
Response of Phenolic Compounds in Lippia graveolens Kunth Irrigated with Aquaculture Wastewater and Steiner Solution

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
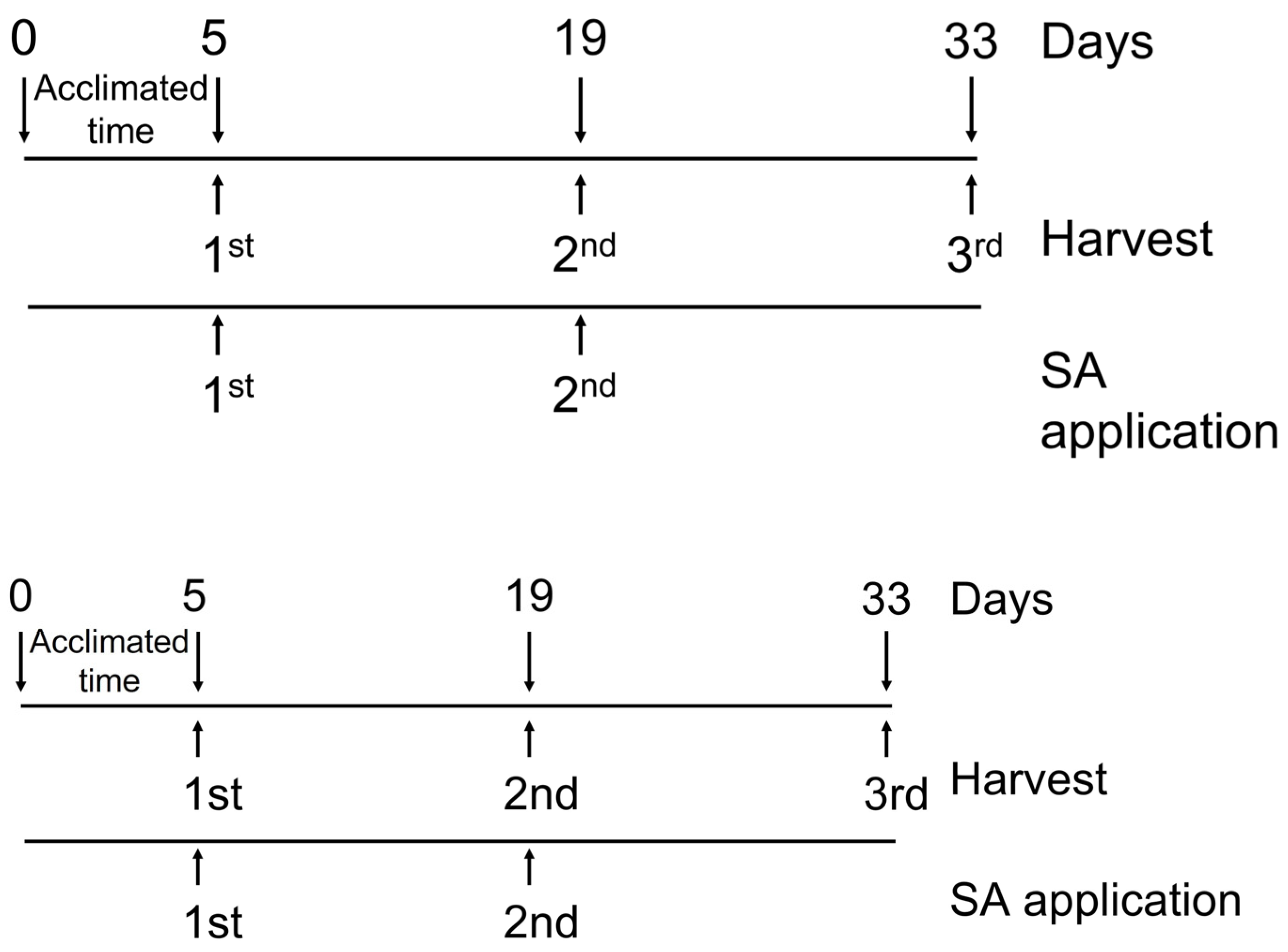
2.1. Plant Material and Culture Conditions
2.2. Irrigation
2.3. Elicitation
2.4. N and P Determination
2.5. Vegetative Sample
2.6. Phenolic Compounds Extraction
2.7. Phenolic Content Determination
2.8. Flavonoid Content Determination
2.9. Antioxidant Capacity
2.9.1. DPPH Assay
2.9.2. FRAP Assay
2.10. Statistical Analysis
3. Results
3.1. Water Quality (N and P Determination)
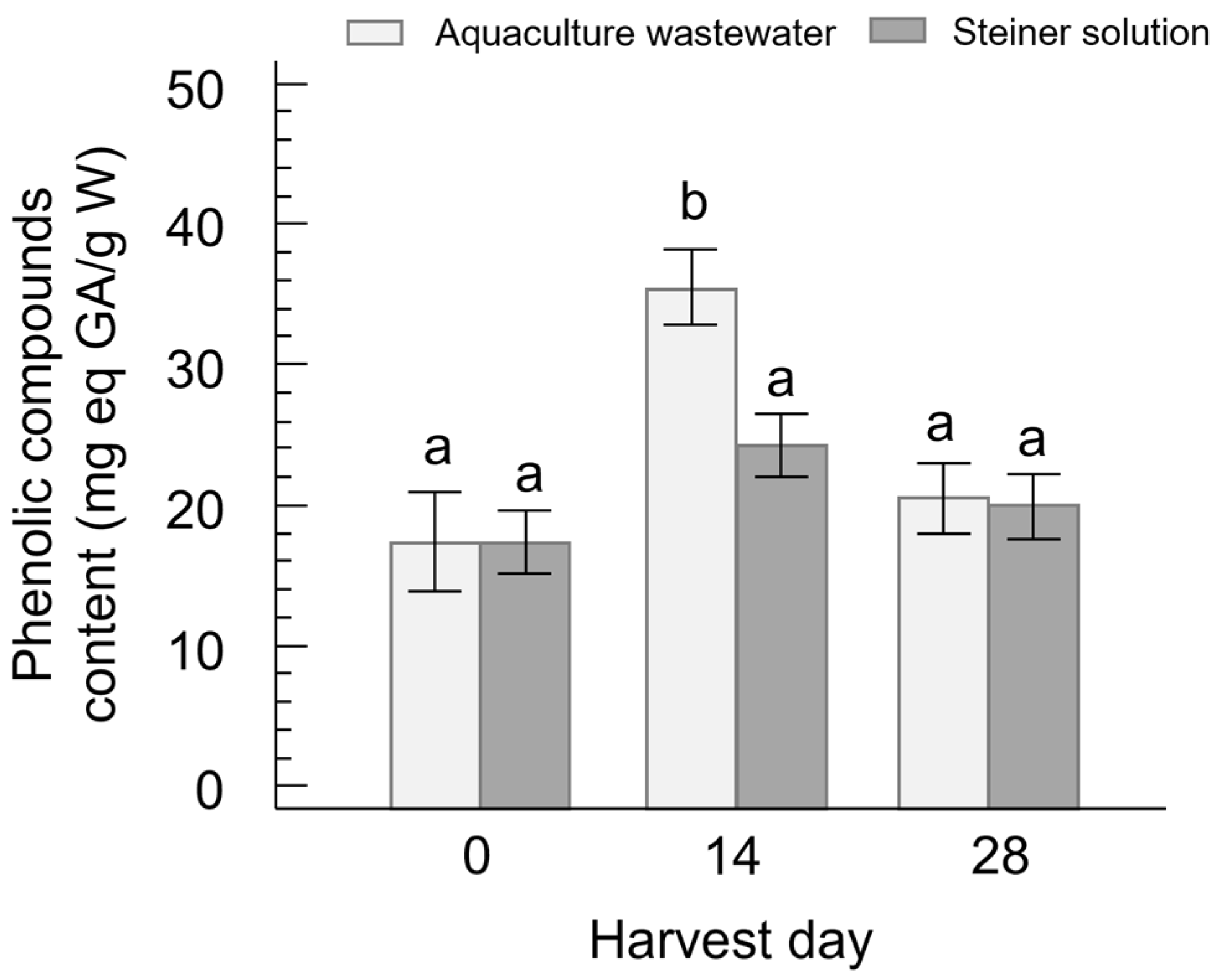
3.2. Phenolic Content
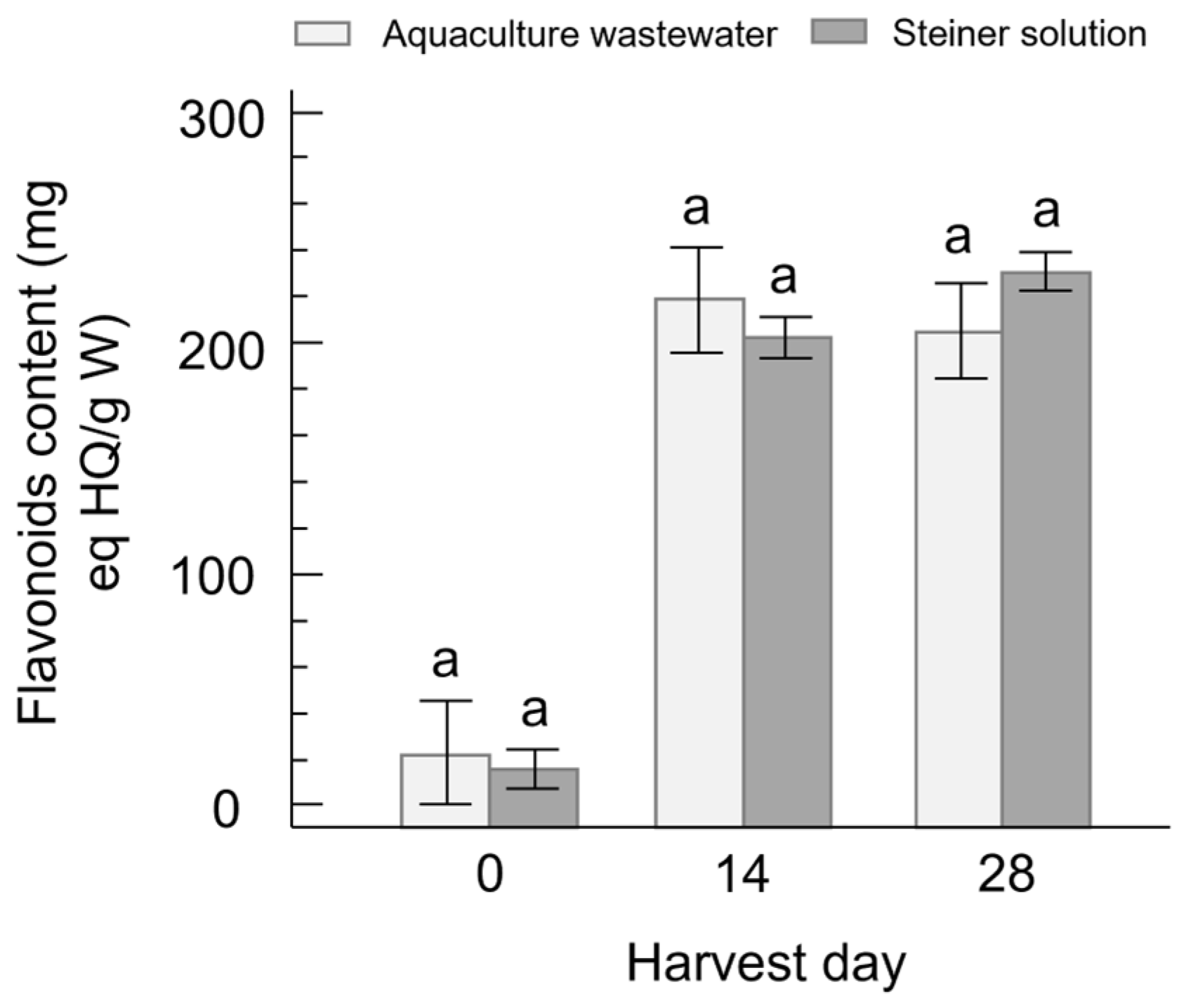
3.3. Flavonoid Content
3.4. Antioxidant Activity
3.4.1. DPPH Assay
3.4.2. FRAP Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martínez-Natarén, D.; Parra-Tabla, V.; Dzib, G.; Acosta-Arriola, V.; Canul Puc, K.; Calvo Irabién, L.M. Essential oil Yield Variation Within and Among Wild Populations of Mexican Oregano (Lippia graveolens H.B.K.-Verbenaceae), and its Relation to Climatic and Edaphic Conditions. J. Essent. Oil Bear. Plants 2012, 15, 589–601. [Google Scholar] [CrossRef]
- García-Pérez, E.; Castro-Álvarez, F.F.; Gutiérrez-Uribe, J.A.; García-Lara, S. Revisión de la producción, composición fitoquímica y propiedades nutracéuticas del orégano mexicano. Rev. Mex. De Cienc. Agrícolas 2012, 3, 339–353. [Google Scholar]
- Ercioglu, E.; Velioglu, H.M.; Boyaci, I.H. Determination of terpenoid contents of aromatic plants using NIRS. Talanta 2018, 178, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Mikallou, M.; Petropoulos, S.; Tzortzakis, N. Profiling of Essential Oils Components and Polyphenols for Their Antioxidant Activity of Medicinal and Aromatic Plants Grown in Different Environmental Conditions. Agronomy 2020, 10, 727. [Google Scholar] [CrossRef]
- Calvo-Irabién, L.M.; Parra-Tabla, V.; Acosta-Arriola, V.; Escalante-Erosa, F.; Dias-Vera, L.; Dzib, G.R.; Peña-Rodríguez, M.L. Phytochemical diversity of the essencial oils of mexican oregano (Lippia graveolens KUNTH) populations along an Edapho-Climatic gradient. Chem. Biodivers. 2014, 11, 1010–1021. [Google Scholar] [CrossRef]
- Martínez-Natarén, D.A.; Parra-Tabla, V.; Ferrer-Ortega, M.M.; Calvo-Irabién, M.L. Genetic diversity and genetic structure in wild populations of Mexican oregano (Lippia graveolens H.B.K) and its relationship with the chemical composition of the essencial oil. Plant Syst. Evol. 2014, 300, 535–547. [Google Scholar] [CrossRef]
- Tezara, W.; Coronel, I.; Herrera, A.; Dzib, G.; Canul-Puc, K.; Calvo-Irabién, L.M.; Gonzales-Meler, M. Photosynthetic capacity and terpene production in populations of Lippia graveolens (Mexican oregano) growing wild and in a common garden in Yucatán peninsula. Ind. Crops Prod. 2014, 57, 1–9. [Google Scholar] [CrossRef]
- Hashim, M.; Ahmad, B.; Drouet, S.; Hano, C.; Abbasi, B.H.; Anjum, S. Comparative Effects of Different Light Sources on the Production of Key Secondary Metabolites in Plants In Vitro Cultures. Plants 2021, 10, 1521. [Google Scholar] [CrossRef]
- Ramak, P.; Kazempour Osaloo, S.; Ebrahimzadeh, H.; Sharifi, M.; Behmanesh, M. Inhibition of the mevalonate pathway enhances carvacrol biosynthesis and DXR gene expression in shoot cultures of Satureja khuzistanica Jamzad. J. Plant Physiol. 2013, 170, 1187–1193. [Google Scholar] [CrossRef]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Khan, F.A.; Naushin, F. Chapter 9—Role and Regulation of Plants Phenolics in Abiotic Stress Tolerance: An Overview. In Plant Signaling Molecules; Khan, M.I.R., Reddy, P.S., Ferrante, A., Khan, N.A., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 157–168. [Google Scholar] [CrossRef]
- Lv, Z.-Y.; Sun, W.-J.; Jiang, R.; Chen, J.-F.; Ying, X.; Zhang, L.; Chen, W.-S. Phytohormones jasmonic acid, salicylic acid, gibberellins, and abscisic acid are key mediators of plant secondary metabolites. World J. Trad. Chin. Med. 2021, 7, 307–325. [Google Scholar] [CrossRef]
- Bueno-Duran, A.Y.; Cervantes-Martínez, J.; Obledo-Vázquez, E.N. Composition of essencial oil from Lippia graveolens. Relationship between spetral light quality and thymol and carvacrol content. J. Essenc. Oil Res. 2013, 26, 153–160. [Google Scholar] [CrossRef]
- Clarenc Aarland, R.; Castellanos-Hernández, O.A.; Rodríguez-Sahagún, A.; Acevedo-Hernández, G.J. Efecto del estrés salino sobre la morfología y fitoquímica de orégano mexicano (Lippia graveolens Kunth) cultivado in vitro. Biotecnia 2020, 22, 131–137. [Google Scholar] [CrossRef]
- Sęczyk, Ł.; Król, B.; Kołodziej, B. In vitro bioaccessibility and activity of Greek oregano (Origanum vulgare L. ssp. hirtum (link) Ietswaart) compounds as affected by nitrogen fertilization. J. Sci. Food Agric. 2020, 100, 2410–2417. [Google Scholar] [CrossRef]
- Tom, A.P.; Jayakumar, J.S.; Biju, M.; Somarajan, J.; Ibrahim, M.A. Aquaculture wastewater treatment technologies and their sustainability: A review. Energy Nexus 2021, 4, 100022. [Google Scholar] [CrossRef]
- Braglia, R.; Costa, P.; Di Marco, G.; D’Agostino, A.; Redi, E.L.; Scuderi, F.; Gismondi, A.; Canini, A. Phytochemicals and quality level of food plants grown in an aquaponics system. J. Sci. Food Agric. 2022, 102, 844–850. [Google Scholar] [CrossRef]
- Kimera, F.; Sewilam, H.; Fouad, W.M.; Suloma, A. Efficient utilization of aquaculture effluents to maximize plant growth, yield, and essential oils composition of Origanum majorana cultivation. Ann. Agric. Sci. 2021, 66, 1–7. [Google Scholar] [CrossRef]
- Kimera, F.; Sewilam, H.; Fouad, W.M.; Suloma, A. Sustainable production of Origanum syriacum L. using fish effluents improved plant growth, yield, and essential oil composition. Heliyon 2021, 7, e06423. [Google Scholar] [CrossRef]
- Azizi, A.; Yan, F.; Honermeier, B. Herbage yield, essential oil content and composition of three oregano (Origanum vulgare L.) populations as affected by soil moisture regimes and nitrogen supply. Ind. Crops Prod. 2009, 29, 554–561. [Google Scholar] [CrossRef]
- Ali, B. Salicylic acid: An efficient elicitor of secondary metabolite production in plants. Biocatal. Agric. Biotechnol. 2021, 31, 101884. [Google Scholar] [CrossRef]
- Cappellari, L.d.R.; Santoro, M.V.; Schmidt, A.; Gershenzon, J.; Banchio, E. Improving Phenolic Total Content and Monoterpene in Mentha x piperita by Using Salicylic Acid or Methyl Jasmonate Combined with Rhizobacteria Inoculation. Int. J. Mol. Sci. 2020, 21, 50. [Google Scholar] [CrossRef] [PubMed]
- Saharkhiz, M.J.; Goudarzi, T. Foliar Application of Salicylic acid Changes Essential oil Content and Chemical Compositions of Peppermint (Mentha piperita L.). J. Essent. Oil Bear. Plants 2014, 17, 435–440. [Google Scholar] [CrossRef]
- Alamer, K.H.; Fayez, K.A. Impact of salicylic acid on the growth and physiological activities of parsley plants under lead toxicity. Physiol. Mol. Biol. Plants 2020, 26, 1361–1373. [Google Scholar] [CrossRef]
- Gharib, F.A.E.-L. Effect of salicylic acid on the growth, metabolic activities and oil content of basil and marjoram. Int. J. Agric. Biol. 2006, 4, 485–492. [Google Scholar]
- Momeni, M.; Pirbalouti, A.G.; Mousavi, A.; Badi, H.N. Effect of Foliar Applications of Salicylic Acid and Chitosan on the Essential Oil of Thymbra spicata L. under Different Soil Moisture Conditions. J. Essent. Oil Bear. Plants 2020, 23, 1142–1153. [Google Scholar] [CrossRef]
- Karlidag, H.; Yildirim, E.; Turan, M. Exogenous applications of salicylic acid affect quality and yield of strawberry grown under antifrost heated greenhouse conditions. J. Plant Nutr. Soil Sci. 2009, 172, 270–276. [Google Scholar] [CrossRef]
- Haider Abbasi, B.; Ullah, M.A.; Nadeem, M.; Tungmunnithum, D.; Hano, C. Exogenous application of salicylic acid and gibberellic acid on biomass accumulation, antioxidant and anti-inflammatory secondary metabolites production in multiple shoot culture of Ajuga integrifolia Buch. Ham. ex D.Don. Ind. Crops Prod. 2020, 145, 112098. [Google Scholar] [CrossRef]
- Figueroa Pérez, M.G.; Rocha-Guzmán, N.E.; Mercado-Silva, E.; Loarca-Piña, G.; Reynoso-Camacho, R. Effect of chemical elicitors on peppermint (Mentha piperita) plants and their impact on the metabolite profile and antioxidant capacity of resulting infusions. Food Chem. 2014, 156, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Corral-Aguayo, R.D.; Yahia, E.M.; Carrillo-Lopez, A.; González-Aguilar, G. Correlation between some nutritional components and the total antioxidant capacity measured with six different assays in eight horticultural crops. J. Agric. Food Chem. 2008, 56, 10498–10504. [Google Scholar] [CrossRef]
- Moon, J.-K.; Shibamoto, T. Antioxidant Assays for Plant and Food Components. J. Agric. Food Chem. 2009, 57, 1655–1666. [Google Scholar] [CrossRef]
- Król, B.; Sęczyk, Ł.; Kołodziej, B.; Paszko, T. Biomass production, active substance content, and bioaccessibility of Greek oregano (Origanum vulgare ssp. hirtum (Link) Ietswaart) following the application of nitrogen. Ind. Crops Prod. 2020, 148, 112271. [Google Scholar] [CrossRef]
- Stumpf, B.; Yan, F.; Honermeier, B. Influence of nitrogen fertilization on yield and phenolic compounds in wheat grains (Triticum aestivum L. ssp. aestivum). J. Plant Nutr. Soil Sci. 2019, 182, 111–118. [Google Scholar] [CrossRef]
- Félix-Cuencas, L.; García-Trejo, J.F.; López-Tejeida, S.; León-Ramírez, J.J.d.; Soto-Zarazúa, G.M. Effect of three productive stages of tilapia (Oreochromis niloticus) under hyper-intensive recirculation aquaculture system on the growth of tomato (Solanum lycopersicum). Lat. Am. J. Aquat. Res. 2021, 49, 689–701. [Google Scholar]
- Heimler, D.; Romani, A.; Ieri, F. Plant polyphenol content, soil fertilization and agricultural management: A review. Eur. Food Res. Technol. 2017, 243, 1107–1115. [Google Scholar] [CrossRef]
- An, C.; Mou, Z. Salicylic Acid and its Function in Plant ImmunityF. JIPB 2011, 53, 412–428. [Google Scholar] [CrossRef]
- Rahmani, N.; Taherkhani, T.; Zandi, P.; Aghdam, A. Effect of regulated deficit irrigation and nitrogen levels on flavonoid content and extract performance of marigold (Calendula officinalis L.). J. Biol. Res. 2012, 3, 2624–2630. [Google Scholar]
- Deng, B.; Li, Y.; Lei, G.; Liu, G. Effects of nitrogen availability on mineral nutrient balance and flavonoid accumulation in Cyclocarya paliurus. Plant Physiol. Biochem. 2019, 135, 111–118. [Google Scholar] [CrossRef]
- Margna, U. Control at the level of substrate supply—An alternative in the regulation of phenylpropanoid accumulation in plant cells. Phytochemistry 1977, 16, 419–426. [Google Scholar] [CrossRef]
- Chepel, V.; Lisun, V.; Skrypnik, L. Changes in the Content of Some Groups of Phenolic Compounds and Biological Activity of Extracts of Various Parts of Heather (Calluna vulgaris (L.) Hull) at Different Growth Stages. Plants 2020, 9, 926. [Google Scholar]
- Michalska, A.; Wojdyło, A.; Bogucka, B. The influence of nitrogen and potassium fertilisation on the content of polyphenolic compounds and antioxidant capacity of coloured potato. J. Food Compos. Anal. 2016, 47, 69–75. [Google Scholar] [CrossRef]
- Matłok, N.; Stępień, A.E.; Gorzelany, J.; Wojnarowska-Nowak, R.; Balawejder, M. Effects of Organic and Mineral Fertilization on Yield and Selected Quality Parameters for Dried Herbs of Two Varieties of Oregano (Origanum vulgare L.). Appl. Sci. 2020, 10, 5503. [Google Scholar] [CrossRef]
- Biesiada, A.; Sokół-Łętowska, A.; Kucharska, A. The Effect of Nitrogen Fertilization on Yielding and Antioxidant Activity of Lavender (Lavandula angustifolia Mill.). Acta Sci. Pol. Hortorum Cultus 2008, 7, 33–40. [Google Scholar]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Week 1 | Week 2 | Week 3 | Week 4 | |||||
|---|---|---|---|---|---|---|---|---|
| SS 1 | AQW 2 | SS | AQW | SS | AQW | SS | AQW | |
| NO3− | 127.3 ± 4.24 | 108.2 ± 2.22 | 207.7 ± 1.23 | 139.2 ± 6.07 | 290.3 ± 4.32 | 122.3 ± 3.37 | 235.4 ± 2.15 | 103.8 ± 4.57 |
| NO2− | 1.361 ± 0.04 | 0.159 ± 0.07 | 0.313 ± 0.04 | 0.403 ± 0.06 | 3.176 ± 0.28 | 0.562 ± 0.08 | 0.515 ± 0.02 | 0.163 ± 0.02 |
| PO43− | 17.79 ± 0.14 | 24.16 ± 0.27 | 43.09 ± 0.35 | 9.75 ± 1.03 | 114.9 ± 3.18 | 12.52 ± 1.64 | 102.2 ± 6.41 | 10.97 ± 2.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nieto-Ramírez, M.I.; Feregrino-Pérez, A.A.; Aguirre Becerra, H.; Parra-Pacheco, B.; Oviedo-Olvera, M.V.; García-Trejo, J.F. Response of Phenolic Compounds in Lippia graveolens Kunth Irrigated with Aquaculture Wastewater and Steiner Solution. Int. J. Plant Biol. 2023, 14, 483-492. https://doi.org/10.3390/ijpb14020037
Nieto-Ramírez MI, Feregrino-Pérez AA, Aguirre Becerra H, Parra-Pacheco B, Oviedo-Olvera MV, García-Trejo JF. Response of Phenolic Compounds in Lippia graveolens Kunth Irrigated with Aquaculture Wastewater and Steiner Solution. International Journal of Plant Biology. 2023; 14(2):483-492. https://doi.org/10.3390/ijpb14020037
Chicago/Turabian StyleNieto-Ramírez, María Isabel, Ana Angélica Feregrino-Pérez, Humberto Aguirre Becerra, Benito Parra-Pacheco, Mónica Vanessa Oviedo-Olvera, and Juan Fernando García-Trejo. 2023. "Response of Phenolic Compounds in Lippia graveolens Kunth Irrigated with Aquaculture Wastewater and Steiner Solution" International Journal of Plant Biology 14, no. 2: 483-492. https://doi.org/10.3390/ijpb14020037