

Production of Recombinant Biopharmaceuticals in Chlamydomonas reinhardtii
 , and
, and
Abstract
:1. Introduction
2. Chlamydomonas reinhardtii: The Microalga
3. Chlamydomonas reinhardtii Genomic Knowledge
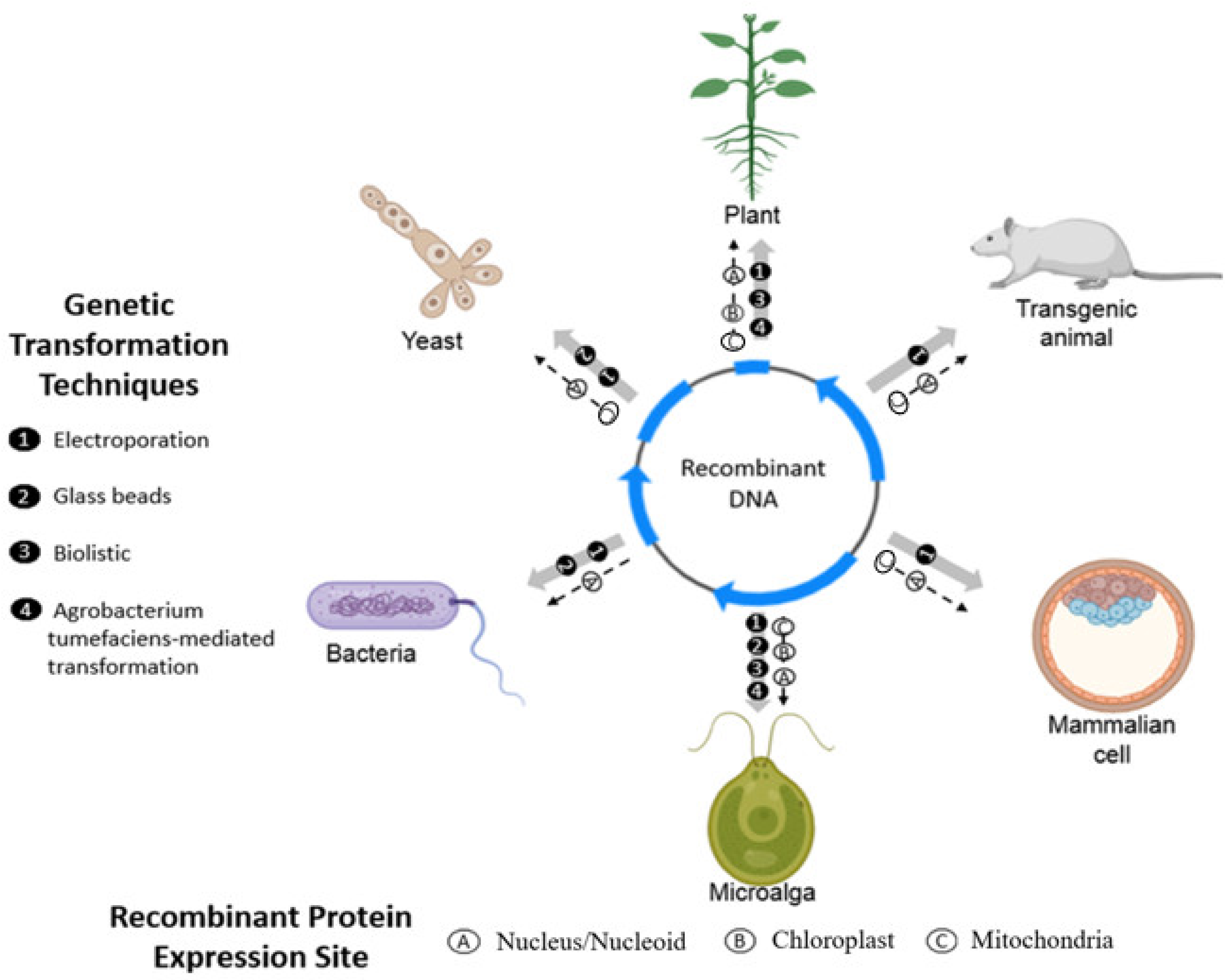
3.1. Transformation Methods in Chlamydomonas reinhardtii
3.2. Chloroplast Gene Regulation and Nuclear Gene Regulation

{kind=link}
| Heterologous Protein | *** (%) from Total Soluble Protein | Expression (mg/L) | Vector |
|---|---|---|---|
| 14FN3 (fourteenth human fibronectin type III domain) (ch) ** | 3 | UN * | psbA/D1-deficient strain (Rasala et al., 2010) [43] |
| VEGF (human vascular endothelial growth factor) (ch) ** | 2 | UN * | psbA/D1-deficient strain (Rasala et al., 2010) [43] |
| HMGB1 (high mobility group protein B1) (ch) ** | 2.5 | UN * | psbA/D1-deficient strain (Rasala et al., 2010) [43] |
| Endolysins—Pal and Cpl1 | 1 | 1.3 mg/g of cell dry weight | psbH insertion site/pSRSap vector/psaA promoter (Stoffels et al., 2017) [64] |
| Growth Hormone | UN * | 0.5 | psbH insertion site/pASapI vector/psaA and atpA promoter (Wannathong et al., 2016) [65] |
| D2 fibronectin-binding domain of Staphylococcus aureus fused with the cholera toxin B subunit (CTB) | 0.7 | 1.6 mg/g of cell dry weight | tscA insertion site/p463 and p464 vectors/rbcL and atpA promoters(Dreesen, Hamri and Fussenegger 2010) [66] |
| 14FN3 (fourteenth human fibronectin type III domain) (ch) ** | 0.21 | 0.04 | 16S atpA/photosynthetic strain (Rasala et al., 2010) [43] |
| Xylanase (nu) ** | 0.25 | UN * | Ble 2A expression system (Rasala et al., 2012) [67] |
| Immunotoxin (αCD22HCH23PE40) (ch) ** | 0.2–0.3 | UN * | psbA/D1-deficient strain (Tran et al., 2013) [42] |
| CD22 (Single chain antibody (scFv) directed against the CD22 cell surface antigen) (ch) ** | 0.6–0.7 | UN * | psbA/D1-deficient strain (Tran et al., 2013) [42] |
| CD22Gel (CD22-scFv genetically linked to gelonin, a ribosome-inactivating protein from G. multiflorum) (ch) ** | 0.2–0.3 | UN * | psbA/D1-deficient strain (Tran et al., 2013) [42] |
| CD22CH23Gel (CD22-scFv genetically fused to the hinge and constant domains of an IgG1 and to gelonin to create a chimeric protein homodimer) (ch) ** | 0.1–0.2 | UN * | psbA/D1-deficient strain (Tran et al., 2013) [42] |
| M-SAA (bovine mammary-associated serum amyloid) (ch) ** | 5 | UN * | psbA/D1-deficient strain (Manuell et al., 2007) [62] |
| Immunotoxin (αCD22HCH23PE40) (ch) ** | 3.0–4.0 | 0.3–0.4 | psbA/D1-deficient strain (Manuell et al., 2007) [62] |
| CtxB-Pfs25 [a chimeric protein consisting of the 25-kDa Plasmodium falciparum surface protein (Pfs25) fused to the subunit of the cholera toxin (CtxB)] (ch) ** | 0.09 | UN * | psbA/D1-deficient strain (Gregory et al., 2013) [54] |
| PFs25 (25-kDa Plasmodium falciparum surface protein) (ch) ** | 0.5 | 125 | psbA/D1-deficient strain (Gregory et al., 2013) [54] |
| PFs28 (28-kDa Plasmodium falciparum surface protein) (ch) ** | 0.2 | 50 | psbA/D1-deficient strain (Gregory et al., 2012) [68] |
| NCQ synthetic protein containing bioactive peptides that present different activities (antihypertensive, antioxidant, antimicrobial, opioid, and hypocholesterolemic) (ch) ** | 0.16–2.4 | UN * | tscA insertion site/rbcL or atpA promoters (Campos-Quevedo et al., 2013) [60] |
| Human glutamic acid decarboxylase 65 (Hgad65) (ch) ** | 0.25–0.3 | UN * | psbA exon V—5s rRNA intergenic region/pXW-GAD-His vector/rbcL promoter (Wang et al., 2008) [59] |
| A fusion protein comprising foot and mouth disease virus VP1 protein and the cholera toxin B subunit (CTBVP1) (ch) ** | 3–4 | ≈40 **** | chlL insertion site/pACTBVP1 vector/atpA promoter (Sun et al., 2003) [56] |
| A fusion protein comprising the D2 fibronectin binding domain of staphylococcus aureus and cholera toxin B subunit (CTBD2) (ch) ** | 0.7 | 260 | tscA insertion site/rbcL promoter (Dreesen, Hamri and Fussenegger 2010) [66] |
| Structural protein E2 from classical swine fever virus (ch) ** | 1.5–2.0 | UN * | chlL insertion site/p64E2 vector/atpA promoter (He et al., 2007) [58] |
| The human gene for erythropoietin (nu) ** | UN * | 0.1 | random insertion site/Expression cassette in the plasmid pEPO6/HSP70A/RBCS2 tandem promoter (Eichler-Stahlberg et al., 2009) [47] |
| HSV8-1sc and HSV8-scFv, a human IgA anti-herpesmonoclonal, large single-chain (Lsc) antibody and a single-chain fragment variable (scFv)antibody (ch) ** | 0.5 | UN * | p322-atpA-HSV8/or p322-rbcL-HSV8 vectors/atpA or rbcL promoters (Mayfield, Franklin, and Lerner 2003) [69] |
| Human tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) (ch) ** | 0.43–0.67 | UN * | chlL insertion site/p64TRAIL vector/atpA promoter (Yang et al., 2006) [57] |
| Full-length IgG1 human monoclonal antibody (mAb) against anthrax protective antigen 83 (PA83); potentially blocks the effects of anthrax toxin (ch) ** | UN * | 100 µg of purified protein per 1 g of dry algal biomass | psbA/D1-deficient strain (Tran et al., 2009) [41] |
| White spot syndrome virus VP28 protein (ch) ** | 0.2 to 20.9% total cellular protein (0.1 to 10.5%TSP) | UN * | psbA and upstream of 16S insertion sites/pBA155 or pSR229 vectors/psbA and atpA promoters (Surzycki et al., 2009) [70] |
| P. falciparum surface protein Pfs48/45 C-terminal domain (ch) ** | UN * | UN * | psbA/D1 deficient strain (Jones et al., 2013) [53] |
| Human Papillomavirus type 16 E7 protein, attenuated mutant (E7GGG) (ch) ** | 0.12 | UN * | Insertion site/pCG2 vector/psbD promoter (Demurtas et al., 2013) [61] |
| Angiotensin II fused to Hepatitis B virus capsid antigen (HbcAg) (nu) ** | 0.05 | UN * | Random insertion site/HbcAgI vector cassette/CaMV 35S promoter (Soria-Guerra et al., 2014) [71] |
4. Heterologous Protein Expression Systems
5. Chlamydomonas reinhardtii Potential for Pharmaceutical Proteins Biotechnology
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Walker, T.L.; Purton, S.; Becker, D.K.; Collet, C. Microalgae as bioreactors. Plant Cell Rep. 2005, 24, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Abdul Razack, S.; Duraiarasan, S.; Mani, V. Biosynthesis of silver nanoparticle and its application in cell wall disruption to release carbohydrate and lipid from C. vulgaris for biofuel production. Biotechnol. Rep. 2016, 11, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, S.R.; dos Santos, P.V.; Giraldi, L.A.; Zaiat, M.; Calijuri, M.C. Anaerobic phototrophic processes of hydrogen production by different strains of microalgae Chlamydomonas sp. FEMS Microbiol. Lett. 2018, 365, fny073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, G.; Yate, C.; Ramos, F.A.; Cala, M.P.; Restrepo, S.; Baena, S. Production of Polyunsaturated Fatty Acids and Lipids from Autotrophic, Mixotrophic and Heterotrophic cultivation of Galdieria sp. strain USBA-GBX-832. Sci. Rep. 2019, 9, 10791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ötleş, S.; Pire, R. Fatty acid composition of Chlorella and Spirulina microalgae species. J. AOAC Int. 2001, 84, 1708–1714. [Google Scholar] [CrossRef] [Green Version]
- Abou-Shanab, R.A.I.; Matter, I.A.; Kim, S.N.; Oh, Y.K.; Choi, J.; Jeon, B.H. Characterization and identification of lipid-producing microalgae species isolated from a freshwater lake. Biomass Bioenergy 2011, 35, 3079–3085. [Google Scholar] [CrossRef]
- Cezare-Gomes, E.A.; Mejia-da-Silva, L.d.C.; Pérez-Mora, L.S.; Matsudo, M.C.; Ferreira-Camargo, L.S.; Singh, A.K.; Carvalho, J.C.M. Potential of Microalgae Carotenoids for Industrial Application. Appl. Biochem. Biotechnol. 2019, 188, 602–634. [Google Scholar] [CrossRef]
- Wang, D.; Ning, K.; Li, J.; Hu, J.; Han, D.; Wang, H.; Zeng, X.; Jing, X.; Zhou, Q.; Su, X.; et al. Nannochloropsis Genomes Reveal Evolution of Microalgal Oleaginous Traits. PLoS Genet. 2014, 10, e1004094. [Google Scholar] [CrossRef]
- Wang, L.; Yang, L.; Wen, X.; Chen, Z.; Liang, Q.; Li, J.; Wang, W. Rapid and high-efficiency transformation of Chlamydomonas reinhardtii by square-wave electroporation. Biosci Rep. 2019, 39, BSR20181210. [Google Scholar] [CrossRef] [Green Version]
- Satjarak, A.; Paasch, A.E.; Graham, L.E.; Kim, E. Complete chloroplast genome sequence of phagomixotrophic green alga Cymbomonas tetramitiformis. Genome Announc. 2016, 4, e00551-16. [Google Scholar] [CrossRef]
- Scaife, M.A.; Nguyen, G.T.D.T.; Rico, J.; Lambert, D.; Helliwell, K.E.; Smith, A.G. Establishing Chlamydomonas reinhardtii as an industrial biotechnology host. Plant J. 2015, 82, 532–546. [Google Scholar] [CrossRef]
- Rasala, B.A.; Mayfield, S.P. Photosynthetic biomanufacturing in green algae; production of recombinant proteins for industrial, nutritional, and medical uses. Photosynth Res. 2015, 123, 227–239. [Google Scholar] [CrossRef]
- Harris, E.H.; Stern, D.B.; Witman, G.B. (Eds.) Chapter 8—Chlamydomonas in the Laboratory. In The Chlamydomonas Sourcebook; Academic Press: London, UK, 2009; Volume 2, pp. 241–302. [Google Scholar]
- Sasso, S.; Stibor, H.; Mittag, M.; Grossman, A. The natural history of model organisms from molecular manipulation of domesticated Chlamydomonas reinhardtii to survival in nature. Elife 2018, 7, e39233. [Google Scholar] [CrossRef]
- Ding, G.; Yaakob, Z.; Takriff, M.S.; Salihon, J. Biomass production and nutrients removal by a newly-isolated microalgal strain Chlamydomonas sp in palm oil mill effluent (POME). Int. J. Hydrogen Energy 2016, 41, 4888–4895. [Google Scholar] [CrossRef]
- Hong, J.W.; Jeong, J.; Kim, S.H.; Kim, S.; Yoon, H.S. Isolation of a Korean domestic microalga, Chlamydomonas reinhardtii KNUA021, and analysis of its biotechnological potential. J. Microbiol. Biotechnol. 2013, 23, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.K.; Cho, D.H.; Suh, S.I.; Lee, C.J.; Kim, H.S.; Suh, H.H. Isolation and Characterization of the Indigenous Microalgae Chlamydomonas reinhardtii K01 as a Potential Resource for Lipid Production and Genetic Modification. J. Life Sci. 2022, 32, 202–209. [Google Scholar]
- Contreras-Flores, C.; Peña-Castro, J.M.; Flores-Cotera, L.B.; Cañizares-Villanueva, R.O. Avances en el diseño conceptual de fotobiorreactores para el cultivo de microalgas. Interciencia 2003, 28, 450–456. [Google Scholar]
- Yan, N.; Fan, C.; Chen, Y.; Hu, Z. The Potential for Microalgae as Bioreactors to Produce Pharmaceuticals. Int. J. Mol. Sci. 2016, 17, 962. [Google Scholar] [CrossRef] [Green Version]
- Rosales-Mendoza, S.; Paz-Maldonado, L.M.T.; Soria-Guerra, R.E. Chlamydomonas reinhardtii as a viable platform for the production of recombinant proteins: Current status and perspectives. Plant Cell Rep. 2012, 31, 479–494. [Google Scholar] [CrossRef]
- Fields, F.J.; Ostrand, J.T.; Mayfield, S.P. Fed-batch mixotrophic cultivation of Chlamydomonas reinhardtii for high-density cultures. Algal Res. 2018, 33, 109–117. [Google Scholar] [CrossRef]
- Arias, C.A.D.; Matsudo, M.C.; Ferreira-Camargo, L.S.; Molino, J.V.D.; Mayfield, S.P.; Carvalho, J.C.M. Semicontinuous system for the production of recombinant mCherry protein in Chlamydomonas reinhardtii. Biotechnol. Prog. 2021, 37, e3101. [Google Scholar] [CrossRef] [PubMed]
- Ajayan, K.; Anjula, K.; Syamasurya, A.; Harilal, C. Energy efficient technology for enhanced growth and lipid production in Chlamydomonas reinhardtii through additional reflector coated LED photo-bioreactor. Biochem. Eng. J. 2019, 144, 81–88. [Google Scholar]
- Duman, E.; Kose, A.; Celik, Y.; Oncel, S. Design of a horizontal-dual bladed bioreactor for low shear stress to improve hydrodynamic responses in cell cultures: A pilot study in Chlamydomonas reinhardtii. Biochem. Eng. J. 2021, 169, 107970. [Google Scholar] [CrossRef]
- Baki, A.E.; Baroty, E. Optimization cultivtion of Chlamydomonas reinhardtii in a tubular photobioreactor (2000 Liter) for biomass and green bioenergy (biodiesel) production. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 1439–1457. [Google Scholar]
- Banerjee, S.; Ray, A.; Das, D. Optimization of Chlamydomonas reinhardtii cultivation with simultaneous CO2 sequestration and biofuels production in a biorefinery framework. Sci. Total Environ. 2021, 762, 143080. [Google Scholar] [CrossRef]
- Kong, Q.X.; Li, L.; Martinez, B.; Chen, P.; Ruan, R. Culture of Microalgae Chlamydomonas reinhardtii in Wastewater for Biomass Feedstock Production. Appl. Biochem. Biotechnol. 2010, 160, 9–18. [Google Scholar] [CrossRef]
- Gimpel, J.A.; Hyun, J.S.; Schoepp, N.G.; Mayfield, S.P. Production of recombinant proteins in microalgae at pilot greenhouse scale. Biotechnol. Bioeng. 2015, 112, 339–345. [Google Scholar] [CrossRef]
- Torres-Tiji, Y.; Fields, F.; Yang, Y.; Heredia, V.; Horn, S.J.; Keremane, S.R.; Jin, M.M.; Mayfield, S.P. Optimized production of a bioactive human recombinant protein from the microalgae Chlamydomonas reinhardtii grown at high density in a fed-batch bioreactor. Algal Res. 2022, 66, 102786. [Google Scholar] [CrossRef]
- Harris, E.H. Chlamydomonas as a model organism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 363–406. [Google Scholar] [CrossRef] [Green Version]
- Silflow, C.D.; Lefebvre, P.A. Assembly and motility of eukaryotic cilia and flagella. Lessons from Chlamydomonas reinhardtii. Plant Physiol. 2001, 127, 1500–1507. [Google Scholar] [CrossRef]
- Specht, E.; Miyake-Stoner, S.; Mayfield, S. Micro-algae come of age as a platform for recombinant protein production. Biotechnol. Lett. 2010, 32, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Tam, L.W.; Lefebvre, P.A. Cloning of flagellar genes in Chlamydomonas reinhardtii by DNA insertional mutagenesis. Genetics 1993, 135, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Cross, F.R.; Umen, J.G. The Chlamydomonas cell cycle. Plant J. 2015, 82, 370–392. [Google Scholar] [CrossRef]
- Franklin, S.; Ngo, B.; Efuet, E.; Mayfield, S.P. Development of a GFP reporter gene for Chlamydomonas reinhardtii chloroplast. Plant J. 2002, 30, 733–744. [Google Scholar] [CrossRef]
- Maul, J.E.; Lilly, J.W.; Cui, L.; dePamphilis, C.W.; Miller, W.; Harris, E.H.; Stern, D.B. The Chlamydomonas reinhardtii Plastid Chromosome: Islands of Genes in a Sea of Repeats. Plant Cell 2002, 14, 2659–2679. [Google Scholar] [CrossRef]
- Gallaher, S.D.; Fitz-Gibbon, S.D.; Strenkert, D.; Purvine, S.O.; Pellegrini, M.; Merchant, S.S. High-throughput sequencing of the chloroplast and mitochondrion of Chlamydomonas reinhardtii to generate improved de novo assemblies, analyze expression patterns and transcript speciation, and evaluate diversity among laboratory strains and wild isolate. Plant J. 2018, 93, 545–565. [Google Scholar] [CrossRef] [Green Version]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terri, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Mayfield, S.P. Protein disulfide isomerase as a regulator of chloroplast translational activation. Science 1997, 278, 1954–1957. [Google Scholar] [CrossRef]
- Schroda, M. The Chlamydomonas genome reveals its secrets: Chaperone genes and the potential roles of their gene products in the chloroplast. Photosynth. Res. 2004, 82, 221–240. [Google Scholar] [CrossRef]
- Tran, M.; Zhou, B.; Pettersson, P.L.; Pettersson, P.L.; Gonzalez, M.J.; Mayfield, S.P. Synthesis and assembly of a full-length human monoclonal antibody in algal chloroplasts. Biotechnol. Bioeng. 2009, 104, 663–673. [Google Scholar] [CrossRef]
- Tran, M.; Van, C.; Barrera, D.J.; Pettersson, P.L.; Peinado, C.D.; Bui, J.; Mayfield, S.P. Production of unique immunotoxin cancer therapeutics in algal chloroplasts. Proc. Natl. Acad. Sci. USA 2013, 110, E15–E22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasala, B.A.; Muto, M.; Lee, P.A.; Jager, M.; Cardoso, R.M.F.; Behnke, C.A.; Kirk, P.; Hokanson, C.A.; Crea, R.; Mendez, M.; et al. Production of therapeutic proteins in algae, analysis of expression of seven human proteins in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2010, 8, 719–733. [Google Scholar] [CrossRef] [Green Version]
- Engel, B.; Schaffer, M.; Cuellar, L.; Villa, E.; Plitzko, J.M.; Baumeister, W. Native architecture of the Chlamydomonas chloroplast revealed by in situ cryo-electron tomography. Elife 2015, 4, e04889. [Google Scholar] [CrossRef]
- Cutolo, E.A.; Mandalà, G.; Dall’osto, L.; Bassi, R. Harnessing the Algal Chloroplast for Heterologous Protein Production. Microorganisms 2022, 10, 743. [Google Scholar] [CrossRef] [PubMed]
- Doron, L.; Segal, N.; Shapira, M. Transgene Expression in Microalgae—From Tools to Applications. Front. Plant. Sci. 2016, 7, 505. [Google Scholar] [CrossRef]
- Eichler-Stahlberg, A.; Weisheit, W.; Ruecker, O.; Heitzer, M. Strategies to facilitate transgene expression in Chlamydomonas reinhardtii. Planta 2009, 229, 873–883. [Google Scholar] [CrossRef]
- Boynton, J.E.; Gillham, N.W.; Harris, E.H.; Hosler, J.P.; Johnson, A.M.; Jones, A.R.; Randolph-Anderson, D.L.; Robertson, D.; Klein, T.M.; Shark, K.B.; et al. Chloroplast transformation in Chlamydomonas with high velocity microprojectiles. Science 1988, 240, 1534–1538. [Google Scholar] [CrossRef]
- Dauvillée, D.; Delhaye, S.; Gruyer, S.; Slomianny, C.; Moretz, S.E.; d’Hulst, C.; Long, C.A.; Ball, S.G.; Tomavo, S. Engineering the chloroplast targeted malarial vaccine antigens in Chlamydomonas starch granules. PLoS ONE 2010, 5, e15424. [Google Scholar] [CrossRef]
- Lehtimäki, N.; Koskela, M.M.; Mulo, P. Posttranslational modifications of chloroplast proteins: An emerging field. Plant Physiol. 2015, 168, 768–775. [Google Scholar] [CrossRef] [Green Version]
- Heitzer, M.; Zschoernig, B. Construction of modular tandem expression vectors for the green alga Chlamydomonas reinhardtii using the Cre/lox-system. Biotechniques 2007, 43, 324–332. [Google Scholar] [CrossRef]
- Carrier, G.; Garnier, M.; Le Cunff, L.; Bougaran, G.; Probert, I.; De Vargas, C.; Corre, E.; Cadoret, J.-P.; Saint-Jean, B. Comparative transcriptome of wild type and selected strains of the microalgae Tisochrysis lutea provides insights into the genetic basis, lipid metabolism and the life cycle. PLoS ONE 2014, 9, e86889. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.S.; Luong, T.; Hannon, M.; Tran, M.; Gregory, J.A.; Shen, Z.; Briggs, S.P.; Mayfield, S.P. Heterologous expression of the C-terminal antigenic domain of the malaria vaccine candidate Pfs48/45 in the green algae Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2013, 97, 1987–1995. [Google Scholar] [CrossRef] [PubMed]
- Gregory, J.A.; Topol, A.B.; Doerner, D.Z.; Mayfield, S. Alga-produced cholera toxin-Pfs25 fusion proteins as oral vaccines. Appl. Environ. Microbiol. 2013, 79, 3917–3925. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Hu, H.; Gao, Y.; Xu, X.; Gao, H. Microalgae as platforms for production of recombinant proteins and valuable compounds: Progress and prospects. J. Ind. Microbiol. Biotechnol. 2011, 38, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Qian, K.; Su, N.; Chang, H.; Liu, J.; Shen, G. Foot-and-mouth disease virus VP1 protein fused with cholera toxin B subunit expressed in Chlamydomonas reinhardtii chloroplast. Biotechnol. Lett. 2003, 25, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Zonggi, Y.; Yinü, L.; Feng, C.; Dong, L.; Zhifang, Z.; Yanxin, L.; Dexian, Z.; Yong, W.; Guifang, S. Expression of human soluble TRAIL in Chlamydomonas reinhardtii chloroplast. Chin. Sci. Bull. 2006, 51, 1703–1709. [Google Scholar] [CrossRef]
- He, D.M.; Qian, K.X.; Shen, G.F.; Zhang, Z.-F.; Li, Y.-N.; Su, Z.-L.; Shao, H.-B. Recombination and expression of classical swine fever virus (CSFV) structural protein E2 gene in Chlamydomonas reinhardtii chroloplasts. Colloids Surf. B Biointerfaces 2007, 55, 26–30. [Google Scholar] [CrossRef]
- Wang, X.; Brandsma, M.; Tremblay, R.; Maxwell, D.; Jevnikar, A.M.; Huner, N.; Ma, S. A novel expression platform for the production of diabetes-associated autoantigen human glutamic acid decarboxylase (hGAD65). BMC Biotechnol. 2008, 8, 87. [Google Scholar] [CrossRef] [Green Version]
- Campos-Quevedo, N.; Rosales-Mendoza, S.; Paz-Maldonado, L.M.T.; Martínez-Salgado, L.; Guevara-Arauza, J.C.; Soria-Guerra, R.E. Production of milk-derived bioactive peptides as precursor chimeric proteins in chloroplasts of Chlamydomonas reinhardtii. Plant Cell Tissue Organ Cult. 2013, 113, 217–225. [Google Scholar] [CrossRef]
- Demurtas, O.C.; Massa, S.; Ferrante, P.; Venuti, A.; Franconi, R.; Giuliano, G. A Chlamydomonas-Derived Human Papillomavirus 16 E7 Vaccine Induces Specific Tumor Protection. PLoS ONE 2013, 8, e61473. [Google Scholar] [CrossRef] [Green Version]
- Manuell, A.L.; Beligni, M.V.; Elder, J.H.; Siefker, D.T.; Tran, M.; Weber, A.; McDonald, T.L.; Mayfield, S.P. Robust expression of a bioactive mammalian protein in Chlamydomonas chloroplast. Plant Biotechnol. J. 2007, 5, 402–412. [Google Scholar] [CrossRef]
- Ferreira-Camargo, L.S.; Tran, M.; Beld, J.; Burkart, M.; Mayfield, S.P. Selenocystamine improves protein accumulation in chloroplasts of eukaryotic green algae. AMB Express 2015, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Stoffels, L.; Taunt, H.N.; Charalambous, B.; Purton, S. Synthesis of bacteriophage lytic proteins against Streptococcus pneumoniae in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2017, 15, 1130–1140. [Google Scholar] [CrossRef] [Green Version]
- Wannathong, T.; Waterhouse, J.C.; Young, R.E.; Economou, C.K.; Purton, S. New tools for chloroplast genetic engineering allow the synthesis of human growth hormone in the green alga Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2016, 100, 5467–5477. [Google Scholar] [CrossRef]
- Dreesen, I.A.J.; Hamri, G.C.E.; Fussenegger, M. Heat-stable oral alga-based vaccine protects mice from Staphylococcus aureus infection. J. Biotechnol. 2010, 145, 273–280. [Google Scholar] [CrossRef]
- Rasala, B.A.; Lee, P.A.; Shen, Z.; Briggs, S.P.; Mendez, M.; Mayfield, S.P. Robust expression and secretion of Xylanase1 in Chlamydomonas reinhardtii by fusion to a selection gene and processing with the FMDV 2A peptide. PLoS ONE 2012, 7, e43349. [Google Scholar] [CrossRef] [Green Version]
- Gregory, J.A.; Li, F.; Tomosada, L.M.; Cox, C.J.; Topol, A.B.; Vinetz, J.M.; Mayfield, S. Algae-produced Pfs25 elicits antibodies that inhibit malaria transmission. PLoS ONE 2012, 7, e37179. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, S.P.; Franklin, S.E.; Lerner, R.A. Expression and assembly of a fully active antibody in algae. Proc. Natl. Acad. Sci. USA 2003, 100, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Surzycki, R.; Greenham, K.; Kitayama, K.; Dibal, F.; Wagner, R.; Rochaix, J.-D.; Ajam, T.; Surzycki, S. Factors effecting expression of vaccines in microalgae. Biologicals 2009, 37, 133–138. [Google Scholar] [CrossRef]
- Soria-Guerra, R.E.; Ramírez-Alonso, J.I.; Ibáñez-Salazar, A.; Govea-Alonso, D.O.; Paz-Maldonado, L.M.T.; Bañuelos-Hernández, B.; Korban, S.S.; Rosalez-Mendoza, S.S. Expression of an HBcAg-based antigen carrying angiotensin II in Chlamydomonas reinhardtii as a candidate hypertension vaccine. Plant Cell Tissue Organ Cult. 2014, 116, 133–139. [Google Scholar] [CrossRef]
- Cristóbal, J.; Matos, C.T.; Aurambout, J.P.; Manfredi, S.; Kavalov, B. Environmental sustainability assessment of bioeconomy value chains. Biomass Bioenergy 2016, 89, 159–171. [Google Scholar] [CrossRef]
- Golden, J.S.; Handfield, R.B.; Daystar, J.; McConnell, T.E. An Economic Impact Analysis of the US Biobased Products Industry: A Report to the Congress of the United States of America. Ind. Biotechnol. 2015, 11, 201–209. [Google Scholar] [CrossRef]
- Chacón-Lee, T.L.; González-Marino, G.E. Microalgae for “Healthy” Foods—Possibilities and Challenges. Compr. Rev. Food Sci. Food Saf. 2010, 9, 655–675. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Zhu, Z.; Qian, G.; Li, Y.; Meng, X.; Xue, J.; Chen, Q.; Sun, W.; Shi, Y. Inducing hairy roots by Agrobacterium rhizogenes-mediated transformation in tartary buckwheat (Fagopyrum tataricum). J. Vis Exp. 2020, 157, e60828. [Google Scholar] [CrossRef] [PubMed]
- Morin, N.; Czerwiec, Q.; Nicaud, J.M.; Neuvéglise, C.; Rossignol, T. Transforming Candida hispaniensis, a promising oleaginous and flavogenic yeast. Yeast 2020, 37, 348–355. [Google Scholar] [CrossRef]
- Gasser, B.; Mattanovich, D. Antibody production with yeasts and filamentous fungi: On the road to large scale. Biotechnol. Lett. 2007, 201, 201–212. [Google Scholar] [CrossRef]
- Aizemberg, R.; Terrazas, W.; Ferreira-Dias, S.; Valentini, S.R.; Gattás, E.A.L. Optimal conditions for Biomass and Recombinant Glycerol Kinase Production Using the Yeast Pichia pastoris. Food Technol. Biotechnol. 2011, 49, 329–335. [Google Scholar]
- Potvin, G.; Zhang, Z. Strategies for high-level recombinant protein expression in transgenic microalgae: A review. Biotechnol. Adv. 2010, 28, 910–918. [Google Scholar] [CrossRef]
- Fingerut, E.; Gutter, B.; Meir, R.; Heliahoo, D.; Pitcovski, J. Vaccine and adjuvant activity of recombinant subunit B of E. coli enterotoxin produced in yeast. Vaccine 2005, 23, 4685–4696. [Google Scholar] [CrossRef]
- Serrano, A.G. Estudio de la Producción Heteróloga de una Lipasa del Hongo Rhizopus Oryzae en la Levadura Metilotrófica Pichia Pastoris. Ph.D. Thesis, Universitat Autònoma de Barcelona, Barcelona, Spain, 2001. [Google Scholar]
- Walsh, G. Biopharmaceutical benchmarks 2014. Nat. Biotechnol. 2014, 32, 992–1000. [Google Scholar] [CrossRef]
- Schirrmann, T.; Al-Halabi, L.; Dübel, S.; Hust, M. Production systems for recombinant antibodies. Front. Biosci. 2008, 13, 4576–4594. [Google Scholar] [CrossRef] [Green Version]
- Lerouge, P.; Cabanes-Macheteau, M.; Rayon, C.; Fischette-Lainé, A.C.; Gomord, V.; Faye, L. N-glycoprotein biosynthesis in plants: Recent developments and future trends. Plant Mol. Biol. 1998, 38, 31–48. [Google Scholar] [CrossRef]
- Park, J.J.; Wang, H.; Gargouri, M.; Deshpande, R.R.; Skepper, J.N.; Holguin, F.O.; Juergens, M.T.; Shachar-Hill, Y.; Hicks, L.M.; Gang, D.R. The response of Chlamydomonas reinhardtii to nitrogen deprivation: A systems biology analysis. Plant J. 2015, 81, 611–624. [Google Scholar] [CrossRef]
- Ariás Gomez, C.L. Caracterización de la Ruta de N-Glicosilación de Proteínas en el Sistema de Endomembranas del Alga Verde Chlamydomonas reinhardtii: Distribución al Cloroplasto de N-Glicoproteínas. Ph.D. Thesis, Universidad Autónoma de Madrid, Madrid, Spain, 2012. [Google Scholar]
- Rybicki, E.P. Plant-produced vaccines: Promise and reality. Drug Discov. Today 2009, 14, 16–24. [Google Scholar] [CrossRef]
- Kirk, D.D.; McIntosh, K.; Walmsley, A.M.; Peterson, R.K.D. Risk analysis for plant-made vaccines. Transgenic Res. 2005, 14, 449–462. [Google Scholar] [CrossRef]
- Busconi, M.; Baldi, G.; Lorenzoni, C.; Fogher, C. Gene flow from transgenic rice to red rice (Oryza sativa L.) in the field. Plant Biol. 2014, 16, 22–27. [Google Scholar] [CrossRef]
- Yao, J.; Weng, Y.; Dickey, A.; Wang, K.Y. Plants as factories for human pharmaceuticals: Applications and challenges. Int. J. Mol. Sci. 2015, 16, 28549–28565. [Google Scholar] [CrossRef]
- Mayfield, S.P.; Franklin, S.E. Expression of human antibodies in eukaryotic micro-algae. Vaccine 2005, 23, 1828–1832. [Google Scholar] [CrossRef]
- Ren, H.; Dong, B.; Fan, Z.; Meng, D. Prokaryotic expression and purification of Chlamydomonas reinhardtii intraflagellar transport protein 46(IFT46) and preparation of polyclonal antibody. Shengwu Gongcheng Xuebao/Chin. J. Biotechnol. 2016, 12, 1124–1132. [Google Scholar] [CrossRef]
- Lara, A. Produccio n de proteinas recombinantes en Escherichia coli. Rev. Mex Ing. Quim. 2011, 10, 209–223. [Google Scholar]
- Preiss, S.; Schrader, S.; Johanningmeier, U. Rapid, ATP-dependent degradation of a truncated D1 protein in the chloroplast. Eur. J. Biochem. 2001, 268, 4562–4569. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arias, C.A.D.; Oliveira, C.F.M.d.; Molino, J.V.D.; Ferreira-Camargo, L.S.; Matsudo, M.C.; Carvalho, J.C.M.d. Production of Recombinant Biopharmaceuticals in Chlamydomonas reinhardtii. Int. J. Plant Biol. 2023, 14, 39-52. https://doi.org/10.3390/ijpb14010004
Arias CAD, Oliveira CFMd, Molino JVD, Ferreira-Camargo LS, Matsudo MC, Carvalho JCMd. Production of Recombinant Biopharmaceuticals in Chlamydomonas reinhardtii. International Journal of Plant Biology. 2023; 14(1):39-52. https://doi.org/10.3390/ijpb14010004
Chicago/Turabian StyleArias, Cesar Andres Diaz, Caroline Frere Martiniuc de Oliveira, João Vitor Dutra Molino, Livia Seno Ferreira-Camargo, Marcelo Chuei Matsudo, and João Carlos Monteiro de Carvalho. 2023. "Production of Recombinant Biopharmaceuticals in Chlamydomonas reinhardtii" International Journal of Plant Biology 14, no. 1: 39-52. https://doi.org/10.3390/ijpb14010004