
Identification of CoDREB Genes for Drought and Cold Tolerance in Camellia oleifera

1
Key Laboratory of Cultivation and Protection for Non-Wood Forest Trees of Ministry of Education, The Key Laboratory of Non-Wood Forest Products of Forestry Ministry, Central South University of Forestry and Technology, Changsha 410004, China
2
Engineering Technology Research Center of Southern Hilly and Mountainous Ecological Non-Wood Forest Industry of Hunan Province, Changsha 410004, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Plant Biol. 2023, 14(1), 228-241; https://doi.org/10.3390/ijpb14010019
Submission received: 15 January 2023
/
Revised: 7 February 2023
/
Accepted: 9 February 2023
/
Published: 13 February 2023
(This article belongs to the Topic Tolerance to Drought and Salt Stress in Plants)
Abstract
:DREB is a plant-specific transcription factor family that plays a pleiotropic regulatory role in response to abiotic stresses such as drought and cold. In this study, we identified 51 DREB genes of Camellia oleifera. These CoDREBs ranged from 88 to 518 amino acids (average/median 259/237 aa). The predicted molecular weights (MW) of the CoDREB proteins ranged from 9.7 kDa to 59.6 kDa, and the isoelectric points (pI) ranged from 4.62 to 10.44. A gene structure analysis showed that 43/51 (84.3%) CoDREBs were intronless, and the number of exons varied from one to three. Then, we focused on the response of CoDREB genes in terms of plant drought and cold acclimation. Under short-/long-term drought stress, CoDREB1.2/4.1/4.4/4.8/4.12/4.15/5.1/5.3/5.5/6.2 have different regulations in response to long-term drought response, and CoDREB1.4/2.5/4.6/4.1/6.3/6.5 specifically in the short term. Additionally, in response to mild/severe drought and followed by recovery, we found that CoDREBs may be involved in a complex drought-responsive regulatory network. Under cold stress, CoDREB5.2 and CoDREB6.5 are significantly up-regulated, and CoDREB may participate in the regulation of the low-temperature response of C. oleifera.
1. Introduction
Agricultural security and sustainability are jeopardized by a variety of potential risks, such as global environment/climate changes, poverty, the Russo-Ukrainian War (interlinked global crises and conflicts), and even the COVID-19 pandemic. To maintain high levels of production in the agricultural sector, plant breeders/researchers are focusing on predictable potential hazards, such as environmental changes that cause abiotic stresses (unfavorable growth conditions) during plant growth, which inevitably affect crop yield every year. During long-term natural selection pressures, sessile plants have evolved a complex set of molecular and physiological mechanisms to cope with abiotic/biotic stresses and breeders expanding numerous tools and mining molecular resources to develop new improved crop varieties [1,2]. Therefore, it is critical to understand underlying mechanisms regarding desired plant breeding goals.
Camellia oleifera Abel. (commonly known as tea oil, Theaceae) is one of four major woody oil plants that are widely distributed in subtropical mountain areas of the Yangtze River basin and South China, with elevations ranging from about 200 to 2000 m [3,4]. The plantation area of C. oleifera spans approximately 4.4 million hectares, with an annual output of over 2.6 million tons of seeds and, as a result, oil yields of more than 0.65 million tons [5]. Precipitation in the main tea-oil-producing areas is mostly concentrated from March to June, accounting for 60% to 70% of the total precipitation during the year; precipitation from July to September (the annual dry season in South China) is relatively low, accounting for about 20% of the annual precipitation, and is susceptible to high temperatures and drought, while the critical period for tea oil synthesis/accumulation is from July to August [6]. Accordingly, despite being a drought-resistant tree, (seasonal) drought stress is regarded as the major threat to C. oleifera growth [7], causing wilting; decreases in photosynthesis, oil contents, flower buds, and fruits setting; even the death of drought-sensitive cultivars that is mainly induced by cell death caused by reactive oxygen species (ROS) accumulation [8,9]. Plants have developed drought-resistance strategies that involve a series of morphological, biochemical, physiological, and molecular factors [10,11]. Similar to tea and other crops, tea oil also modifies its leaf structure based on the degree of drought in order to meet the needs for survival and growth [12], increasing peroxidase (POD) activity and malonaldehyde (MDA) content [7]. Moreover, due to recent advances in high-throughput sequencing, a transcriptomic analysis of C. oleifera can be used to identify core differentially expressed genes (DEGs) in response to various degrees of drought stress [5,7,13]. However, research on C. oleifera-specific drought-resistant regulatory networks/mechanisms remains far from being used in molecular assistance breeding or simply providing genetic resources to speed up the breeding process.
Dehydration-responsive element-binding (DREB) proteins bind to specific DNA motifs (DRE/CRT element, A/GCCGAC), and are found to be important in activating the expression of target stress-inducible genes [14,15,16]. As reported, the overexpression of DREB2 gene can improve the drought tolerance of the Arabidopsis [14]. Additionally, DREB1 is rapidly and transiently induced by cold stress, and the overexpression of DREB1 leads to increased stress tolerance in rice and Arabidopsis [14,17]. To date, DREB has been widely identified in the plant kingdom and the role of DREB has frequently been reviewed. Gene discovery leads to improved drought stress tolerance in plants via gene transfer. The present study comprehensively integrates the latest C. oleifera transcriptome studies on various levels of drought stress (both short-/long-term and severe). Simultaneously, as C. oleifera is perennial non-wood economic species and a cold-area-adaptive subspecies, cold-responsive DREB is an important resource for future molecule assistance breeding. A publicly available RNA-seq dataset related to C. oleifera was also analyzed to identify potential DREBs, and preliminarily characterized DREBs may be involved in combined drought and cold stress responses. We identified 18 DREBs involved in short-/long-term drought responses, both together and each individually, and cross-validated the function of genes involved in mild/severe drought stresses. CoDREB5.2 and CoDREB6.5 were identified as key genes for cold adaptation. Finally, the combined results suggest that CoDREB5.2 and CoDREB6.5 may be involved in both drought and cold stress adaptation.
2. Materials and Methods
2.1. Homologs Sequence Identification of DREB Genes in C. oleifera
To aid in the comprehensive identification and analysis of DREB gene family in C. oleifera, sequences of putative DREB genes were obtained from National Center for Biotechnological Information (NCBI, https://www.ncbi.nlm.nih.gov/), accessed on 14 March 2022. GenBank and The Arabidopsis International Resource (TAIR, https://www.arabidopsis.org/), accessed on 14 March 2022. The chromosome-scale reference genome of diploid progenitor C. oleifera “Nanyongensis” (CON, 2n = 2x = 30) was downloaded from Zenodo.org released on 9 December 2021 (https://zenodo.org/record/5768785#.Y7b3H3ZBwQ8, accessed on 14 March 2022) [18]. CoDREB genes were identified with a released procedure: CoDREB genes were first searched with TBtools “BLAST GUI Wrapper” [19] with AtDREBs to obtain candidate C. oleifera DREB-family members; the HMM (Hidden Markov Model) profile PF00847 (AP2 domain) was retrieved from the Pfam database (https://pfam.sanger.ac.uk/), accessed on 14 March 2022. to identify putative DREB genes with highly conserved amino acid residues 14th Valine (V) and 19th glutamic acid/valine/leucine/alanine (E/V/L/A) [20]. The molecular weight (MW) and isoelectric point (pI) were predicted using the ProtParam tool (https://web.expasy.org/protparam/), accessed on 14 March 2022.
2.2. Phylogenetic Analysis of DREB Proteins and Nomenclature
CoDREB and its homologs were imported into MEGAX (version 10.2.6, https://www.megasoftware.net), accessed on 25 March 2022 and the sequence alignment was performed using the default “ClustalW” settings [21]. Phylogenetic trees were then constructed using the “Neighbor-joining” algorithm with default parameters, and the ‘.nwk’ phylogenetic tree was made using iTol online (https://itol.embl.de), accessed on 25 March 2022 [22]. According to Arabidopsis A1–A6 nomenclature, CoDREB genes have been designated based on their location on the phylogenetic tree.
2.3. Gene Structure and MEME Conserved Motif Analysis
Conserved motifs of CoDREB proteins were predicted using the MEME suite version 5.4.1 (https://meme-suite.org/meme/index.html), accessed on 25 March 2022 [23] with default parameters except the “motifs should find” was set to 10. The coding sequences (CDS) and the structure of all genes were graphically displayed with TBtools function “Gene Structure View” [19]. The chromosome-scale reference genome annotated structure information “gff” file of C. oleifera was downloaded from Zenodo.org released on 9 December 2021 (https://zenodo.org/record/5768785#.Y7b3H3ZBwQ8, accessed on 14 March 2022) [18].
2.4. RNA-Seq Data
Raw data of RNA-seq were downloaded from The European Nucleotide Archive (ENA, https://www.ebi.ac.uk/ena/browser/home), accessed on 29 October 2022 [24] through using “IBM aspera” (https://www.ibm.com/products/aspera), accessed on 29 October 2022 that was installed through “Bioconda” [25]. Short-/long-term drought stress RNA-seq data were “PRJNA355046” and “PRJNA875963” [5,7]; “PRJNA309526” for mild and severe drought stress [13] and “PRJNA292037” for cold acclimation of wild C. oleifera cultivars at series location and elevation [4]. Raw data (raw reads) in fastq format were first qualified with FastQC (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/), accessed on 29 October 2022 for Q20, Q30, GC-content and sequence duplication levels; the data were then processed in Hisat2 version 2.2.1 [26] for read alignment to the C. oleifera reference genome. The reads were subjected to fragments per kilobase of transcript per million fragments mapped (FPKM) conversion to obtain the expression value of genes and transcripts. In-house R scripts were used to analyze gene expression and generate heatmaps. A heatmap was generated using ggplot2, reshape2, gplots, and dplyr packages in R version 4.1.2 [27].
3. Results
3.1. Genome-Wide Identification and Phylogenetic Analysis of DREB Genes in C. oleifera
A total of 198 proteins containing AP2/ERF domain(s) (PF00847) were originally obtained in the diploid C. oleifera var. The “Nanyongensis” (CON) (2n = 2x = 30) genome was identified via stepwise procedures, including local BLAST and HMM searches, which were processed at an E-value cutoff of 1E-5 [18] (Figure 1A). These genes contained at least one ap2 domain. The 14th valine (V) and 19th glutamic acid/valine/leucine/alanine (E/V/L/A), used to confirm conserved amino acid residues, were processed to identify 51 candidate DREB genes in C. oleifera. These CoDREBs ranged from 88 to 518 amino acids (average/median 259/237 aa). The predicted molecular weights (MW) of the CoDREB proteins ranged from 9.7 kDa to 59.6 kDa, and the isoelectric points (pI) ranged from 4.62 to 10.44 (Table S1).
3.2. Phylogenetic Relationship, Conserved Motifs and Genes Structure Analysis and Nomenclature of CoDREBs
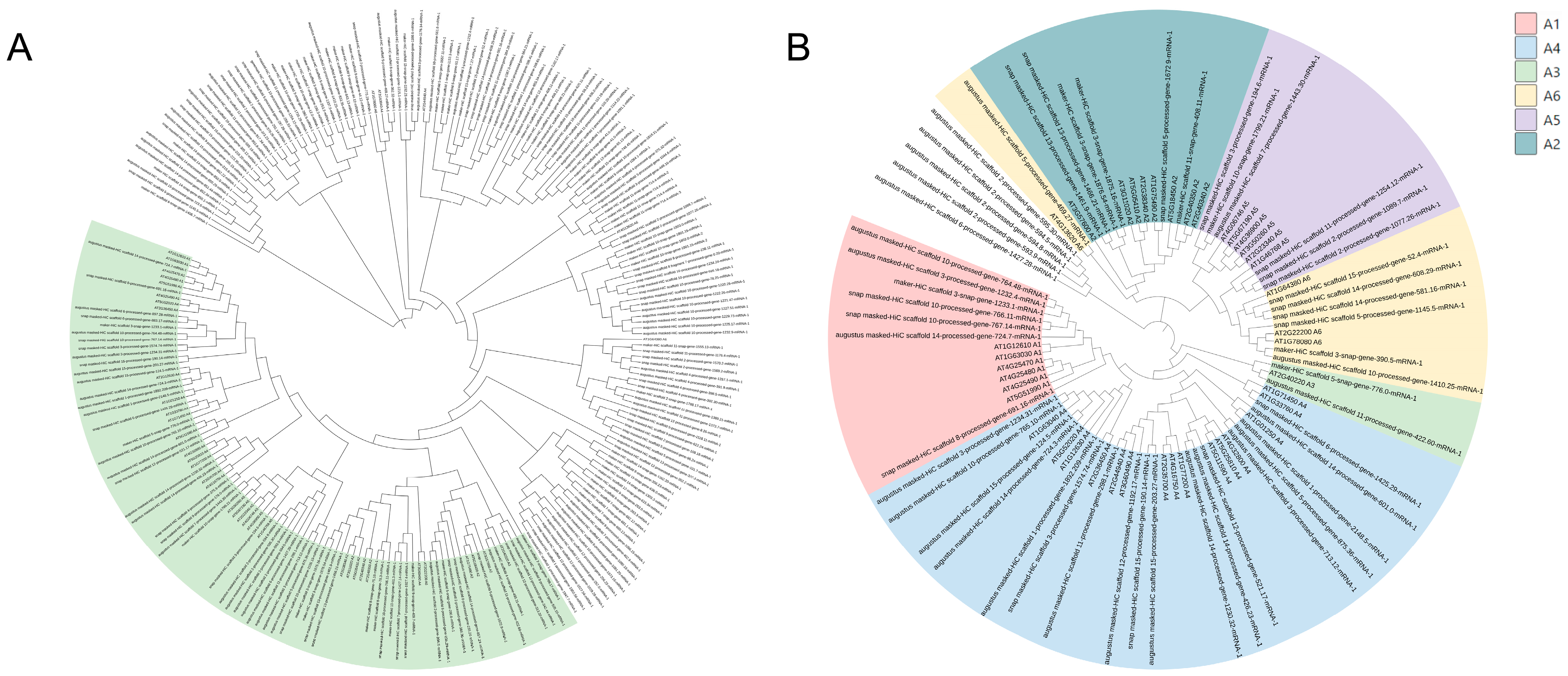
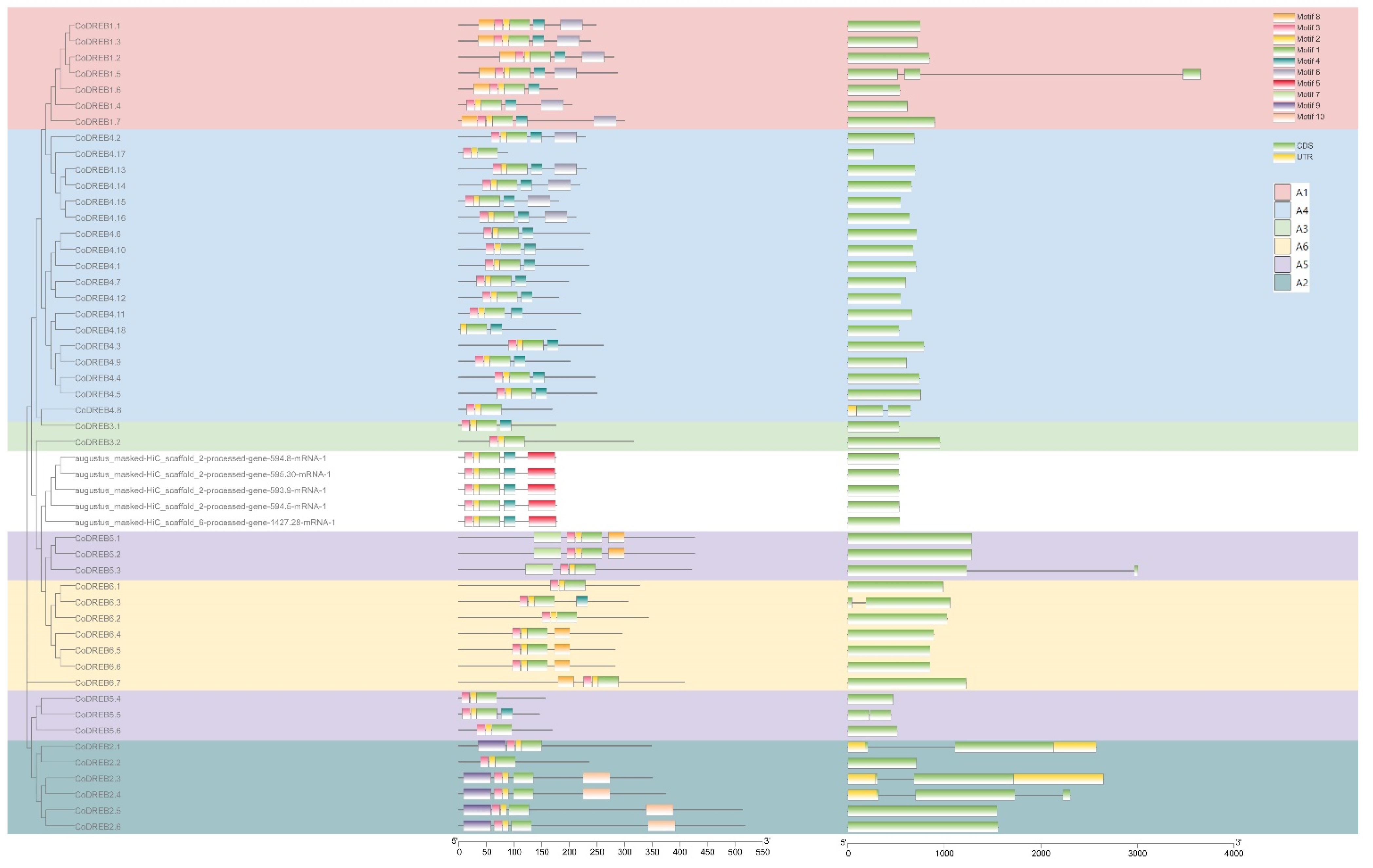
To investigate the evolutionary relationships of CoDREB genes, multiple sequence alignments of 51 putative CoDREBs and 40 AtDREBs were processed, followed by a phylogenetic analysis in MEGAX. The results show that 51 CoDREB genes can be classified into six groups, namely A-1 (with seven members), A-2 (6), A3 (2), A-4 (18), A-5 (6) and A-6 (7) (Figure 1B). Five genes that are evolutionarily distant from the main groups were not classified and are almost never expressed in RNA-seq treatments (Figure S1). Furthermore, in silico conserved motif prediction (MEME suite) and the exon–intro structure were analyzed to overview the structure characteristics of CoDREBs. A total of 10 motifs were predicted and named as Motifs 1 to 10 (Figure 2). All proteins included Motif 1 and 2, which consist of the AP2 structural domain. None of genes contained all 10 motifs. However, the motif diversity is relatively conserved in all groups, with slight differences among subfamilies. The previous five non-classified genes have highly conserved motifs, especially regarding Motif 5. A gene structure analysis showed that most CoDREBs were intronless, 43/51 (84.3%), and the number of exons varied from one to three (Figure 2). Additionally, we suggested a uniform nomenclature for CoDREB, excluding those unclassified genes based on phylogenetic and structural studies to facilitate the future investigation of expression. (Table 1). The “Seq id” values from the GFF file, gene loci, and 14th and 19th amino acid residues are all listed. Interestingly, the 19th leucine was only found in the A-5 and A-6 subfamilies (Table 1).
3.3. Expression Profiles of 46 CoDREB Genes in Response to Short-/Long-Term Drought Stress
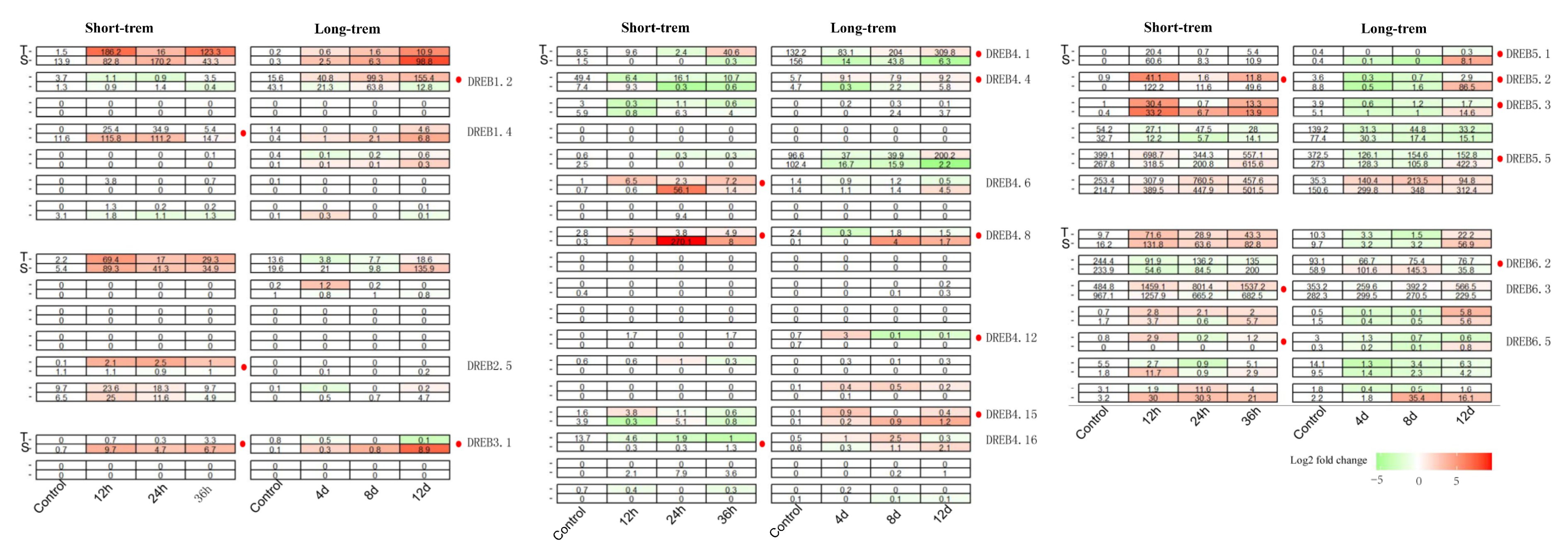
DREB is a plant-specific transcription factor family that has been widely studied for its pleiotropic regulatory roles in response to abiotic stresses, such as drought and cold. However, similar research on tea oil is currently scarce. Thus, to gain insights into the transcriptional regulation of CoDREB genes involved in drought response, a publicly available RNA-seq dataset of C. oleifera was studied during long-term drought (0 d to 12 d, PRJNA875963) and comparatively short-term drought (0 to 36 h, PRJNA355046). Details on data processing could be found in the Section 2. The overall alignment ratio of reads was quantified using a quality control package “fastQC” that revealed ratios between 78.58% and 85.02%. Both experiments compared drought-tolerant (T) and drought-sensitive (S) cultivars of C. oleifera. The expression pattern of CoDREBs was group-plotted using a heatmap (Figure 3). In general, almost two-thirds of the CoDREB genes were up-/down-regulated during drought stresses. However, half of these genes similarly induce/reduce the response to drought stress, demonstrating a general drought adaptation pathway. Instead, we focused on genes that are differentially regulated in expression level between cultivars under long-/short-term drought stress, or even both types of stress. CoDREB1.2/4.1/4.4/4.8/4.12/4.15/5.1/5.3/5.5/6.2 were only differentially regulated in response to long-term drought stress, CoDREB1.4/2.5/4.6/4.16/6.3/6.5 were specifically regulated in response to short-term stress, and only CoDREB3.1/4.8/5.2 were identified for both types of stress (Figure 3). All 18 CoDREBs were identified and labeled, and the changes in CoDREB expression patterns between drought-tolerant and -sensitive cultivars may play an important role in regulating drought adaptation in C. oleifera.
3.4. CoDREBs in Response to Mild, Severe Drought and Followed by Recovery
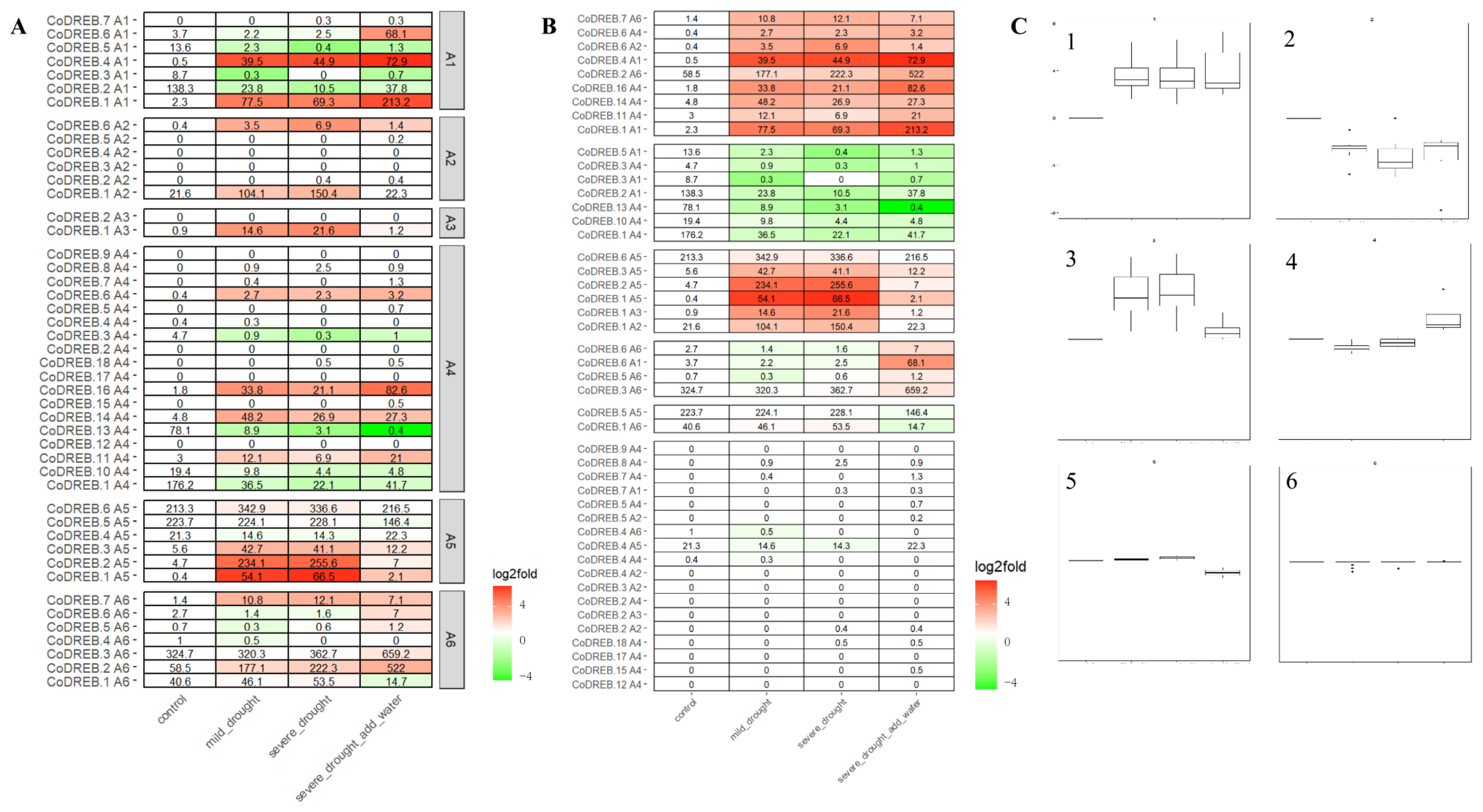
Despite the negative consequences of drought stress, severe environmental circumstances can lead to interesting adaptations in plants that allow them to survive and reproduce [28]. In this scenario, to extend our understanding of possible biological functions of DREBs, the transcriptome data of C. oleifera subjected to mild, severe drought stress and followed by recovery were analyzed. As illustrated in Figure 4A, the expression pattern is relatively varied in each CoDREB subfamily, indicating that the CoDREBs may be involved in a complex drought-responsive regulatory network. We further classified the 46 CoDREBs into six clades based on expression profiling; Clades 1–5 were significantly differentially expressed during drought stress, and Clade 6 was not (Figure 4B,C).
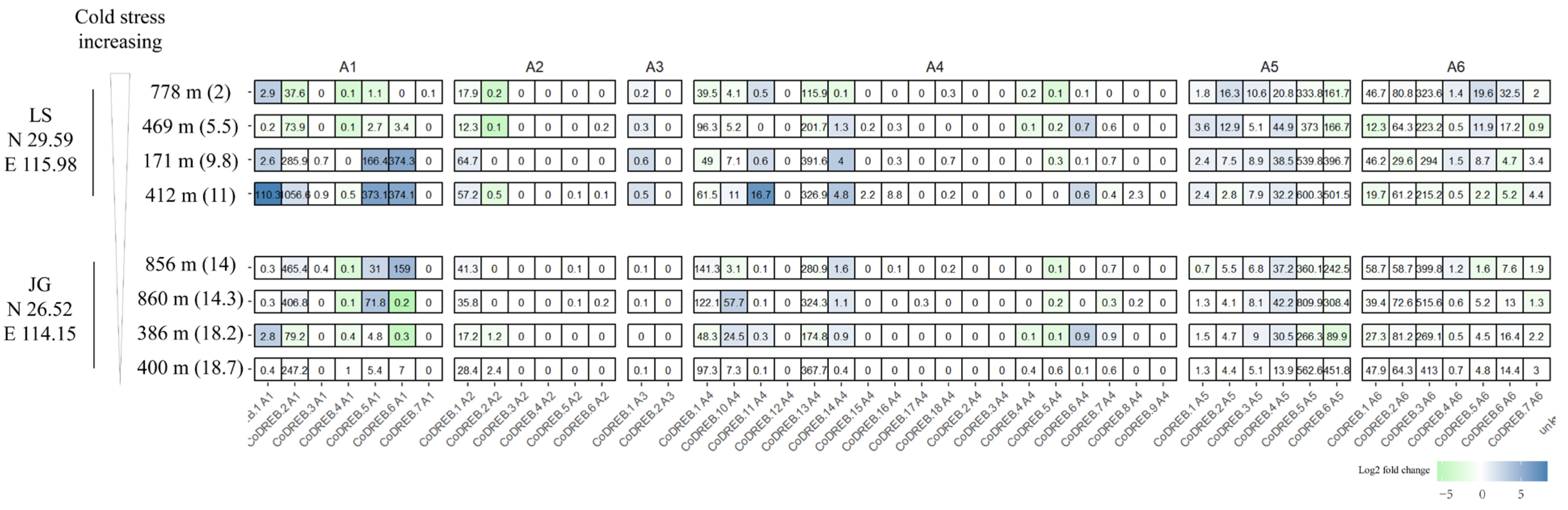
3.5. Transcriptome Analysis of Wild C. oleifera Grown from Different Latitudes and Elevations to Discover CoDREBs Involved in Cold Acclimation
To investigate the response of CoDREB genes regarding cold stress acclimation and preliminarily explore the potential candidate of DREB involved in cold- and drought-stress-related “crosstalk”, we analyzed the transcriptomes of wild tea oil cultivars grown in different locations and elevations [4]. Evidently, cold stress increases with latitude and elevation (Figure 5). C. oleifera wild cultivars were harvested in November or December when the perennial plant’s cold adaptation occurs. A schematic complex transcriptional regulatory network is involved in this adaptation process, in which DREB1 functions as a master regulator [29]. In this study, CoDREB5.2 and CoDREB 6.5 were significantly up-regulated with a temperature decrease from 9.8 to 2, and this might be involved in the cold acclimation of tea oil. (Figure 5).
4. Discussion
The AP2/EREBP superfamily was originally thought to contain plant-specific transcription factors until several AP2 domain-containing HNH endonucleases were found in cyanobacteria (Trichodesmiumerythraeum), ciliate (Tetrahymena thermophile), and viruses (Enterobacteria phage Rb49 and Bacteriophage Felix 01) [30]. The AP2/ERF transcription factors are known to regulate diverse processes of plant development and stress responses (reviewed by [31]). With the same AP2 domain, the valine (14th V) was present in all DREB members but relatively inconsistent at the 19th E/V/L/A, with four candidates identified, whereas alanine (14th A) and aspartic acid (19th D) were conserved in the ERF proteins [32,33,34,35]. With the availability of numerous plant whole-genome sequences, DREB has been widely identified and investigated in many plant species. However, limited studies have been undertaken on non-timber wood species with DREB genes, and even fewer studies have integrated different perspectives to analyze the changes in expression during abiotic stress. Herein, with the release of the CON genome [18], we first identified 51 DREB genes in C. oleifera. In comparison to Arabidopsis (57, genome size 125 Mb), rice (56, 480 Mb), Vitis vinifera (38, 400 Mb), alfalfa (172, 800 Mb), jujube (25, 440 Mb) and Saccharum spontaneum (83, 3.36 Gb) [36,37,38,39,40], only 51 CoDREBs with a genome size of 2.97 Gb were identified. This might be because the aligned diploid CON genome was previously identified as a wild progenitor of cultivated tetraploid/polyploidy C. oleifera [18], demanding an in-depth genome sequencing of cultivated polyploid C. oleifera. Additionally, this finding might be supported by the discovery of DREB genes in tea (Camellia sinensis), in which only 45 CsDREB genes were identified prior to the recent publication of high-resolution genome sequencing data [41,42,43]. Furthermore, the de novo assembly of C. oleifera RNA-seq raw reads resulted in 66,570 unigenes, which is a value 1.43-fold higher than the CON-released 46,617 genes [13,18]. This result cross-validates the limitation of the CON for a cultivated C. oleifera transcriptome experiment. We simultaneously analyzed four different RNA-seq datasets used to standardize gene assembly and alignment within the same framework.
With the 51 identified putative CoDREB genes, the structure analysis of CoDREBs revealed that motif diversity is relatively conserved in all sub-families with slight differences among these subfamilies (Figure 2). Additionally, we obtained a few group-specific motifs (Figure 2), such as Motif 4/5/9/10. Even with a conserved 14th V, CoDREBs containing Motif 5 were phylogenetically separated from the other CoDREBs (Figure 1). Moreover, four genes in this subgroup were nearly rarely expressed in response to drought or cold stress (Figure S1). Taken together, these genes may be involved in different abiotic stress responses, as the DREB family was first identified in response to drought and cold stress, as well as salt stress [33]. For genes induced during short-/long-term drought stress, groups with diverse expression patterns ranging between tolerant (T) and sensitive (S) were separated from the rest (Figure 3). We identified 18 CoDREBs as the candidate drought-responsive genes that potentially participate in a regulatory network. CoDREB1.4/2.5/4.6/4.16/6.3/6.5 were specifically and differently regulated between T/S cultivars in response to relatively short-term drought stress. During this period, KEGG enrichment analysis revealed that both biochemical pathways and the signal transduction pathway were activated, including “biosynthesis of secondary metabolites”, “vitamin B6 metabolism”, “anthocyanin biosynthesis”, and “phosphatidylinositol signaling system” [7]. Chlorophyll content (Chl), peroxidase (POD) activity, malonaldehyde (MDA) concentration, and soluble sugar content were all found to be different between the two cultivars; therefore, the antioxidant capacities of T and S cultivars differed. For long-term drought stress, fewer biochemical and physiological parameters were measured [5]. PIF7, a DREB1 negative regulator under circadian control in Arabidopsis, significantly decreased in sensitive (S) cultivars but was highly expressed in tolerant (T) cultivars; this may indicate the involvement of the circadian clock system in drought stress response. The perception of drought stress response signaling is rapid, and most dehydration-responsive genes are induced by the plant hormones abscisic acid (ABA) and ethylene (ET) [44]. Therefore, because the most enriched pathways and metabolites are involved in antioxidants but not signal transduction, we hypothesized that the genes encoding these 18 CoDREBs are critical for drought acclimation but not for drought signal perception. Furthermore, additional specialized and in-depth experiments with CoDREBs during drought stress are still required.
To understand the possible biological functions of CoDREBs involved in drought tolerance, transcriptome data of C. oleifera subjected to mild and severe drought stresses and followed by recovery were analyzed. Evidently, the expression patterns of CoDREBs are inconsistent within subfamilies (Figure 4A). In another transcription factor gene family in alfalfa [45], cold-responsive bHLH genes with similar expression patterns are clustered with promoter region transcription factor-binding sites (TFBS), rather than structural and conserved motifs (TFBS). Thus, we further classified the 46 CoDREBs into six clades based on expression profiling; Clades 1–5 were significantly differentially expressed during drought stress and Clade 6 was not (Figure 4B,C). We deduced that the genes in Clades 1–5 may be involved in multiple biological processes that have been activated by changing the expression patterns of anti-oxidants in dehydrated plants [46,47,48,49]. For instance, genes in Clade 1 that are constitutively expressed both during mild/severe drought stresses and after recovery are expected to be involved in ROS-scavenging activities that require a specific duration of time to eliminate the accumulated dose after drought water recovery. Moreover, Clade 3 genes are sensitive to external drought stresses, suggesting that they may be engaged in the signal transduction regulatory network. All of these findings show that DREB genes are involved in multiple processes of drought stress adaptation and are worthy of future research.
Regarding cold stress, stomatal opening is inhibited and demonstrates marked changes in lipid composition, mainly in membranes, as well as for stomatal closure during drought stress [50]. Both cold and drought stresses are accompanied by phytohormone accumulation and other signaling components, such as ROS, nitric oxidase (NO), and Ca2+, while the mechanisms themselves require extensive research. In reality, plants grown in dry and semi-arid regions, such as tea and tea oil, are simultaneously exposed to combined stresses, such as cold and drought stresses [51]. The DREB gene family was reported to be involved in cold and drought adaptation. Thus, combined with previous drought RNA-seq data, we adaptively analyzed CoDREB expression patterns in cultivars under long-term cold/freezing conditions (Figure 5). Collectively, CoDREB5.2 and CoDREB6.5 that are significantly up-regulated with a temperature decrease from 9.8 to 2 might be involved in tea oil cold acclimation. Interestingly, CoDREB5.2 and 6.5 were both characterized as drought-responsive elements in a previous analysis. We suggest that these two CoDREB genes could be interesting molecular targets available for the improvement of plant resistance to drought and cold stresses. On the other hand, we predicted that CoDREB5.2 and 6.5 might be involved in cold–drought stress signal crosstalk.
Drought and cold stresses are extreme conditions faced by Camellia oleifera during its growth and affect the growth and development of Camellia oleifera. Improving the drought tolerance and cold tolerance of Camellia oleifera has always been a major focus of research. Through the analysis of the structure and biological function of the CoDREB gene, we found that this gene can be used to improve the ability of Camellia oleifera to adapt to drought and cold stresses.
5. Conclusions
Overall, our study found that the changes in CoDREB expression patterns between drought-tolerant and -sensitive cultivars play an important role in regulating drought adaptation in C. oleifera. A transcriptome analysis of wild C. oleifera grown from different latitudes and elevations also found that CoDREBs were involved in regulating C. oleifera to adapt to cold stress. Drought and cold stresses refer to extreme weather conditions that hurt plants and other living organisms. Both of these can cause physical and physiological damage to the organisms they affect. Gene discovery leads to improved drought/cold stress tolerance in plants via gene transfer. With the improvement of the transgenic system of C. oleifera, it will increase our understanding of the function of CoDREB in resisting the drought and cold stresses of C. oleifera. CoDREB is a new potential target of engineering trees used to resist extreme weather conditions and promote the development of the C. oleifera industry.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijpb14010019/s1, Figure S1: Expression analysis of short-/long-term drought stress with five unknown genes. Table S1: The Molecular weight (MW) and isoelectric point (pI) of 51 CoDREBs.
Author Contributions
J.L. and J.Y. contributed to the design and conception of the study, data collection and analysis, and write up of the final paper. Y.W. and P.G. participated in all phases of the study, experimental design, data collection, analysis and write up the first draft of the paper, Y.W. and P.G. made equal contributions to this work, S.S. provided technical assistance to Y.W. and P.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the following three projects: Changsha Natural Science Foundation, project number: kq2202281; Hunan Forestry Science and Technology Innovation Project, project number: XLK202101-2; China Postdoctoral Science Foundation, project number: 2021M703653.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lenaerts, B.; Collard, B.C.Y.; Demont, M. Review: Improving global food security through accelerated plant breeding. Plant Sci. 2019, 287, 110207. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.S.; Vanderschuren, H.; Qaim, M.; Mahfouz, M.M.; Kohli, A.; Mansoor, S.; Tester, M. New plant breeding technologies for food security. Science 2019, 363, 1390–1391. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Lu, J.; Zhang, Z.; Ma, L.; Liu, C.; Chen, Y. Global transcriptome and correlation analysis reveal cultivar-specific molecular signatures associated with fruit development and fatty acid determination in Camellia oleifera Abel. Int. J. Genom. 2020, 2022, 6162802. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, X.; Huang, X.; Duan, S.; Long, C.; Chen, J.; Rong, J. Leaf transcriptome analysis of a subtropical evergreen broadleaf plant, wild oil-tea camellia (Camellia oleifera), revealing candidate genes for cold acclimation. BMC Genom. 2017, 18, 211. [Google Scholar] [CrossRef]
- He, Z.; Liu, C.; Zhang, Z.; Wang, R.; Chen, Y. Integration of MRNA and MiRNA analysis reveals the differentially regulatory network in two different Camellia oleifera cultivars under drought stress. Front. Plant Sci. 2022, 13, 1001357. [Google Scholar] [CrossRef]
- Zhou, G.; Wei, X.; Wu, Y.; Liu, S.; Huang, Y.; Yan, J.; Zhang, D.; Zhang, Q.; Liu, J.; Meng, Z. Quantifying the hydrological responses to climate change in an intact forested small watershed in southern China. Glob. Change Biol. 2011, 17, 3736–3746. [Google Scholar] [CrossRef]
- Dong, B.; Wu, B.; Hong, W.; Li, X.; Li, Z.; Xue, L.; Huang, Y. Transcriptome analysis of the tea oil camellia (Camellia oleifera) reveals candidate drought stress genes. PLoS ONE 2017, 12, e0181835. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Muhammad, I.; Shalmani, A.; Ali, M.; Yang, Q.H.; Ahmad, H.; Li, F.B. Mechanisms regulating the dynamics of photosynthesis under abiotic stresses. Front. Plant Sci. 2021, 11, 615942. [Google Scholar] [CrossRef] [PubMed]
- Hamanishi, E.T.; Campbell, M.M. Genome-wide responses to drought in forest trees. Forestry 2011, 84, 273–283. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, X.; Zhang, Y.; Du, Z.; Chen, X.; Kong, X.; Sun, W.; Chen, C. Drought stress modify cuticle of tender tea leaf and mature leaf for transpiration barrier enhancement through common and distinct modes. Sci. Rep. 2020, 10, 6696. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, H.Y.; Yang, X.N.; Zhan, J.J.; Zhou, H.; Wang, C.; Yu, Y.; Lu, X.Y.; Chen, Y.Z.; Tian, Y. Transcriptomic analysis of Camellia oleifera in response to drought stress using high throughput RNA-seq. Russ. J. Plant Physiol. 2017, 64, 728–737. [Google Scholar] [CrossRef]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef]
- Kasuga, M.; Liu, Q.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nat. Biotechnol. 1999, 17, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Jaglo-Ottosen, K.R.; Gilmour, S.J.; Zarka, D.G.; Schabenberger, O.; Thomashow, M.F. Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science 1998, 280, 104–106. [Google Scholar] [CrossRef]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought, high-salt and cold-responsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef]
- Lin, P.; Wang, K.; Wang, Y.; Hu, Z.; Yan, C.; Huang, H.; Ma, X.; Cao, Y.; Long, W.; Liu, W.; et al. The genome of Oil-Camellia and population genomics analysis provide insights into seed Oil Domestication. Genome Biol. 2022, 23, 14. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Sonnhammer, E.L.; Eddy, S.R.; Birney, E.; Bateman, A.; Durbin, R. Pfam: Multiple sequence alignments and HMM-profiles of protein domains. Nucleic Acids Res. 1998, 26, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (ITOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Leinonen, R.; Akhtar, R.; Birney, E.; Bower, L.; Cerdeno-Tarraga, A.; Cheng, Y.; Cleland, I.; Faruque, N.; Goodgame, N.; Gibson, R. The european nucleotide archive. Nucleic Acids Res. 2011, 39, D28–D31. [Google Scholar] [CrossRef] [PubMed]
- Grüning, B.; Dale, R.; Sjödin, A.; Chapman, B.A.; Rowe, J.; Tomkins-Tinch, C.H.; Valieris, R.; Köster, J.; The Bioconda Team. Bioconda: Sustainable and comprehensive software distribution for the life sciences. Nat. Methods 2018, 15, 475–476. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; pp. 12–21. [Google Scholar]
- Silva, E.C.D.; Albuquerque, M.; Neto, A.A.; Junior, S. Drought and its consequences to plants—From individual to ecosystem. In Responses of Organisms to Water Stress; IntechOpen: London, UK, 2013; pp. 17–47. [Google Scholar]
- Kidokoro, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant cold-stress responses. Trends Plant Sci. 2022, 27, 922–935. [Google Scholar] [CrossRef]
- Magnani, E.; Sjölander, K.; Hake, S. From endonucleases to transcription factors: Evolution of the AP2 DNA binding domain in plants. Plant Cell 2004, 16, 2265–2277. [Google Scholar] [CrossRef]
- Xu, Z.S.; Chen, M.; Li, L.C.; Ma, Y.Z. Functions and application of the AP2/ERF transcription Factor family in crop improvement: Functions of the AP2/EREBP family. J. Integr. Plant Biol. 2011, 53, 570–585. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, T. Expansion and stress responses of the AP2/EREBP super family in cotton. BMC Genom. 2017, 18, 118. [Google Scholar]
- Sun, J.; Peng, X.; Fan, W.; Tang, M.; Liu, J.; Shen, S. Functional analysis of BpDREB2 gene involved in salt and drought response from a woody plant Broussonetia papyrifera. Gene 2014, 535, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, X.; Liu, Y.; Gao, H.; Wang, Z.; Sun, G. CkDREB gene in caragana korshinskii is involved in the regulation of stress response to multiple abiotic stresses as an AP2/EREBP transcription factor. Mol. Biol. Rep 2011, 38, 2801–2811. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Sun, J.; Liu, Y.; Chen, F.; Shen, S. Isolation and functional characterization of the JcERF gene, a putative AP2/EREBP domain-containing transcription factor, in the woody oil plant Jatropha curcas. Plant Mol. Biol. 2007, 63, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Jia, J.; Kong, D.; Zhang, Z.; Song, S.; Li, Y.; Pang, X. Genome-wide identification and analysis of the DREB genes and their expression profiles under abiotic stresses in Chinese Jujube (Ziziphus jujuba Mill.). J. For. Res. 2019, 30, 1277–1287. [Google Scholar] [CrossRef]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and Rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef]
- Li, Z.; Wang, G.; Liu, X.; Wang, Z.; Zhang, M.; Zhang, J. Genome-wide identification and expression profiling of DREB genes in Saccharum spontaneum. BMC Genom. 2021, 22, 456. [Google Scholar] [CrossRef] [PubMed]
- Sheng, S.; Guo, X.; Wu, C.; Xiang, Y.; Duan, S.; Yang, W.; Li, W.; Cao, F.; Liu, L. Genome-wide identification and expression analysis of DREB genes in Alfalfa (Medicago sativa) in response to cold stress. Plant Signal. Behav. 2022, 17, 2081420. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lv, H.; Li, L.; Liu, J.; Mu, S.; Li, X.; Gao, J. Genome-wide analysis of the AP2/ERF transcription factors family and the expression patterns of DREB genes in Moso Bamboo (Phyllostachys edulis). PLoS ONE 2015, 10, e0126657. [Google Scholar] [CrossRef]
- Wu, Z.J.; Li, X.H.; Liu, Z.W.; Li, H.; Wang, Y.X.; Zhuang, J. Transcriptome-based discovery of AP2/ERF transcription factors related to temperature stress in tea plant (Camellia sinensis). Funct. Integr. Genom. 2015, 15, 741–752. [Google Scholar] [CrossRef]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Xia, E.; Lu, Y.; Tai, Y.; She, G. Draft genome sequence of Camellia sinensis Var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, E4151–E4158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, S.; Shi, L.; Gong, D.; Zhang, S.; Zhao, Q.; Zhan, D.; Vasseur, L.; Wang, Y.; Yu, J. Haplotype-resolved genome assembly provides insights into evolutionary history of the tea plant Camellia sinensis. Nat. Genet. 2021, 53, 1250–1259. [Google Scholar] [CrossRef]
- Perumal, V. Signaling and Communication in Plants, 1st ed.; Springer: Dordrecht, The Netherlands, 2020; pp. 249–267. [Google Scholar]
- Li, G.; Jin, L.; Sheng, S. Genome-wide identification of BHLH transcription factor in medicago sativa in response to cold stress. Genes 2022, 13, 2371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lei, L.; Lai, J.; Zhao, H.; Song, W. Effects of drought stress and water recovery on physiological responses and gene expression in maize seedlings. BMC Plant Biol. 2018, 18, 68. [Google Scholar] [CrossRef]
- Selote, D.S.; Bharti, S.; Khanna-Chopra, R. Drought acclimation reduces O2− accumulation and lipid peroxidation in wheat seedlings. Biochem. Biophys. Res. Commun. 2004, 314, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. The Water-Water Cycle in Chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Hayano-Kanashiro, C.; Calderón-Vázquez, C.; Ibarra-Laclette, E.; Herrera-Estrella, L.; Simpson, J. Analysis of gene expression and physiological responses in three Mexican maize landraces under drought stress and recovery irrigation. PLoS ONE 2009, 4, e7531. [Google Scholar] [CrossRef]
- Agurla, S.; Gahir, S.; Munemasa, S.; Murata, Y.; Raghavendra, A.S. Mechanism of stomatal closure in plants exposed to drought and cold stress. In Survival Strategies in Extreme Cold and Desiccation: Adaptation Mechanisms and Their Applications; Springer: Singapore, 2018. [Google Scholar]
- Zheng, C.; Wang, Y.; Ding, Z.; Zhao, L. Global transcriptional analysis reveals the complex relationship between tea quality, leaf senescence and the responses to cold-drought combined stress in Camellia sinensis. Front. Plant Sci. 2016, 7, 1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Phylogenetic analysis of CoDREBs using BLAST and other programs; a total of 198 proteins containing AP2/ERF domain(s) were obtained in the diploid C. oleifera var. “Nanyong” (CON) (2n = 2x = 30) genome. (A) Multiple sequence alignments were performed for 51 CoDREBs and 40 AtDREBs, and phylogenetic analysis was performed in MEGAX. A total of 51 CoDREB genes could be classified into six groups, namely A1 (with 7 members), A2 (with 6 members), A3 (with 2 members), A4 (with 18 members), A5 (with 6 members) and A6 (with 7 members). (B).
Figure 1.
Phylogenetic analysis of CoDREBs using BLAST and other programs; a total of 198 proteins containing AP2/ERF domain(s) were obtained in the diploid C. oleifera var. “Nanyong” (CON) (2n = 2x = 30) genome. (A) Multiple sequence alignments were performed for 51 CoDREBs and 40 AtDREBs, and phylogenetic analysis was performed in MEGAX. A total of 51 CoDREB genes could be classified into six groups, namely A1 (with 7 members), A2 (with 6 members), A3 (with 2 members), A4 (with 18 members), A5 (with 6 members) and A6 (with 7 members). (B).

Figure 2.
We used MEME suite version 5.4.1 to predict the silico conserved motif prediction and exon–intro structure of CoDREBs protein; a total of 10 motifs were predicted and named as Motifs 1 to 10.
Figure 2.
We used MEME suite version 5.4.1 to predict the silico conserved motif prediction and exon–intro structure of CoDREBs protein; a total of 10 motifs were predicted and named as Motifs 1 to 10.

Figure 3.
Expression analysis of short-/long-term drought stress with CoDREBs, T: Drought-tolerant cultivars of C. oleifera, S: Drought-sensitive cultivars of C. oleifera, Long-term: 0 d to 12 d, Short-term: 0 to 36 h.
Figure 3.
Expression analysis of short-/long-term drought stress with CoDREBs, T: Drought-tolerant cultivars of C. oleifera, S: Drought-sensitive cultivars of C. oleifera, Long-term: 0 d to 12 d, Short-term: 0 to 36 h.

Figure 4.
Expression analysis of mild, severe drought stress and severe drought with CoDREB expression. (A) According to the expression profile, 46 CoDREBs are divided into six Clades. (B,C). Color means the up/down of gene expression, red means the up and green means the down. The darker the color, the higher the expression.
Figure 4.
Expression analysis of mild, severe drought stress and severe drought with CoDREB expression. (A) According to the expression profile, 46 CoDREBs are divided into six Clades. (B,C). Color means the up/down of gene expression, red means the up and green means the down. The darker the color, the higher the expression.

Figure 5.
Expression analysis of wild C. oleifera cultivars grown in different locations and elevations with CoDREBs; (2)–(18.7) indicates the temperature, LS: Lu Mountain, JG: Jinggang Mountain, N: Latitude, E: Longitude. Color means the up/down of gene expression, blue means the up and green means the down. The darker the color, the higher the expression.
Figure 5.
Expression analysis of wild C. oleifera cultivars grown in different locations and elevations with CoDREBs; (2)–(18.7) indicates the temperature, LS: Lu Mountain, JG: Jinggang Mountain, N: Latitude, E: Longitude. Color means the up/down of gene expression, blue means the up and green means the down. The darker the color, the higher the expression.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The seq-id, gene loci, and 14th valine (V) and 19th glutamic acid/valine/leucine/alanine (E/V/L/A) amino acid residues of 51 CoDREBs.
Table 1.
The seq-id, gene loci, and 14th valine (V) and 19th glutamic acid/valine/leucine/alanine (E/V/L/A) amino acid residues of 51 CoDREBs.
| Symbol | Seq Id | 14th | 19th | Source | Strand | Start | End |
|---|---|---|---|---|---|---|---|
| CoDREB1.1 | augustus_masked-HiC_scaffold_10-processed-gene-764.48-mRNA-1 | V | E | HiC_scaffold_10 | − | 76409702 | 76410451 |
| CoDREB1.2 | augustus_masked-HiC_scaffold_3-processed-gene-1232.4-mRNA-1 | V | E | HiC_scaffold_3 | − | 123283087 | 123283932 |
| CoDREB1.3 | snap_masked-HiC_scaffold_10-processed-gene-766.11-mRNA-1 | V | E | HiC_scaffold_10 | − | 76623619 | 76624338 |
| CoDREB1.4 | augustus_masked-HiC_scaffold_14-processed-gene-724.7-mRNA-1 | V | E | HiC_scaffold_14 | + | 72478180 | 72478797 |
| CoDREB1.5 | maker-HiC_scaffold_3-snap-gene-1233.1-mRNA-1 | V | E | HiC_scaffold_3 | − | 123362118 | 123365776 |
| CoDREB1.6 | snap_masked-HiC_scaffold_10-processed-gene-767.14-mRNA-1 | V | E | HiC_scaffold_10 | − | 76705366 | 76705905 |
| CoDREB1.7 | snap_masked-HiC_scaffold_8-processed-gene-691.16-mRNA-1 | V | E | HiC_scaffold_8 | − | 69154923 | 69155825 |
| CoDREB2.1 | maker-HiC_scaffold_11-snap-gene-408.11-mRNA-1 | V | E | HiC_scaffold_11 | + | 40854256 | 40856829 |
| CoDREB2.2 | snap_masked-HiC_scaffold_5-processed-gene-1672.9-mRNA-1 | V | E | HiC_scaffold_5 | + | 167280494 | 167281204 |
| CoDREB2.3 | maker-HiC_scaffold_3-snap-gene-1875.16-mRNA-1 | V | E | HiC_scaffold_3 | − | 187547455 | 187550105 |
| CoDREB2.4 | maker-HiC_scaffold_3-snap-gene-1876.54-mRNA-1 | V | E | HiC_scaffold_3 | − | 187645199 | 187647503 |
| CoDREB2.5 | snap_masked-HiC_scaffold_13-processed-gene-1466.21-mRNA-1 | V | E | HiC_scaffold_13 | − | 146604874 | 146606418 |
| CoDREB2.6 | snap_masked-HiC_scaffold_13-processed-gene-1461.9-mRNA-1 | V | E | HiC_scaffold_13 | + | 146118529 | 146120085 |
| CoDREB3.1 | maker-HiC_scaffold_5-snap-gene-776.0-mRNA-1 | V | E | HiC_scaffold_5 | + | 77602969 | 77603618 |
| CoDREB3.2 | augustus_masked-HiC_scaffold_11-processed-gene-422.60-mRNA-1 | V | E | HiC_scaffold_11 | + | 42215560 | 42216513 |
| CoDREB4.1 | augustus_masked-HiC_scaffold_14-processed-gene-1230.32-mRNA-1 | V | E | HiC_scaffold_14 | − | 123064670 | 123065377 |
| CoDREB4.10 | snap_masked-HiC_scaffold_12-processed-gene-521.17-mRNA-1 | V | E | HiC_scaffold_12 | - | 52181645 | 52182322 |
| CoDREB4.11 | augustus_masked-HiC_scaffold_3-processed-gene-713.12-mRNA-1 | V | A | HiC_scaffold_3 | + | 71361910 | 71362575 |
| CoDREB4.12 | augustus_masked-HiC_scaffold_14-processed-gene-601.0-mRNA-1 | V | E | HiC_scaffold_14 | + | 60115087 | 60115632 |
| CoDREB4.13 | augustus_masked-HiC_scaffold_3-processed-gene-1234.31-mRNA-1 | V | E | HiC_scaffold_3 | − | 123398472 | 123399167 |
| CoDREB4.14 | augustus_masked-HiC_scaffold_10-processed-gene-765.10-mRNA-1 | V | E | HiC_scaffold_10 | − | 76529118 | 76529777 |
| CoDREB4.15 | augustus_masked-HiC_scaffold_15-processed-gene-124.5-mRNA-1 | V | E | HiC_scaffold_15 | + | 12454548 | 12455093 |
| CoDREB4.16 | augustus_masked-HiC_scaffold_14-processed-gene-724.3-mRNA-1 | V | E | HiC_scaffold_14 | + | 72440566 | 72441204 |
| CoDREB4.17 | augustus_masked-HiC_scaffold_1-processed-gene-1892.209-mRNA-1 | V | E | HiC_scaffold_1 | + | 189269071 | 189269337 |
| CoDREB4.18 | augustus_masked-HiC_scaffold_5-processed-gene-875.36-mRNA-1 | V | A | HiC_scaffold_5 | − | 87523685 | 87524215 |
| CoDREB4.2 | snap_masked-HiC_scaffold_3-processed-gene-1574.74-mRNA-1 | V | E | HiC_scaffold_3 | − | 157422246 | 157422935 |
| CoDREB4.3 | augustus_masked-HiC_scaffold_12-processed-gene-1192.17-mRNA-1 | V | E | HiC_scaffold_12 | − | 119269647 | 119270435 |
| CoDREB4.4 | snap_masked-HiC_scaffold_15-processed-gene-190.14-mRNA-1 | V | E | HiC_scaffold_15 | + | 19021440 | 19022183 |
| CoDREB4.5 | augustus_masked-HiC_scaffold_15-processed-gene-203.27-mRNA-1 | V | E | HiC_scaffold_15 | − | 20340240 | 20340995 |
| CoDREB4.6 | augustus_masked-HiC_scaffold_14-processed-gene-426.23-mRNA-1 | V | E | HiC_scaffold_14 | + | 42621443 | 42622156 |
| CoDREB4.7 | augustus_masked-HiC_scaffold_1-processed-gene-2148.5-mRNA-1 | V | E | HiC_scaffold_1 | + | 214865752 | 214866351 |
| CoDREB4.8 | snap_masked-HiC_scaffold_6-processed-gene-1425.29-mRNA-1 | V | E | HiC_scaffold_6 | − | 142541920 | 142542450 |
| CoDREB4.9 | augustus_masked-HiC_scaffold_11-processed-gene-298.1-mRNA-1 | V | E | HiC_scaffold_11 | − | 29817495 | 29818103 |
| CoDREB5.1 | snap_masked-HiC_scaffold_2-processed-gene-1077.26-mRNA-1 | V | L | HiC_scaffold_2 | − | 107715224 | 107716507 |
| CoDREB5.2 | snap_masked-HiC_scaffold_2-processed-gene-1089.7-mRNA-1 | V | L | HiC_scaffold_2 | + | 108932798 | 108934081 |
| CoDREB5.3 | snap_masked-HiC_scaffold_11-processed-gene-1254.12-mRNA-1 | V | L | HiC_scaffold_11 | + | 125398486 | 125401490 |
| CoDREB5.4 | augustus_masked-HiC_scaffold_7-processed-gene-1443.30-mRNA-1 | V | E | HiC_scaffold_7 | − | 144370313 | 144370783 |
| CoDREB5.5 | maker-HiC_scaffold_10-snap-gene-1799.21-mRNA-1 | V | E | HiC_scaffold_10 | + | 179923482 | 179923929 |
| CoDREB5.6 | snap_masked-HiC_scaffold_3-processed-gene-194.6-mRNA-1 | V | E | HiC_scaffold_3 | − | 19425828 | 19426337 |
| CoDREB6.1 | snap_masked-HiC_scaffold_5-processed-gene-1145.5-mRNA-1 | V | L | HiC_scaffold_5 | + | 114517410 | 114518396 |
| CoDREB6.2 | augustus_masked-HiC_scaffold_10-processed-gene-1410.25-mRNA-1 | V | L | HiC_scaffold_10 | − | 141067137 | 141068168 |
| CoDREB6.3 | maker-HiC_scaffold_3-snap-gene-390.5-mRNA-1 | V | L | HiC_scaffold_3 | − | 39027029 | 39028091 |
| CoDREB6.4 | snap_masked-HiC_scaffold_15-processed-gene-52.4-mRNA-1 | V | L | HiC_scaffold_15 | + | 5199287 | 5200177 |
| CoDREB6.5 | snap_masked-HiC_scaffold_14-processed-gene-581.16-mRNA-1 | V | L | HiC_scaffold_14 | − | 58133946 | 58134797 |
| CoDREB6.6 | snap_masked-HiC_scaffold_14-processed-gene-608.29-mRNA-1 | V | L | HiC_scaffold_14 | + | 60806950 | 60807801 |
| CoDREB6.7 | augustus_masked-HiC_scaffold_5-processed-gene-469.27-mRNA-1 | V | L | HiC_scaffold_5 | − | 46960303 | 46961529 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Y.; Guo, P.; Sheng, S.; Li, J.; Yan, J. Identification of CoDREB Genes for Drought and Cold Tolerance in Camellia oleifera. Int. J. Plant Biol. 2023, 14, 228-241. https://doi.org/10.3390/ijpb14010019
AMA Style
Wang Y, Guo P, Sheng S, Li J, Yan J. Identification of CoDREB Genes for Drought and Cold Tolerance in Camellia oleifera. International Journal of Plant Biology. 2023; 14(1):228-241. https://doi.org/10.3390/ijpb14010019
Chicago/Turabian StyleWang, Ying, Purui Guo, Song Sheng, Jian’an Li, and Jindong Yan. 2023. "Identification of CoDREB Genes for Drought and Cold Tolerance in Camellia oleifera" International Journal of Plant Biology 14, no. 1: 228-241. https://doi.org/10.3390/ijpb14010019