
The Na+/H+ Exchanger NHX1 Controls H+ Accumulation in the Vacuole to Influence Sepal Color in Hydrangea macrophylla

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sequencing
2.2. RNA Extraction and cDNA Synthesis
2.3. Full-Length Amplification of the HmNHX1 Coding DNA Sequence (CDS)
2.4. Gel Extraction
2.5. Bioinformatic Analysis
2.6. Tissue-Specific Expression Analysis of HmNHX1
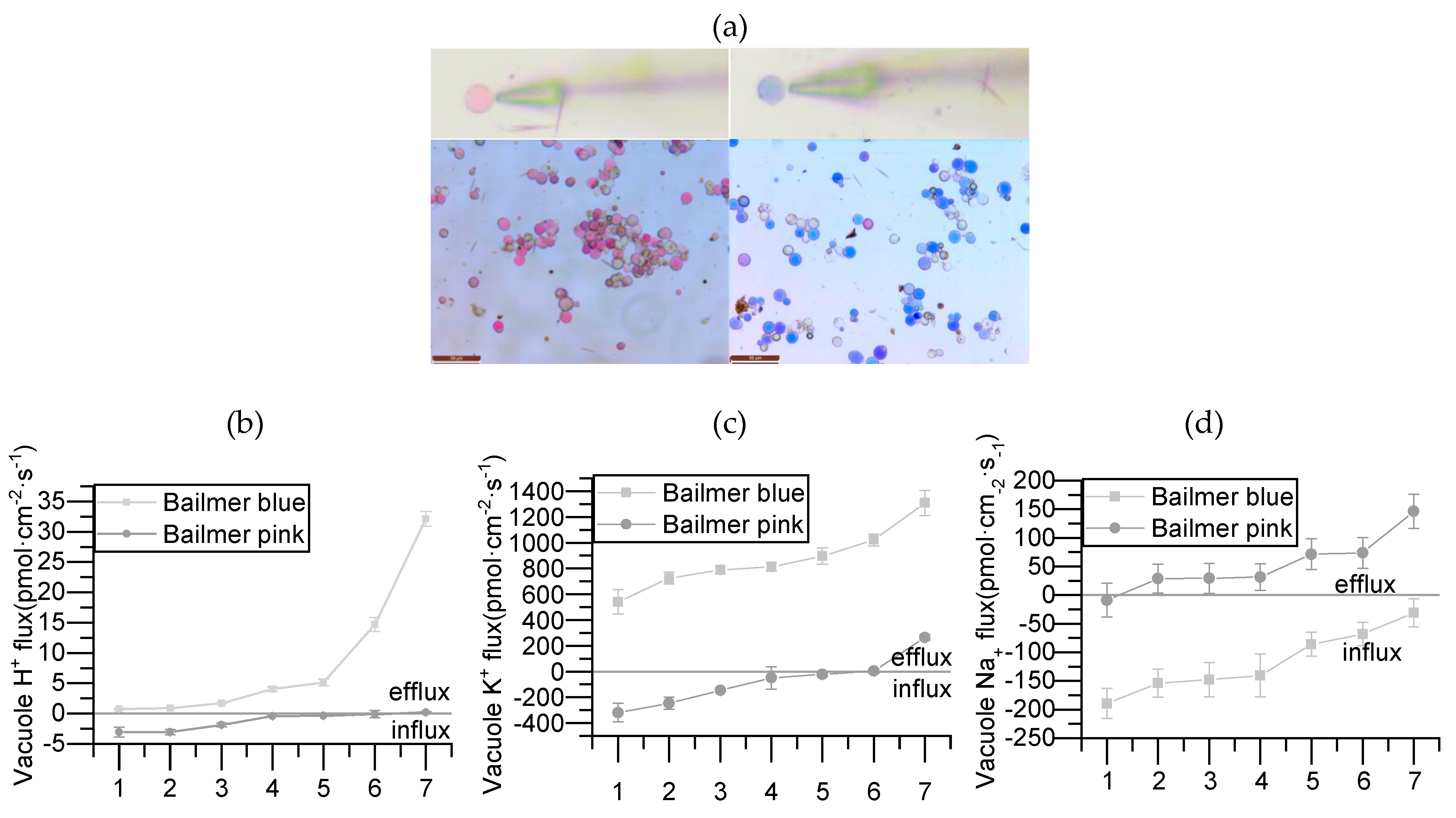
2.7. Ion Flux Measurements of Bailmer Vacuoles
3. Results
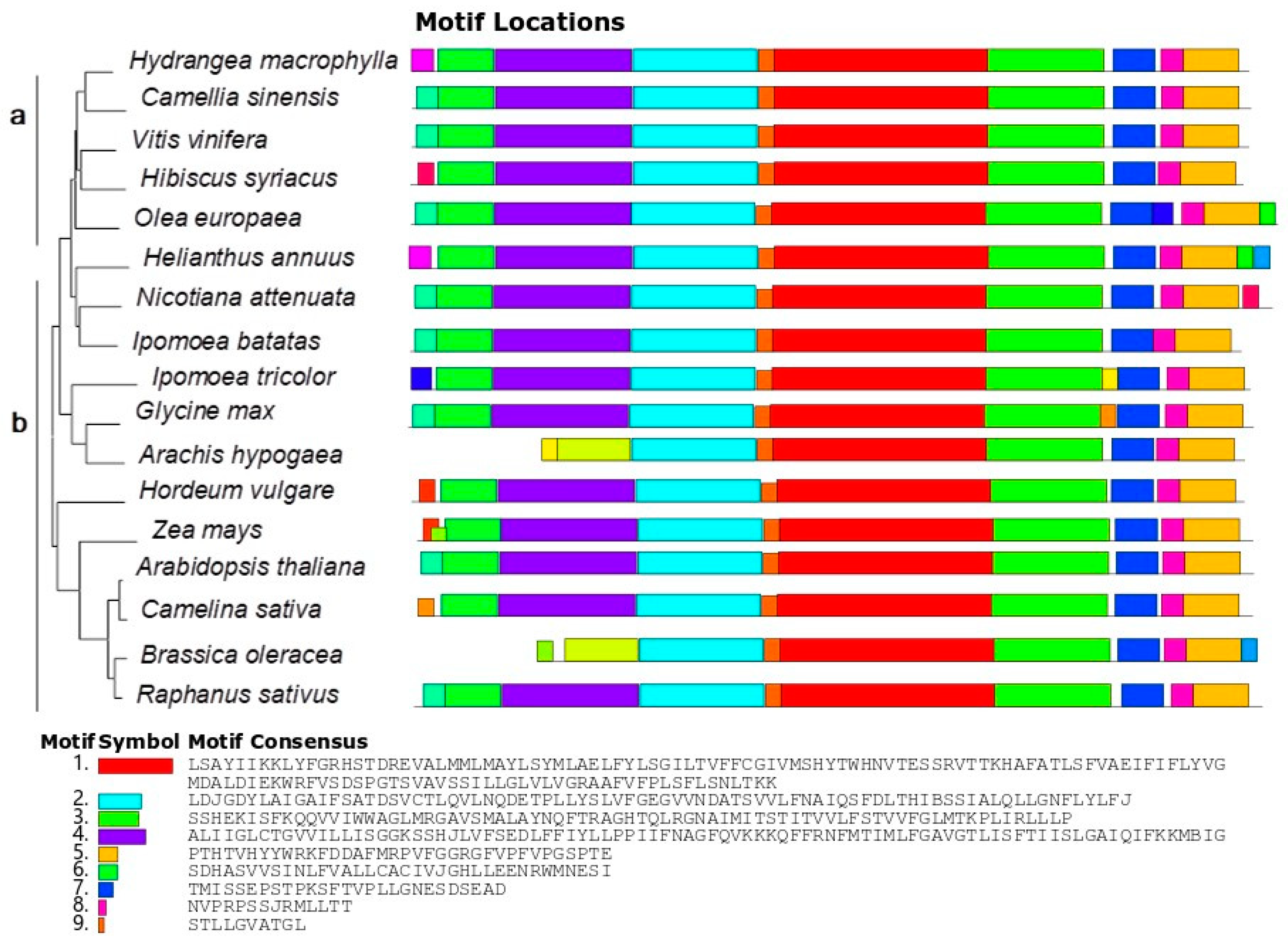
3.1. HmNHX1 Bioinformatic Analysis
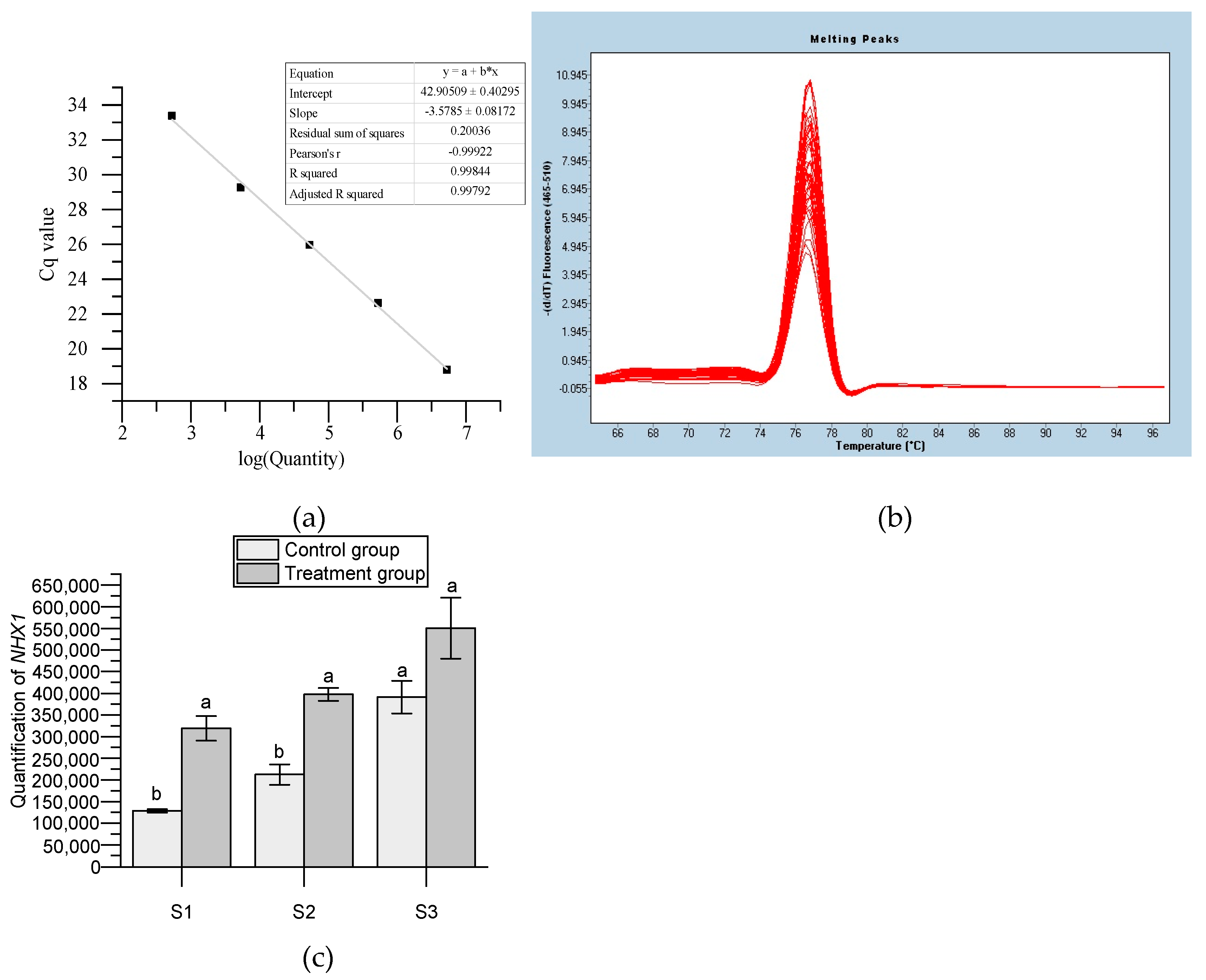
3.2. Expression Analysis of HmNHX1
3.3. Relationship between NHX1 and Ion Flux Measurements
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galopin, G.; Codarin, S.; Viemont, J.D.; Morel, P. Architectural Development of Inflorescence in Hydrangea macrophylla cv. Hermann Dienemann. Hortscience 2008, 43, 361–365. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Aoki, D.; Fukushima, K.; Yoshida, K. Direct Mapping of Hydrangea Blue-complex in Sepal Tissues of Hydrangea macrophylla. Sci. Rep. 2019, 9, 5450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, H.D. Curious chemistry guides hydrangea colors. Am. Sci. 2014, 102, 444. [Google Scholar] [CrossRef]
- Yoshida, K.; Toyama-Kato, Y.; Kameda, K.; Tadao, K. Sepal Color Variation of Hydrangea macrophylla and Vacuolar pH Measured with a Proton-Selective Microelectrode. Plant Cell Physiol. 2003, 44, 262–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Oyama, K.; Yoshida, K. Direct Observation of Hydrangea Blue-Complex Composed of 3-O-Glucosyldelphinidin, Al3+ and 5-O-Acylquinic Acid by ESI-Mass Spectrometry. Molecules 2018, 23, 1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukada-Tanaka, S.; Inagaki, Y.; Yamaguchi, T.; Saito, N.; Iida, S. Colour-Enhancing Protein in Blue Petals. Spectacular Morning Glory Blooms Rely on a Behind-the-Scenes Proton Exchanger. Nature 2000, 407, 581. [Google Scholar] [CrossRef]
- Quattrocchio, F.; Verweij, W.; Kroon, A.; Spelt, C.; Mol, J.; Koes, R. PH4 of petunia is an R2R3 MYB protein that activates vacuolar acidification through interactions with Basic-Helix-Loop-Helix transcription factors of the anthocyanin pathway. Plant Cell 2006, 18, 1274–1291. [Google Scholar] [CrossRef] [Green Version]
- Ryoji, T.; Noriko, Y.; Nobuyuki, Y. A MYB transcription factor controls flower color in soybean. J. Hered. 2013, 104, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Asen, S.; Stewart, R.N.; Norris, K.H. Co-Pigmentation of Anthocyanins in Plant Tissues and Its Effect on Color. Phytochemistry 1972, 11, 1139–1144. [Google Scholar] [CrossRef]
- Asen, S.; Stewart, R.N.; Norris, K.H. Anthocyanin, Flavonol Copigments, and pH Responsible for Larkspur Flower Color. Phytochemistry 1975, 14, 2677–2682. [Google Scholar] [CrossRef]
- Verweij, W.; Spelt, C.; Di Sansebastiano, G.P.; Vermeer, J.; Reale, L.; Ferranti, F.; Koes, R.; Quattrocchio, F. An H+ P-ATPase on the Tonoplast Determines Vacuolar pH and Flower Colour. Nat. Cell Biol. 2008, 10, 1456–1462. [Google Scholar] [CrossRef]
- Brett, C.L.; Donowitz, M.; Rao, R. Evolutionary Origins of Eukaryotic Sodium/Proton Exchangers. Am. J. Physiol. Cell Physiol. 2005, 288, C223–C239. [Google Scholar] [CrossRef] [Green Version]
- Darley, C.P.; Van Wuytswinkel, O.C.M.; Karel, V.; Mager, W.H.; De Boer, A.H. Arabidopsis thaliana and Saccharomyces cerevisiae NHX1 genes encode amiloride sensitive electroneutral Na+/H+ exchangers. Biochem. J. 2000, 351, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.H.; Zhao, X.Y.; Li, W.; Luo, Y.; Zhang, X.S. Overexpression of AeNHX1, a root-specific vacuolar Na+/H+ antiporter from Agropyron elongatum, confers salt tolerance to Arabidopsis and Festuca plants. Plant Cell Rep. 2007, 26, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Borrero, V.B.; Leidi, E.O.; Andrés, Z.; Rubio, L.; De Luca, A.; Fernández, J.A.; Cubero, B.; Pardo, J.M. Ion Exchangers NHX1 and NHX2 Mediate Active Potassium Uptake into Vacuoles to Regulate Cell Turgor and Stomatal Function in Arabidopsis. Plant Cell 2012, 24, 1127–1142. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Kondo, T.; Okazaki, Y.; Katou, K. Cause of blue petal colour. Nature 1995, 373, 291. [Google Scholar] [CrossRef]
- Yoshida, K.; Mori, M.; Kondo, T. Blue Flower Color Development by Anthocyanins: From Chemical Structure to cell. Nat. Prod. Rep. 2009, 26, 884–915. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Bao, H.; Guo, J.; Jia, W.; Tai, F.; Nie, L.; Jiang, P.; Feng, J.; Lv, S.; Li, Y. Na+/H+ Exchanger 1 Participates in Tobacco Disease Defence Against Phytophthora Parasitica Var. Nicotianae by Affecting Vacuolar pH and Priming the Antioxidative System. J. Exp. Bot. 2014, 65, 6107–6122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koes, R.E.; Quattrocchio, F.; Mol, J. The flavonoid biosynthetic pathway in plants: Function and evolution. BioEssays 1994, 16, 123–132. [Google Scholar] [CrossRef]
- Holton, T.A.; Cornish, E.C. Genetics and Biochemistry of Anthocyanin Biosynthesis. Plant Cell 1995, 7, 1071–1083. [Google Scholar] [CrossRef]
- Alfenito, M.R.; Souer, E.; Goodman, C.D.; Buell, R.; Mol, J.; Kose, R.; Walbot, V. Functional Complementation of Anthocyanin Sequestration in the Vacuole by Widely Divergent Glutathione S-transferases. Plant Cell 1998, 10, 1135–1149. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Dong, X.; Xue, C.; Liu, Z.; Cao, F. Exploring the Molecular Mechanism of Blue Flower Color Formation in Hydrangea macrophylla cv. “Forever Summer”. Front. Plant Sci. 2021, 12, 585665. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Tamura, H.; Kawai, T.; Hoshino, T.; Kondo, T. Chemistry of metalloanthocyanins. Ann. N. Y. Acad. Sci. 1987, 471, 155–173. [Google Scholar] [CrossRef]
- Griesbach, R.J. Correlation of pH and Light Intensity on Flower Color in Potted Eustoma grandiflorum Grise. HortScience 1992, 27, 817–818. [Google Scholar] [CrossRef]
- Arkin, I.T.; Xu, H.F.; Jensen, M.Ø.; Arbely, E.; Bennett, E.R.; Bowers, K.J.; Chow, E.; Dror, R.O.; Eastwood, M.P.; Flitman-Tene, R.; et al. Mechanism of Na+/H+ antiporting. Science 2007, 317, 799–803. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Fukada-Tanaka, S.; Inagaki, Y.; Saito, N.; Yonekura-Sakakibara, K.; Tanaka, Y.; Kusumi, T.; Iida, S. Genes Encoding the Vacuolar Na+/H+ Exchanger and Flower Coloration. Plant Cell Physiol. 2001, 42, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Matthew, G.; Wendy, S.; Mark, T.; Stephen, D.T. Simultaneous Flux and Current Measurement from Single Plant Protoplasts Reveals a Strong Link between K+ Fluxes and Current, but No Link between Ca2+ Fluxes and Current. Plant J. 2006, 46, 134–144. [Google Scholar] [CrossRef]
- Rauf, M.; Shahzad, K.; Ali, R.; Ahmad, M.; Habib, I.; Mansoor, S.; Berkowitz, G.A.; Saeed, N.A. Cloning and Characterization of Na+/H+ Antiporter (LfNHX1) Gene from a Halophyte Grass Leptochloa fusca for Drought and Salt Tolerance. Mol. Biol. Rep. 2014, 41, 1669–1682. [Google Scholar] [CrossRef]
- Ma, J.F.; Ryan, P.R.; Delhaize, E. Aluminium Tolerance in Plants and the Complexing Role of organic acids. Trends Plant Sci. 2001, 6, 273–278. [Google Scholar] [CrossRef]
- Negishi, T.; Oshima, K.; Hattori, M.; Kanai, M.; Mano, S.; Nishimura, M.; Yoshida, K. Tonoplast- and Plasma Membrane-Localized Aquaporin-Family Transporters in Blue Hydrangea Sepals of Aluminum Hyperaccumulating Plant. PLoS ONE 2012, 7, e43189. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Yuan, S.; Qi, H.; Chu, Z.; Liu, C. The Na+/H+ Exchanger NHX1 Controls H+ Accumulation in the Vacuole to Influence Sepal Color in Hydrangea macrophylla. Int. J. Plant Biol. 2023, 14, 266-275. https://doi.org/10.3390/ijpb14010022
Zhang G, Yuan S, Qi H, Chu Z, Liu C. The Na+/H+ Exchanger NHX1 Controls H+ Accumulation in the Vacuole to Influence Sepal Color in Hydrangea macrophylla. International Journal of Plant Biology. 2023; 14(1):266-275. https://doi.org/10.3390/ijpb14010022
Chicago/Turabian StyleZhang, Gaitian, Suxia Yuan, Hui Qi, Zhiyun Chu, and Chun Liu. 2023. "The Na+/H+ Exchanger NHX1 Controls H+ Accumulation in the Vacuole to Influence Sepal Color in Hydrangea macrophylla" International Journal of Plant Biology 14, no. 1: 266-275. https://doi.org/10.3390/ijpb14010022