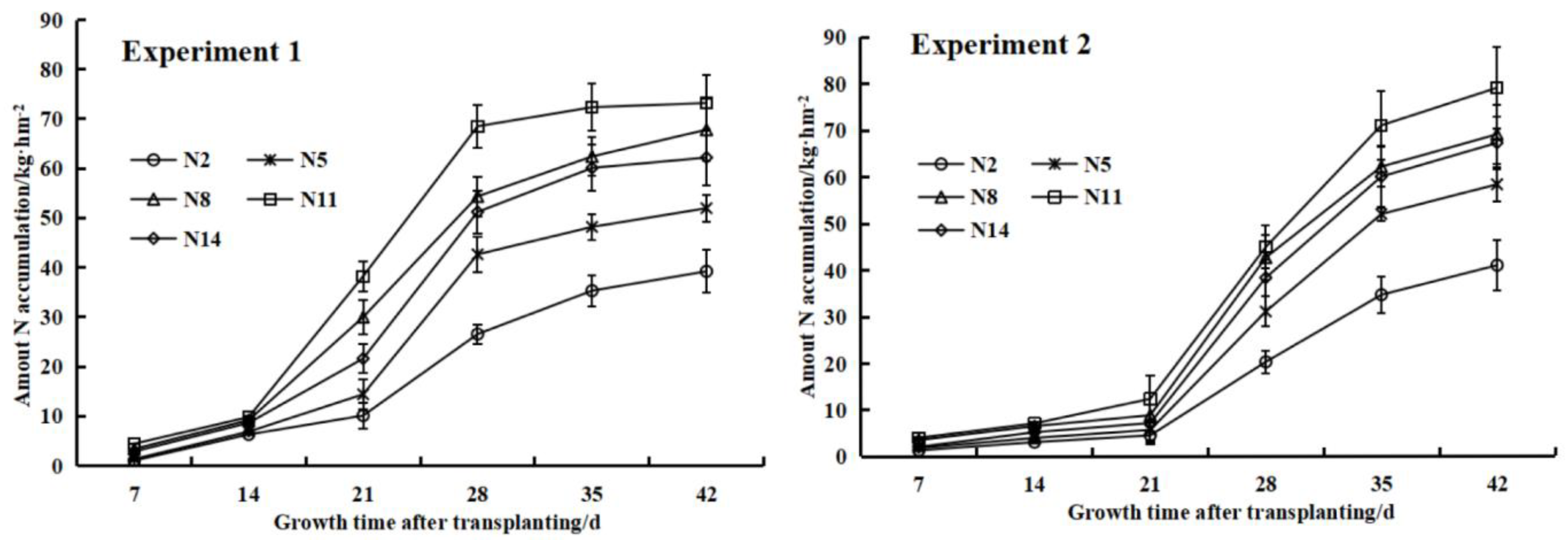
3.5. The Effect of Nitrogen Treatments on the Aboveground Nitrogen Accumulation of Endive and Its Dynamic Simulation
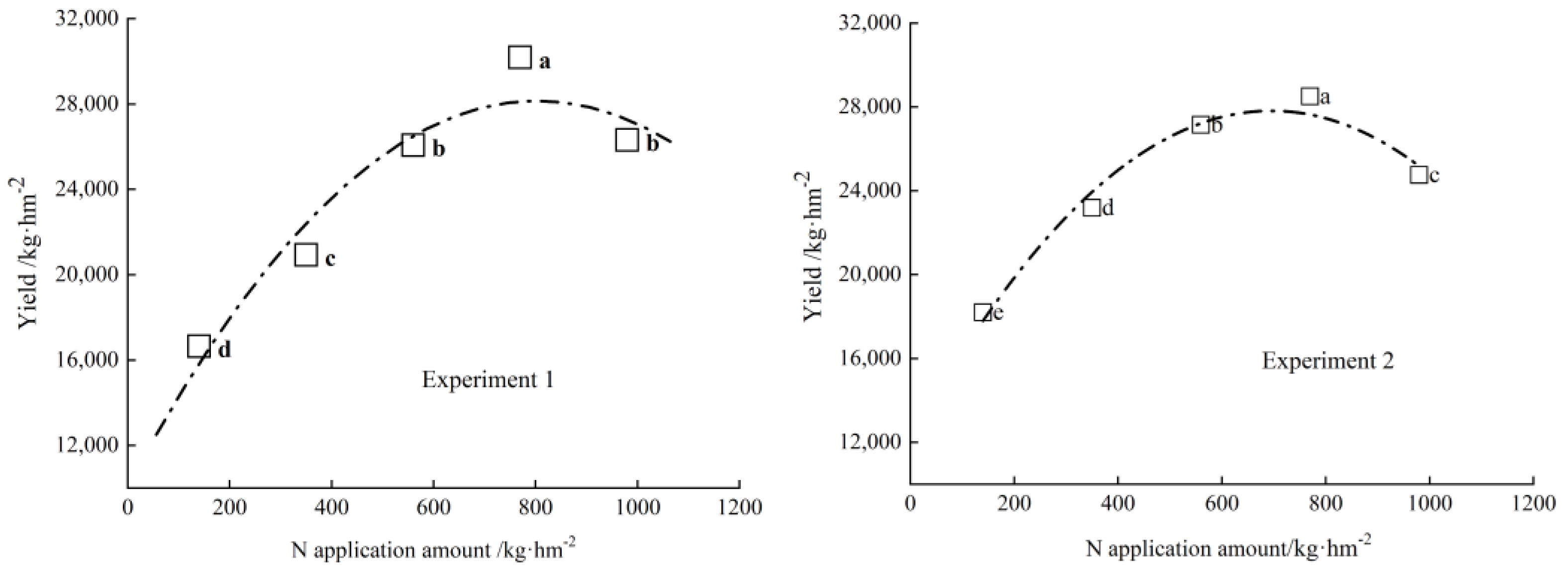
3.6. The Effect of Rates of Nitrogen Application on the Yield of Endive
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koudela, M.; Petříková, K. Nutritional composition and yield of endive cultivars–Cichorium endivia L. Hortic. Sci. 2007, 34, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Zhu, R.H. Physiological Response and Transcriptome Analysis of Chicory Seedlings under Cadmium Stress. Ph.D. Thesis, Sichuan Agricultural University, Ya’an, China, 2020. [Google Scholar]
- Hawrylak-Nowak, B. Comparative effects of selenite and selenate on growth and selenium accumulation in lettuce plants under hydroponic conditions. Plant Growth Regul. 2013, 70, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, A.; Dong, Q.; Wang, X.; Gui, H.; Zhang, H.; Zhang, X.; Song, M. Variations in Nitrogen Metabolism are Closely Linked with Nitrogen Uptake and Utilization Efficiency in Cotton Genotypes under Various Nitrogen Supplies. Plants 2020, 9, 250. [Google Scholar] [CrossRef] [Green Version]
- Boyle, E. Nitrogen pollution knows no bounds. Science 2017, 356, 700–701. [Google Scholar] [CrossRef]
- Tavassoli, A.; Ghanbari, A.; Ahmadian, A. Effect of zinc and manganese nutrition on fruit yield and nutrient concentrations in greenhouse tomato in hydroponic culture. J. Soil Plant Interact. Isfahan Univ. Technol. 2010, 1, 1–7. [Google Scholar]
- Chen, J.; Tang, L.; Liu, X.J.; Cao, W.; Zhu, Y. Modeling plant nitrogen uptake and grain protein accumulation in rice. Sci. Agric. Sin. 2011, 44, 1997–2004. [Google Scholar]
- Kim, Y.B.; Bae, J.H.; Park, M.H. Effects of supplemental lighting on growth and yield of sweet pepper (Capsicum annuum L.) in hydroponic culture under low levels of natural light in winter. Hortic. Sci. Technol. 2011, 29, 317–325. [Google Scholar]
- Diao, M.; Dai, J.F.; Luo, W.H.; Yuan, C.M.; Bu, C.X.; Xian, K.M.; Zhang, S.F.; Xu, R. Growth and yield prediction model of greenhouse sweet pepper. Trans. CSAE 2009, 25, 241–246. [Google Scholar]
- Stuart, N.W. Adaptation of the micro-Kjeldahl method for the determination of nitrogen in plant tissues. Plant Physiol. 1936, 11, 173. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ma, F.; Diao, M.; Fan, H.; Cui, J.; Jia, B.; He, H.; Liu, Q. Dynamic simulation of plant growth, nitrogen accumulation, and utilization in processed tomato plants under different nitrogen application levels. Chin. J. Appl. Ecol. 2014, 25, 1043–1050. [Google Scholar]
- Xin, W.; Zhang, L.; Zhang, W.; Gao, J.; Yi, J.; Zhen, X.; Du, M.; Zhao, Y.; Chen, L. Morphological and Physiological Characteristics of Rice Cultivars with Higher Yield and Nitrogen Use Efficiency at Various Nitrogen Rates. Agronomy 2022, 12, 358. [Google Scholar] [CrossRef]
- Zhao, Q.Z.; Chen, J.R.; Liu, H.; Qiao, J.F.; Gao, T.M.; Yang, H.X.; Wang, J.H. The relationship between key enzyme activities of nitrogen assimilation in rice and changes in leaf color. Chin. Agric. Sci. 2008, 09, 2607–2616. [Google Scholar]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification. Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Yu, B.; Qin, S.J.; Lv, D.D. Appropriate amount of zinc supply significantly improves carbon and nitrogen absorption and assimilation efficiency of Pingyi sweet tea seedlings. J. Plant Nutr. Fertil. 2021, 27, 97–108. [Google Scholar]
- Ji, C.; Li, J.; Jiang, C.; Zhang, L.; Shi, L.; Xu, F.; Cai, H. Zinc and nitrogen synergistic act on root-to-shoot translocation and preferential distribution in rice. J. Adv. Res. 2021, 35, 187–198. [Google Scholar] [CrossRef]
- Zhang, H.Z.; Xu, H.Y. Research progress of related enzymes in the process of nitrogen assimilation in plants. North. Hortic. 2011, 20, 180–183. [Google Scholar]
- Yin, H.; Li, B.; Wang, X.; Xi, Z. Effect of ammonium and nitrate supplies on nitrogen and sucrose metabolism of Cabernet Sauvignon (Vitis vinifera cv.). J. Sci. Food Agric. 2020, 100, 5239–5250. [Google Scholar] [CrossRef]
- Wang, H.; Yang, Z.; Yu, Y.; Chen, S.; He, Z.; Wang, Y.; Jiang, L.; Wang, G.; Yang, C.; Liu, B.; et al. Drought Enhances Nitrogen Uptake and Assimilation in Maize Roots. Agron. J. 2017, 109, 39–46. [Google Scholar] [CrossRef]
- Xu, H.C.; Shang, J.; Liu, M.H.; Zhang, F.; Li, Y.Y. Research progress on enzymes related to nitrogen metabolism. Anhui Agric. Sci. 2022, 50, 17–20. [Google Scholar]
- Liu, X.J.; Zhang, J.X.; Ye, F.; Qi, P. Effects of nitrogen application on nitrogen metabolism and nitrogen accumulation in alfalfa. J. Nucl. Agric. 2015, 29, 1399–1405. [Google Scholar]
- Diao, Z.W.; Yu, X.; Wang, Y.B.; Li, C.F.; Gai, Z.J.; Zhang, Y.F.; Ma, F.M. Study on the coupling regulation of nitrate reductase and nitrite reductase in sugar beet. J. Nucl. Agric. 2014, 28, 138–145. [Google Scholar]
- Li, D.F.; Li, Z.Y.; Li, S.Q.; Wang, Q.J.; Shao, M.G.; Li, F.G. Effects of nitrogen application on nitrate nitrogen and nitrate reductase activity in winter wheat plants of different varieties. Northwest Bot. 2006, 1, 104–109. [Google Scholar]
- Chen, H.; Wang, L.; Zhao, C.L.; Xiao, F.; Zheng, Y.F.; Liu, S.C. Effects of nitrogen and red-blue composite light ratio on nitrite reductase activity and gene expression in amaranth seedlings. J. China Agric. Univ. 2021, 26, 61–71. [Google Scholar]
- Kyaing, S.M.; Gu, L.H.; Cheng, H.M. The role of nitrate reductase and nitrite reductase in plant. Curr. Biotechnol. 2011, 1, 159–164. [Google Scholar]
- Zhang, Y.Y.; Song, X.F.; Xu, J.B.; Zhou, Q.; Liu, H.Q.; Wang, Y.; Zhang, Z.Y.; Song, H.L. Effects of different nitrogen concentrations on the growth and physiological indicators of hyacinth, especially the activities of key enzymes in nitrogen metabolism. Chin. J. Environ. 2021, 30, 39–46. [Google Scholar]
- Zhang, T.; Shi, Y.; Piao, F.; Sun, Z. Effects of different LED sources on the growth and nitrogen metabolism of lettuce. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 134, 231–240. [Google Scholar]
- Jabeen, N.; Ahmad, R. Growth response and nitrogen metabolism of sunflower (Helianthus annuus L.) to vermicompost and biogas slurry under salinity stress. J. Plant Nutr. 2017, 40, 104–114. [Google Scholar]
- Wani, A.S.; Tahir, I.; Ahmad, S.S.; Dar, R.A.; Nisar, S. Efficacy of 24-epibrassinolide in improving the nitrogen metabolism and antioxidant system in chickpea cultivars under cadmium and/or NaCl stress. Sci. Hortic. 2017, 225, 48–55. [Google Scholar] [CrossRef]
- Forde, B.G.; Lea, P.J. Glutamate in plants: Metabolism, regulation, and signaling. J. Exp. Bot. 2007, 58, 2339–2358. [Google Scholar] [CrossRef]
- Zhang, C.; Peng, S.; Peng, X.; Chavez, A.Q.; Bennett, J. Response of glutamine synthetase isoforms to nitrogen sources in rice (Oryza sativa L.) roots. Plant Sci. 1997, 125, 163–170. [Google Scholar] [CrossRef]
- Wang, J.F.; Wang, Z.Z.; Gu, F.X.; Mu, H.M.; Wang, Y.; Duan, J.Z.; Feng, W.; Wang, Y.H.; Guo, T.C. Effects of nitrogen density regulation on carbon and nitrogen metabolism and yield of two winter wheat cultivars. Sci. Agric. Sin. 2021, 54, 4070–4083. [Google Scholar]
- Kishore Kumarkumar, R.; Bulle, M.; Wany, A.; Gupta, K.J. An overview of important enzymes involved in nitrogen assimilation of plants. Nitrogen Metab. Plants 2020, 2057, 1–13. [Google Scholar]
- Wang, F.; Gao, J.; Yong, J.W.; Liu, Y.; Cao, D.; He, X. Glutamate over-accumulation may serve as an endogenous indicator of tricarboxylic acid (TCA) cycle suppression under NH4+ nutrition in wheat (Triticum aestivum L.) seedlings. Environ. Exp. Bot. 2020, 177, 104130. [Google Scholar] [CrossRef]
- Ashraf, M.; Shahzad, S.M.; Imtiaz, M.; Rizwan, M.S. Salinity effects on nitrogen metabolism in plants—Focusing on the activities of nitrogen metabolizing enzymes: A review. J. Plant Nutr. 2018, 41, 1065–1081. [Google Scholar] [CrossRef]
- Moda, L.R.; de Mello Prado, R.; de Souza Júnior, J.P.; da Silva Júnior, G.B.; dos Santos, L.C.N.; Soares, M.V.G.; Kadyampakeni, D.M. Response of orange seedlings to the proportion of nitrate-ammonium in the nutrient solution and the benefits of phosphorus in ammonia toxicity. Sci. Hortic. 2021, 285, 110166. [Google Scholar] [CrossRef]
- Wang, J.; Nan, Z.; Christensen, M.J.; Zhang, X.; Tian, P.; Zhang, Z. Effect of Epichloë gansuensis endophyte on the nitrogen metabolism, nitrogen use efficiency, and stoichiometry of Achnatherum inebrians under nitrogen limitation. J. Agric. Food Chem. 2018, 66, 4022–4031. [Google Scholar] [CrossRef] [PubMed]
- Jing, B.; Diao, M.; Zhang, K.; Guo, P.; Wan, W.; Niu, N. Optimal fertilization effect of processed tomato based on critical nitrogen concentration. Soil Fertil. Sci. China 2020, 06, 205–212. [Google Scholar]
- Cui, Z.; Zhang, F.; Chen, X.; Miao, Y.; Li, J.; Shi, L.; Xu, J.; Ye, Y.; Liu, C.; Yang, Z.; et al. On-farm evaluation of an in-season nitrogen management strategy based on soil Nmin test. Field Crop. Res. 2008, 105, 48–55. [Google Scholar]
- Imran, M.; Sun, X.; Hussain, S.; Ali, U.; Rana, M.S.; Rasul, F.; Saleem, M.H.; Moussa, M.G.; Bhantana, P.; Afzal, J.; et al. Molybdenum-induced effects on nitrogen metabolism enzymes and elemental profile of winter wheat (Triticum aestivum L.) under different nitrogen sources. Int. J. Mol. Sci. 2019, 20, 3009. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhang, Z.; Tian, H.; Mo, Z.; Ashraf, U.; Duan, M.; Wang, Z.; Wang, S.; Tang, X.; Pan, S. Roles of nitrogen deep placement on grain yield, nitrogen use efficiency, and antioxidant enzyme activities in mechanical pot-seedling transplanting rice. Agronomy 2020, 10, 1252. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Compound | Chemical Formula | Dosage (mg/L) |
|---|---|---|---|
| Liquid A | Calcium nitrate | Ca(NO3)2·4H2O | 472 |
| Potassium nitrate | KNO3 | 267 | |
| Ammonium nitrate | NH4NO3 | 53 | |
| Liquid B | Potassium dihydrogen phosphate | KH2PO4 | 100 |
| Potassium sulfate | K2SO4 | 116 | |
| Magnesium sulfate heptahydrate | MgSO4·7H2O | 246 | |
| Liquid C | Chelated iron | NaFe-EDTA | 13 |
| Boric acid | H3BO3 | 2.86 | |
| Manganese sulfate | MnSO4·4H2O | 2.13 | |
| Zinc sulfate | ZnSO4·7H2O | 0.22 | |
| Copper sulfate | CuSO4·5H2O | 0.05 | |
| Ammonium molybdate | (NH4)6Mo7O2·4H2O | 0.02 |
| Treatment | Dosage of Salt Compounds (mg·L−1) | Nitrogen Content (mmol·L−1) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Calcium Nitrate Tetrahydrate | Potassium Nitrate | Ammonium Nitrate | Dipotassium Phosphate | Magnesium Sulfate Heptahydrate | Potassium Dihydrogen Phosphate | Potassium Sulfate | Calcium Sulfate Dihydrate | ||
| N2 | 236 | 296 | 246 | 0 | 116 | 172 | 2 | ||
| N5 | 472 | 101 | 129 | 246 | 0 | 197 | 5 | ||
| N8 | 472 | 267 | 53 | 246 | 100 | 116 | 8 | ||
| N11 | 472 | 267 | 173 | 246 | 100 | 116 | 11 | ||
| N14 | 472 | 267 | 293 | 246 | 100 | 116 | 14 | ||
| Indicators | Treatments | Growth to 7 d | Growth to 14 d | Growth to 21 d | Growth to 28 d | Growth to 35 d |
|---|---|---|---|---|---|---|
| Dry-matter production/(g·m−2) | N2 | 4.291 ± 0.27 d | 15.353 ± 0.87 d | 29.034 ± 1.18 d | 79.785 ± 4.11 d | 106.163 ± 7.20 c |
| N5 | 4.774 ± 0.14 c | 16.574 ± 0.78 d | 52.272 ± 2.67 c | 129.344 ± 4.69 c | 141.954 ± 4.22 b | |
| N8 | 5.518 ± 0.34 b | 22.323 ± 0.88 b | 85.533 ± 3.96 a | 152.069 ± 4.03 b | 178.852 ± 6.55 a | |
| N11 | 6.549 ± 0.34 a | 24.233 ± 1.46 a | 70.719 ± 4.20 b | 165.573 ± 3.98 a | 184.283 ± 8.87 a | |
| N14 | 5.562 ± 0.19 b | 20.551 ± 0.71 | 54.344 ± 4.86 c | 128.902 ± 3.52 c | 149.174 ± 7.49 b | |
| N concentration (%) | N2 | 3.233 ± 0.37 b | 3.687 ± 0.08 c | 3.526 ± 0.10 c | 3.454 ± 0.21 bc | 3.529 ± 0.16 b |
| N5 | 3.696 ± 0.16 a | 3.714 ± 0.14 c | 3.535 ± 0.15 c | 3.136 ± 0.28 c | 3.448 ± 0.34 b | |
| N8 | 3.817 ± 0.09 a | 4.084 ± 0.16 b | 3.906 ± 0.36 b | 3.365 ± 0.29 c | 3.562 ± 0.22 b | |
| N11 | 3.825 ± 0.13 a | 4.152 ± 0.24 b | 4.337 ± 0.22 a | 3.733 ± 0.27 ab | 3.869 ± 0.228 a | |
| N14 | 3.585 ± 0.14 a | 4.510 ± 0.29 a | 4.443 ± 0.26 a | 3.861 ± 0.30 a | 4.021 ± 0.18 a | |
| Total protein/(g·m−2) | N2 | 0.868 ± 0.12 c | 3.553 ± 0.76 b | 6.403 ± 0.63 e | 17.111 ± 2.90 c | 23.462 ± 2.57 d |
| N5 | 1.102 ± 0.1 bc | 3.845 ± 0.62 b | 8.336 ± 1.01 d | 26.784 ± 2.42 b | 30.504 ± 4.43 c | |
| N8 | 1.313 ± 0.10 ab | 5.704 ± 0.85 a | 17.184 ± 2.01 b | 31.990 ± 5.09 b | 39.650 ± 5.76 ab | |
| N11 | 1.562 ± 0.47 a | 6.277 ± 0.34 a | 23.121 ± 1.43 a | 41.641 ± 4.49 a | 43.994 ± 2.44 a | |
| N14 | 1.265 ± 0.34 ab | 5.966 ± 1.00 a | 14.524 ± 2.06 c | 31.133 ± 3.82 b | 37.472 ± 3.98 b |
| Treatment | Experiment 1 | Experiment 2 | ||
|---|---|---|---|---|
| Dry Matter Accumulation | Nitrogen Accumulation | Dry Matter Accumulation | Nitrogen Accumulation | |
| /(kg·hm−2) | /(kg·hm−2) | /(kg·hm−2) | /(kg·hm−2) | |
| N2 | 1191.932 e | 38.680 e | 1302.671 e | 47.732 d |
| N5 | 1497.270 d | 53.456 d | 1660.137 d | 64.895 c |
| N8 | 1840.474 b | 70.075 b | 1943.472 b | 77.281 b |
| N11 | 1950.402 a | 75.803 a | 2041.075 a | 85.800 a |
| N14 | 1596.333 c | 63.961 c | 1772.532 c | 77.520 b |
| Experiment | Treatment | Fitting Equation | CorrelationCoefficient | T1 | T2 | Tm | Vm | T2 − T1 |
|---|---|---|---|---|---|---|---|---|
| R2 | (d) | (d) | (d) | (kg·hm−2·d−1) | (d) | |||
| Experiment 1 | N2 | DMT = 1199.241/(1 + 436.771e−0.2389T) | 0.998 ** | 19.93 | 30.95 | 25.44 | 71.642 | 11.02 |
| N5 | DMT = 1473.714/(1 + 71474e−0.4843T) | 0.996 ** | 20.36 | 25.8 | 23.08 | 178.450 | 5.44 | |
| N8 | DMT = 1833.875/(1 + 918.172e−0.3014T) | 0.999 ** | 18.27 | 27 | 22.63 | 138.191 | 8.73 | |
| N11 | DMT = 1915.492/(1 + 2940.9e−0.3714T) | 0.998 ** | 17.96 | 25.05 | 21.5 | 177.862 | 7.09 | |
| N14 | DMT = 1578.544/(1 + 921.942e−0.2939T) | 0.998 ** | 18.75 | 27.71 | 23.23 | 115.985 | 8.96 | |
| Experiment 2 | N2 | DMT = 1397.910/(1 + 1046.10e−0.2226T) | 0.998 ** | 25.32 | 37.15 | 31.24 | 77.794 | 11.83 |
| N5 | DMT = 1769.764/(1 + 481.605e−0.1984T) | 0.999 ** | 24.5 | 37.78 | 31.14 | 87.771 | 13.28 | |
| N8 | DMT = 2038.854/(1 + 307.342e−0.1869T) | 0.998 ** | 23.59 | 37.68 | 30.64 | 95.290 | 14.09 | |
| N11 | DMT = 2238.432/(1 + 195.10e−0.1672T) | 0.997 ** | 23.66 | 39.41 | 31.53 | 93.595 | 15.75 | |
| N14 | DMT = 1891.533/(1 + 488.02e−0.1988T) | 0.998 ** | 24.51 | 37.76 | 31.14 | 94.010 | 13.25 |
| Experiment | Treatment | Fitting Equation | Correlation Coefficient | T1 | T2 | Tm | Vm | T2 − T1 |
|---|---|---|---|---|---|---|---|---|
| R2 | (d) | (d) | (d) | (kg·hm−2·d−1) | (d) | |||
| Experiment 1 | N2 | DMT = 39.651/(1 + 571.58e−0.2556T) | 0.997 ** | 19.68 | 29.99 | 24.83 | 2.5343 | 10.31 |
| N5 | DMT = 52.33/(1 + 3055.1e−0.3363T) | 0.997 ** | 19.95 | 27.78 | 23.86 | 4.3993 | 7.83 | |
| N8 | DMT = 68.87/(1 + 289.191e−0.2432T) | 0.999 ** | 17.89 | 28.72 | 23.3 | 4.1872 | 10.83 | |
| N11 | DMT = 73.56/(1 + 1555.7e−0.3490T) | 0.999 ** | 17.29 | 24.83 | 21.06 | 6.4181 | 7.54 | |
| N14 | DMT = 63.68/(1 + 448.630e−0.2633T) | 0.996 ** | 18.19 | 28.19 | 23.19 | 4.192 | 10 | |
| Experiment 2 | N2 | DMT = 51.29/(1 + 1011.62e−0.2213T) | 0.998 ** | 25.32 | 37.23 | 31.27 | 2.8368 | 11.91 |
| N5 | DMT = 68.66/(1 + 751.121e−0.2192T) | 0.999 ** | 24.2 | 36.22 | 30.21 | 3.7618 | 12.02 | |
| N8 | DMT = 80.92/(1 + 507.254e−0.2148T) | 0.998 ** | 22.87 | 35.14 | 29 | 4.3443 | 12.27 | |
| N11 | DMT = 93.10/(1 + 274.872e−0.1878T) | 0.996 ** | 22.89 | 36.91 | 29.9 | 4.372 | 14.02 | |
| N14 | DMT = 82.45/(1 + 727.723e−0.2186T) | 0.998 ** | 24.12 | 36.16 | 30.14 | 4.5069 | 12.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Chai, Y.; Guan, S.; Tuerti, T.; Liu, H.; Zhang, Z.; Diao, M. Effects of Different Levels of Nitrogen Supply on Key Enzyme Activities of Nitrogen Metabolism and Growth Stimulation of Endive (Cichorium endivia L.). Sustainability 2022, 14, 15159. https://doi.org/10.3390/su142215159
Ma Y, Chai Y, Guan S, Tuerti T, Liu H, Zhang Z, Diao M. Effects of Different Levels of Nitrogen Supply on Key Enzyme Activities of Nitrogen Metabolism and Growth Stimulation of Endive (Cichorium endivia L.). Sustainability. 2022; 14(22):15159. https://doi.org/10.3390/su142215159
Chicago/Turabian StyleMa, Yadong, Yaqian Chai, Sihui Guan, Tayier Tuerti, Huiying Liu, Zhijun Zhang, and Ming Diao. 2022. "Effects of Different Levels of Nitrogen Supply on Key Enzyme Activities of Nitrogen Metabolism and Growth Stimulation of Endive (Cichorium endivia L.)" Sustainability 14, no. 22: 15159. https://doi.org/10.3390/su142215159