
Beta-Thalassemia Minor and SARS-CoV-2: Physiopathology, Prevalence, Severity, Morbidity, and Mortality

Abstract
:1. Introduction
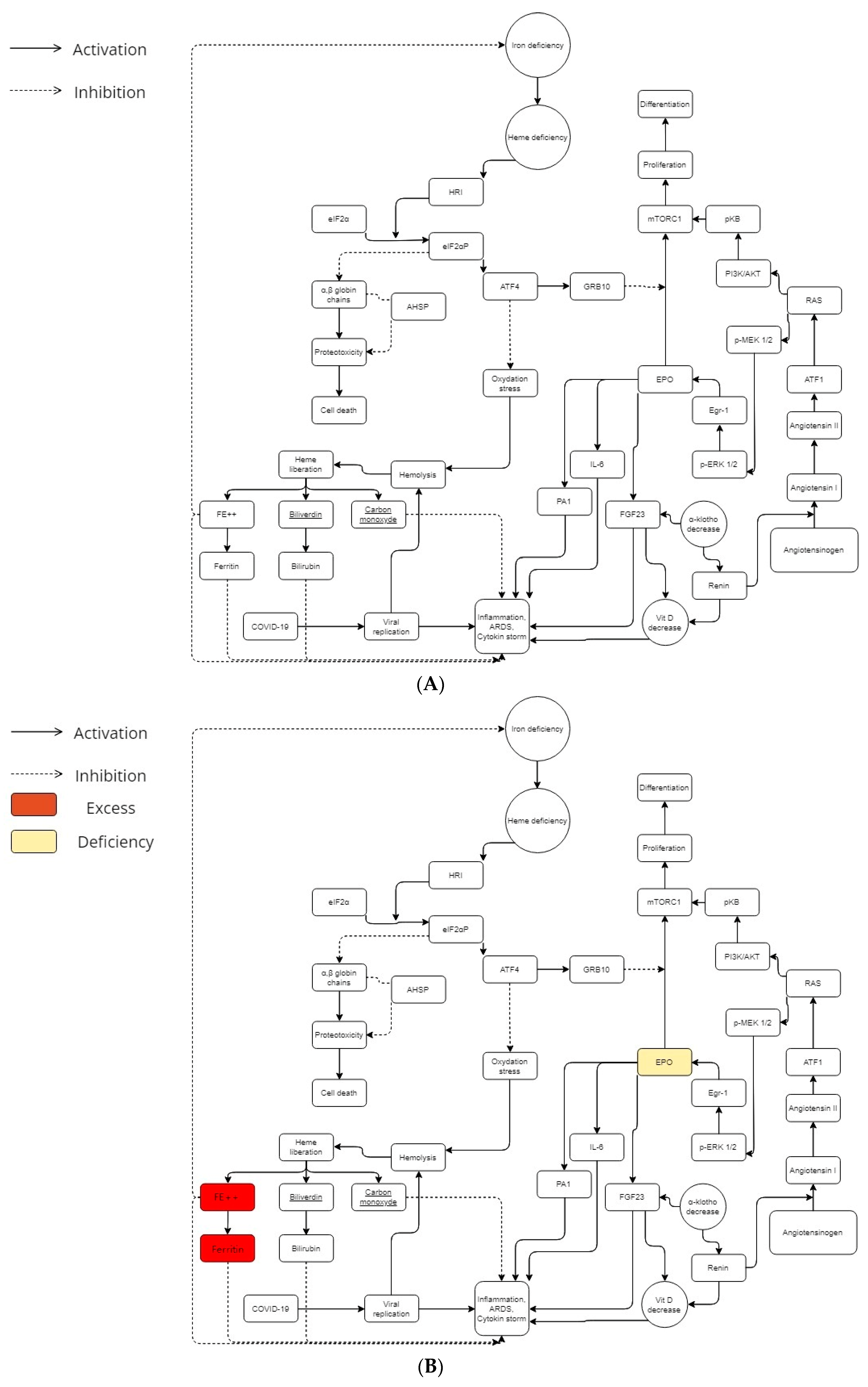
2. Physiopathology
2.1. Beta-Thalassemia Minor
2.2. Iron, Heme, and Redox
2.3. Erythropoietin and ACE Way
2.4. BCL11A Gene Possible Imputation
3. Review
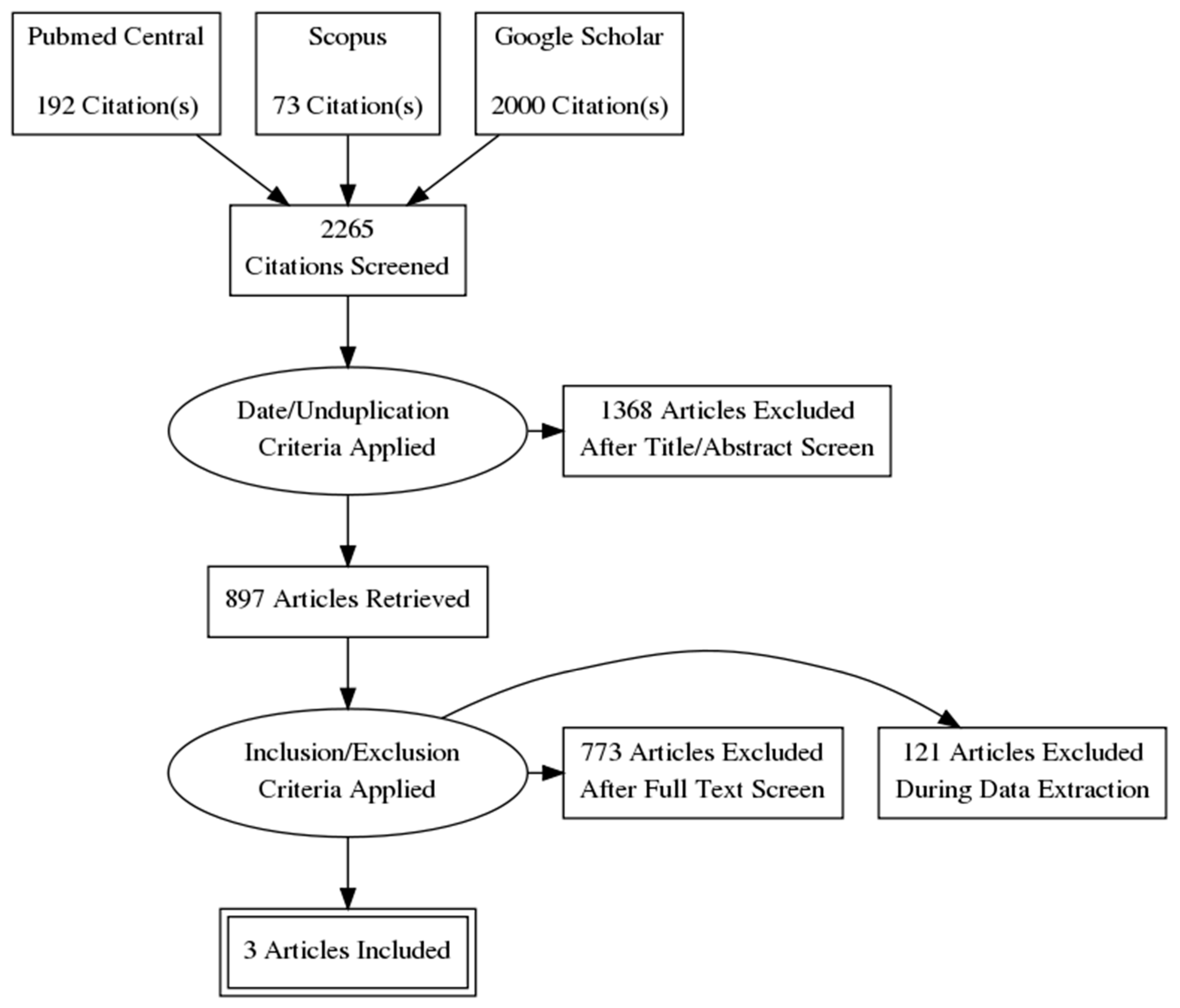
3.1. Methods
3.1.1. Risk of Bias (Quality) Assessment
3.1.2. Strategy for Data Synthesis
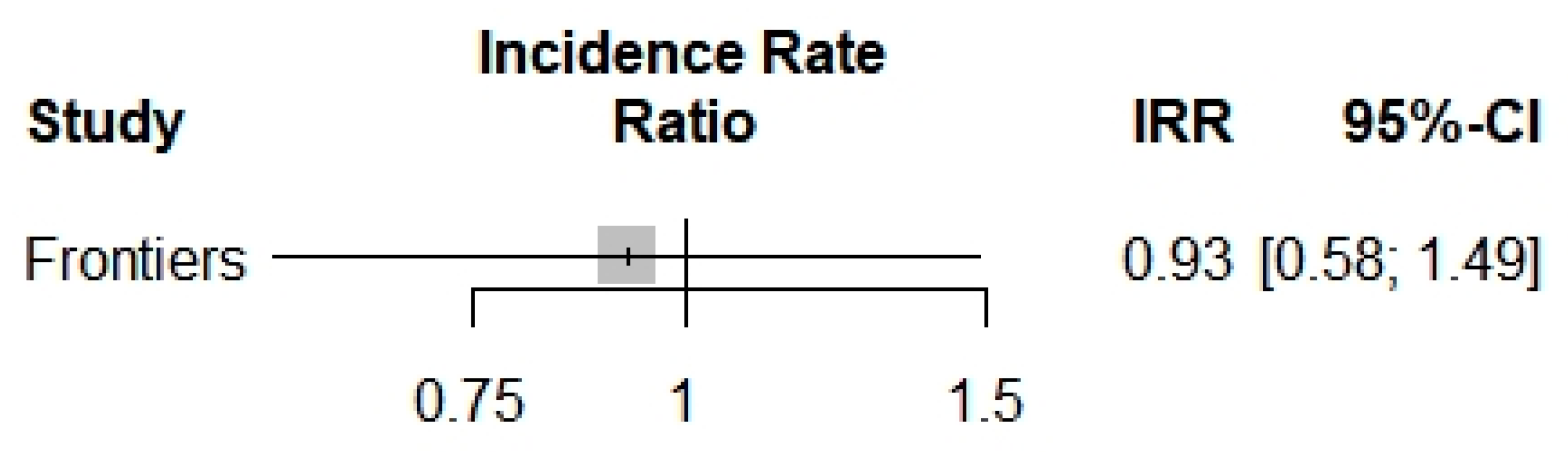
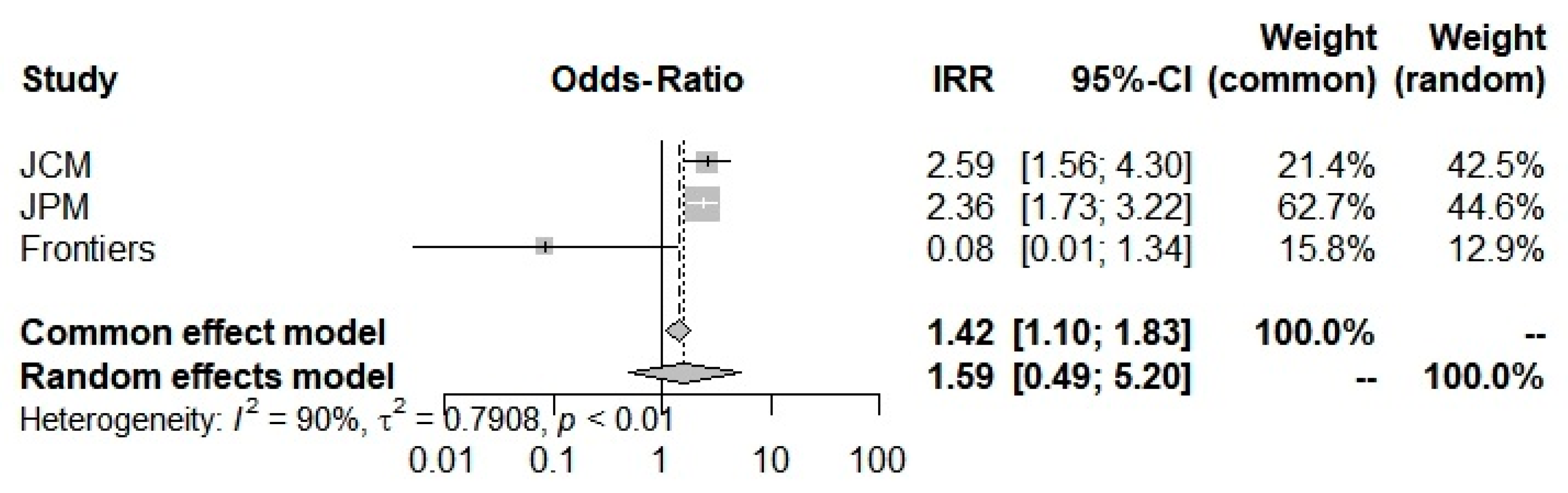
3.2. Results
4. Discussion
Funding
Conflicts of Interest
References
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.; Lau, E.H.; Wong, J.Y.; et al. Early transmission dynamics in Wuhan, China, of novel coronavirus–infected pneumonia. N. Engl. J. Med. 2020, 382, 29. [Google Scholar] [CrossRef] [PubMed]
- Lansiaux, E.; Pébaÿ, P.P.; Picard, J.-L.; Son-Forget, J. COVID-19: Beta-thalassemia subjects immunised? Obs. Study 2020, 142, 109827. [Google Scholar] [CrossRef] [PubMed]
- Drouin, E. Beta-thalassemia may protect against COVID 19. Med. Hypotheses 2020, 143, 110014. [Google Scholar] [CrossRef]
- Torti, L.; Maffei, L.; Sorrentino, F.; De Fabritiis, P.; Miceli, R.; Abruzzese, E. Impact of SARS CoV-2 in Hemoglobinopathies with Immune Disfunction and Epidemiology. A Protective Mechanism from Beta Chain Hemoglobin Defects? Mediterr. J. Hematol. Infect. Dis. 2020, 12, e2020052. [Google Scholar] [CrossRef]
- Karimi, M.; Haghpanah, S.; Azarkeivan, A.; Zahedi, Z.; Zarei, T.; Tavakoli, M.A.; Bazrafshan, A.; Shirkavand, A.; De Sanctis, V. Prevalence and mortality in β-thalassaemias due to outbreak of novel coronavirus disease (COVID-19): The nationwide Iranian experience. Br. J. Haematol. 2020, 190, e137–e140. [Google Scholar] [CrossRef] [PubMed]
- Littera, R.; Campagna, M.; Deidda, S.; Angioni, G.; Cipri, S.; Melis, M.; Firinu, D.; Santus, S.; Lai, A.; Porcella, R.; et al. Human Leukocyte Antigen Complex and Other Immunogenetic and Clinical Factors Influence Susceptibility or Protection to SARS-CoV-2 Infection and Severity of the Disease Course. The Sardinian Experience. Front. Immunol. 2020, 11, 605688. [Google Scholar] [CrossRef] [PubMed]
- Haghpanah, S.; Hosseini-Bensenjan, M.; Sayadi, M.; Karimi, M. Incidence Rate of COVID-19 Infection in Hemoglobinopathies: A Systematic Review and Meta-analysis. Hemoglobin 2021, 45, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Park, K.C.; Donovan, K.; McKechnie, S.; Ramamurthy, N.; Klenerman, P.; Swietach, P. Single-cell oxygen saturation imaging shows that gas exchange by red blood cells is not impaired in COVID-19 patients. Br. J. Haematol. 2020, 190, e229–e232. [Google Scholar] [CrossRef]
- Liu, W.; Li, H. COVID-19: Attacks the 1-beta Chain of Hemoglobin to Disrupt Respiratory Function and Escape Immunity. Camb. Open Engag. 2022, preprint. [Google Scholar]
- Chen, J.-J. Regulation of protein synthesis by the heme-regulated eIF2α kinase: Relevance to anemias. Blood 2007, 109, 2693–2699. [Google Scholar] [CrossRef]
- Lu, L.; Han, A.-P.; Chen, J.-J. Translation Initiation Control by Heme-Regulated Eukaryotic Initiation Factor 2α Kinase in Erythroid Cells under Cytoplasmic Stresses. Mol. Cell. Biol. 2001, 21, 7971–7980. [Google Scholar] [CrossRef] [PubMed]
- Han, A.; Yu, C.; Lu, L.; Fujiwara, Y.; Browne, C.; Chin, G.; Fleming, M.; Leboulch, P.; Orkin, S.H.; Chen, J. Heme-regulated eIF2alpha kinase (HRI) is required for translational regulation and survival of erythroid precursors in iron deficiency. EMBO J. 2001, 20, 6909–6918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.-J. Translational control by heme-regulated eIF2α kinase during erythropoiesis. Curr. Opin. Hematol. 2014, 21, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-J.; Zhang, S. Heme-regulated eIF2α kinase in erythropoiesis and hemoglobinopathies. Blood 2019, 134, 1697–1707. [Google Scholar] [CrossRef]
- Pittala, V.; Salerno, L.; Romeo, G.; Modica, M.; Siracusa, M. A Focus on Heme Oxygenase-1 (HO-1) Inhibitors. Curr. Med. Chem. 2013, 20, 3711–3732. [Google Scholar] [CrossRef]
- Pechlaner, R.; Willeit, P.; Summerer, M.; Santer, P.; Egger, G.; Kronenberg, F.; Demetz, E.; Weiss, G.; Tsimikas, S.; Witztum, J.L.; et al. Heme Oxygenase-1 Gene Promoter Microsatellite Polymorphism Is Associated with Progressive Atherosclerosis and Incident Cardiovascular Disease. Arter. Thromb. Vasc. Biol. 2015, 35, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Yamada, N.; Yamaya, M.; Okinaga, S.; Nakayama, K.; Sekizawa, K.; Shibahara, S.; Sasaki, H. Microsatellite Polymorphism in the Heme Oxygenase-1 Gene Promoter Is Associated with Susceptibility to Emphysema. Am. J. Hum. Genet. 2000, 66, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, I.; Krögler, J.; Endler, G.; Kaufmann, S.; Mustafa, S.; Exner, M.; Mannhalter, C.; Wagner, O.; Pehamberger, H. A microsatellite polymorphism in theheme oxygenase-1 gene promoter is associated with risk for melanoma. Int. J. Cancer 2006, 119, 1312–1315. [Google Scholar] [CrossRef]
- Hirai, H.; Kubo, H.; Yamaya, M.; Nakayama, K.; Numasaki, M.; Kobayashi, S.; Suzuki, S.; Shibahara, S.; Sasaki, H. Microsatellite polymorphism in heme oxygenase-1 gene promoter is associated with susceptibility to oxidant-induced apoptosis in lymphoblastoid cell lines. Blood 2003, 102, 1619–1621. [Google Scholar] [CrossRef]
- Guenegou, A.; Leynaert, B.; Bénessiano, J.; Pin, I.; Demoly, P.; Neukirch, F.; Boczkowski, J.; Aubier, M. Association of lung function decline with the heme oxygenase-1 gene promoter microsatellite polymorphism in a general population sample. Results from the European Community Respiratory Health Survey (ECRHS), France. J. Med. Genet. 2006, 43, e43. [Google Scholar] [CrossRef] [Green Version]
- Exner, M.; Schillinger, M.; Minar, E.; Mlekusch, W.; Schlerka, G.; Haumer, M.; Mannhalter, C.; Wagner, O. Heme Oxygenase-1 Gene Promoter Microsatellite Polymorphism is Associated with Restenosis after Percutaneous Transluminal Angioplasty. J. Endovasc. Ther. 2001, 8, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Song, F.; Li, X.; Rong, S.; Yang, W.; Wang, D.; Xu, J.; Fu, J.; Zhao, Y.; Liu, L. Association between Heme Oxygenase-1 Gene Promoter Polymorphisms and Type 2 Diabetes Mellitus: A HuGE Review and Meta-Analysis. Am. J. Epidemiol. 2010, 172, 631–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, M.I.; Jiang, J.; Silver, N.; Best, S.; Menzel, S.; Mijovic, A.; Colella, S.; Ragoussis, J.; Garner, C.; Weiss, M.J.; et al. alpha-Haemoglobin stabilising protein is a quantitative trait gene that modifies the phenotype of beta-thalassaemia. Br. J. Haematol. 2006, 133, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Zhou, S.; Kihm, A.J.; Katein, A.M.; Yu, X.; Gell, D.A.; Mackay, J.P.; Adachi, K.; Foster-Brown, L.; Louden, C.S.; et al. Loss of α-hemoglobin–stabilizing protein impairs erythropoiesis and exacerbates β-thalassemia. J. Clin. Investig. 2004, 114, 1457–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockman, J.A.; Weiner, L.S.; Simon, G.E.; Stuart, M.J.; Oski, F.A. The measurement of free erythrocyte porphyrin (FEP) as a simple means of distinguishing iron deficiency from beta-thalassemia trait in subjects with microcytosis. J. Lab. Clin. Med. 1975, 85, 113–119. [Google Scholar]
- Bajwa, H.; Basit, H. Thalassemia. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK545151/ (accessed on 5 November 2021).
- Vargas-Vargas, M.; Cortés-Rojo, C. Ferritin levels and COVID-19. Rev. Panam. Salud Pública 2020, 44, e72. [Google Scholar] [CrossRef] [PubMed]
- Millet, G.P.; Debevec, T.; Brocherie, F.; Burtscher, M.; Burtscher, J. Altitude and COVID-19: Friend or foe? A narrative review. Physiol. Rep. 2020, 8, e14615. [Google Scholar] [CrossRef]
- Papadopoulos, K.I.; Sutheesophon, W.; Manipalviratn, S.; Aw, T.-C. Age and genotype dependent erythropoietin protection in COVID-19. World J. Stem Cells 2021, 13, 1513–1529. [Google Scholar] [CrossRef]
- Papadopoulos, K.I.; Sutheesophon, W.; Manipalviratn, S.; Aw, T.C. A Southeast Asian Perspective on the COVID-19 Pandemic: Hemoglobin E (HbE)-Trait Confers Resistance Against COVID-19. Med. Sci. Monit. Basic Res. 2021, 27, e929207. [Google Scholar] [CrossRef]
- Dhangadamajhi, G.; Mohapatra, B.N.; Kar, S.K.; Ranjit, M. Gene polymorphisms in angiotensin I converting enzyme (ACE I/D) and angiotensin II converting enzyme (ACE2 C→T) protect against cerebral malaria in Indian adults. Infect. Genet. Evol. 2010, 10, 337–341. [Google Scholar] [CrossRef]
- Brown, N.J.; Blais, C.; Gandhi, S.K.; Adam, A. ACE Insertion/Deletion Genotype Affects Bradykinin Metabolism. J. Cardiovasc. Pharmacol. 1998, 32, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Elliott, H.L.; Morton, J.J.; Connell, J.M.C. Enhanced Pressor Response to Angiotensin I in Normotensive Men with the Deletion Genotype (DD) for Angiotensin-Converting Enzyme. Hypertension 1995, 25, 1266–1269. [Google Scholar] [CrossRef] [PubMed]
- Brines, M.; Cerami, A. Erythropoietin-mediated tissue protection: Reducing collateral damage from the primary injury response. J. Intern. Med. 2008, 264, 405–432. [Google Scholar] [CrossRef] [PubMed]
- Dore, F.; Bonfigli, S.; Gaviano, E.; Pardini, S.; Cianciulli, P.; Papa, G.; Longinotti, M. Serum erythropoietin levels in thalassemia intermedia. Ann. Hematol. 1993, 67, 183–186. [Google Scholar] [CrossRef]
- Manor, D.; Fibach, E.; Goldfarb, A.; Rachmilewitz, E.A. Erythropoietin activity in the serum of beta thalassemic patients. Scand. J. Haematol. 2009, 37, 221–228. [Google Scholar] [CrossRef]
- Zores, F.; Rebeaud, M. COVID and the Renin-Angiotensin System: Are Hypertension or Its Treatments Deleterious? Front. Cardiovasc. Med. 2020, 7, 71. [Google Scholar] [CrossRef]
- Menzel, S.; Garner, C.; Gut, I.; Matsuda, F.; Yamaguchi, M.; Heath, S.; Foglio, M.; Zelenika, D.; Boland, A.; Rooks, H.; et al. A QTL influencing F cell production maps to a gene encoding a zinc-finger protein on chromosome 2p15. Nat. Genet. 2007, 39, 1197–1199. [Google Scholar] [CrossRef]
- Uda, M.; Galanello, R.; Sanna, S.; Lettre, G.; Sankaran, V.G.; Chen, W.; Usala, G.; Busonero, F.; Maschio, A.; Albai, G.; et al. Genome-wide association study shows BCL11A associated with persistent fetal hemoglobin and amelioration of the phenotype of -thalassemia. Proc. Natl. Acad. Sci. USA 2008, 105, 1620–1625. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lai, Y.; Shi, L. BCL11A Down-Regulation Induces γ-Globin in Human β-Thalassemia Major Erythroid Cells. Hemoglobin 2018, 42, 225–230. [Google Scholar] [CrossRef]
- Lettre, G.; Sankaran, V.G.; Bezerra, M.A.C.; Araújo, A.S.; Uda, M.; Sanna, S.; Cao, A.; Schlessinger, D.; Costa, F.F.; Hirschhorn, J.N.; et al. DNA polymorphisms at the BCL11A, HBS1L-MYB, and β- globin loci associate with fetal hemoglobin levels and pain crises in sickle cell disease. Proc. Natl. Acad. Sci. USA 2008, 105, 11869–11874. [Google Scholar] [CrossRef] [Green Version]
- Sotiriou, S.; Samara, A.A.; Vamvakopoulou, D.; Vamvakopoulos, K.-O.; Sidiropoulos, A.; Vamvakopoulos, N.; Janho, M.B.; Gourgoulianis, K.I.; Boutlas, S. Susceptibility of β-Thalassemia Heterozygotes to COVID-19. J. Clin. Med. 2021, 10, 3645. [Google Scholar] [CrossRef] [PubMed]
- Sotiriou, S.; Samara, A.A.; Lachanas, K.E.; Vamvakopoulou, D.; Vamvakopoulos, K.-O.; Vamvakopoulos, N.; Janho, M.B.; Perivoliotis, K.; Donoudis, C.; Daponte, A.; et al. Vulnerability of β-Thalassemia Heterozygotes to COVID-19: Results from a Cohort Study. J. Pers. Med. 2022, 12, 352. [Google Scholar] [CrossRef] [PubMed]
- Laredo, M.; Curis, E.; Masson-Fron, E.; Voicu, S.; Mégarbane, B. Does COVID-19 alter the oxyhemoglobin dissociation curve?—An observational cohort study using a mixed-effect modelling. Clin. Chem. Lab. Med. 2021, 59, e416–e419. [Google Scholar] [CrossRef] [PubMed]
- Munn, Z.; Moola, S.; Riitano, D.; Lisy, K. The development of a critical appraisal tool for use in systematic reviews addressing questions of prevalence. Int. J. Health Policy Manag. 2014, 3, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.P.T.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, M.; Smith, G.D.; Schneider, M.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| References | 10 | 11 | 6 |
| Timeline | Prospective | Retrospective | Prospective |
| Duration (days) | 61 days | 181 days | 30 days |
| Country | Greece | Greece | Italy |
| Study type | Cohort study | Cohort study | Case-control |
| Total patients | 255 | 760 | 801 |
| Total patients affected by COVID-19 | 255 | 760 | 182 |
| B-thal heterozygote patients | 45 | 189 | NR |
| B-thal heterozygote patients affected by COVID-19 | 45 | 189 | 19 |
| Severity | |||
| Total patients | |||
| Mild affected by COVID-19 | 68 | 190 | 143 |
| Moderate affected by COVID | 113 | 373 | 39 |
| Severe and Critical affected by COVID-19 | 74 | 197 | |
| B-thal heterozygotes | |||
| Mild affected by COVID-19 | 5 | 15 | 19 |
| Moderate affected by COVID | 19 | 66 | 0 |
| Severe and Critical affected by COVID-19 | 21 | 56 | |
| Mortality | |||
| Total deceased | 70 | 189 | NR |
| B-thal heterozygote deceased | 20 | 53 | NR |
| Total deceased by COVID-19 | 70 | 189 | NR |
| B-thal heterozygote deceased by COVID-19 | 20 | 53 | NR |
| ICU admission | |||
| Total ICU | 53 | NR | 39 |
| B-thal heterozygote ICU | 11 | NR | 0 |
| Comorbidities in the general population | |||
| Male | 153 | 448 | 70 |
| Age: mean ± SD | 61.56 (±16.597) | 62.21 (±16.42) | 53.2 ± 18.1 |
| Diabetes mellitus | 53 | 156 | 6 |
| Hypertension | 142 | 420 | 27 |
| Cardiovascular disease | 82 | 227 | NR |
| Cancer | 29 | 85 | 4 |
| Chronic Pulmonary Disease (emphysema or/and bronchitis) | 32 | 94 | 2 |
| Autoimmune disease (Hashimoto thyroiditis, rheumatoid arthritis, systemic lupus, and autoimmune hepatitis) | NR | NR | 22 |
| Hypercholesterolemia | 113 | NR | 18 |
| Stroke | 50 | 142 | 14 |
| Comorbidities in B-thal minor population | |||
| Male | NR | NR | NR |
| Age: mean ± SD | NR | NR | NR |
| Diabetes mellitus | NR | NR | NR |
| Hypertension | NR | NR | NR |
| Cardiovascular disease | NR | NR | NR |
| Cancer | NR | NR | NR |
| Chronic Pulmonary Disease (emphysema or/and bronchitis) | NR | NR | NR |
| Autoimmune disease (Hashimoto thyroiditis, rheumatoid arthritis, systemic lupus, and autoimmune hepatitis) | NR | NR | NR |
| Hypercholesterolemia | NR | NR | NR |
| Stroke | NR | NR | NR |
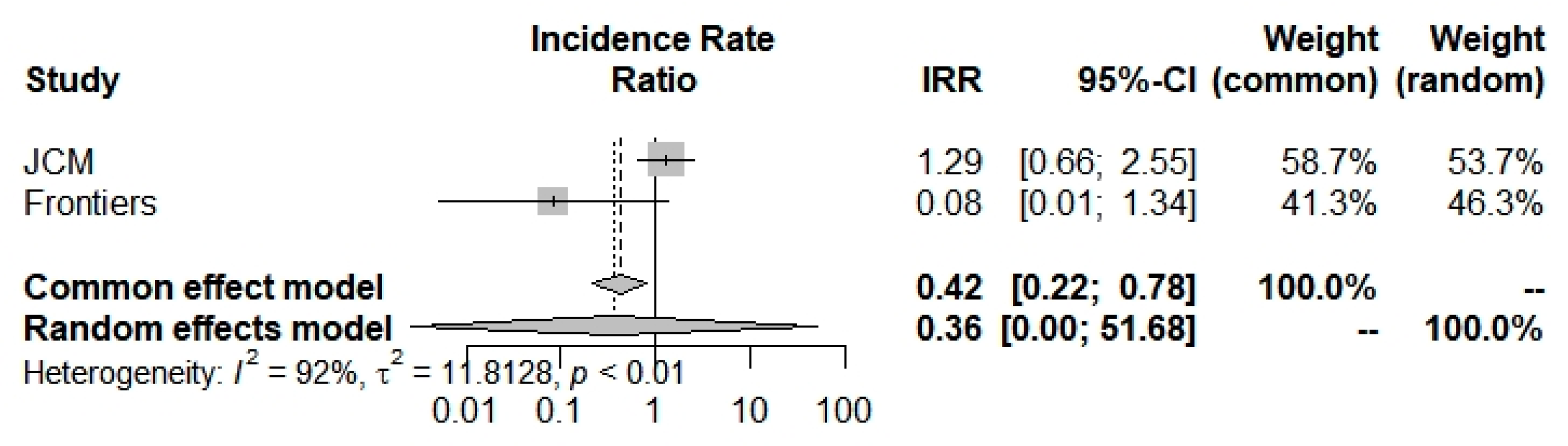
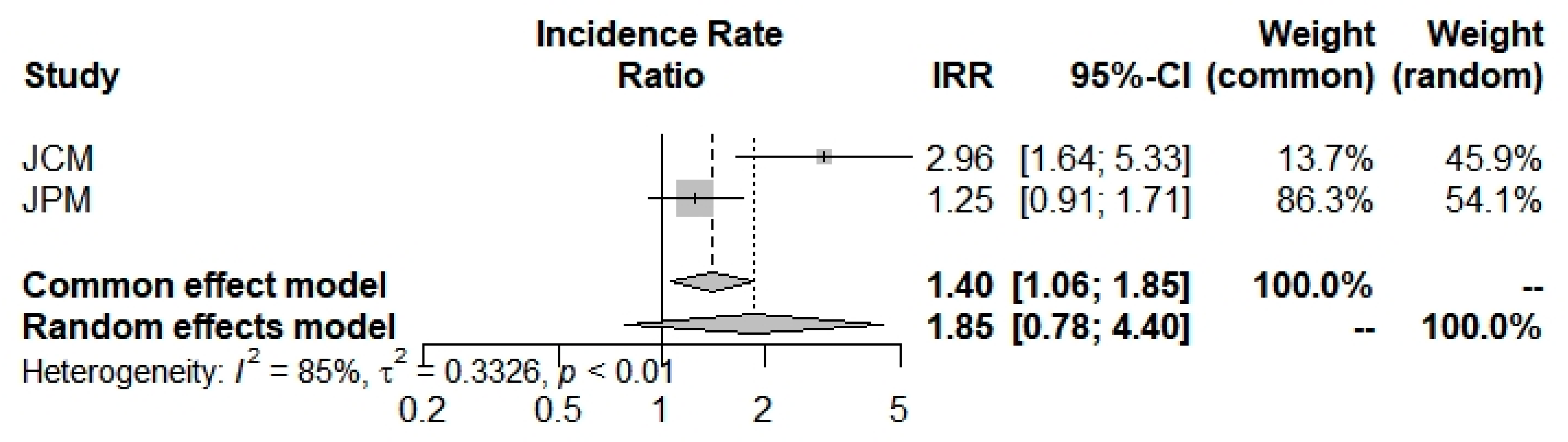
| Incidence Rate (95% CI) | Severity Rate (95% CI) | ICU Admission Rate (95% CI) | Mortality Rate (95% CI) | ||
|---|---|---|---|---|---|
| Overall | index | 0.9250 (0.5752; 1.4877) | 1.5933 (0.4884; 5.1981) | 0.3620 (0.0025; 51.6821) | 1.8542 (0.7819; 4.3970) |
| z | −0.32 | 0.77 | −0.40 | 1.40 | |
| p value | 0.7479 | 0.4400 | 0.6881 | 0.1610 | |
| I2 (%) | - | 90.0% (73.1–96.2%) | 91.7% (71.0–97.6%) | 85.1% (39.7%; 96.3%) | |
| p value | - | 0.0001 | 0.0005 | 0.0095 | |
| Egger’s test | 95% CI for bias | - | (−4.65; −0.12) | - | - |
| p-value | - | 0.3138047 | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lansiaux, E.; Drouin, E.; Bolm, C. Beta-Thalassemia Minor and SARS-CoV-2: Physiopathology, Prevalence, Severity, Morbidity, and Mortality. Thalass. Rep. 2023, 13, 21-32. https://doi.org/10.3390/thalassrep13010003
Lansiaux E, Drouin E, Bolm C. Beta-Thalassemia Minor and SARS-CoV-2: Physiopathology, Prevalence, Severity, Morbidity, and Mortality. Thalassemia Reports. 2023; 13(1):21-32. https://doi.org/10.3390/thalassrep13010003
Chicago/Turabian StyleLansiaux, Edouard, Emmanuel Drouin, and Carsten Bolm. 2023. "Beta-Thalassemia Minor and SARS-CoV-2: Physiopathology, Prevalence, Severity, Morbidity, and Mortality" Thalassemia Reports 13, no. 1: 21-32. https://doi.org/10.3390/thalassrep13010003