
Fruit Structure in Amphicarpic Annual Gymnarrhena micrantha (Asteraceae, Gymnarrheneae) in Relation to the Species Biology


Abstract
:1. Introduction
2. Material and Methods
2.1. Fruit Material
2.2. Scanning Electron Microscopy (SEM) and Stereomicroscopy
2.3. Light Microscopy
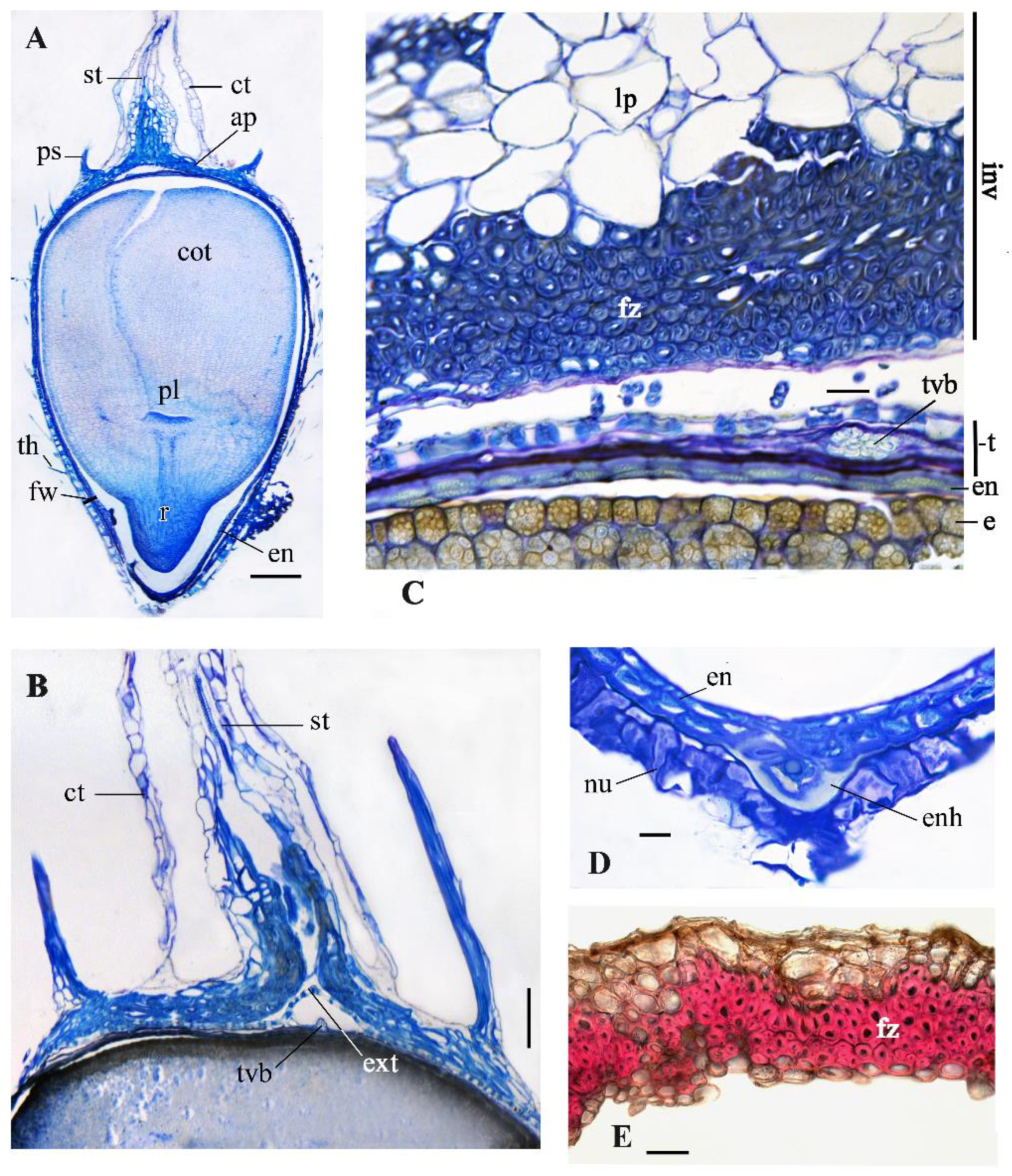
3. Results
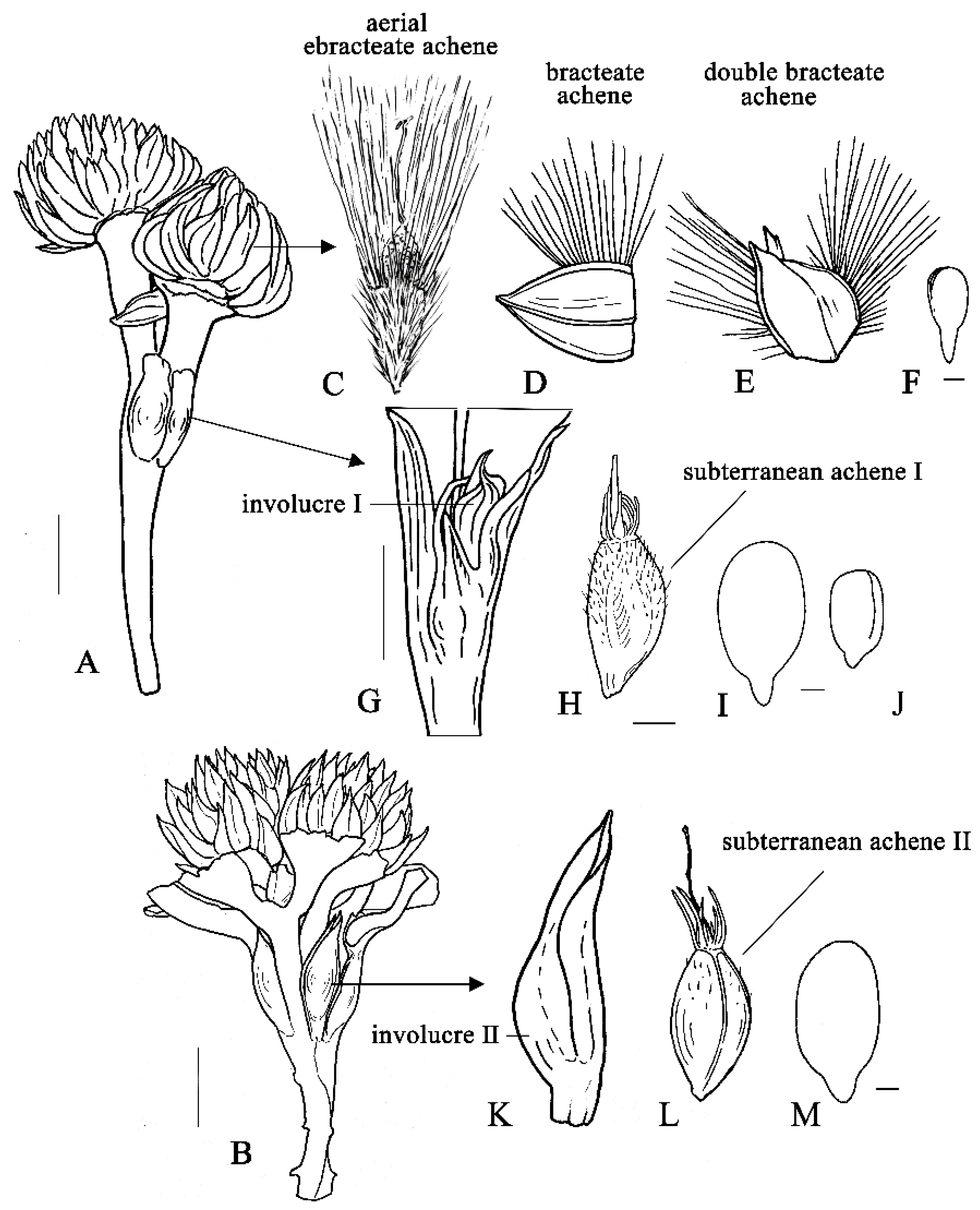
3.1. The Types of Heteromorphic Fruits
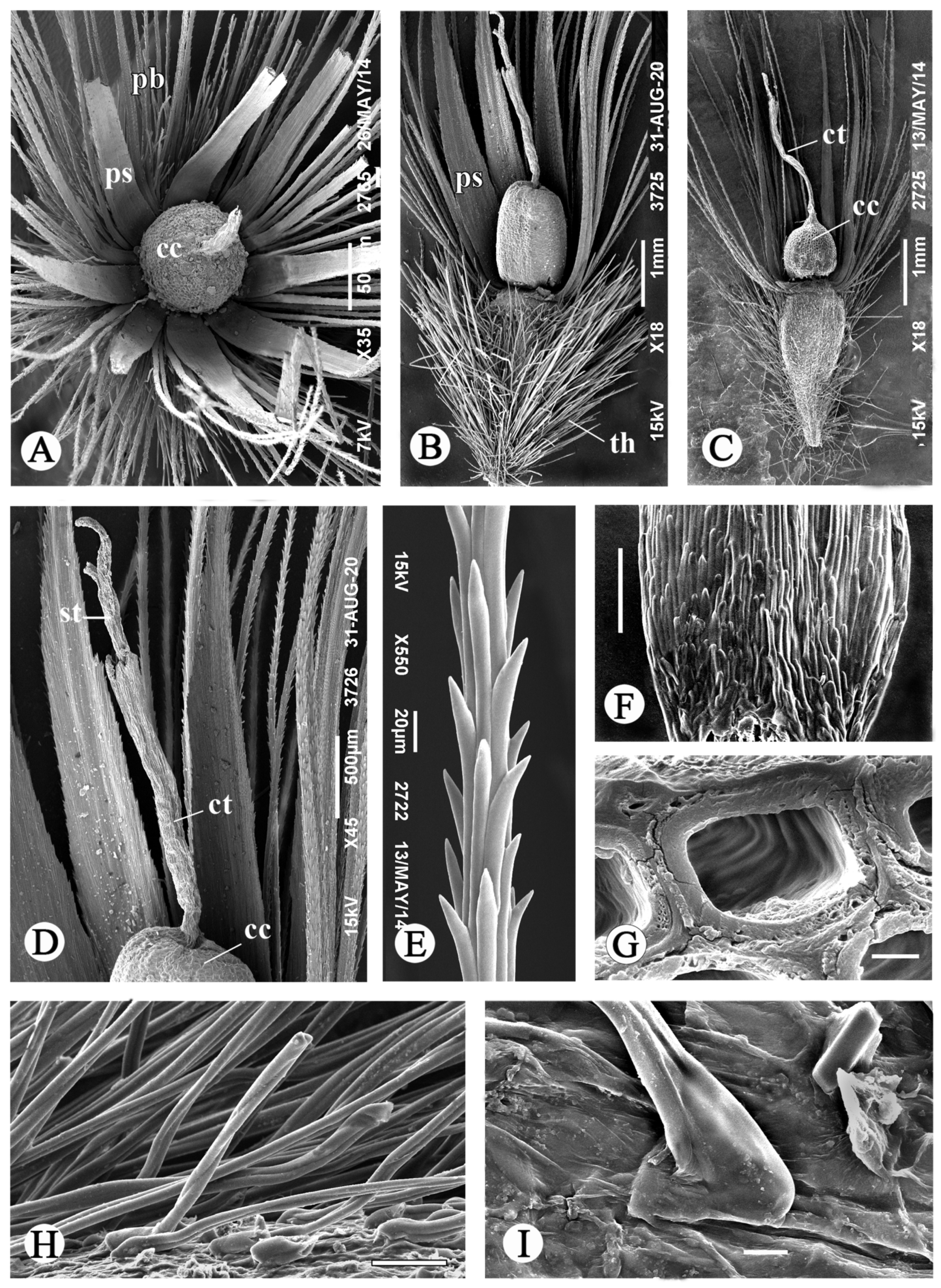
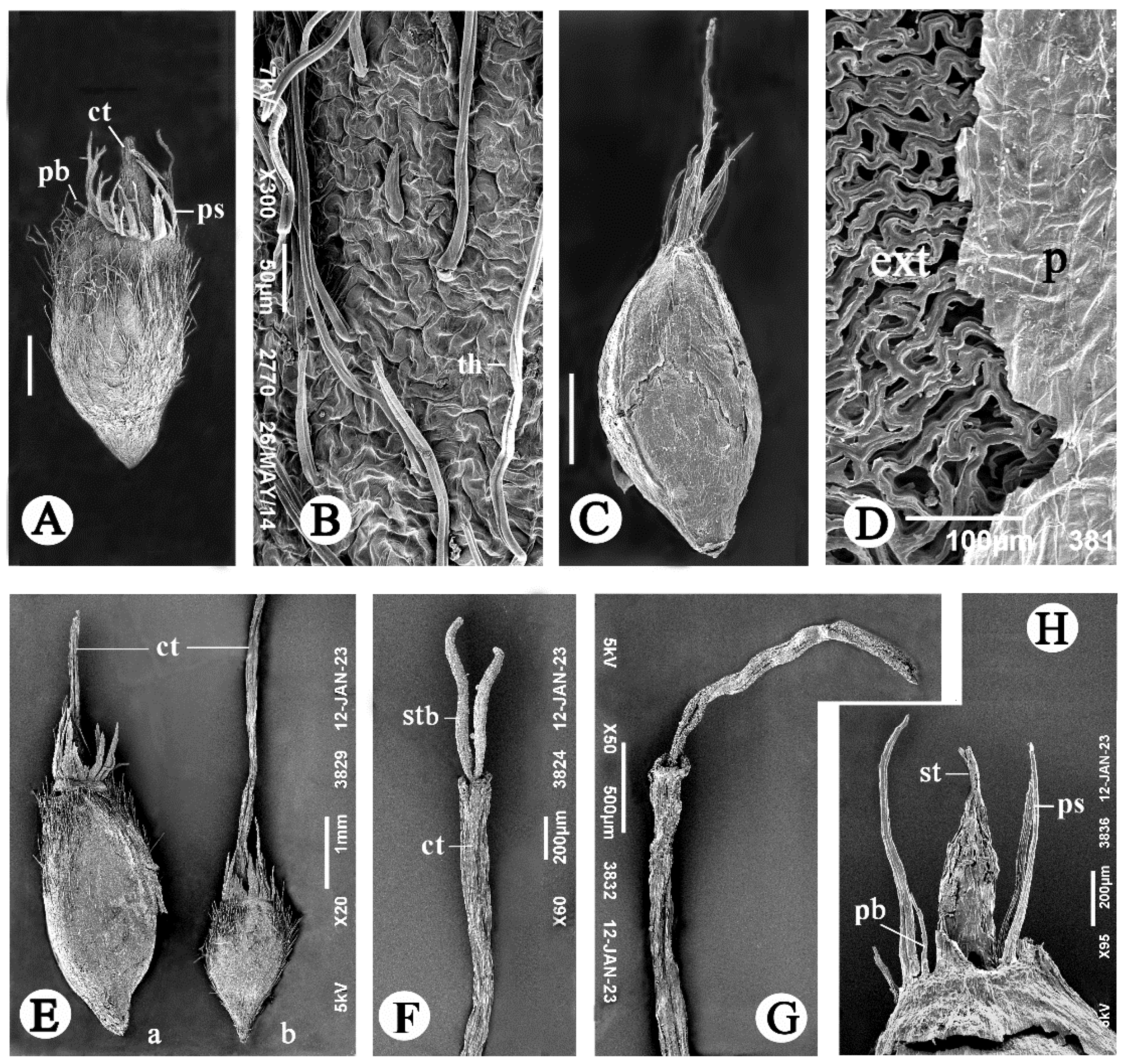
3.1.1. The Aerial Achenes
3.1.2. The Subterranean Achenes
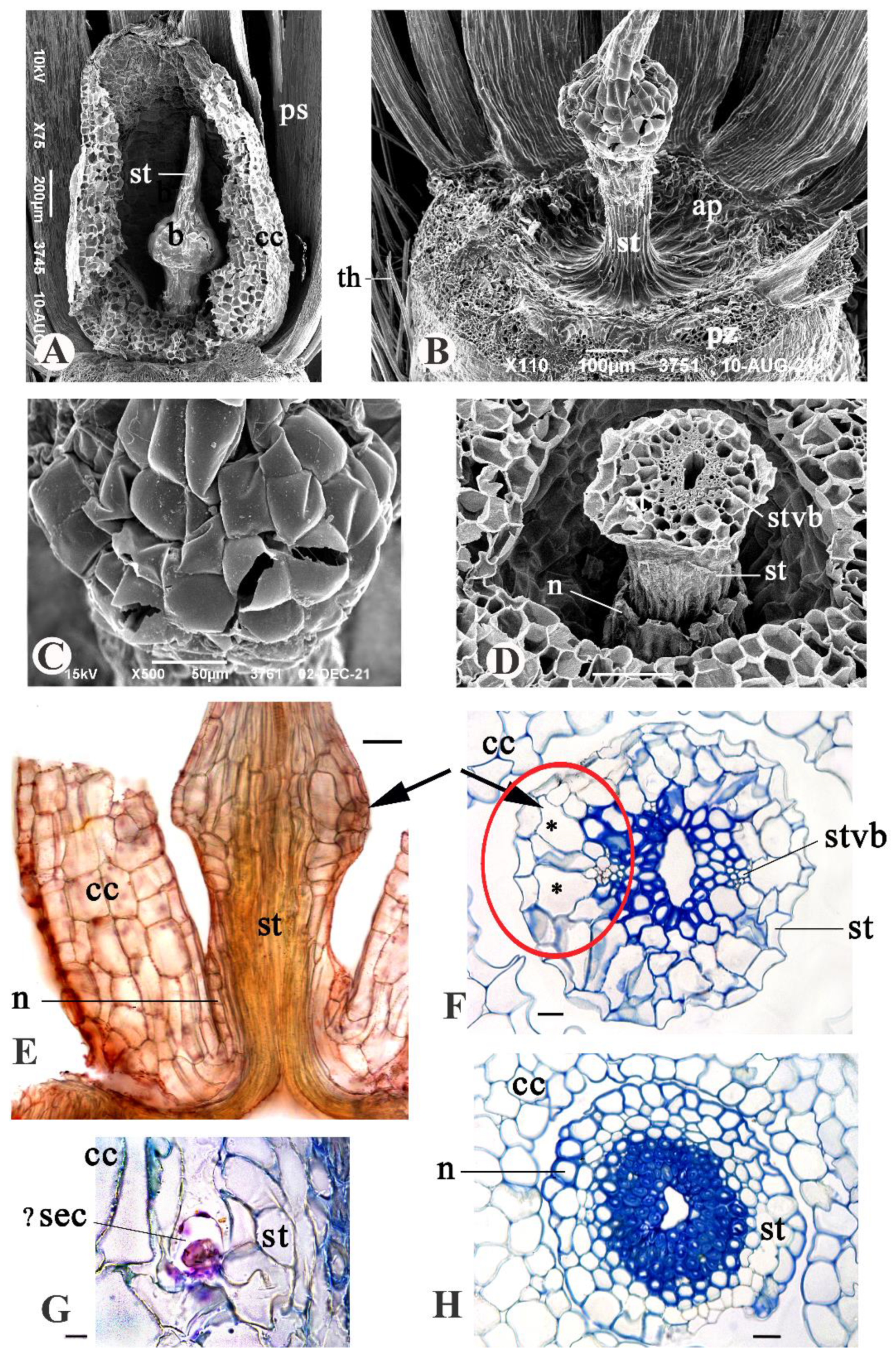
3.2. Anatomical Structure of the Aerial Achenes
3.2.1. The Apical Plate
3.2.2. The Pappus
3.2.3. The Persistent Corolla and the Style
3.2.4. The Basal Achene Region
3.3. Anatomical Structure of Subterranean Fruits
3.4. Fruit Wall Structure
3.4.1. The Pericarp
3.4.2. The Seed Coat (Testa)
3.4.3. The Endosperm
4. Discussion
4.1. Fruit Diversity in Gymnarrhena
4.2. From Which Kind of Flowers Do Subterranean Fruits Develop?
4.3. Structural Differences of Heteromorphic Fruits of Gymnarrhena
4.4. Possible Adaptive Significance of Aerial Achene Structural Features
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zohary, M. Die verbreitungsökologischen Verhältnisse der Pflanzen Palästinas. I. Die antitelechorischen Erscheinungen. Beih. Bot. Centralbl. 1937, 56, 1–155. [Google Scholar]
- van der Pijl, L. Principles of Dispersal in Higher Plants, 3rd ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1982. [Google Scholar] [CrossRef]
- Cheplik, G.P. The ecology of amphicarpic plants. Trends Ecol. Evol. 1987, 2, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Baskin, J.M.; Baskin, C.C.; Cheplick, G.P.; Yang, X.; Huang, Z. Amphicarpic plants: Definition, ecology, geographic distribution, systematics, life history, evolution and use in agriculture. Biol. Rev. 2020, 95, 1442–1466. [Google Scholar] [CrossRef] [PubMed]
- Funk, V.A.; Fragman-Sapir, O. Gymnarrheneae (Gimnarrhenoideae). In Systematics, Evolution, and Biogeography of Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; IAPT: Vienna, Austria, 2009; pp. 328–332. ISBN 978-3-9501754-3-1. [Google Scholar]
- Susanna, A.; Baldwin, B.G.; Bayer, R.J.; Bonifacino, J.M.; Garcia-Jacas, N.; Keeley, S.C.; Mandel, J.R.; Ortiz, S.; Robinson, H.; Stuessy, T.F. The classification of the Compositae: A tribute to Vicki Ann Funk (1947–2019). Taxon 2020, 69, 807–814. [Google Scholar] [CrossRef]
- Kilian, N.; Birgit Gemeinholzer, B.; Lack, H.W. Cichorieae. In Systematics, Evolution, and Biogeography of Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; IAPT: Vienna, Austria, 2009; pp. 343–383. ISBN 978-3-9501754-3-1. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Elsevier/Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Burtt, B.L. Aspects of diversification in the capitulum. In The Biology and Chemistry of the Compositae; Heywood, V.H., Harborne, J.B., Turner, B.L., Eds.; Academic Press: London, UK, 1977; Volume 1, pp. 41–60. ISBN 0123468019. [Google Scholar]
- Imbert, E. Ecological consequences and ontogeny of seed heteromorphism. Perspect. Plant Ecol. Evol. Syst. 2002, 5, 13–36. [Google Scholar] [CrossRef]
- Roth, I. Fruits of Angiosperms; Encyclopedia of Plant Anatomy, Bd 10, Teil 1; G. Borntraeger: Berlin, Germany; Stuttgart, Germany, 1977; ISBN 978-3-443-14010-6. [Google Scholar]
- Voytenko, V.F. Typology and evolution of heterocarpy forms in tribe Lactuceae (Asteraceae). Bot. Zhurn. 1989, 74, 1241–1257. [Google Scholar]
- Matilla, A.; Gallardo, M.; Puga-Hermida, M.I. Structural, physiological and molecular aspects of heterogeneity in seeds: A review. Seed Sci. Res. 2005, 15, 63–76. [Google Scholar] [CrossRef]
- Marzinek, J.; Nakajima, J.N.; Marques, D.; De-Paula, O.C. Heterocarpy in Dipterocypselinae (Asteraceae): Morphology, anatomy and systematic significance. Afr. J. Bot. 2022, 147, 263–274. [Google Scholar] [CrossRef]
- Grimbach, P. Vergleichende Anatomie verschiedenartiger Früchte und Samen bei derselben Spezies. Beib. Bot. Jahrb. 1913, 51, 1–52. [Google Scholar]
- Zhang, K.; Baskin, J.M.; Baskin, C.C.; Yang, X.; Huang, Z. Lack of divergence in seed ecology of two Amphicarpaea (Fabaceae) species disjunct between eastern Asia and eastern North America. Am. J. Bot. 2015, 102, 860–869. [Google Scholar] [CrossRef]
- Song, T.; Zhou, M.; Yuan, Y.; Yu, J.; Cai, H.; Li, J.; Chen, Y.; Bai, Y.; Zhou, G.; Cui, G. Chromosome-Scale reference genome of Amphicarpaea edgeworthii: A new resource for amphicarpic plants research and complex flowering pattern. Front. Plant Sci. 2021, 12, 770660. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, X.; Han, K.; Li, R.; Xu, G.; Han, Y.; Cui, F.; Fan, S.; Seim, I.; Fan, G.; et al. Insights into amphicarpy from the compact genome of the legume Amphicarpaea edgeworthii. Plant Biotechnol. J. 2021, 19, 952–965. [Google Scholar] [CrossRef]
- Evenari, M. Zur Keimungsökologie zweier Wüstenpflanzen. Mitt. Fl.-Soziol. Arbeitsgem. 1963, 10, 70–81. [Google Scholar]
- Koller, D.; Roth, N. Studies of the ecological and physiological significance of amphicarpy in Gymnarrhena micrantha (Compositae). Am. J. Bot. 1964, 51, 26–35. [Google Scholar] [CrossRef]
- Ruiz de Clavijo, E. The ecological significance of fruit heteromorphism in the amphicarpic species Catananche lutea (Asteraceae). Int. J. Plant Sci. 1995, 156, 824–833. [Google Scholar] [CrossRef]
- Ruiz de Clavijo, E.; Jimenez, M.J. The influence of achene type and plant density on growth and biomass allocation in the heterocarpic annual Catananche lutea (Asteraceae). Int. J. Plant Sci. 1998, 159, 637–647. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Seed germination in cleistogamous species: Theoretical considerations and a literature survey of experimental results. Seed Sci. Res. 2017, 27, 84–98. [Google Scholar] [CrossRef]
- Zeide, B. Reproductive behavior of plants in time. Am. Nat. 1978, 112, 636–639. [Google Scholar] [CrossRef]
- Venable, D.L. The evolutionary ecology of seed heteromorphism. Am. Nat. 1985, 126, 577–595. [Google Scholar] [CrossRef]
- Mandák, B. Seed heteromorphism and plant life cycle: A review of literature. Preslia 1997, 69, 129–159. [Google Scholar]
- Barker, N.P. A review and survey of basicarpy, geocarpy, and amphicarpy in the African and Madagascan flora. Ann. Mo. Bot. Gard. 2005, 92, 445–462. [Google Scholar]
- Sadech, A.; Guterman, H.; Gersani, M.; Ovadia, O. Plastic bet-hedging in an amphicarpic annual: An integrated strategy under variable conditions. Evol. Ecol. 2009, 23, 373–388. [Google Scholar] [CrossRef]
- Botschantzev, V.P.; Gorelova, T.G.; Kamelin, R.V. New records of Badchys flora. III. Bot. Zhurn. 1981, 66, 132–134. (In Russian) [Google Scholar]
- Hoffmann, O. Compositae. In Die Naturlichen Pflanzenfamilien; Engler, A., Prantl, K., Eds.; Engelmann: Leipzig, Germany, 1894; Volume IV, pp. 87–391. [Google Scholar]
- Zohary, M. Über einen neuen Fall von Amphikarpie bei Gymnarrhena micrantha Desf. Feddes Repert. 1930, 61, 95–97. [Google Scholar]
- Zamski, Y.E.; Ucko, O.; Koller, D. The mechanism of root contraction in Gymnarrhena micrantha, a desert plant. New Phytol. 1983, 95, 29–35. [Google Scholar] [CrossRef]
- Gutterman, Y. Seed dispersal by rain (ombrohydrochory) in some of the flowering desert plants in the deserts of Israel and the Sinai Peninsula. Mitt. Inst. Allg. Bot. Hambg. 1990, 23b, 841–852. [Google Scholar]
- Gutterman, Y. Strategies of seed dispersal and germination in plants inhabiting deserts. Bot. Rev. 1994, 60, 373–425. [Google Scholar] [CrossRef]
- Gutterman, Y. Maternal effects on seed during development. In Seeds. The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI Publishing: Oxford, UK; New York, NY, USA, 2000; pp. 59–84. [Google Scholar] [CrossRef]
- Bremer, K. Asteraceae. Cladistic & Classification; Timber Press: Portland, OR, USA, 1994; ISBN 10-0881922757. [Google Scholar]
- Jeffrey, C. Gymnarrheneae (Asteraceae). In The Families and Genera of Vascular Plants, Volume 8; Flowering Plants: Eudicots Asterales; Kadereit, J.W., Jeffrey, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 147–148. [Google Scholar] [CrossRef]
- Shamso, E.M.; Hosni, H.A.; Ahmed, D.; Shaltout, K. Achene characteristics of Some Taxa of Asteraceae from the Northwestern Mediterranean Coast of Egypt. Egypt. J. Bot. 2021, 61, 1–31. [Google Scholar] [CrossRef]
- van Rheede van Oudtshoorn, K.; van Rooyen, M.W. Dispersal Biology of Desert Plants; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar] [CrossRef]
- Prozina, M.N. Botanical Microtechniques; Vysschaya Schkola: Moscow, Russia, 1960. (In Russian) [Google Scholar]
- Corner, E.J.H. The Seeds of Dicotyledons: Volume 1; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1976. [Google Scholar]
- Dittrich, M. Die Bedeutung morphologischer und anatomischer Achänen-Merkmale für die Systematik der Tribus Echinopeae Cass. und Carlineae Cass. Boissiera 1996, 51, 1–102. [Google Scholar]
- Häffner, E. On the phylogeny of the subtribe Carduinae (tribe Cardueae, Compositae). In Englera; Botanischer Garten und Botanisches Museum: Berlin, Germany, 2000; Volume 21, pp. 1–209. ISBN 3-921800-46-3. [Google Scholar]
- Roque, N.; Keil, D.J.; Susanna, A. Illustrated glossary of Compositae. In Systematics, Evolution, and Biogeography of Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; IAPT: Vienna, Austria, 2009; pp. 781–806. ISBN 978-3-9501754-3-1. [Google Scholar]
- Hess, R. Vergleichende Untersuchungen über die Zwillingshaare er Kompositen. Bot. Jb. 1938, 68, 435–496. [Google Scholar]
- Grau, J. Die Testa der Mutisieae und ihre systematische Bedeutung. Mitt Bot. Staatssamml. München 1980, 16, 269–332. [Google Scholar]
- Stuessy, T.F.; Spooner, D.M. The adaptive and phylogenetic significance of receptacular bracts in the Compositae. Taxon 1988, 37, 114–126. [Google Scholar] [CrossRef]
- Sun, H.Z.; Lu, J.J.; Tan, D.Y.; Baskin, J.M.; Baskin, C.C. Dormancy and germination characteristics of the trimorphic achenes of Garhadiolus papposus (Asteraceae), an annual ephemeral from the Junggar Desert, China. Afr. J. Bot. 2009, 75, 537–545. [Google Scholar] [CrossRef]
- Sheldon, J.C.; Burrows, F.M. The dispersal effectiveness of the achene-pappus units of selected Compositae in steady winds with convection. New Phytol. 1973, 72, 665–675. [Google Scholar] [CrossRef]
- Marzinek, J.; Oliveira, D.M.T. Structure and ontogeny of the pericarp of six Eupatorieae (Asteraceae) with ecological and taxonomic considerations. An. Acad. Bras. Ciênc. 2010, 82, 279–291. [Google Scholar] [CrossRef]
- Talieff, W.I. von. Ueber das Hygroscopysche Gewebe des Compositenpappus. Proc. Soc. Nat. Sci. Imp. Kazan Univ. 1894, 27, 3–38. [Google Scholar]
- Fahn, A. Plant Anatomy, 3rd ed.; Pergamon Press Ltd.: Oxford, UK, 1982; ISBN 0-08-028030-7. [Google Scholar]
- Boesewinkel, F.D.; Bouman, F. The seed: Structure. In Embryology of Angiosperms; Johri, B.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 567–610. ISBN 10-3540127399. [Google Scholar]
- Werker, E. Seed Anatomy; Encyclopedia of Plant Anatomy Bd. 10, Teil 3; G. Borntraeger: Berlin, Germany; Stuttgart, Germany, 1997; ISBN 3-443-14024-6. [Google Scholar]
- Kozo-Poljanski, B.M. The Mimicry Problem in Botany; Voronezh St. University: Voronezh, Russia, 1939. (In Russian) [Google Scholar]
- Mani, M.S.; Saravanan, J.M. Pollination Ecology and Evolution in Compositae (Asteraceae); Oxford and IBH Publishing Co.: New Delhi, India; Calcutta, India, 1999; ISBN 1578080584. [Google Scholar]
- Evert, R.F. Esau’s Plant Anatomy. Meristems, Cells, and Tissues of Plant Body, Their Structure, Function, and Development, 3rd ed.; J. Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Del-Claro, K.; Guillermo-Ferreira, R.; Almeida, E.M.; Zardini, H.; Torezan-Silingardi, H.M. Ants visiting the post-floral secretions of pericarpial nectaries in Palicourea rigida (Rubiaceae) provide protection against leaf herbivores but not against seed parasites. Sociobiology 2013, 60, 217–221. [Google Scholar] [CrossRef]
- Sanz-Veiga, P.A.; Re Jorge, L.; Benitez-Vieyra, S.; Amorim, F.W. Pericarpial nectary-visiting ants do not provide fruit protection against pre-dispersal seed predators regardless of ant species composition and resource availability. PLoS ONE 2017, 12, e0188445. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | Aerial Achene | Subterranean Achene I | (?)Subterranean Achene II |
|---|---|---|---|
| Delamination of the fruit wall | - | - | + |
| Pericarp hairs | |||
| Density | densely arranged | rare | very rare |
| Length | 500–2000 | <500 | <500 |
| Seed coat in the middle of the seed | |||
| Length of exotesta cells | 70–135 | 110–170 | 102–170 |
| Thickness of anticlinal cell walls in exotesta | 8–12 | 9–18 | 12–18 |
| Orientation of crystals along the seed | ± | - | - |
| Endosperm in the middle of the seed | |||
| Cells length in the outer layer | 20–40 | 25–50 | 27–60 |
| Thickness of anticlinal cell walls in the outer layer | 1.5–6.0 | 1.5–4.0 (5.0) | 1.5–3.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kravtsova, T. Fruit Structure in Amphicarpic Annual Gymnarrhena micrantha (Asteraceae, Gymnarrheneae) in Relation to the Species Biology. Int. J. Plant Biol. 2023, 14, 397-417. https://doi.org/10.3390/ijpb14020032
Kravtsova T. Fruit Structure in Amphicarpic Annual Gymnarrhena micrantha (Asteraceae, Gymnarrheneae) in Relation to the Species Biology. International Journal of Plant Biology. 2023; 14(2):397-417. https://doi.org/10.3390/ijpb14020032
Chicago/Turabian StyleKravtsova, Tatyana. 2023. "Fruit Structure in Amphicarpic Annual Gymnarrhena micrantha (Asteraceae, Gymnarrheneae) in Relation to the Species Biology" International Journal of Plant Biology 14, no. 2: 397-417. https://doi.org/10.3390/ijpb14020032