
Physiological and Morphological Response of Argania spinosa (L.) Skeels to a Competing Liana: Case Study under Arid Climate

 ,
,
Abstract
:1. Introduction
2. Material and Methods
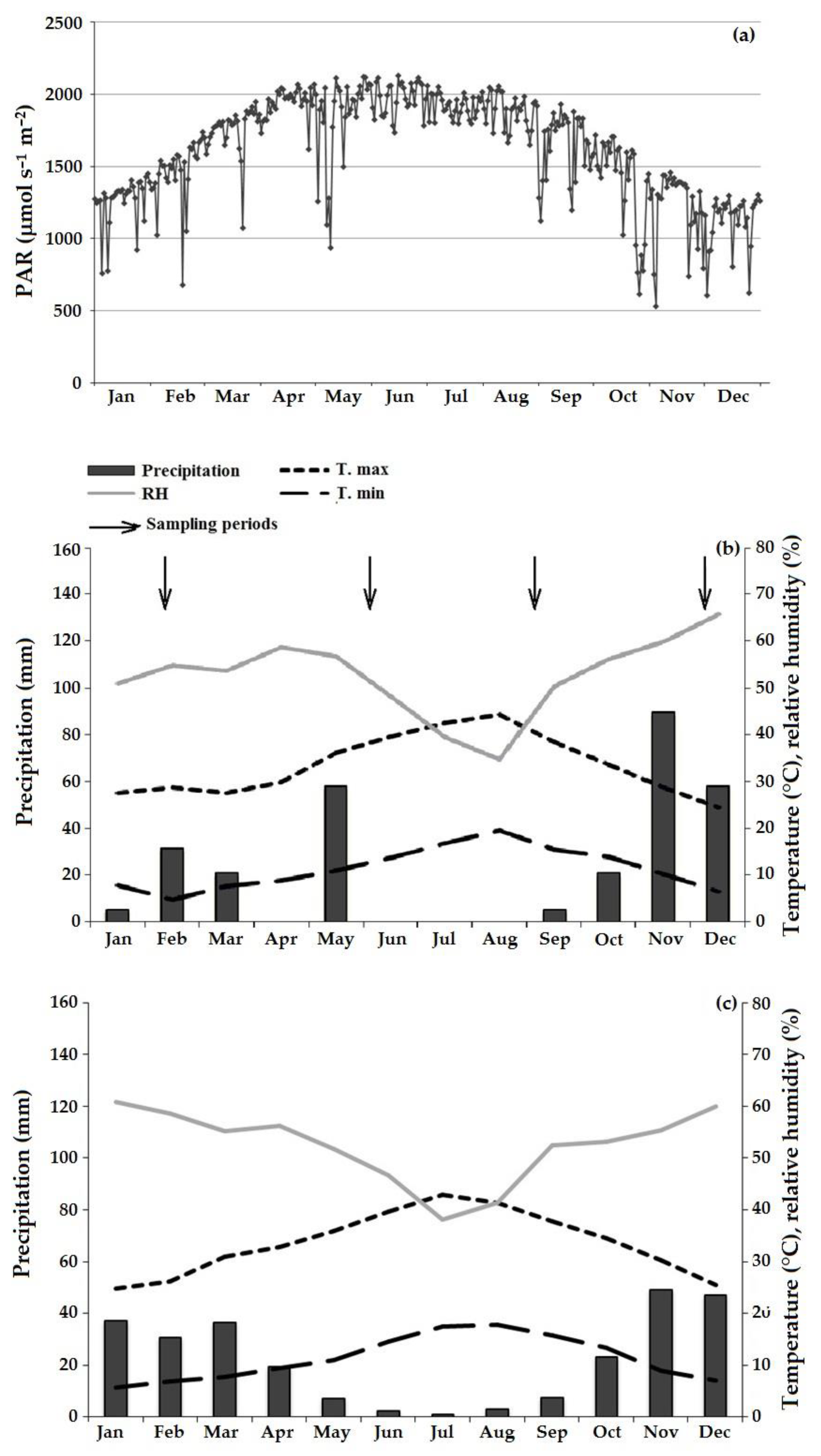
2.1. Study Site and Plant Material
2.2. Gas Exchange
2.3. Water Status
2.4. Photosynthetic Pigment Content
2.5. Leaf Dry Mass per Unit Area
2.6. Statistical Analysis
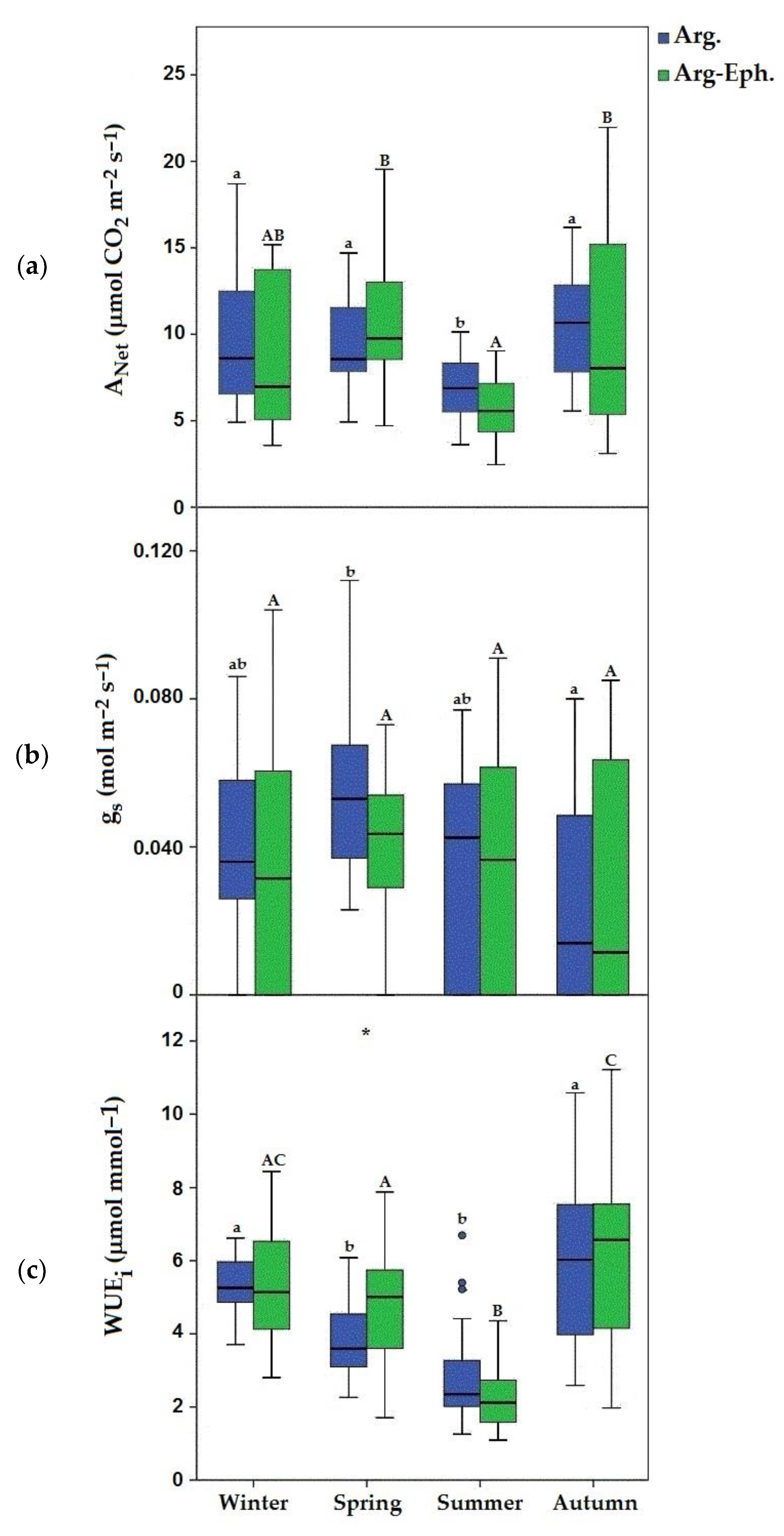
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Callaway, R.M. Positive interactions among plants. Bot. Rev. 1995, 61, 306–349. [Google Scholar] [CrossRef]
- Tilman, D. Resource Competition and Community Structure; Monographs in Population Biology 17; Princeton University Press: Princeton, NJ, USA, 1982. [Google Scholar]
- Schoener, T.W. Field experiments on interspecific competition. Am. Nat. 1983, 122, 240–285. [Google Scholar] [CrossRef]
- Estrada-Villegas, S.; Pedraza Narvaez, S.S.; Sanchez, A.; Schnitzer, S.A. Lianas significantly reduce tree performance and biomass accumulation across tropical forests: A global meta-Analysis. Front. For. Glob. Chang. 2022, 4, 812066. [Google Scholar] [CrossRef]
- Burnham, R.J. Climbing Plants in the Fossil Record: Paleozoic to Present. In The Ecology of Lianas; Schnitzer, S.A., Bongers, F., Burnham, R.J., Putz, F.E., Eds.; Wiley-Blackwell: Oxford, UK, 2015; pp. 205–220. [Google Scholar]
- Londré, R.; Schnitzer, S. The distribution of lianas and their change in abundance in temperate forests over the past 45 years. Ecology 2006, 87, 2973–2978. [Google Scholar] [CrossRef] [Green Version]
- Schnitzer, S.A.; Mangan, S.A.; Dalling, J.W.; Baldeck, C.; Hubbell, S.P.; Ledo, A.; Muller Landau, H.; Tobin, M.; Aguilar, S.; Brassfield, D.; et al. Liana abundance, diversity, and distribution on Barro Colorado Island, 499 Panama. PLoS ONE 2012, 7, e52114. [Google Scholar] [CrossRef] [Green Version]
- Schnitzer, S. Increasing Liana Abundance in Neotropical Forests: Causes and Consequences. In Ecology of Lianas; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 451–464. [Google Scholar] [CrossRef]
- Laurance, W.F.; Andrade, A.S.; Magrach, A.; Camargo, J.L.; Valsko, J.J.; Campbell, M.; Fearnside, P.M.; Edwards, W.; Lovejoy, T.E.; Laurance, S.G. Long-term changes in liana abundance and forest dynamics in undisturbed Amazonian forests. Ecology 2014, 95, 1604–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marvin, D.C.; Winter, K.; Burnham, R.J.; Schnitzer, S.A. No evidence that elevated CO2 gives tropical lianas an advantage over tropical trees. Glob. Chang. Biol. 2015, 21, 2055–2069. [Google Scholar] [CrossRef] [Green Version]
- Parolari, A.J.; Paul, K.; Griffing, A.; Condit, R.; Perez, R.; Aguilar, S.; Schnitzer, S.A. Liana abundance and diversity increase with rainfall seasonality along a precipitation gradient in Panama. Ecography 2019, 42, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Stewart, T.E.; Schnitzer, S.A. Blurred lines between competition and parasitism. Biotropica 2017, 49, 433–438. [Google Scholar] [CrossRef]
- Van der Heijden, G.M.F.; Phillips, O.L. Liana infestation impacts tree growth in a lowland tropical moist Forest. Biogeosciences 2009, 6, 2217–2226. [Google Scholar] [CrossRef] [Green Version]
- Schnitzer, S.A.; Van Der Heijden, G.M.F.; Mascaro, J.; Carson, W.P. Lianas in gaps reduce carbon accumulation in a tropical forest. Ecology 2014, 95, 3008–3017. [Google Scholar] [CrossRef] [Green Version]
- Van der Heijden, G.M.F.; Powers, S.J.; Schnitzer, S.A. Lianas reduce carbon accumulation in tropical forests. Proc. Natl. Acad. Sci. USA 2015, 112, 13267–13271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledo-Aceves, T. Above- and Belowground Competition between Lianasand Trees. In Ecology of Lianas; Schnitzer, S.A., Bongers, F., Burnham, R.J., Putz, F.E., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 147–163. [Google Scholar]
- Jones, I.L.; Peres, C.A.; Benchimol, M.; Bunnefeld, L.; Dent, D.H.; Zang, R. Woody lianas increase in dominance and maintain compositional integrity across an Amazonian dam-induced fragmented landscape. PLoS ONE 2017, 12, e0185527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-Villegas, S.; Schnitzer, S.A. A comprehensive synthesis of liana removal experiments in tropical forests. Biotropica 2018, 50, 729–739. [Google Scholar] [CrossRef]
- Becknell, J.M.; Vargas, G.G.; Wright, L.A.; Woods, N.F.; Medvigy, D.; Powers, J.S. Increasing Liana Abundance and Associated Reductions in Tree Growth in Secondary Seasonally Dry Tropical Forest. Front. For. Glob. Chang. 2022, 5, 838357. [Google Scholar] [CrossRef]
- Kainer, K.A.; Wadt, L.H.O.; Staudhammer, C.L. Testing a silvicultural recommendation: Brazil nut responses 10 years after liana cutting. J. Appl. Ecol. 2014, 51, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Leon, M.; Martinez-Izquierdo, L.; Powers, J.S.; Schnitzer, S.A. Lianas reduce community-level canopy tree reproduction in a Panamanian forest. J. Ecol. 2018, 106, 737–745. [Google Scholar] [CrossRef] [Green Version]
- Grauel, W.T.; Putz, F.E. Effects of lianas on growth and regeneration of Prioria copaifera in Darien, Panama. For. Ecol. Manag. 2004, 190, 99–108. [Google Scholar] [CrossRef]
- Schnitzer, S.A.; Carson, W.P. Lianas suppress tree regeneration and diversity in tree fall gaps. Ecol. Lett. 2010, 13, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.D.; Schnitzer, S.A.; Wright, S.J.; Muller Landau, H.C.; Jongejans, E.; Comita, L.S.; de Kroon, H.; Condit, R.; Hubbell, S.P. Tree species vary widely in their tolerance for liana infestation: A case study of differential host response to generalist parasites. J. Ecol. 2018, 106, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Ingwell, L.L.; Wright, S.J.; Becklund, K.K.; Hubbell, S.P.; Schnitzer, S.A. The impact of lianas on 10 years of tree growth and mortality on Barro Colorado Island, Panama. J. Ecol. 2010, 98, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Schnitzer, S.A. Testing ecological theory with lianas. New Phytol. 2018, 220, 366–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konapala, G.; Mishra, A.K.; Wada, Y.; Mann, M.E. Climate change will affect global water availability through compounding changes in seasonal precipitation and evaporation. Nat. Commun. 2020, 11, 3044. [Google Scholar] [CrossRef] [PubMed]
- Mitchard, E.T.A. The tropical forest carbon cycle and climate change. Nature 2018, 559, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Villegas, S.; Hall, J.S.; van Breugel, M.; Schnitzer, S.A. Lianas do not reduce tree biomass accumulation in young successional tropical dry forests. Oecologia 2021, 195, 1019–1029. [Google Scholar] [CrossRef]
- Brandes, A.F.; Rizzieri, Y.C.; Tamaio, N.; Pace, M.R.; Barros, C.F. A global review on wood growth rings in lianas. Dendrochronologia 2022, 71, 125920. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A. Distribution and effects on tree growth of lianas and woody hemiepiphytes in a Costa Rican tropical wet forest. J. Trop. Ecol. 1990, 6, 321–331. [Google Scholar] [CrossRef]
- Dillenburg, L.R.; Whigham, D.F.; Teramura, A.H.; Forseth, I.N. Effects of below- and aboveground competition from vines Lonicera japonica and Parthenocissus quinquefolia on the growth of the tree host Liquidambar styraciflua. Oecologia 1993, 93, 48–54. [Google Scholar] [CrossRef]
- Dillenburg, L.R.; Teramura, A.H.; Forseth, I.N.; Whigham, D.F. Photosynthetic and biomass allocation responses of Liquidambar styraciflua (Hamamelidaceae) to vine competition. Am. J. Bot. 1995, 82, 454–461. [Google Scholar] [CrossRef]
- Barker, M.G.; Pérez-Salicrup, D. Tree Comparative water relations of mature mahogany (Swietenia macrophylla) trees with and without lianas in a subhumid, seasonally dry forest in Bolivia. Physiology 2000, 20, 1167–1174. [Google Scholar] [CrossRef]
- Cai, Z.Q.; Poorter, L.; Cao, K.F.; Bongers, F. Seedling growth strategies in Bauhinia species: Comparing lianas and trees. Ann. Bot. 2007, 100, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Bongers, F.; Cao, K.; Cai, Z. Above- and below-ground competition in high and low irradiance: Tree seedling responses to a competing liana Byttneria grandifolia. J. Trop. Ecol. 2008, 24, 517–524. [Google Scholar] [CrossRef]
- Alvarez-Cansino, L.; Schnitzer, S.A.; Reid, J.P.; Powers, J.S. Liana competition with tropical trees varies seasonally but not with tree species identity. Ecology 2015, 96, 39–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnitzer, S.A.A. Mechanistic explanation for global patterns of liana abundance and distribution. Am. Nat. 2005, 166, 262–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dörken, V.M. Leaf-morphology and leaf-anatomy in Ephedra altissima DESF. (Ephedraceae, Gnetales) and their evolutionary relevance. Feddes Repert. 2012, 123, 243–255. [Google Scholar] [CrossRef]
- Michon, G.; Nasi, R.; Balent, G. Public policies and management of rural forests: Lasting alliance or fool’s dialogue? Ecol. Soc. 2013, 18, 30. [Google Scholar] [CrossRef] [Green Version]
- Lybbert, T.J.; Magnan, N.; Aboudrare, A. Household and local forest impacts of Morocco’s argan oil bonanza. Environ. Dev. Econ. 2010, 15, 439–464. [Google Scholar] [CrossRef]
- Morton, J.F.; Voss, G.L. The Argan tree (Argania sideroxylon, Sapotaceae), a desert source of edible oil. Econ. Bot. 1987, 41, 221–233. [Google Scholar] [CrossRef]
- Chakhchar, A.; Lamaoui, M.; Wahbi, S.; Ferradous, A.; El Mousadik, A.; Ibnsouda-Koraichi, S.; Filali-Maltouf, A.; El Modafar, C. Leaf water status, osmoregulation and secondary metabolism as a model for depicting drought tolerance in Argania spinosa. Acta Physiol. Plant 2015, 37, 1–16. [Google Scholar] [CrossRef]
- Chakhchar, A.; Wahbi, S.; Lamaoui, M.; Ferradous, A.; El Mousadik, A.; Ibnsouda-Koraichi, S.; Filali-Maltouf, A.; El Modafar, C. Physiological and biochemical traits of drought tolerance in Argania spinosa. J. Plant Interact. 2015, 10, 252–261. [Google Scholar] [CrossRef]
- Chakhchar, A.; Lamaoui, M.; Aissam, S.; Ferradous, A.; Wahbi, S.; El Mousadik, A.; Ibnsouda-Koraichi, S.; Filali-Maltouf, A.; El Modafar, C. Differential physiological and antioxidative responses to drought stress and recovery among four contrasting Argania spinosa ecotypes. J. Plant Interact. 2016, 11, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Chakhchar, A.; Haworth, M.; El Modafar, C.; Lauteri, M.; Mattioni, C.; Wahbi, S.; Centritto, M. An assessment of genetic diversity and drought tolerance in argan tree (Argania spinosa) populations: Potential for the development of improved drought tolerance. Front. Plant. Sci. 2017, 8, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakhchar, A.; Lamaoui, M.; Aissam, S.; Ferradous, A.; Wahbi, S.; El Mousadik, A.; Ibnsouda-Koraichi, S.; Filali-Maltouf, A.; El Modafar, C. Physiological and Biochemical Mechanisms of Drought Stress Tolerance in the Argan Tree. In Plant Metabolites and Regulation Under Environmental Stress; Ahmad, P., Ahanger, M.A., Singh, V.P., Tripathi, D.K., Alam, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 311–322. [Google Scholar] [CrossRef]
- Chakhchar, A.; Lamaoui, M.; Aissam, S.; Ferradous, A.; Wahbi, S.; El Mousadik, A.; Ibnsouda-Koraichi, S.; Filali-Maltouf, A.; El Modafar, C. Physiological and carbohydrate metabolism traits for discrimination of drought-tolerant elite ecotypes of Argania spinosa. Plant Physiol. Rep. 2019, 24, 388–398. [Google Scholar] [CrossRef]
- Chakhchar, A.; Ben Salah, I.; El Kharrassi, Y.; Filali-Maltouf, A.; El Modafar, C.; Lamaoui, M. Agro-fruit-forest systems based on Argan tree in Morocco: A review of recent results. Front. Plant Sci. 2022, 12, 783615. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Barradas, M.C.; Zunzunegui, M.; Ain-Lhout, F.; Jáuregui, J.; Boutaleb, S.; Álvarez-Cansino, L.; Esquivias, M.P. Seasonal physiological responses of Argania spinosa tree from Mediterranean to semi-arid climate. Plant Soil 2010, 337, 217–231. [Google Scholar] [CrossRef]
- Díaz-Barradas, M.C.; Zunzunegui, M.; Esquivias, M.P.; Boutaleb, S.; Valera-Burgos, J.; Tagma, T.; Ain-Lhout, F. Some secrets of Argania spinosa water economy in a semiarid climate. Nat. Prod. Commun. 2013, 8, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Ait Bihi, M.; Ain-Lhout, F.; Hatimi, A.; Fahmi, F.; Tahrouch, S. Ecophysiological Response and Morphological Adjustment of Argania spinosa L. Skeels under Contrasting Climates: Case Study of Marginal Populations. Int. J. Plant Biol. 2021, 12, 9404. [Google Scholar] [CrossRef]
- NaitDouch, A.; Boukhalef, L.; El Asbahani, A.; Al-Namazi, A.A.; El Mehrach, K.; Bouqbis, L.; Touaf, M.; Ain-Lhout, F. Photosynthetic Behavior of Argania spinosa (L.) Skeels Induced under Grazed and Ungrazed Conditions. Sustainability 2022, 14, 12081. [Google Scholar] [CrossRef]
- Alados, C.L.; El Aich, A. Stress assessment of argan (Argania spinosa L. Skeels) in response to land use across an aridity gradient: Translational asymmetry and branch fractal dimension. J. Arid Environ. 2008, 72, 338–349. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classifcation updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- M’Hirit, O.; Benzyane, M.; Benchekroum, F.; El Yousfi, S.M.; Bendaanoun, M. L’Arganier: Une Espèce Forestière à Usages Multiples; Mardaga: Sprimont, Belgium, 1998. [Google Scholar]
- NASA. Power Data Access Viewer. Natural. 2020. Available online: https://power.larc.nasa.gov/data-access-iewer/ (accessed on 15 August 2022).
- Ordiales-Plaza, R. Midebmp; ver 4.2; Estación Experimental de Zonas Áridas, CSIC, Consejo Superior de Investigaciones Científicas: Almería, Spain, 2000. [Google Scholar]
- Garnier, E.; Shipley, B.; Roumet, C.; Laurent, G.A. Standardized protocol for the determination of specific leaf area and dry matter content. Funct. Ecol. 2001, 15, 688–695. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Louati, M.; Ucarli, C.; Arikan, B.; Ghada, B.; Hannachi, A.S.; Turgut-Kara, N. Genetic, morphological, and biochemical diversity of argan tree (Argania spinosa L.) (sapotaceae) in Tunisia. Plants 2019, 8, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, E.N.; Ribeiro, R.V.; Ferreira-Silva, S.L.; Viégas, R.A.; Silveira, J.A.G. Comparative effects of salinity and water stress on photosynthesis, water relations and growth of Jatropha curcas plants. J. Arid Environ. 2010, 74, 1130–1137. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Cescatti, A.; Rodeguiero, M.; Tosens, T. Complex adjustments of photosynthetic potentials and internal diffusion conductance to current and previous light availabilities and leaf age in Mediterranean evergreen species Quercus ilex. Plant Cell Environ. 2006, 29, 1159–1178. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Prot. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Lennon, A.M.; Lewis, V.R.; Farrell, A.D.; Umaharan, P. Photochemical responses to light in sun and shade leaves of Theobroma cacao L. (West African Amelonado). Sci. Hortic. 2021, 276, 109747. [Google Scholar] [CrossRef]
- Hikosaka, K.; Terashima, I.A. model of the acclimation of photosynthesis in the leaves of C3 plants to sun and shade with respect to nitrogen use. Plant Cell Environ. 1995, 18, 605–618. [Google Scholar] [CrossRef]
- Kitajima, K.; Hogan, K.P. Increases of chlorophyll a/b ratios during acclimation of tropical woody seedlings to nitrogen limitation and high light. Plant Cell Environ. 2003, 26, 857–865. [Google Scholar] [CrossRef]
- Nyongesah Maina, J.; Wang, Q. Seasonal Response of Chlorophyll a/b Ratio to Stress in a Typical Desert Species: Haloxylonammodendron. Arid Land Res. Manag. 2015, 29, 321–334. [Google Scholar] [CrossRef]
- Pishchik, V.N.; Filippova, P.S.; Mirskaya, G.V.; Khomyakov, Y.V.; Vertebny, V.E.; Dubovitskaya, V.I.; Ostankova, Y.V.; Semenov, A.V.; Chakrabarty, D.; Zuev, E.V.; et al. Epiphytic PGPB Bacillus megaterium AFI1 and Paenibacillus nicotianae AFI2 Improve Wheat Growth and Antioxidant Status under Ni Stress. Plants. 2021, 10, 2334. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, N.; Attard, E. Autochthonous and International Olive Cultivars in Malta: Morphometric characteristics and Physicochemical Analysis. SSRG Int. J. Agric. Environ. Sci. 2021, 8, 15–22. [Google Scholar] [CrossRef]
- Maslova, T.G.; Popova, I.A. Adaptive properties of the plant pigment systems. Photosynthetica. 1993, 29, 195–203. [Google Scholar]
- Kyparissis, A.; Drilias, P.; Manetas, Y. Seasonal fluctuations in photoprotective (xanthophyll cycle) and photoselective (chlorophylls) capacity in eight Mediterranean plant species belonging to two different growth forms. Aust. J. Plant Physiol. 2000, 27, 265–272. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Damesin, C.; Rambal, S.; Joffre, R. Between-tree variations in leaf delta13C of Quercus pubescens and Quercus ilex among Mediterranean habitats with different water availability. Oecologia. 1997, 111, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Bacon, M.A. Water Use Efficiency in Plant Biology; Blackwell Publishing Ltd.: Oxford, UK; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Shen, H.; Dong, S.; Li, S.; Xiao, J.; Han, Y.; Yang, M.; Zhang, J.; Gao, X.; Xu, Y.; Li, Y.; et al. Grazing enhances plant photosynthetic capacity by altering soil nitrogen in alpine grasslands on the Qinghai-Tibetan plateau. Agric. Ecosyst. Environ. 2019, 280, 161–168. [Google Scholar] [CrossRef]
- Aïachi Mezghani, M.; Mguidiche, A.; Allouche Khebour, F.; Zouari, I.; Attia, F.; Provenzano, G. Water Status and Yield Response to Deficit Irrigation and Fertilization of Three Olive Oil Cultivars under the Semi-Arid Conditions of Tunisia. Sustainability. 2019, 11, 4812. [Google Scholar] [CrossRef]
- Villar, R.J.; Ruiz-Robleto, J.L.; Ubera, J.L.; Poorter, H. Exploring variation in leaf mass per area (LMA) from leaf to cell: An anatomical analysis of 26 woody species. Am. J. Bot. 2013, 100, 1969–1980. [Google Scholar] [CrossRef]
- Alonso-Forn, D.; Sancho-Knapik, D.; Ferrio, J.P.; Peguero-Pina, J.J.; Bueno, A.; Onoda, Y.; Cavender-Bares, J.; Ülo, N.; Jansen, S.; Riederer, M. Revisiting the functional basis of sclerophylly within the leaf economics spectrum of oaks: Different roads to Rome. Curr. For. Rep. 2020, 6, 260–281. [Google Scholar] [CrossRef]
- Jordan, G.J.; Dillon, R.A.; Weston, P.H. Solar radiation as a factor in the evolution of scleromorphic leaf anatomy in Proteaceae. Am. J. Bot. 2005, 92, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü. Does the touch of cold make evergreen leaves tougher? Tree Physiol. 2016, 36, 267–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratani, L.; Bombelli, A. Leaf anatomy, inclination, and gas exchange relationships in evergreen sclerophyllous and drought semi-deciduous shrub species. Photosynthetica 1999, 37, 573–585. [Google Scholar] [CrossRef]
- Hassiotou, F.; Renton, M.; Ludwig, M.; Evans, J.R.; Veneklaas, E.J. Photosynthesis at an extreme end of the leaf trait spectrum: How does it relate to high leaf dry mass per area and associated structural parameters? J. Exp. Bot. 2010, 61, 3015–3028. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trunk Perimeter (m) | Tree Height (m) | Canopy Size (m3) | Leaf Area (cm2) | Liana Canopy Cover (%) | |
|---|---|---|---|---|---|
| Arg. | 1.44 ± 0.04 | 4.26 ± 0.33 | 112.95 ± 6.53 | 1.94 ± 0.29 | - |
| Arg-Eph. | 1.51 ± 0.08 | 4.21 ± 0.37 | 119.93 ± 15.96 | 1.85 ± 0.35 | 10–20% |
| Winter | Spring | Summer | Autumn | |||||
|---|---|---|---|---|---|---|---|---|
| Arg. | Arg-Eph. | Arg. | Arg-Eph. | Arg. | Arg-Eph. | Arg. | Arg-Eph. | |
| RWC | 68.7 ± 2.41 a | 70.63 ± 5.56 AB | 71.85 ± 5.58 a | 72.60 ± 4.90 B | 68.51 ± 4.00 a | 65.41 ± 8.51 A | 82.35 ± 3.58 b | 83.07 ± 4.72 C |
| Chl a | 40.58 ± 14.74 a * | 54.16 ± 13.55 A * | 48.55 ± 9.01 a | 56.39 ± 12.64 A | 39.03 ± 11.72 a | 35.80 ± 10.54 B | 50.12 ± 8.66 a | 48.90 ± 8.96 A |
| Chl b | 17.70 ± 7.34 a * | 24.96 ± 6.89 A * | 28.99 ± 7.24 b | 24.24 ± 7.29 A | 17.27 ± 6.42 a | 20.69 ± 8.90 A | 21.97 ± 5.50 ab | 25.70 ± 7.10 A |
| Chl a/b | 2.44 ± 0.42 a | 2.25 ± 0.27 AB | 1.77 ± 0.24 b * | 2.72 ± 0.76 B * | 2.41 ± 0.45 a * | 1.94 ± 0.33 A * | 2.37 ± 0.31 a | 2.08 ± 0.63 AB |
| Car | 14.08 ± 5.15 a * | 18.97 ± 6.02 A * | 17.15 ± 2.93 a | 18.61 ± 5.13 A | 13.04 ± 3.96 a * | 16.46 ± 5.20 A * | 15.24 ± 3.11 a | 15.92 ± 2.66 A |
| LMA | 117.81 ± 13.37 a | 131.12 ± 35.10 AB | 133.87 ± 20.83 b * | 149.1 ± 34.63 B * | 138.96 ± 21.35 b | 144.25 ± 25.70 B | 110.15 ± 19.78 a | 113.8 ± 20.31 A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boukhalef, L.; Nait Douch, A.; Bouqbis, L.; El Asbahani, A.; Touaf, M.; Ain-Lhout, F. Physiological and Morphological Response of Argania spinosa (L.) Skeels to a Competing Liana: Case Study under Arid Climate. Int. J. Plant Biol. 2023, 14, 1-13. https://doi.org/10.3390/ijpb14010001
Boukhalef L, Nait Douch A, Bouqbis L, El Asbahani A, Touaf M, Ain-Lhout F. Physiological and Morphological Response of Argania spinosa (L.) Skeels to a Competing Liana: Case Study under Arid Climate. International Journal of Plant Biology. 2023; 14(1):1-13. https://doi.org/10.3390/ijpb14010001
Chicago/Turabian StyleBoukhalef, Laila, Aicha Nait Douch, Laila Bouqbis, Abdelhafed El Asbahani, Mourad Touaf, and Fatima Ain-Lhout. 2023. "Physiological and Morphological Response of Argania spinosa (L.) Skeels to a Competing Liana: Case Study under Arid Climate" International Journal of Plant Biology 14, no. 1: 1-13. https://doi.org/10.3390/ijpb14010001