
Nano-Priming against Abiotic Stress: A Way Forward towards Sustainable Agriculture

 ,
,  , , , ,
, , , ,  , and
, and 
Abstract
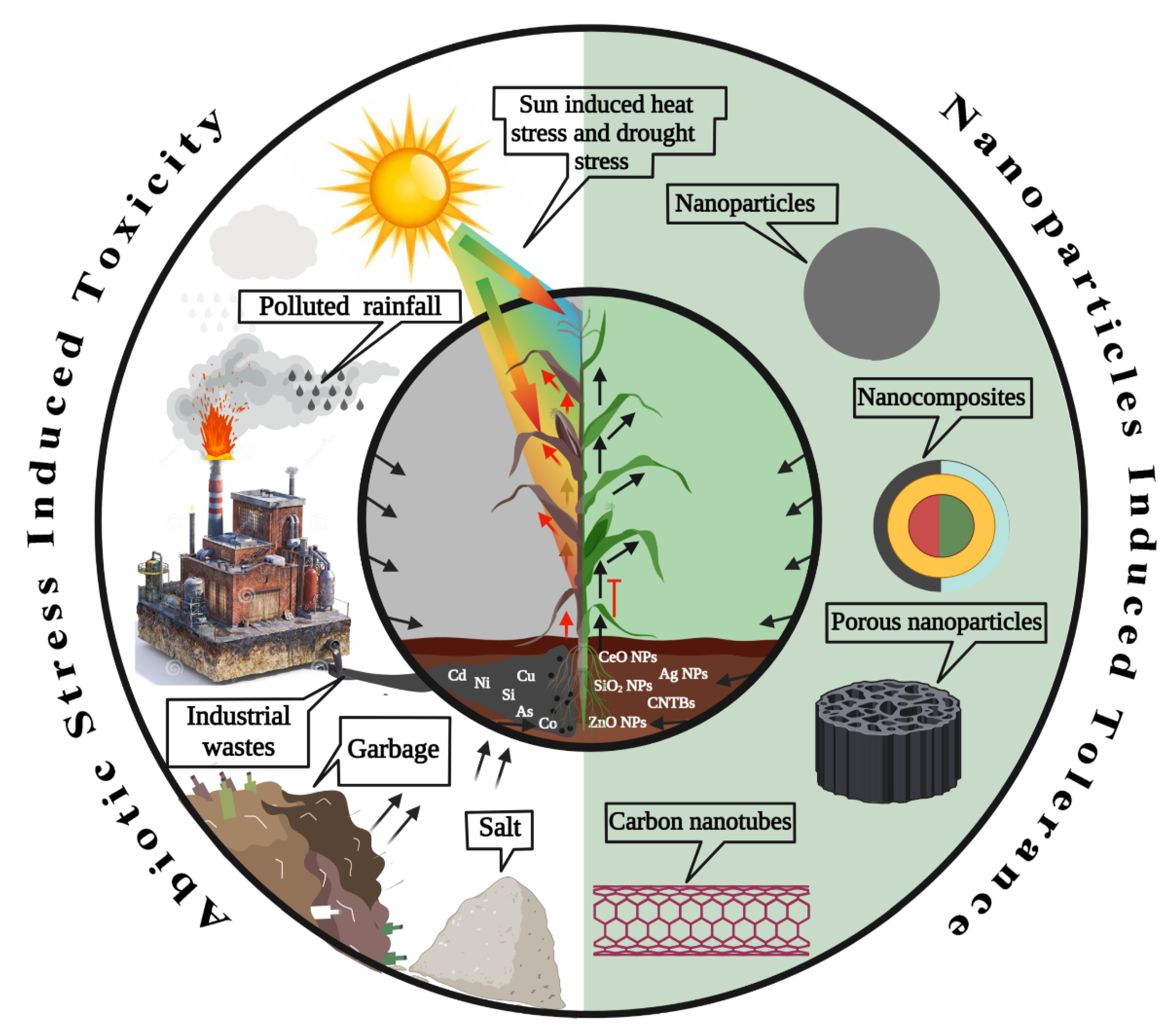
:1. Introduction
2. Nanotechnology and Agriculture
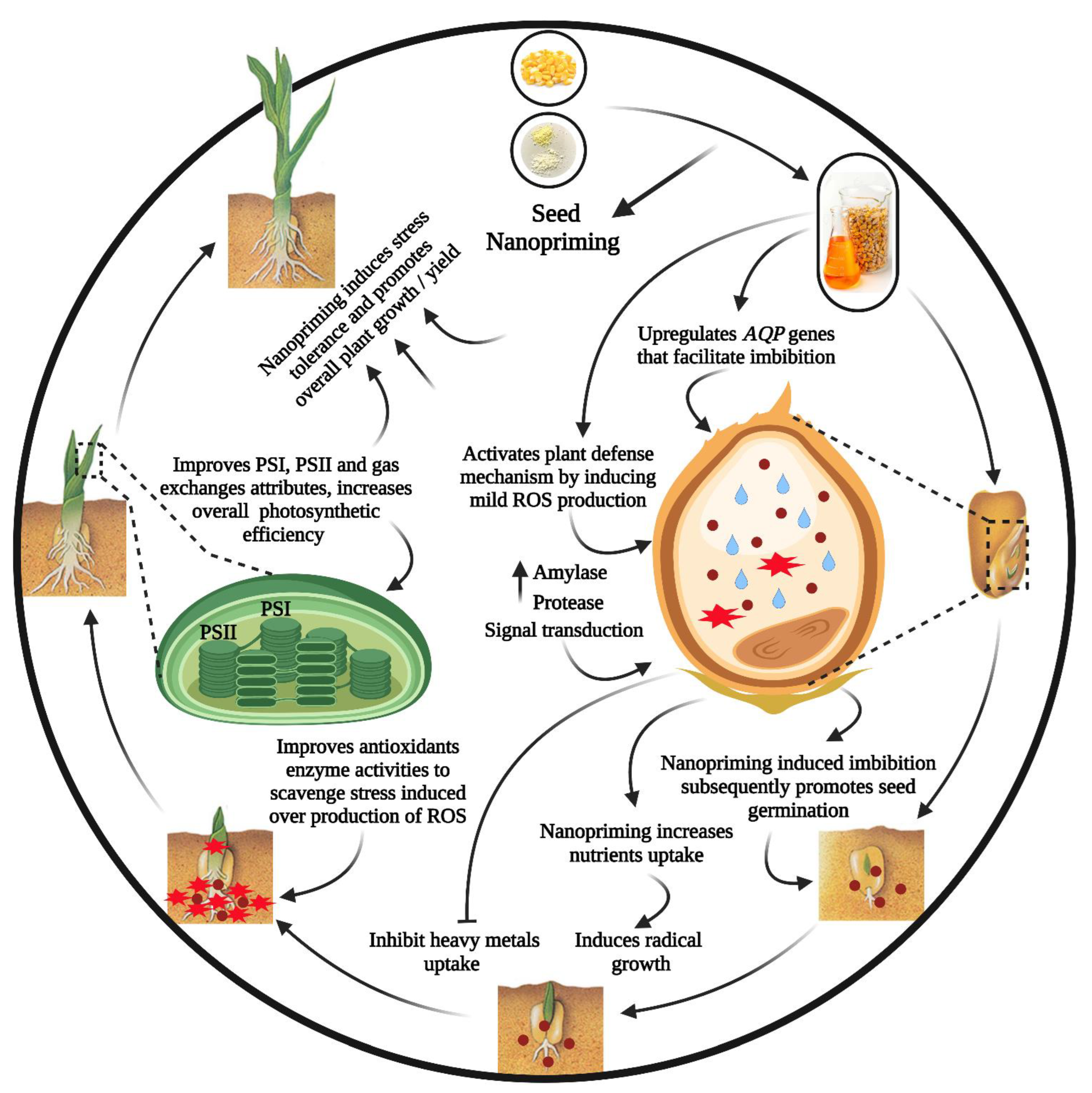
3. Seed Nano-Priming
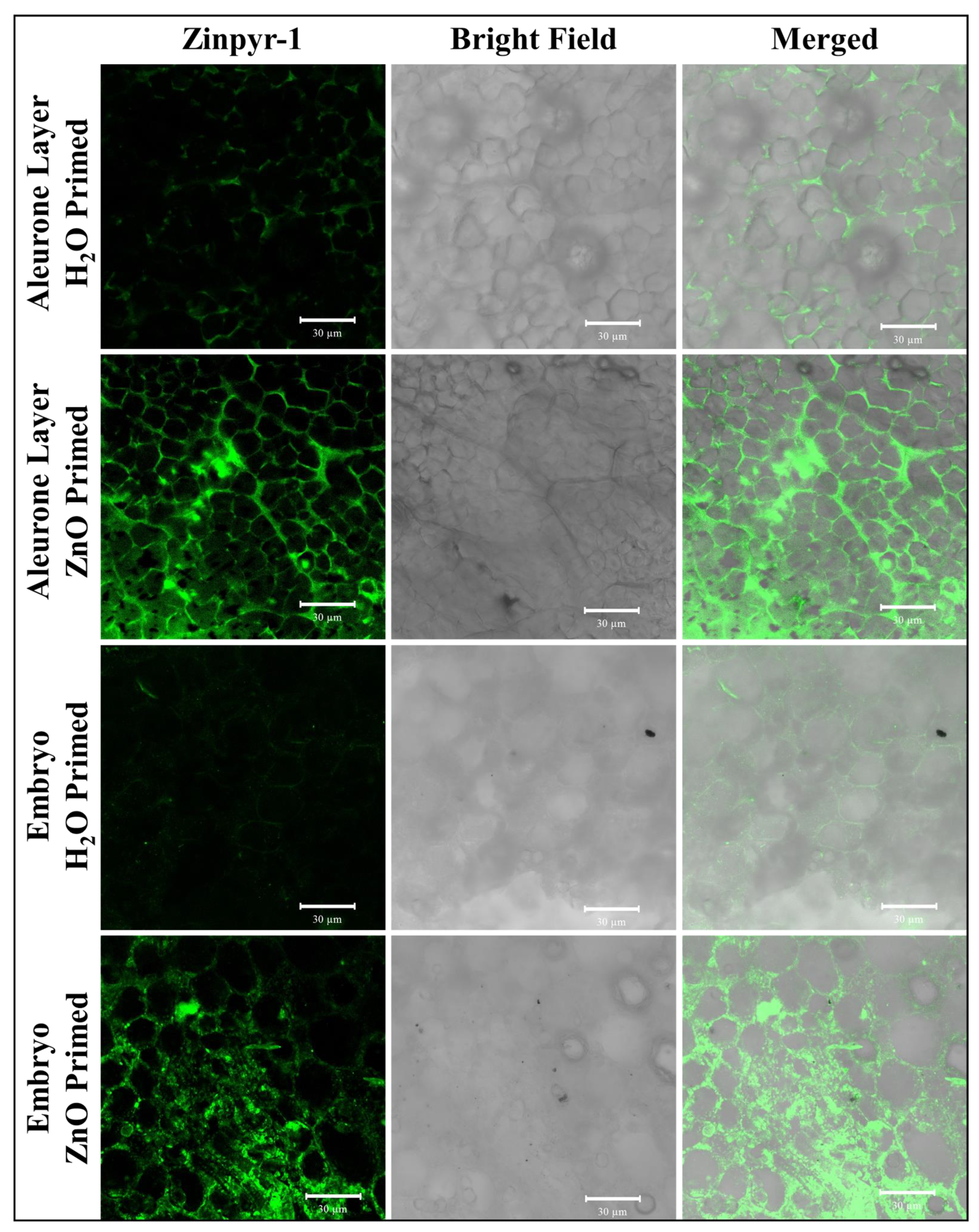
4. Effect on Seed Germination
5. Effects on Plant Growth and Physiology under Abiotic Stress
6. Effects on Plant Metabolism under Abiotic Stress
7. Priming-Induced Molecular Responses against Abiotic Stresses
8. Effects on Photosynthesis under Abiotic Stress
9. Effects on Nutrient Uptake and Regulation
10. Effects on Plant Antioxidant Defense Systems
11. Crosstalk with Plant Growth-Promoting Microbiota
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mueller, N.D.; Gerber, J.S.; Johnston, M.; Ray, D.K.; Ramankutty, N.; Foley, J.A. Closing Yield Gaps through Nutrient and Water Management. Nature 2012, 490, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.M.; Demokritou, P.; Dokoozlian, N.; Hendren, C.O.; Karn, B.; Mauter, M.S.; Sadik, O.A.; Safarpour, M.; Unrine, J.M.; Viers, J. Nanotechnology for Sustainable Food Production: Promising Opportunities and Scientific Challenges. Environ. Sci. Nano. 2017, 4, 767–781. [Google Scholar] [CrossRef]
- Afridi, M.S.; Javed, M.A.; Ali, S.; de Medeiros, F.H.V.; Ali, B.; Salam, A.; Sumaira; Marc, R.A.; Alkhalifah, D.H.M.; Selim, S.; et al. New Opportunities in Plant Microbiome Engineering for Increasing Agricultural Sustainability under Stressful Conditions. Front. Plant Sci. 2022, 13, 899464. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and Biotic Stress Combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Zhao, L.; Lu, L.; Wang, A.; Zhang, H.; Huang, M.; Wu, H.; Xing, B.; Wang, Z.; Ji, R. Nano-Biotechnology in Agriculture: Use of Nanomaterials to Promote Plant Growth and Stress Tolerance. J. Agric. Food Chem. 2020, 68, 1935–1947. [Google Scholar] [CrossRef]
- Connor, R. The United Nations World Water Development Report 2015: Water for a Sustainable World; UNESCO Publishing: Paris, France, 2015; Volume 1, ISBN 9231000713. [Google Scholar]
- Shabala, S.; Bose, J.; Hedrich, R. Salt Bladders: Do They Matter? Trends Plant Sci. 2014, 19, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Salam, A.; Ali, A.; Afridi, M.S.; Ali, S.; Ullah, Z. Agrobiodiversity: Effect of Drought Stress on the Eco-Physiology and Morphology of Wheat. In Biodiversity, Conservation and Sustainability in Asia; Springer: Cham, Switzerland, 2022; pp. 597–618. [Google Scholar]
- Hayat, K.; Menhas, S.; Hayat, S.; Salam, A.; Aftab, T.; Zhou, Y.; Afridi, M.S.; Khan, A.A.; Zhou, P. Stress-Tolerant Species of Medicinal Plants and Phytoremediation Potential. In Environmental Challenges and Medicinal Plants: Sustainable Production Solutions under Adverse Conditions; Aftab, T., Ed.; Springer: Cham, Switzerland, 2022; pp. 433–448. [Google Scholar]
- Fenner, K.; Canonica, S.; Wackett, L.P.; Elsner, M. Evaluating Pesticide Degradation in the Environment: Blind Spots and Emerging Opportunities. Science 2013, 341, 752–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anwar, T.; Qureshi, H.; Perveen, N.; Mahmood, S.; Zulqurnain Haider, M.; Mumtaz, S.; Nawaz, H.; Khan, S.A.; Hafeez, A.; Imran Tipu, M.; et al. Herbicidal Effectiveness of Wild Poisonous Plant Rhazya Stricta Using Different Media by the Sandwich Method. Pak. J. Bot. 2022. [Google Scholar] [CrossRef]
- Khan, A.R.; Azhar, W.; Wu, J.; Ulhassan, Z.; Salam, A.; Zaidi, S.H.R.; Yang, S.; Song, G.; Gan, Y. Ethylene Participates in Zinc Oxide Nanoparticles Induced Biochemical, Molecular and Ultrastructural Changes in Rice Seedlings. Ecotoxicol. Environ. Saf. 2021, 226, 112844. [Google Scholar] [CrossRef]
- Javaid, M.H.; Khan, A.R.; Salam, A.; Neelam, A.; Azhar, W.; Ulhassan, Z.; Gan, Y. Exploring the Adaptive Responses of Plants to Abiotic Stresses Using Transcriptome Data. Agriculture 2022, 12, 211. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, Mechanism and Health Effects of Some Heavy Metals. Interdiscip. Toxicol. 2014, 7, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, S.; Modi, P.; Dave, A.; Vijapura, A.; Patel, D.; Patel, M. Effect of Abiotic Stress on Crops. In Sustainable Crop Production; Chapter 1; Hasanuzzaman, M., Filho, M.C.M.T., Fujita, M., Nogueira, T.A.R., Eds.; IntechOpen: Rijeka, Croatia, 2020; ISBN 978-1-78985-318-6. [Google Scholar]
- Reddy, P.V.L.; Hernandez-Viezcas, J.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Lessons Learned: Are Engineered Nanomaterials Toxic to Terrestrial Plants? Sci. Total Environ. 2016, 568, 470–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chand Mali, S.; Raj, S.; Trivedi, R. Nanotechnology a Novel Approach to Enhance Crop Productivity. Biochem. Biophys. Rep. 2020, 24, 100821. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Singh, S.S.; Bahuguna, A.; Yadav, B.; Barthwal, A.; Nandan, R.; Khatana, R.; Pandey, A.; Thakur, R.; Singh, H.; et al. Nanotechnology: An Efficient Tool in Plant Nutrition Management. In Ecosystem Services: Types, Management and Benefits; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2022; ISBN 978-1-68507-614-6. [Google Scholar]
- Shahid, M.; Naeem-Ullah, U.; Khan, W.; Saeed, D.S.; Razzaq, K. Application of Nanotechnology for Insect Pests Management: A Review. J. Innov. Sci. 2021, 7, 28–39. [Google Scholar] [CrossRef]
- Elmer, W.; Ma, C.; White, J. Nanoparticles for Plant Disease Management. Curr. Opin. Environ. Sci. Health 2018, 6, 66–70. [Google Scholar] [CrossRef]
- Kah, M.; Kookana, R.S.; Gogos, A.; Bucheli, T.D. A Critical Evaluation of Nanopesticides and Nanofertilizers against Their Conventional Analogues. Nat. Nanotechnol. 2018, 13, 677–684. [Google Scholar] [CrossRef]
- Liu, R.; Lal, R. Synthetic Apatite Nanoparticles as a Phosphorus Fertilizer for Soybean (Glycine max). Sci. Rep. 2014, 4, 5686. [Google Scholar] [CrossRef] [Green Version]
- Delfani, M.; Baradarn Firouzabadi, M.; Farrokhi, N.; Makarian, H. Some Physiological Responses of Black-Eyed Pea to Iron and Magnesium Nanofertilizers. Commun. Soil Sci. Plant Anal. 2014, 45, 530–540. [Google Scholar] [CrossRef]
- Changmei, L.; Chaoying, Z.; Junqiang, W.; Guorong, W.; Mingxuan, T. Research of the Effect of Nanometer Materials on Germination and Growth Enhancement of Glycine Max and Its Mechanism. Soybean Sci. 2002, 21, 168–171. [Google Scholar]
- Dola, D.B.; Mannan, M.A.; Sarker, U.; Mamun, M.A.A.; Islam, T.; Ercisli, S.; Saleem, M.H.; Ali, B.; Pop, O.L.; Marc, R.A. Nano-Iron Oxide Accelerates Growth, Yield, and Quality of Glycine Max Seed in Water Deficits. Front. Plant Sci. 2022, 13, 992535. [Google Scholar] [CrossRef]
- Cañas, J.E.; Long, M.; Nations, S.; Vadan, R.; Dai, L.; Luo, M.; Ambikapathi, R.; Lee, E.H.; Olszyk, D. Effects of Functionalized and Nonfunctionalized Single-Walled Carbon Nanotubes on Root Elongation of Select Crop Species. Environ. Toxicol. Chem. 2008, 27, 1922–1931. [Google Scholar] [CrossRef] [PubMed]
- Khodakovskaya, M.V.; de Silva, K.; Biris, A.S.; Dervishi, E.; Villagarcia, H. Carbon Nanotubes Induce Growth Enhancement of Tobacco Cells. ACS Nano 2012, 6, 2128–2135. [Google Scholar] [CrossRef] [PubMed]
- Yadav, J.; Jasrotia, P.; Kashyap, P.L.; Bhardwaj, A.K.; Kumar, S.; Singh, M.; Singh, G.P. Nanopesticides: Current Status and Scope for Their Application in Agriculture. Plant Prot. Sci. 2022, 58, 1–17. [Google Scholar] [CrossRef]
- Taha, R.A. Nanotechnology and Its Application in Agriculture. Adv. Plants Agric. Res. 2016, 3, 224–232. [Google Scholar] [CrossRef]
- Gajbhiye, M.; Kesharwani, J.; Ingle, A.; Gade, A.; Rai, M. Fungus-Mediated Synthesis of Silver Nanoparticles and Their Activity against Pathogenic Fungi in Combination with Fluconazole. Nanomedicine 2009, 5, 382–386. [Google Scholar] [CrossRef]
- Sharon, M.; Choudhary, A.K.; Kumar, R. Nanotechnology in a Gricultural Diseases and food safety. J. Phytol. 2010, 2, 83–92. [Google Scholar]
- Cioffi, N.; Torsi, L.; Ditaranto, N.; Tantillo, G.; Ghibelli, L.; Sabbatini, L.; Bleve-Zacheo, T.; D’Alessio, M.; Zambonin, P.G.; Traversa, E. Copper Nanoparticle/Polymer Composites with Antifungal and Bacteriostatic Properties. Chem. Mater. 2005, 17, 5255–5262. [Google Scholar] [CrossRef]
- Esteban-Tejeda, L.; Malpartida, F.; Esteban-Cubillo, A.; Pecharromán, C.; Moya, J.S. Antibacterial and Antifungal Activity of a Soda-Lime Glass Containing Copper Nanoparticles. Nanotechnology 2009, 20, 505701. [Google Scholar] [CrossRef]
- Holzinger, M.; Le Goff, A.; Cosnier, S. Nanomaterials for Biosensing Applications: A Review. Front. Chem. 2014, 2, 63. [Google Scholar] [CrossRef] [Green Version]
- Chhipa, H. Applications of Nanotechnology in Agriculture, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; Volume 46, ISBN 9780128149928. [Google Scholar]
- Do Espirito Santo Pereira, A.; Caixeta Oliveira, H.; Fernandes Fraceto, L.; Santaella, C. Nanotechnology Potential in Seed Priming for Sustainable Agriculture. Nanomaterials 2021, 11, 267. [Google Scholar] [CrossRef]
- Malik, A.; Mor, V.S.; Tokas, J.; Punia, H.; Malik, S.; Malik, K.; Sangwan, S.; Tomar, S.; Singh, P.; Singh, N.; et al. Biostimulant-Treated Seedlings under Sustainable Agriculture: A Global Perspective Facing Climate Change. Agronomy 2021, 11, 14. [Google Scholar] [CrossRef]
- An, J.; Hu, P.; Li, F.; Wu, H.; Shen, Y.; White, J.C.; Tian, X.; Li, Z.; Giraldo, J.P. Emerging Investigator Series: Molecular Mechanisms of Plant Salinity Stress Tolerance Improvement by Seed Priming with Cerium Oxide Nanoparticles. Environ. Sci. Nano 2020, 7, 2214–2228. [Google Scholar] [CrossRef]
- Mahakham, W.; Sarmah, A.K.; Maensiri, S.; Theerakulpisut, P. Nanopriming Technology for Enhancing Germination and Starch Metabolism of Aged Rice Seeds Using Phytosynthesized Silver Nanoparticles. Sci. Rep. 2017, 7, 8263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Cota-Ruiz, K.; Hernández-Viezcas, J.A.; Valdés, C.; Medina-Velo, I.A.; Turley, R.S.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Manganese Nanoparticles Control Salinity-Modulated Molecular Responses in Capsicum Annuum L. Through Priming: A Sustainable Approach for Agriculture. ACS Sustain. Chem. Eng. 2020, 8, 1427–1436. [Google Scholar] [CrossRef]
- Abbasi Khalaki, M.; Moameri, M.; Asgari Lajayer, B.; Astatkie, T. Influence of Nano-Priming on Seed Germination and Plant Growth of Forage and Medicinal Plants. Plant Growth. Regul. 2021, 93, 13–28. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan, M.; Ali, Q.; Ali, S. Seed Priming with Silicon Nanoparticles Improved the Biomass and Yield While Reduced the Oxidative Stress and Cadmium Concentration in Wheat Grains. Environ. Sci. Pollut. Res. 2019, 26, 7579–7588. [Google Scholar] [CrossRef]
- Kasote, D.M.; Lee, J.H.J.; Jayaprakasha, G.K.; Patil, B.S. Seed Priming with Iron Oxide Nanoparticles Modulate Antioxidant Potential and Defense-Linked Hormones in Watermelon Seedlings. ACS Sustain. Chem. Eng. 2019, 7, 5142–5151. [Google Scholar] [CrossRef]
- Pirzada, T.; de Farias, B.V.; Mathew, R.; Guenther, R.H.; Byrd, M.V.; Sit, T.L.; Pal, L.; Opperman, C.H.; Khan, S.A. Recent Advances in Biodegradable Matrices for Active Ingredient Release in Crop Protection: Towards Attaining Sustainability in Agriculture. Curr. Opin. Colloid. Interface Sci. 2020, 48, 121–136. [Google Scholar] [CrossRef]
- De La Torre-Roche, R.; Cantu, J.; Tamez, C.; Zuverza-Mena, N.; Hamdi, H.; Adisa, I.O.; Elmer, W.; Gardea-Torresdey, J.; White, J.C. Seed Biofortification by Engineered Nanomaterials: A Pathway to Alleviate Malnutrition? J. Agric. Food Chem. 2020, 68, 12189–12202. [Google Scholar] [CrossRef]
- Salam, A.; Khan, A.R.; Liu, L.; Yang, S.; Azhar, W.; Ulhassan, Z.; Zeeshan, M.; Wu, J.; Fan, X.; Gan, Y. Seed Priming with Zinc Oxide Nanoparticles Downplayed Ultrastructural Damage and Improved Photosynthetic Apparatus in Maize under Cobalt Stress. J. Hazard. Mater. 2022, 423, 127021. [Google Scholar] [CrossRef]
- Manjaiah, K.M.; Mukhopadhyay, R.; Paul, R.; Datta, S.C.; Kumararaja, P.; Sarkar, B. Clay Minerals and Zeolites for Environmentally Sustainable Agriculture. In Modified Clay and Zeolite Nanocomposite Materials; Elsevier: Amsterdam, The Netherlands, 2019; pp. 309–329. [Google Scholar]
- Acharya, P.; Jayaprakasha, G.K.; Crosby, K.M.; Jifon, J.L.; Patil, B.S. Green-Synthesized Nanoparticles Enhanced Seedling Growth, Yield, and Quality of Onion (Allium cepa L.). ACS Sustain. Chem. Eng. 2019, 7, 14580–14590. [Google Scholar] [CrossRef]
- Abid, M.; Hakeem, A.; Shao, Y.; Liu, Y.; Zahoor, R.; Fan, Y.; Suyu, J.; Ata-Ul-Karim, S.T.; Tian, Z.; Jiang, D. Seed Osmopriming Invokes Stress Memory against Post-Germinative Drought Stress in Wheat (Triticum aestivum L.). Environ. Exp. Bot. 2018, 145, 12–20. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed Priming to Alleviate Salinity Stress in Germinating Seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Patil, B.D.; Manjare, M.R. Effect of Seed Priming on Germination and Bulb Yield in Onion (Allium cepa L.). Ecol. Environ. Conserv. 2013, 19, 243–246. [Google Scholar]
- Mehmood, S.; Khatoon, Z.; Amna; Ahmad, I.; Muneer, M.A.; Kamran, M.A.; Ali, J.; Ali, B.; Chaudhary, H.J.; Munis, M.F.H. Bacillus sp. PM31 Harboring Various Plant Growth-Promoting Activities Regulates Fusarium Dry Rot and Wilt Tolerance in Potato. Arch. Agron. Soil Sci. 2021, 2021, 1–15. [Google Scholar] [CrossRef]
- Badawy, A.A.; Abdelfattah, N.A.H.; Salem, S.S.; Awad, M.F.; Fouda, A. Efficacy Assessment of Biosynthesized Copper Oxide Nanoparticles (CuO-NPs) on Stored Grain Insects and Their Impacts on Morphological and Physiological Traits of Wheat (Triticum Aestivum L.) Plant. Biology 2021, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Mohanlall, V.; Odayar, K.; Odhav, B. The Role of Nanoparticles on the Plant Growth of Orthodox and Recalcitrant Seeds. Adv. Compos. Biocomposites Nanocomposites 2013, 1, 287–304. [Google Scholar]
- Karpachev, V.V.; Spiridonov, J.J.; Voropaeva, N.L.; Tkachev, A.G.; Shachnev, N.V.; Figovsky, O.L. Pre-Sowing Seed Treatment Nanotechnology with Environment-Friendly Nanotube-Based Nanochips. Int. Lett. Nat. Sci. 2016, 58, 29–34. [Google Scholar] [CrossRef]
- Aslani, F.; Bagheri, S.; Muhd Julkapli, N.; Juraimi, A.S.; Hashemi, F.S.G.; Baghdadi, A. Effects of Engineered Nanomaterials on Plants Growth: An Overview. Sci. World J. 2014, 2014, 641759. [Google Scholar] [CrossRef] [Green Version]
- Das, C.K.; Srivastava, G.; Dubey, A.; Verma, S.; Saxena, M.; Roy, M.; Sethy, N.K.; Bhargava, K.; Singh, S.K.; Sarkar, S. The Seed Stimulant Effect of Nano Iron Pyrite Is Compromised by Nano Cerium Oxide: Regulation by the Trace Ionic Species Generated in the Aqueous Suspension of Iron Pyrite. RSC Adv. 2016, 6, 67029–67038. [Google Scholar] [CrossRef]
- Srivastava, G.; Das, C.K.; Das, A.; Singh, S.K.; Roy, M.; Kim, H.; Sethy, N.; Kumar, A.; Sharma, R.K.; Singh, S.K.; et al. Seed Treatment with Iron Pyrite (FeS2) Nanoparticles Increases the Production of Spinach. RSC Adv. 2014, 4, 58495–58504. [Google Scholar] [CrossRef]
- Das, C.K.; Srivastava, G.; Dubey, A.; Roy, M.; Jain, S.; Sethy, N.K.; Saxena, M.; Harke, S.; Sarkar, S.; Misra, K.; et al. Nano-Iron Pyrite Seed Dressing: A Sustainable Intervention to Reduce Fertilizer Consumption in Vegetable (Beetroot, Carrot), Spice (Fenugreek), Fodder (Alfalfa), and Oilseed (Mustard, Sesamum) Crops. Nanotechnol. Environ. Eng. 2016, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Shaw, A.K.; Hossain, Z. Impact of Nano-CuO Stress on Rice (Oryza sativa L.) Seedlings. Chemosphere 2013, 93, 906–915. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, T.; Kundu, S.; Biswas, A.K.; Kundu, S.; Tarafdar, J.C.; Rao, A.S. Effect of Copper Oxide Nano Particle on Seed Germination of Selected Crops. J. Agric. Sci. Technol. 2012, 2, 815–823. [Google Scholar]
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.-S.P. Alleviation of the Effect of Salinity on Growth and Yield of Strawberry by Foliar Spray of Selenium-Nanoparticles. Environ. Pollut. 2019, 253, 246–258. [Google Scholar] [CrossRef]
- Hu, T.; Li, H.; Li, J.; Zhao, G.; Wu, W.; Liu, L.; Wang, Q.; Guo, Y. Absorption and Bio-Transformation of Selenium Nanoparticles by Wheat Seedlings (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babajani, A.; Iranbakhsh, A.; Oraghi Ardebili, Z.; Eslami, B. Differential Growth, Nutrition, Physiology, and Gene Expression in Melissa Officinalis Mediated by Zinc Oxide and Elemental Selenium Nanoparticles. Environ. Sci. Pollut. Res. 2019, 26, 24430–24444. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.D.; Raja, K.; Natarajan, N.; Govindaraju, K.; Subramanian, K.S. Invigouration Treatment of Metal and Metal Oxide Nanoparticles for Improving the Seed Quality of Aged Chilli Seeds (Capsicum annum L.). Mater. Chem. Phys. 2020, 242, 122492. [Google Scholar] [CrossRef]
- Itroutwar, P.D.; Govindaraju, K.; Tamilselvan, S.; Kannan, M.; Raja, K.; Subramanian, K.S. Seaweed-Based Biogenic ZnO Nanoparticles for Improving Agro-Morphological Characteristics of Rice (Oryza sativa L.). J. Plant Growth. Regul. 2020, 39, 717–728. [Google Scholar] [CrossRef]
- Savassa, S.M.; Duran, N.M.; Rodrigues, E.S.; De Almeida, E.; Van Gestel, C.A.M.; Bompadre, T.F.V.; de Carvalho, H.W.P. Effects of ZnO Nanoparticles on Phaseolus Vulgaris Germination and Seedling Development Determined by X-Ray Spectroscopy. ACS Appl. Nano Mater. 2018, 1, 6414–6426. [Google Scholar] [CrossRef]
- Dhingra, P.; Sharma, S.; Singh, K.H.; Kushwaha, H.S.; Barupal, J.K.; Haq, S.; Kothari, S.L.; Kachhwaha, S. Seed Priming with Carbon Nanotubes and Silicon Dioxide Nanoparticles Influence Agronomic Traits of Indian Mustard (Brassica Juncea) in Field Experiments. J. King Saud Univ. Sci. 2022, 34, 102067. [Google Scholar] [CrossRef]
- Sonawane, H.; Arya, S.; Math, S.; Shelke, D. Myco-Synthesized Silver and Titanium Oxide Nanoparticles as Seed Priming Agents to Promote Seed Germination and Seedling Growth of Solanum Lycopersicum: A Comparative Study. Int. Nano. Lett. 2021, 11, 371–379. [Google Scholar] [CrossRef]
- Kasote, D.M.; Lee, J.H.J.; Jayaprakasha, G.K.; Patil, B.S. Manganese Oxide Nanoparticles as Safer Seed Priming Agent to Improve Chlorophyll and Antioxidant Profiles in Watermelon Seedlings. Nanomaterials 2021, 11, 1016. [Google Scholar] [CrossRef]
- Del Buono, D.; Luzi, F.; Puglia, D. Lignin Nanoparticles: A Promising Tool to Improve Maize Physiological, Biochemical, and Chemical Traits. Nanomaterials 2021, 11, 846. [Google Scholar] [CrossRef] [PubMed]
- Saharan, V.; Kumaraswamy, R.V.; Choudhary, R.C.; Kumari, S.; Pal, A.; Raliya, R.; Biswas, P. Cu-Chitosan Nanoparticle Mediated Sustainable Approach to Enhance Seedling Growth in Maize by Mobilizing Reserved Food. J. Agric. Food. Chem. 2016, 64, 6148–6155. [Google Scholar] [CrossRef]
- Gomes, D.G.; Pelegrino, M.T.; Ferreira, A.S.; Bazzo, J.H.; Zucareli, C.; Seabra, A.B.; Oliveira, H.C. Seed Priming with Copper-loaded Chitosan Nanoparticles Promotes Early Growth and Enzymatic Antioxidant Defense of Maize (Zea Mays L.) Seedlings. J. Chem. Technol. Biotechnol. 2021, 96, 2176–2184. [Google Scholar] [CrossRef]
- Esper Neto, M.; Britt, D.W.; Jackson, K.A.; Coneglian, C.F.; Inoue, T.T.; Batista, M.A. Early Growth of Corn Seedlings after Seed Priming with Magnetite Nanoparticles Synthetised in Easy Way. Acta Agric. Scand. B Soil Plant Sci. 2021, 71, 91–97. [Google Scholar] [CrossRef]
- Do Espírito Santo Pereira, A.; Oliveira, H.C.; Fraceto, L.F. Polymeric Nanoparticles as an Alternative for Application of Gibberellic Acid in Sustainable Agriculture: A Field Study. Sci. Rep. 2019, 9, 7135. [Google Scholar] [CrossRef] [Green Version]
- Falsini, S.; Clemente, I.; Papini, A.; Tani, C.; Schiff, S.; Salvatici, M.C.; Petruccelli, R.; Benelli, C.; Giordano, C.; Gonnelli, C.; et al. When Sustainable Nanochemistry Meets Agriculture: Lignin Nanocapsules for Bioactive Compound Delivery to Plantlets. ACS Sustain. Chem. Eng. 2019, 7, 19935–19942. [Google Scholar] [CrossRef] [Green Version]
- Sundaria, N.; Singh, M.; Upreti, P.; Chauhan, R.P.; Jaiswal, J.P.; Kumar, A. Seed Priming with Iron Oxide Nanoparticles Triggers Iron Acquisition and Biofortification in Wheat (Triticum aestivum L.) Grains. J. Plant Growth. Regul. 2019, 38, 122–131. [Google Scholar] [CrossRef]
- Chau, N.H.; Doan, Q.H.; Chu, T.H.; Nguyen, T.T.; Dao Trong, H.; Ngo, Q.B. Effects of Different Nanoscale Microelement-Containing Formulations for Presowing Seed Treatment on Growth of Soybean Seedlings. J. Chem. 2019, 2019, 8060316. [Google Scholar] [CrossRef]
- Shukla, P.; Chaurasia, P.; Younis, K.; Qadri, O.S.; Faridi, S.A.; Srivastava, G. Nanotechnology in Sustainable Agriculture: Studies from Seed Priming to Post-Harvest Management. Nanotechnol. Environ. Eng. 2019, 4, 11. [Google Scholar] [CrossRef]
- Panda, D.; Mondal, S. Seed Enhancement for Sustainable Agriculture: An Overview of Recent Trends. Plant Arch. 2020, 20, 2320–2332. [Google Scholar]
- Ismail, G.; Abou-Zeid, H. The Role of Priming with Biosynthesized Silver Nanoparticles in the Response of Triticum aestivum L. to Salt Stress. Egypt. J. Bot. 2018, 58, 73–85. [Google Scholar] [CrossRef]
- Tariq, M.; Choudhary, S.; Singh, H.; Siddiqui, M.A.; Kumar, H.; Amir, A.; Kapoor, N. Role of Nanoparticles in Abiotic Stress. In Technology in Agriculture; Ahmad, F., Sultan, M., Eds.; IntechOpen: Rijeka, Croatia, 2021. [Google Scholar]
- Rai-Kalal, P.; Tomar, R.S.; Jajoo, A. Seed Nanopriming by Silicon Oxide Improves Drought Stress Alleviation Potential in Wheat Plants. Funct. Plant Biol. 2021, 48, 905–915. [Google Scholar] [CrossRef]
- Rahimi, D.; Kartoolinejad, D.; Nourmohammadi, K.; Naghdi, R. Increasing Drought Resistance of Alnus subcordata CA Mey. Seeds Using a Nano Priming Technique with Multi-Walled Carbon Nanotubes. J. For. Sci. 2016, 62, 269–278. [Google Scholar] [CrossRef]
- Yousefi, S.; Kartoolinejad, D.; Naghdi, R. Effects of Priming with Multi-Walled Carbon Nanotubes on Seed Physiological Characteristics of Hopbush (Dodon aeaviscosa L.) under Drought Stress. Int. J. Environ. Stud. 2017, 74, 528–539. [Google Scholar] [CrossRef]
- Abou-Zeid, H.M.; Ismail, G.S.M.; Abdel-Latif, S.A. Influence of Seed Priming with ZnO Nanoparticles on the Salt-Induced Damages in Wheat (Triticum aestivum L.) Plants. J. Plant. Nutr. 2021, 44, 629–643. [Google Scholar] [CrossRef]
- El-Badri, A.M.A.; Batool, M.; Mohamed, I.A.A.; Khatab, A.; Sherif, A.; Wang, Z.; Salah, A.; Nishawy, E.; Ayaad, M.; Kuai, J.; et al. Modulation of Salinity Impact on Early Seedling Stage via Nano-Priming Application of Zinc Oxide on Rapeseed (Brassica napus L.). Plant Physiol. Biochem. 2021, 166, 376–392. [Google Scholar] [CrossRef]
- Khan, M.N. Nano-Titanium Dioxide (Nano-TiO2) Mitigates NaCl Stress by Enhancing Antioxidative Enzymes and Accumulation of Compatible Solutes in Tomato (Lycopersicon Esculentum Mill.). J. Plant Sci. 2016, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Shah, T.; Latif, S.; Saeed, F.; Ali, I.; Ullah, S.; Abdullah Alsahli, A.; Jan, S.; Ahmad, P. Seed Priming with Titanium Dioxide Nanoparticles Enhances Seed Vigor, Leaf Water Status, and Antioxidant Enzyme Activities in Maize (Zea mays L.) under Salinity Stress. J. King Saud Univ. Sci. 2021, 33, 101207. [Google Scholar] [CrossRef]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed Priming for Abiotic Stress Tolerance: An Overview. Acta Physiol. Plant. 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Ivani, R.; Sanaei Nejad, S.H.; Ghahraman, B.; Astaraei, A.R.; Feizi, H. Role of Bulk and Nanosized SiO(2) to Overcome Salt Stress during Fenugreek Germination (Trigonella Foenum- Graceum L.). Plant Signal. Behav. 2018, 13, e1044190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hojjat, S.S.; Kamyab, M. The Effect of Silver Nanoparticle on Fenugreek Seed Germination under Salinity Levels. Russ. Agric. Sci. 2017, 43, 61–65. [Google Scholar] [CrossRef]
- Konate, A.; He, X.; Zhang, Z.; Ma, Y.; Zhang, P.; Alugongo, G.M.; Rui, Y. Magnetic (Fe3O4) Nanoparticles Reduce Heavy Metals Uptake and Mitigate Their Toxicity in Wheat Seedling. Sustainability 2017, 9, 790. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, U.; Luo, X.; Wang, Q.; Shu, K. Are There Unidentified Factors Involved in the Germination of Nanoprimed Seeds? Front. Plant Sci. 2020, 11, 832. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.; Kaur, S.; Dharamvir, K.; Nayyar, H.; Verma, G. Multi-walled Carbon Nanotubes Applied through Seed-priming Influence Early Germination, Root Hair, Growth and Yield of Bread Wheat (Triticum aestivum L.). J. Sci. Food Agric. 2018, 98, 3148–3160. [Google Scholar] [CrossRef] [PubMed]
- Amna; Ali, B.; Azeem, M.A.; Qayyum, A.; Mustafa, G.; Ahmad, M.A.; Javed, M.T.; Chaudhary, H.J. Bio-Fabricated Silver Nanoparticles: A Sustainable Approach for Augmentation of Plant Growth and Pathogen Control. In Sustainable Agriculture Reviews 53; Springer: Berlin/Heidelberg, Germany, 2021; pp. 345–371. [Google Scholar]
- Faryal, S.; Ullah, R.; Khan, M.N.; Ali, B.; Hafeez, A.; Jaremko, M.; Qureshi, K.A. Thiourea-Capped Nanoapatites Amplify Osmotic Stress Tolerance in Zea mays L. by Conserving Photosynthetic Pigments, Osmolytes Biosynthesis and Antioxidant Biosystems. Molecules 2022, 27, 5744. [Google Scholar] [CrossRef] [PubMed]
- Guha, T.; Ravikumar, K.V.G.; Mukherjee, A.; Mukherjee, A.; Kundu, R. Nanopriming with Zero Valent Iron (NZVI) Enhances Germination and Growth in Aromatic Rice Cultivar (Oryza sativa cv. Gobindabhog L.). Plant Physiol. Biochem. 2018, 127, 403–413. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; ur Rehman, M.Z.; Waris, A.A. Zinc and Iron Oxide Nanoparticles Improved the Plant Growth and Reduced the Oxidative Stress and Cadmium Concentration in Wheat. Chemosphere 2019, 214, 269–277. [Google Scholar] [CrossRef]
- Gupta, N.; Singh, P.M.; Sagar, V.; Pandya, A.; Chinnappa, M.; Kumar, R.; Bahadur, A. Seed Priming with ZnO and Fe3O4 Nanoparticles Alleviate the Lead Toxicity in Basella Alba L. through Reduced Lead Uptake and Regulation of ROS. Plants 2022, 11, 2227. [Google Scholar] [CrossRef] [PubMed]
- Ragab, G.; Saad-Allah, K. Seed Priming with Greenly Synthesized Sulfur Nanoparticles Enhances Antioxidative Defense Machinery and Restricts Oxidative Injury under Manganese Stress in Helianthus annuus (L.) Seedlings. J. Plant Growth Regul. 2021, 40, 1894–1902. [Google Scholar] [CrossRef]
- Basit, F.; Ulhassan, Z.; Mou, Q.; Nazir, M.M.; Hu, J.; Hu, W.; Song, W.; Sheteiwy, M.S.; Zhou, W.; Bhat, J.A.; et al. Seed Priming with Nitric Oxide and/or Spermine Mitigate the Chromium Toxicity in Rice (Oryza sativa) Seedlings by Improving the Carbon-Assimilation and Minimising the Oxidative Damages. Funct. Plant Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Zafar, S.; Perveen, S.; Kamran Khan, M.; Shaheen, M.R.; Hussain, R.; Sarwar, N.; Rashid, S.; Nafees, M.; Farid, G.; Alamri, S. Effect of Zinc Nanoparticles Seed Priming and Foliar Application on the Growth and Physio-Biochemical Indices of Spinach (Spinacia oleracea L.) under Salt Stress. PLoS ONE 2022, 17, e0263194. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.; Li, Y.; Fu, C.; Hu, J.; Chen, L.; Yan, J.; Khan, Z.; Wu, H.; Li, Z. CeO2 Nanoparticles Seed Priming Increases Salicylic Acid Level and ROS Scavenging Ability to Improve Rapeseed Salt Tolerance. Global Chall. 2022, 6, 2200025. [Google Scholar] [CrossRef]
- Khan, M.N.; Li, Y.; Khan, Z.; Chen, L.; Liu, J.; Hu, J.; Wu, H.; Li, Z. Nanoceria Seed Priming Enhanced Salt Tolerance in Rapeseed through Modulating ROS Homeostasis and α-Amylase Activities. J. Nanobiotechnol. 2021, 19, 1–19. [Google Scholar] [CrossRef]
- Van Nguyen, D.; Nguyen, H.M.; Le, N.T.; Nguyen, K.H.; Nguyen, H.T.; Le, H.M.; Nguyen, A.T.; Dinh, N.T.T.; Hoang, S.A.; van Ha, C. Copper Nanoparticle Application Enhances Plant Growth and Grain Yield in Maize Under Drought Stress Conditions. J. Plant Growth Regul. 2022, 41, 364–375. [Google Scholar] [CrossRef]
- Waqas Mazhar, M.; Ishtiaq, M.; Hussain, I.; Parveen, A.; Hayat Bhatti, K.; Azeem, M.; Thind, S.; Ajaib, M.; Maqbool, M.; Sardar, T. Seed Nano-Priming with Zinc Oxide Nanoparticles in Rice Mitigates Drought and Enhances Agronomic Profile. PLoS ONE 2022, 17, e0264967. [Google Scholar] [CrossRef]
- Waqas Mazhar, M.; Ishtiaq, M.; Maqbool, M.; Akram, R.; Shahid, A.; Shokralla, S.; Al-Ghobari, H.; Alataway, A.; Dewidar, A.Z.; El-Sabrout, A.M. Seed Priming with Iron Oxide Nanoparticles Raises Biomass Production and Agronomic Profile of Water-Stressed Flax Plants. Agronomy 2022, 12, 982. [Google Scholar] [CrossRef]
- Rahimi, S.; Hatami, M.; Ghorbanpour, M. Silicon-Nanoparticle Mediated Changes in Seed Germination and Vigor Index of Marigold (Calendula Officinalis L.) Compared to Silicate Under PEG-Induced Drought Stress. Gesunde Pflanzen 2021, 73, 575–589. [Google Scholar] [CrossRef]
- Lemmens, E.; Deleu, L.J.; De Brier, N.; De Man, W.L.; De Proft, M.; Prinsen, E.; Delcour, J.A. The Impact of Hydro-Priming and Osmo-Priming on Seedling Characteristics, Plant Hormone Concentrations, Activity of Selected Hydrolytic Enzymes, and Cell Wall and Phytate Hydrolysis in Sprouted Wheat (Triticum aestivum L.). ACS Omega 2019, 4, 22089–22100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonogaki, H. Seed Dormancy and Germination—Emerging Mechanisms and New Hypotheses. Front. Plant Sci. 2014, 5, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourioug, M.; Ezzaza, K.; Bouabid, R.; Alaoui-Mhamdi, M.; Bungau, S.; Bourgeade, P.; Alaoui-Sossé, L.; Alaoui-Sossé, B.; Aleya, L. Influence of Hydro-and Osmo-Priming on Sunflower Seeds to Break Dormancy and Improve Crop Performance under Water Stress. Environ. Sci. Pollut. Res. 2020, 27, 13215–13226. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.-M.; Rekha, K.; Rajakumar, G.; Thiruvengadam, M. Influence of Silver Nanoparticles on the Enhancement and Transcriptional Changes of Glucosinolates and Phenolic Compounds in Genetically Transformed Root Cultures of Brassica Rapa Ssp. Rapa. Bioprocess Biosyst. Eng. 2018, 41, 1665–1677. [Google Scholar] [CrossRef]
- Sosan, A.; Svistunenko, D.; Straltsova, D.; Tsiurkina, K.; Smolich, I.; Lawson, T.; Subramaniam, S.; Golovko, V.; Anderson, D.; Sokolik, A. Engineered Silver Nanoparticles Are Sensed at the Plasma Membrane and Dramatically Modify the Physiology of Arabidopsis Thaliana Plants. Plant J. 2016, 85, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Mirzajani, F.; Askari, H.; Hamzelou, S.; Schober, Y.; Römpp, A.; Ghassempour, A.; Spengler, B. Proteomics Study of Silver Nanoparticles Toxicity on Oryza sativa L. Ecotoxicol. Environ. Saf. 2014, 108, 335–339. [Google Scholar] [CrossRef]
- Khan, M.N.; Mobin, M.; Abbas, Z.K.; AlMutairi, K.A.; Siddiqui, Z.H. Role of Nanomaterials in Plants under Challenging Environments. Plant Physiol. Biochem. 2017, 110, 194–209. [Google Scholar] [CrossRef]
- Khodakovskaya, M.; Dervishi, E.; Mahmood, M.; Xu, Y.; Li, Z.; Watanabe, F.; Biris, A.S. Carbon Nanotubes Are Able to Penetrate Plant Seed Coat and Dramatically Affect Seed Germination and Plant Growth. ACS Nano 2009, 3, 3221–3227. [Google Scholar] [CrossRef]
- Saddiq, M.S.; Iqbal, S.; Afzal, I.; Ibrahim, A.M.H.; Bakhtavar, M.A.; Hafeez, M.B.; Jahanzaib; Maqbool, M.M. Mitigation of Salinity Stress in Wheat (Triticum aestivum L.) Seedlings through Physiological Seed Enhancements. J. Plant Nutr. 2019, 42, 1192–1204. [Google Scholar] [CrossRef]
- Siddaiah, C.N.; Prasanth, K.V.H.; Satyanarayana, N.R.; Mudili, V.; Gupta, V.K.; Kalagatur, N.K.; Satyavati, T.; Dai, X.-F.; Chen, J.-Y.; Mocan, A. Chitosan Nanoparticles Having Higher Degree of Acetylation Induce Resistance against Pearl Millet Downy Mildew through Nitric Oxide Generation. Sci. Rep. 2018, 8, 2485. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Singh, N.B.; Hussain, I.; Singh, H.; Yadav, V. Synthesis and Characterization of Copper Oxide Nanoparticles and Its Impact on Germination of Vigna radiata (L.) R. Wilczek. Trop. Plant Res. 2017, 4, 246–253. [Google Scholar] [CrossRef]
- Rastogi, A.; Tripathi, D.K.; Yadav, S.; Chauhan, D.K.; Živčák, M.; Ghorbanpour, M.; El-Sheery, N.I.; Brestic, M. Application of Silicon Nanoparticles in Agriculture. 3 Biotech 2019, 9, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narendhran, S.; Rajiv, P.; Sivaraj, R. Toxicity of ZnO Nanoparticles on Germinating Sesamum Indicum (Co-1) and Their Antibacterial Activity. Bull. Mater. Sci. 2016, 39, 415–421. [Google Scholar] [CrossRef] [Green Version]
- Mishra, V.; Mishra, R.K.; Dikshit, A.; Pandey, A.C. Interactions of Nanoparticles with Plants: An Emerging Prospective in the Agriculture Industry. In Emerging Technologies and Management of Crop Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2014; pp. 159–180. [Google Scholar]
- Giraldo, J.P.; Landry, M.P.; Faltermeier, S.M.; McNicholas, T.P.; Iverson, N.M.; Boghossian, A.A.; Reuel, N.F.; Hilmer, A.J.; Sen, F.; Brew, J.A. Plant Nanobionics Approach to Augment Photosynthesis and Biochemical Sensing. Nat. Mater. 2014, 13, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salama, D.M.; Osman, S.A.; Abd El-Aziz, M.E.; Abd Elwahed, M.S.A.; Shaaban, E.A. Effect of Zinc Oxide Nanoparticles on the Growth, Genomic DNA, Production and the Quality of Common Dry Bean (Phaseolus vulgaris). Biocatal. Agric. Biotechnol. 2019, 18, 101083. [Google Scholar] [CrossRef]
- Lahiani, M.H.; Dervishi, E.; Chen, J.; Nima, Z.; Gaume, A.; Biris, A.S.; Khodakovskaya, M. V Impact of Carbon Nanotube Exposure to Seeds of Valuable Crops. ACS Appl. Mater. Interfaces 2013, 5, 7965–7973. [Google Scholar] [CrossRef]
- Hayes, K.L.; Mui, J.; Song, B.; Sani, E.S.; Eisenman, S.W.; Sheffield, J.B.; Kim, B. Effects, Uptake, and Translocation of Aluminum Oxide Nanoparticles in Lettuce: A Comparison Study to Phytotoxic Aluminum Ions. Sci. Total Environ. 2020, 719, 137393. [Google Scholar] [CrossRef]
- Plaksenkova, I.; Kokina, I.; Petrova, A.; Jermaļonoka, M.; Gerbreders, V.; Krasovska, M. The Impact of Zinc Oxide Nanoparticles on Cytotoxicity, Genotoxicity, and MiRNA Expression in Barley (Hordeum vulgare L.) Seedlings. Sci. World J. 2020, 2020, 6649746. [Google Scholar] [CrossRef]
- Whitmarsh, J.; Govindjee, J.A. Concepts in Photobiology: Photosynthesis and Photo-Morphogenesis; Singhal, G.S., Renger, G.R., Sopory, S.K., Irrgang, K.-D., Eds.; Govindjee Narosa: New Delhi, India, 1997; pp. 11–51. [Google Scholar]
- Abdel Latef, A.A.H.; Zaid, A.; Abu Alhmad, M.F.; Abdelfattah, K.E. The Impact of Priming with Al2O3 Nanoparticles on Growth, Pigments, Osmolytes, and Antioxidant Enzymes of Egyptian Roselle (Hibiscus sabdariffa L.) Cultivar. Agronomy 2020, 10, 681. [Google Scholar] [CrossRef]
- Pradhan, S.; Patra, P.; Das, S.; Chandra, S.; Mitra, S.; Dey, K.K.; Akbar, S.; Palit, P.; Goswami, A. Photochemical Modulation of Biosafe Manganese Nanoparticles on Vigna Radiata: A Detailed Molecular, Biochemical, and Biophysical Study. Environ. Sci. Technol. 2013, 47, 13122–13131. [Google Scholar] [CrossRef]
- Hasanpour, H.; Maali-Amir, R.; Zeinali, H. Effect of TiO2 Nanoparticles on Metabolic Limitations to Photosynthesis under Cold in Chickpea. Russ. J. Plant Physiol. 2015, 62, 779–787. [Google Scholar] [CrossRef]
- Samadi, N.; Yahyaabadi, S.; Rezayatmand, Z. Effect of TiO2 and TiO2 Nanoparticle on Germination, Root and Shoot Length and Photosynthetic Pigments of Mentha Piperita. Int. J. Plant Soil Sci. 2014, 3, 408–418. [Google Scholar] [CrossRef]
- Yang, F.; Hong, F.; You, W.; Liu, C.; Gao, F.; Wu, C.; Yang, P. Influence of Nano-Anatase TiO2 on the Nitrogen Metabolism of Growing Spinach. Biol. Trace Elem. Res. 2006, 110, 179–190. [Google Scholar] [CrossRef]
- Grillet, L.; Mari, S.; Schmidt, W. Iron in Seeds–Loading Pathways and Subcellular Localization. Front. Plant Sci. 2014, 4, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, M.; Ma, C.; Hao, Y.; Guo, J.; Rui, Y.; Tang, X.; Zhao, Q.; Fan, X.; Zhang, Z.; Hou, T. Iron Oxide Nanoparticles as a Potential Iron Fertilizer for Peanut (Arachis hypogaea). Front. Plant Sci. 2016, 7, 815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guha, T.; Das, H.; Mukherjee, A.; Kundu, R. Elucidating ROS Signaling Networks and Physiological Changes Involved in Nanoscale Zero Valent Iron Primed Rice Seed Germination Sensu Stricto. Free Radic Biol. Med. 2021, 171, 11–25. [Google Scholar] [CrossRef]
- Astafurova, T.P.; Morgalev, Y.N.; Zotikova, A.P.; Verkhoturova, G.S.; Mikhajlova, S.I.; Burenina, A.A.; Zajtseva, T.A.; Postovalova, V.M.; Tsytsareva, L.K.; Borovikova, G.V. Effect of Nanoparticles of Titanium Dioxide and Aluminum Oxide on Some Morphophysiological Characteristics of Plants. Bull. Tomsk. State Univ. Biol. 2011, 2011, 113–122. [Google Scholar]
- Cao, Z.; Rossi, L.; Stowers, C.; Zhang, W.; Lombardini, L.; Ma, X. The Impact of Cerium Oxide Nanoparticles on the Physiology of Soybean (Glycine max (L.) Merr.) under Different Soil Moisture Conditions. Environ. Sci. Pollut. Res. 2018, 25, 930–939. [Google Scholar] [CrossRef]
- Majumdar, S.; Peralta-Videa, J.R.; Trujillo-Reyes, J.; Sun, Y.; Barrios, A.C.; Niu, G.; Flores-Margez, J.P.; Gardea-Torresdey, J.L. Soil Organic Matter Influences Cerium Translocation and Physiological Processes in Kidney Bean Plants Exposed to Cerium Oxide Nanoparticles. Sci. Total Environ. 2016, 569, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Das, P.; Chetia, B.; Prasanth, R.; Madhavan, J.; Singaravelu, G.; Benelli, G.; Murugan, K. Green Nanosynthesis and Functionalization of Gold Nanoparticles as PTP 1B Inhibitors. J. Clust. Sci. 2017, 28, 2269–2277. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Abu Alhmad, M.F.; Abdelfattah, K.E. The Possible Roles of Priming with ZnO Nanoparticles in Mitigation of Salinity Stress in Lupine (Lupinus Termis) Plants. J. Plant Growth Regul. 2017, 36, 60–70. [Google Scholar] [CrossRef]
- Maswada, H.F.; Djanaguiraman, M.; Prasad, P.V. V Seed Treatment with Nano-iron (III) Oxide Enhances Germination, Seeding Growth and Salinity Tolerance of Sorghum. J. Agron. Crop Sci. 2018, 204, 577–587. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Faisal, M.; Al Sahli, A.A. Nano-silicon Dioxide Mitigates the Adverse Effects of Salt Stress on Cucurbita pepo L. Environ. Toxicol. Chem. 2014, 33, 2429–2437. [Google Scholar] [CrossRef]
- Shang, Y.; Hasan, M.K.; Ahammed, G.J.; Li, M.; Yin, H.; Zhou, J. Applications of Nanotechnology in Plant Growth and Crop Protection: A Review. Molecules 2019, 24, 2558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalia, A.; Kaur, H. Nanofertilizers: An Innovation towards New Generation Fertilizers for Improved Nutrient-Use Efficacy and Environmental Sustainability. In NanoAgroceuticals & NanoPhytoChemicals; CRC Press: Boca Raton, FL, USA, 2018; pp. 45–61. ISBN 1351139282. [Google Scholar]
- Marslin, G.; Sheeba, C.J.; Franklin, G. Nanoparticles Alter Secondary Metabolism in Plants via ROS Burst. Front. Plant Sci. 2017, 8, 832. [Google Scholar] [CrossRef] [Green Version]
- Yatim, N.M.; Shaaban, A.; Dimin, M.F.; Yusof, F.; Abd Razak, J. Effect of Functionalised and Non-Functionalised Carbon Nanotubes-Urea Fertilizer on the Growth of Paddy. Trop. Life Sci. Res. 2018, 29, 17. [Google Scholar] [CrossRef]
- Ghidan, A.Y.; Al-Antary, T.M.; Awwad, A.M.; Ayad, J.Y. Physiological Effect of Some Nanomaterials on Pepper (Capsicum Annuum L.) Plants. Fresenius Environ. Bull. 2018, 27, 7872–7878. [Google Scholar]
- Aubert, T.; Burel, A.; Esnault, M.-A.; Cordier, S.; Grasset, F.; Cabello-Hurtado, F. Root Uptake and Phytotoxicity of Nanosized Molybdenum Octahedral Clusters. J. Hazard. Mater. 2012, 219, 111–118. [Google Scholar] [CrossRef]
- Acharya, P.; Jayaprakasha, G.K.; Crosby, K.M.; Jifon, J.L.; Patil, B.S. Nanoparticle-Mediated Seed Priming Improves Germination, Growth, Yield, and Quality of Watermelons (Citrullus lanatus) at Multi-Locations in Texas. Sci. Rep. 2020, 10, 5037. [Google Scholar] [CrossRef] [Green Version]
- Laware, S.L.; Raskar, S. Effect of Titanium Dioxide Nanoparticles on Hydrolytic and Antioxidant Enzymes during Seed Germination in Onion. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 749–760. [Google Scholar]
- Salachna, P.; Byczyńska, A.; Zawadzińska, A.; Piechocki, R.; Mizielińska, M. Stimulatory Effect of Silver Nanoparticles on the Growth and Flowering of Potted Oriental Lilies. Agronomy 2019, 9, 610. [Google Scholar] [CrossRef] [Green Version]
- Shivay, Y.S.; Singh, U.; Prasad, R.; Kaur, R. Agronomic Interventions for Micronutrient Biofortification of Pulses. Indian J. Agron. 2016, 61, S161–S172. [Google Scholar]
- Umair, A.; Ali, S.; Hayat, R.; Ansar, M.; Tareen, M.J. Evaluation of Seed Priming in Mung Bean (Vigna radiata) for Yield, Nodulation and Biological Nitrogen Fixation under Rainfed Conditions. Afr. J. Biotechnol. 2011, 10, 18122–18129. [Google Scholar]
- Cakmak, I. The Role of Potassium in Alleviating Detrimental Effects of Abiotic Stresses in Plants. J. Plant. Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Arif, M.; Waqas, M.; Nawab, K.; Shahid, M. Effect of Seed Priming in Zn Solutions on Chickpea and Wheat. In Proceedings of the 8th African Crop Science Conference, El-Minia, Egypt, 27–31 October 2007; Volume 8, pp. 237–240. [Google Scholar]
- Harris, D.; Rashid, A.; Miraj, G.; Arif, M.; Yunas, M. ‘On-Farm’Seed Priming with Zinc in Chickpea and Wheat in Pakistan. Plant Soil 2008, 306, 3–10. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Guo, K.; Elbaz, A.A.; Yang, Z.M. Salicylic Acid Alleviates Mercury Toxicity by Preventing Oxidative Stress in Roots of Medicago Sativa. Environ. Exp. Bot. 2009, 65, 27–34. [Google Scholar] [CrossRef]
- Osman, M.S.; Badawy, A.A.; Osman, A.I.; Abdel Latef, A.A.H. Ameliorative Impact of an Extract of the Halophyte Arthrocnemum Macrostachyum on Growth and Biochemical Parameters of Soybean Under Salinity Stress. J. Plant Growth Regul. 2021, 40, 1245–1256. [Google Scholar] [CrossRef]
- Tavili, A.; Zare, S.; Enayati, A. Hydropriming, Ascorbic and Salicylic Acid Influence on Germination of Agropyron Elongatum Host. Seeds under Salt Stress. Res. J. Seed Sci. 2009, 2, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Shinde, S.; Paralikar, P.; Ingle, A.P.; Rai, M. Promotion of Seed Germination and Seedling Growth of Zea Mays by Magnesium Hydroxide Nanoparticles Synthesized by the Filtrate from Aspergillus Niger. Arab. J. Chem. 2020, 13, 3172–3182. [Google Scholar] [CrossRef]
- Yugandhar, P.; Savithramma, N. Green Synthesis of Calcium Carbonate Nanoparticles and Their Effects on Seed Germination and Seedling Growth of Vigna mungo (L.) Hepper. Int. J. Adv. Res. 2013, 1, 89–103. [Google Scholar]
- Nadeem, F.; Farooq, M.; Ullah, A.; Rehman, A.; Nawaz, A.; Naveed, M. Influence of Zn Nutrition on the Productivity, Grain Quality and Grain Biofortification of Wheat under Conventional and Conservation Rice–Wheat Cropping Systems. Arch. Agron. Soil Sci. 2020, 66, 1042–1057. [Google Scholar] [CrossRef]
- Das, C.K.; Jangir, H.; Kumar, J.; Verma, S.; Mahapatra, S.S.; Philip, D.; Srivastava, G.; Das, M. Nano-Pyrite Seed Dressing: A Sustainable Design for NPK Equivalent Rice Production. Nanotechnol. Environ. Eng. 2018, 3, 14. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Gao, B.; Zhang, M. Stimulation of Peanut Seedling Development and Growth by Zero-Valent Iron Nanoparticles at Low Concentrations. PLoS ONE 2015, 10, e0122884. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Kang, Y.-G.; Chang, Y.-S.; Kim, J.-H. Effects of Zerovalent Iron Nanoparticles on Photosynthesis and Biochemical Adaptation of Soil-Grown Arabidopsis Thaliana. Nanomaterials 2019, 9, 1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attia, M.S.; Hashem, A.H.; Badawy, A.A.; Abdelaziz, A.M. Biocontrol of Early Blight Disease of Eggplant Using Endophytic Aspergillus Terreus: Improving Plant Immunological, Physiological and Antifungal Activities. Bot. Stud. 2022, 63, 26. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive Oxygen Species, Abiotic Stress and Stress Combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms of Oxidative Stress in Plants: From Classical Chemistry to Cell Biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S. Interactive Effect of Temperature and Water Stress on Physiological and Biochemical Processes in Soybean. Physiol. Mol. Biol. Plants 2019, 25, 667–681. [Google Scholar] [CrossRef]
- Hatami, M.; Khanizadeh, P.; Bovand, F.; Aghaee, A. Silicon Nanoparticle-Mediated Seed Priming and Pseudomonas spp. Inoculation Augment Growth, Physiology and Antioxidant Metabolic Status in Melissa Officinalis L. Plants. Ind. Crops Prod. 2021, 162, 113238. [Google Scholar] [CrossRef]
- Dos Santos, J.O.; de Fária, M.E.; da Silva, D.M.; de Oliveira Silveira, H.R.; Campos, C.N.; Alves, J.D. Copper (Cu) Stress Affects Carbon and Antioxidant Metabolism in’Coffea Arabica’seedlings. Aust. J. Crop Sci. 2017, 11, 960–967. [Google Scholar] [CrossRef]
- Cui, J.D.; Qiu, J.Q.; Fan, X.W.; Jia, S.R.; Tan, Z.L. Biotechnological Production and Applications of Microbial Phenylalanine Ammonia Lyase: A Recent Review. Crit. Rev. Biotechnol. 2014, 34, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Wang, X.; Saleem, M.H.; Hafeez, A.; Afridi, M.S.; Khan, S.; Ullah, I.; do Amaral Júnior, A.T.; Alatawi, A.; Ali, S. PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes. Plants 2022, 11, 345. [Google Scholar] [CrossRef]
- Afridi, M.S.; Amna; Sumaira; Mahmood, T.; Salam, A.; Mukhtar, T.; Mehmood, S.; Ali, J.; Khatoon, Z.; Bibi, M.; et al. Induction of Tolerance to Salinity in Wheat Genotypes by Plant Growth Promoting Endophytes: Involvement of ACC Deaminase and Antioxidant Enzymes. Plant Physiol. Biochem. 2019, 139, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Riachi, L.G.; De Maria, C.A.B. Peppermint Antioxidants Revisited. Food Chem. 2015, 176, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Heo, S.-I.; Wang, M.-H. Antioxidant and Antidiabetic Activities of Extracts from Cirsium Japonicum Roots. Nutr. Res. Pract. 2008, 2, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamal Kumar, V.; Muthukrishnan, S.; Rajalakshmi, R. Phytostimulatory Effect of Phytochemical Fabricated Nanosilver (AgNPs) on Psophocarpus tetragonolobus (L.) DC. Seed Germination: An Insight from Antioxidative Enzyme Activities and Genetic Similarity Studies. Curr. Plant. Biol. 2020, 23, 100158. [Google Scholar] [CrossRef]
- Afridi, M.S.; van Hamme, J.; Bundschuh, J.; Sumaira; Khan, M.N.; Salam, A.; Waqar, M.; Munis, M.F.H.; Chaudhary, H.J. Biotechnological Approaches in Agriculture and Environmental Management-Bacterium Kocuria rhizophila 14ASP as Heavy Metal and Salt- Tolerant Plant Growth- Promoting Strain. Biologia 2021, 76, 3091–3105. [Google Scholar] [CrossRef]
- Mukhtar, T.; Afridi, M.S.; McArthur, R.; van Hamme, J.D.; Rineau, F.; Mahmood, T.; Amna; Sumaira; Zahid, M.; Salam, A.; et al. Correction: Draft Genome Sequence of Bacillus Safensis SCAL1, an Endophytic Heat-Tolerant Plant Growth-Promoting Bacterium”. Microbiol. Resour. Announc. 2019, 8, e00306-18. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant-Microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A Review on the Plant Microbiome: Ecology, Functions, and Emerging Trends in Microbial Application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Solanki, M.K.; Solanki, A.C.; Rai, S.; Srivastava, S.; Kashyap, B.K.; Divvela, P.K.; Kumar, S.; Yandigeri, M.; Kashyap, P.L.; Srivastava, A.K.; et al. Functional Interplay between Antagonistic Bacteria and Rhizoctonia Solani in the Tomato Plant Rhizosphere. Front. Microbiol. 2022, 13, 990850. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Hafeez, A.; Ahmad, S.; Javed, M.A.; Afridi, M.S.; Dawoud, T.M.; Almaary, K.S.; Muresan, C.C.; Marc, R.A.; Alkhalifah, D.H.M. Bacillus Thuringiensis PM25 Ameliorates Oxidative Damage of Salinity Stress in Maize via Regulating Growth, Leaf Pigments, Antioxidant Defense System, and Stress Responsive Gene Expression. Front. Plant Sci. 2022, 13, 921668. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, H.; Mazher, J.; Azmat, A.; Nosheen, A.; Naz, R.; Hassan, M.N.; Noureldeen, A.; Ahmad, P. Combined Application of Zinc Oxide Nanoparticles and Biofertilizer to Induce Salt Resistance in Safflower by Regulating Ion Homeostasis and Antioxidant Defence Responses. Ecotoxicol. Environ. Saf. 2021, 218, 112262. [Google Scholar] [CrossRef]
- Akhtar, N.; Ilyas, N.; Mashwani, Z.-U.-R.; Hayat, R.; Yasmin, H.; Noureldeen, A.; Ahmad, P. Synergistic Effects of Plant Growth Promoting Rhizobacteria and Silicon Dioxide Nano-Particles for Amelioration of Drought Stress in Wheat. Plant Physiol. Biochem. 2021, 166, 160–176. [Google Scholar] [CrossRef]
- Galal, A. Exogenous Application of Zinc Mitigates the Deleterious Effects in Eggplant Grown under Salinity Stress. J. Plant Nutr. 2019, 42, 915–927. [Google Scholar] [CrossRef]
- Raliya, R.; Tarafdar, J.C.; Biswas, P. Enhancing the Mobilization of Native Phosphorus in the Mung Bean Rhizosphere Using ZnO Nanoparticles Synthesized by Soil Fungi. J. Agric. Food Chem. 2016, 64, 3111–3118. [Google Scholar] [CrossRef]
- Dai, Y.; Chen, F.; Yue, L.; Li, T.; Jiang, Z.; Xu, Z.; Wang, Z.; Xing, B. Uptake, Transport, and Transformation of CeO 2 Nanoparticles by Strawberry and Their Impact on the Rhizosphere Bacterial Community. ACS Sustain. Chem. Eng. 2020, 8, 4792–4800. [Google Scholar] [CrossRef]
- Zeeshan, M.; Hu, Y.X.; Iqbal, A.; Salam, A.; Liu, Y.X.; Muhammad, I.; Ahmad, S.; Khan, A.H.; Hale, B.; Wu, H.Y.; et al. Amelioration of AsV Toxicity by Concurrent Application of ZnO-NPs and Se-NPs Is Associated with Differential Regulation of Photosynthetic Indexes, Antioxidant Pool and Osmolytes Content in Soybean Seedling. Ecotoxicol. Environ. Saf. 2021, 225, 112738. [Google Scholar] [CrossRef]
- Yang, S.; Ulhassan, Z.; Shah, A.M.; Khan, A.R.; Azhar, W.; Hamid, Y.; Hussain, S.; Sheteiwy, M.S.; Salam, A.; Zhou, W. Salicylic Acid Underpins Silicon in Ameliorating Chromium Toxicity in Rice by Modulating Antioxidant Defense, Ion Homeostasis and Cellular Ultrastructure. Plant Physiol. Biochem. 2021, 166, 1001–1013. [Google Scholar] [CrossRef]
- Gul, A.; Salam, A.; Afridi, M.S.; Bangash, N.K.; Ali, F.; Ali, M.Y.; Khan, S.; Mubeeen, R. Effect of Urea, Bio-Fertilizers and Their Interaction on the Growth, Yield and Yield Attributes of Cyamopsis Tetragonoloba. Indian J. Agric. Res. 2019, 53, 423–428. [Google Scholar] [CrossRef]
- Ulhassan, Z.; Khan, A.R.; Sheteiwy, M.S.; Azhar, W.; Hamid, Y.; Hussain, S.; Salam, A.; Kamran, M.A.; Hakeem, K.R.; Aftab, T.; et al. Selenium-Mediated Regulation of Antioxidant Defense System and Improved Heavy Metals Tolerance in Plants. In Antioxidant Defense in Plants; Springer Nature: Singapore, 2022; pp. 369–382. [Google Scholar]
- Ulhassan, Z.; Khan, A.R.; Hamid, Y.; Azhar, W.; Hussain, S.; Sheteiwy, M.S.; Salam, A.; Hakeem, K.R.; Zhou, W. Interaction of Nanoparticles with Soil–Plant System and Their Usage in Remediation Strategies. In Metals Metalloids Soil Plant Water Systems; Elsevier: Amsterdam, The Netherlands, 2022; pp. 287–308. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| S No. | Priming Agent | Concentration/Size | Priming Duration | Target Plant | Effect on Plant Growth and Development | Ref. |
|---|---|---|---|---|---|---|
| 1 | TiO2 NPs, ZnO NPs, and Ag NPs | 750, 1000, 1250 mg kg−1/35–40 nm 750, 1000, 1250 mg kg−1/100 nm 750, 1000, 1250 mg kg−1/85 nm | 3 min/5 times | Capsicum annum L. | Increased seed germination rate, seedling vigor index, root, shoot length, and CAT activity in aged chili seeds. | [65] |
| 2 | ZnO NPs | 5, 10, 25, 50, 100 and 200 mg L−1/15 nm | 12 h | Oryza sativa L. | Enhanced plant Zn contents, improved seed germination, overall growth, and agronomical characteristics. | [66] |
| 3 | ZnO NPs | 1, 10, 100, 1000, 5000 mg L−1/20, 40, 60 nm | 20 min | Phaseolus vulgaris L. | None of the concentrations and sizes harmed the seeds. NPs mostly detected in the seed coat, while 10 mg L−1 entered the inner seed and increased plant weight. | [67] |
| 4 | SiO2 NPs and MWCNTs | 25, 50, 75, 100, 125 μg mL−1/80.75 nm, 54.64 nm | 6 h | Brassica juncea L. | Increased agronomic traits, silique length, and yield per silique. | [68] |
| 5 | Ag NPs and TiO2 NPs | 25, 50, 100, 200, 400 μg mL−1/50–100 nm | 24 h | Solanum lycopersicum L. | Ag NPs at 25 and 50 μg mL−1 increased seed germination and seedling vigor index, whereas TiO2 NPs reduced seed germination and seedling vigor and increased antioxidant activity. | [69] |
| 6 | MnO NPs | 10, 20, 40, 80 mg L−1/22–39 nm | 14 h | Citrullus lanatus L. | Improved chlorophyll contents, phenolic acids, and phytohormones. | [70] |
| 7 | Lignin NPs | 80, 312, 1250, 5000, 20,000 mg L−1/50 nm | 8 h | Zea mays L. | Induced beneficial effects on root and shoot growth, increased chlorophyll, carotenoids, and anthocyanin contents. | [71] |
| 8 | Cu-chitosan NPs | 0.01, 0.04, 0.08, 0.012, and 0.16% (w/v)/150 ± 12.4 nm | 4 h | Zea mays L. | Increased α-amylase and protease enzymes also increased the total protein content in germinating seeds. | [72] |
| 9 | Cu-chitosan NPs | 0.0625 mmol L−1/174.2 ± 1.5 nm | N/A | Zea mays L. | Increased root shoot length, dry mass, leaf area, and gas exchange attributes. | [73] |
| 10 | Fe3O4 NPs | 0, 20, 40, 80, 160 mg L−1/30 nm | 8 h | Zea mays L. | Increased seed germination and vigor, plant length, and biomass. | [74] |
| 11 | Fe2O3 NPs | 50, 100, 150 mg L−1/23 nm | 14 h | Citrullus lanatus T. | Increased root and shoot length as well as non-enzymatic antioxidants. | [43] |
| 12 | nanoCS/TPP-GA3, nanoALG/CS-GA3 | 0.005 mg mL−1 and 0.0005 mg mL−1/195, 450 nm | 12 h | Solanum lycopersicum L. | Increased fruit production and overall productivity up to 4 fold. | [75] |
| 13 | Lignin NCs with GA3 | 0.5, 1, and 1.5 mg mL−1/200–250 nm | 30 Min | Eruca vesicaria and Solanum lycopersicum L. | Improved germination percentage, increased stem and root length as well as their fresh and dry weight both under in-vitro and in-vivo conditions. | [76] |
| 14 | Fe2O3 NPs | 0 ppm to 600 ppm/80 nm | 12 h | Triticum aestivum L. | Increased grain iron content, germination percentage, seed vigor index I, II and shoot length. | [77] |
| 15 | Co and MoO3 NPs | 1 L for 40 kg seeds, 0.5 mL/20 g seed/60–80 nm | 2 h | Glycin max L. | Improved seed vigor indices and plant growth as well as biomass | [78] |
| S No. | Stress Factor | Priming Agent | Concentration/Size | Priming Duration | Target Plant | Stress Mitigation Mechanism | Ref. |
|---|---|---|---|---|---|---|---|
| 1 | Cobalt | ZnO NPs | 500 mg L−1/20 nm | 24 h | Zea mays L. | Increased seed zinc content, decreased oxidative damage, improved antioxidant activity, photosynthetic apparatus, and ultrastructure. | [46] |
| 2 | Cadmium | ZnO NPs and Fe NPs | 25, 50, 75, 100 mg L−1/20–30 nm, 5, 10, 15, 20 mg L−1/50–100 nm | 20 h | Triticum aestivum L. | Increased Zn and Fe content, respectively. The NPs increased plant growth and nutrient contents while decreasing Cd uptake and, consequently, Cd toxicity. | [99] |
| 3 | Lead | ZnO and Fe3O4 NPs | 0, 50, 100, 200, 300 and 500 mg L−1/70 nm, 55 nm | 16 h | Basella alba L. | At 200 mg L−1, both NPs decreased lead uptake and reduced toxicity by reducing H2O2 and MDA content and increasing SOD, POD, CAT, and proline activity. | [100] |
| 4 | Manganese | SNPs | 12.5, 25, 50, 100, 200 µM/23 nm | 18 h | Helianthus annuus L. | Improved antioxidants and phenolic compounds while reducing oxidative damage and lipid peroxidation. | [101] |
| 5 | Chromium | Nitric oxide NPs | 100 μM/N/A | 24 h | Oryza sativa L. | Together with spermidine, nitric oxide reduced chromium accumulation in plants leading to reduce oxidative stress and increased carbon assimilation | [102] |
| 6 | Salt | CeO2 NPs | 500 mg L−1/1.8 nm | 24 h | Gossypium hirsutum L. | NPs localized in cotyledon and root apical meristem. Enhanced nutrient uptake, reduced ROS accumulation, and up-regulated terpene synthase genes. | [38] |
| 7 | Salt | TiO2 NPs | 40, 60, 80 ppm/25 nm | 24 h | Zea mays L. | Enhanced phenylalanine ammonia lyase, potassium ions, and antioxidants while reducing sodium ions, MDA, and electrolyte leakage. | [89] |
| 8 | Salt | ZnO NPs | 50, 100 and 500 mg L−1/12–21.0 nm | 24 h | Triticum aestivum L. | Changed polypeptide patterns which resulted in improved plant growth, photosynthetic pigments, overall photosynthetic efficacy, and leaf ultrastructure. | [86] |
| 9 | Salt | ZnO NPs | 0.1%, 0.2%, 0.3%/N/A | 12 h | Spinacia oleracea L. | Reversed salt stress-induced reduction in biochemical and growth attributes, improved antioxidant defense system. | [103] |
| 10 | Salt | CeO2 NPs | 100 µMol/4.1 nm | 8 h | Brassica napus L. | Improved ROS scavenging ability and up-regulated salicylic acid-related genes. | [104] |
| 11 | Salt | CeO2 NPs | 100 µMol/8.5 nm | 1, 3, 8 h | Brassica napus L. | Increased seed waster and NPs uptake capacity, increased α-amylase activity of seed, and consequently improved plant nutrient content. | [105] |
| 12 | Drought | zero-valent Cu NPs | 5.556 mg L−1/30–40 nm | 8 h | Zea mays L. | It decreased drought-induced damage by enhancing ROS-scavenging enzymatic activity, and light-harvesting pigments. | [106] |
| 13 | Drought | ZnO NPs | 5, 10, 15, 25, 50 ppm/20–30 nm | 24 h | Oryza sativa L. | Increased plant growth attributes, biomass, proline, SOD, POD, and CAT levels while reducing MDA content. | [107] |
| 14 | Drought | Fe2O3 NPs | 0, 25, 50, 75, and 100 ppm/N/A | 24 h | Linum usitatissimum L. | Increased plant growth, biomass, and yield attributes improved antioxidant enzymatic activity while decreasing oxidative damage. | [108] |
| 15 | Drought | Si NPs | 0, 100, 200, 500 mg L−1/25 nm | N/A | Calendula Officinalis L. | Si NPs at higher levels deposited in root cell walls after priming which resulted in enhanced germination indices and subsequent growth attributes under drought stress. | [109] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salam, A.; Afridi, M.S.; Javed, M.A.; Saleem, A.; Hafeez, A.; Khan, A.R.; Zeeshan, M.; Ali, B.; Azhar, W.; Sumaira; et al. Nano-Priming against Abiotic Stress: A Way Forward towards Sustainable Agriculture. Sustainability 2022, 14, 14880. https://doi.org/10.3390/su142214880
Salam A, Afridi MS, Javed MA, Saleem A, Hafeez A, Khan AR, Zeeshan M, Ali B, Azhar W, Sumaira, et al. Nano-Priming against Abiotic Stress: A Way Forward towards Sustainable Agriculture. Sustainability. 2022; 14(22):14880. https://doi.org/10.3390/su142214880
Chicago/Turabian StyleSalam, Abdul, Muhammad Siddique Afridi, Muhammad Ammar Javed, Aroona Saleem, Aqsa Hafeez, Ali Raza Khan, Muhammad Zeeshan, Baber Ali, Wardah Azhar, Sumaira, and et al. 2022. "Nano-Priming against Abiotic Stress: A Way Forward towards Sustainable Agriculture" Sustainability 14, no. 22: 14880. https://doi.org/10.3390/su142214880