
Public Health Importance of Preventive Measures for Salmonella Tennessee and Salmonella Typhimurium Strain LT2 Biofilms
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains’ Preparation
2.2. Bacterial Inoculation, Biofilm Formation, and Treatments
2.3. Study Design and Descriptive and Inferential Statistical Analyses
3. Results and Discussion
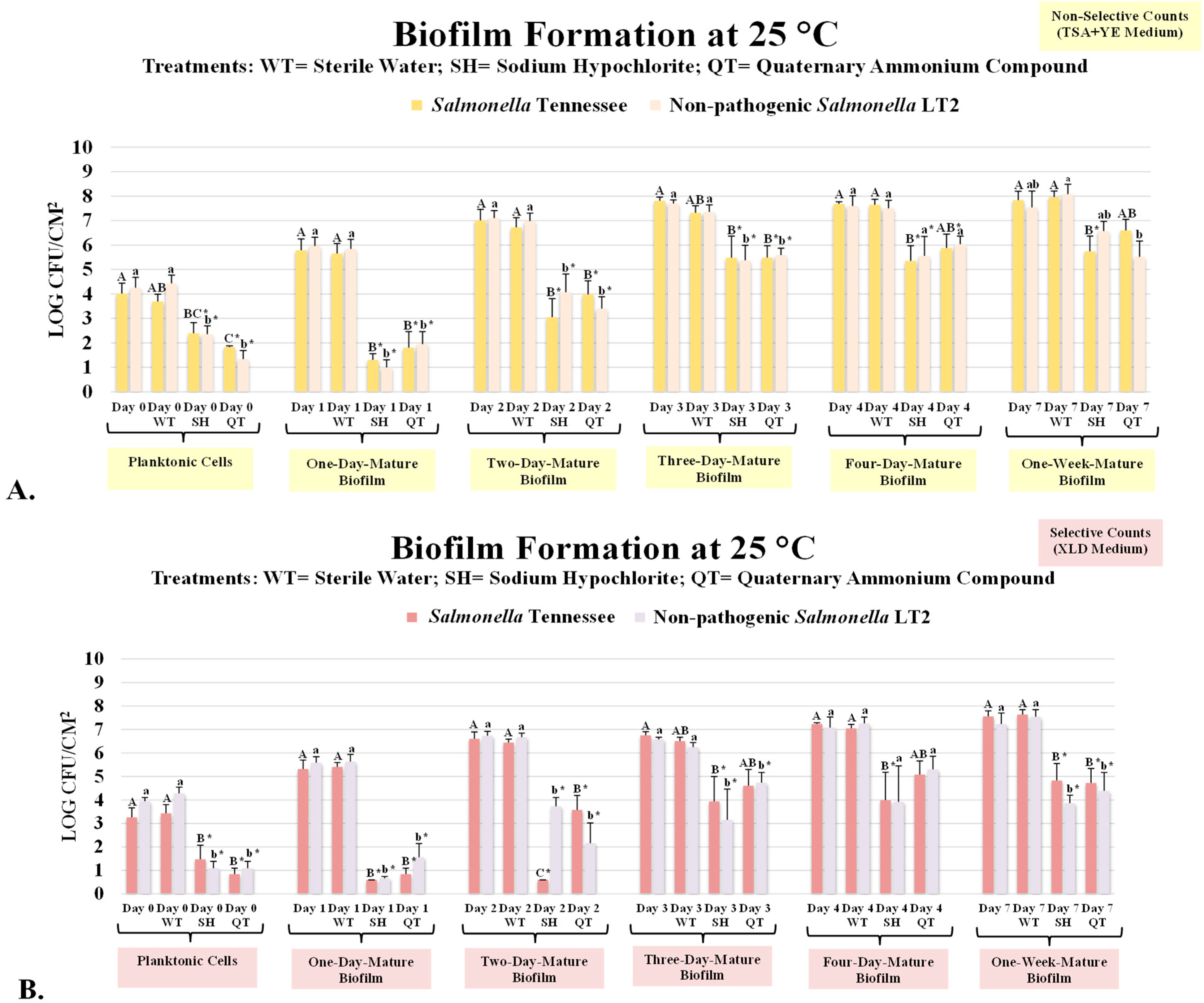
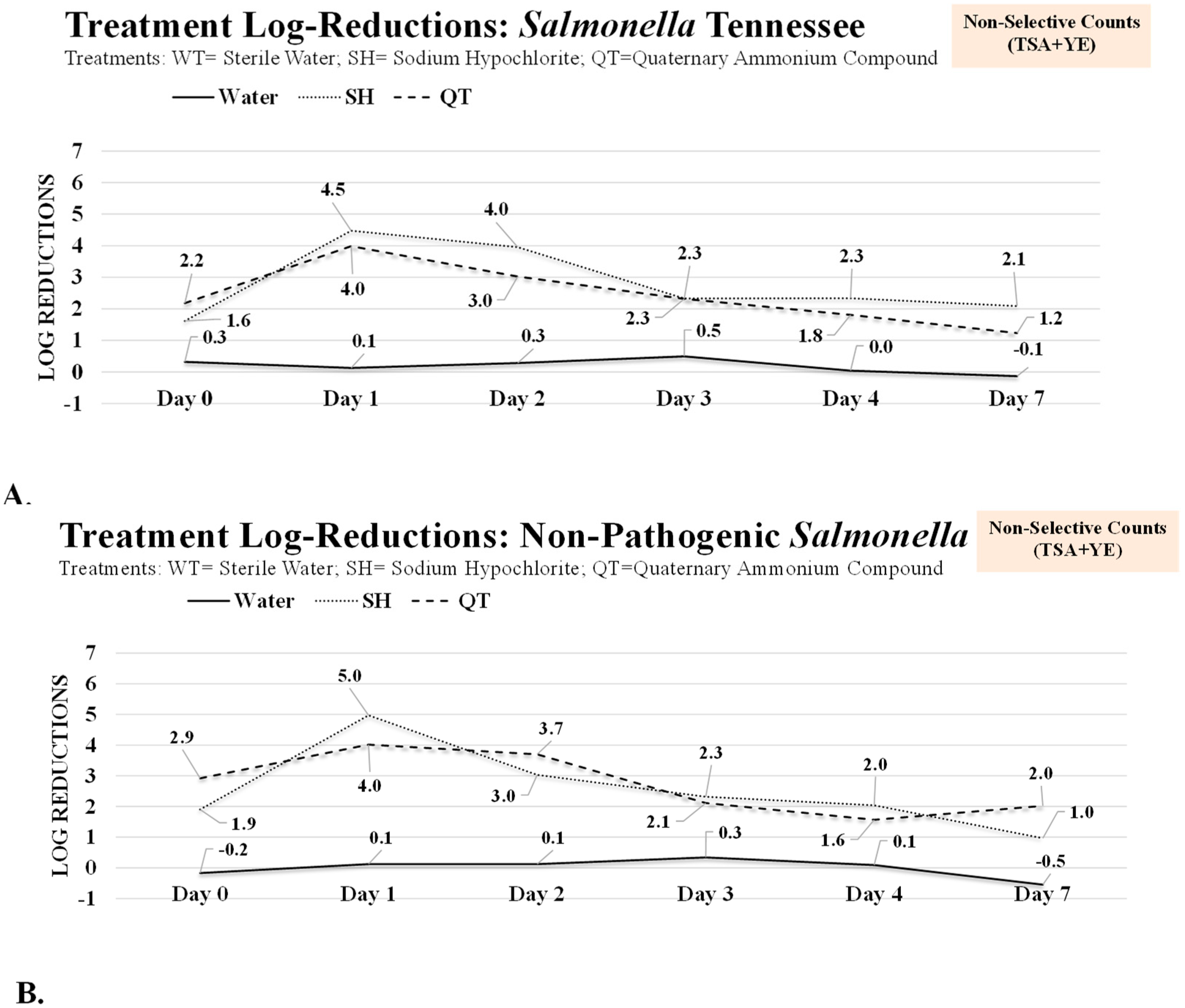
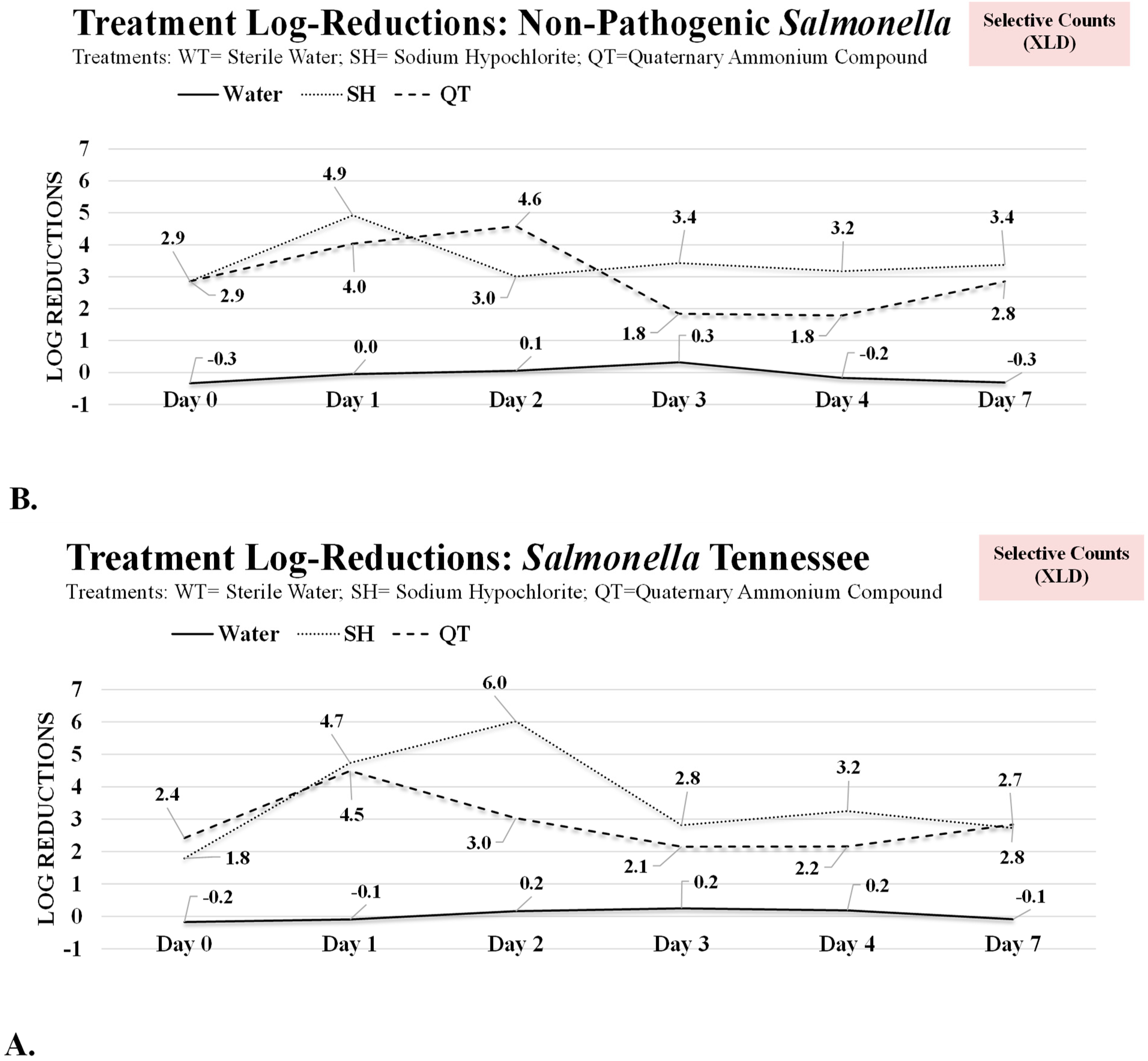
3.1. Antimicrobial Treatment of Planktonic Cells and One-Day and Two-Day Mature Biofilms of Salmonella Tennessee and Salmonella LT2
3.2. Inactivation of Three-, Four-, and Seven-Day Biofilms of Salmonella Tennessee and Salmonella LT2
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eng, S.K.; Pusparajah, P.; Ab Mutalib, N.S.; Ser, H.L.; Chan, K.G.; Lee, L.H. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef]
- Kumar, A.; Allison, A.; Henry, M.; Scales, A.; Fouladkhah, A.C. Development of salmonellosis as affected by bioactive food compounds. Microorganisms 2019, 7, 364. [Google Scholar] [CrossRef]
- Grimont, P.A.; Weill, F.X. Antigenic Formulae of the Salmonella Serovars; WHO Collaborating Centre for Reference and Research on Salmonella: Paris, France, 2007. [Google Scholar]
- Gal-Mor, O.; Boyle, E.C.; Grassl, G.A. Same species, different diseases: How and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front. Microbiol. 2014, 5, 391. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. J. Emerg. Infect. Dis. 2011, 17, 7. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Mahon, B.E.; Jones, T.F.; Griffin, P.M. An assessment of the human health impact of seven leading foodborne pathogens in the United States using disability adjusted life years. Epidemiol. Infect. 2015, 143, 2795–2804. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Luby, S.P.; Mintz, E.D. The global burden of typhoid fever. Bull. World Health Organ. 2004, 82, 346–353. [Google Scholar]
- Buckle, G.C.; Walker, C.L.F.; Black, R.E. Typhoid fever and paratyphoid fever: Systematic review to estimate global morbidity and mortality for 2010. J. Glob. Health 2012, 2, 010401. [Google Scholar] [CrossRef]
- Fouladkhah, A.C.; Thompson, B.; Camp, J.S. Safety of food and water supplies in the landscape of changing climate. Microorganisms 2019, 7, 469. [Google Scholar] [CrossRef]
- Fouladkhah, A.C.; Thompson, B.; Camp, J.S. The threat of antibiotic resistance in changing climate. Microorganisms 2020, 8, 748. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Fouladkhah, A.C. Outbreak history, biofilm formation, and preventive measures for control of Cronobacter sakazakii in infant formula and infant care settings. Microorganisms 2019, 7, 77. [Google Scholar] [CrossRef]
- Allison, A.; Fouladkhah, A.C. Sensitivity of planktonic cells and biofilm of wild-type and pressure-stressed Cronobacter sakazakii and Salmonella enterica serovars to sodium hypochlorite. Food Prot. Trends 2021, 41, 195–203. [Google Scholar] [CrossRef]
- Fouladkhah, A.; Geornaras, I.; Sofos, J.N. Biofilm formation of O157 and Non-O157 Shiga toxin-producing Escherichia coli and multidrug-resistant and susceptible Salmonella Typhimurium and Newport and their inactivation by sanitizers. J. Food Sci. 2013, 78, M880–M886. [Google Scholar] [CrossRef]
- Kabir, M.N.; Aras, S.; Wadood, S.; Chowdhury, S.; Fouladkhah, A.C. Fate and biofilm formation of wild-type and pressure-stressed pathogens of public health concern in surface water and on abiotic surfaces. Microorganisms 2020, 8, 408. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Multistate outbreak of Salmonella serotype Tennessee infections associated with peanut butter—United States, 2006–2007. MMWR 2007, 56, 521–524. [Google Scholar]
- Fouladkhah, A. The Need for evidence-based outreach in the current food safety regulatory landscape. J. Ext. 2017, 55, 20. [Google Scholar]
- Nummer, B.; Gump, D.; Wells, S.; Zimmerman, S.; Montalbano, A. Hazard Analysis and Critical Control Points (HACCP). In Regulatory Foundations for the Food Protection Professional; Springer: New York, NY, USA, 2015. [Google Scholar]
- Public Health Microbiology FoundationSM. Bacterial Multiplication. Available online: https://publichealthmicrobiology.education/fact-sheets (accessed on 29 December 2022).
- de Moraes, M.H.; Chapin, T.K.; Ginn, A.; Wright, A.C.; Parker, K.; Hoffman, C.; Pascual, D.W.; Danyluk, M.D.; Teplitski, M. Development of an avirulent Salmonella surrogate for modeling pathogen behavior in pre-and postharvest environments. Appl. Environ. Microbiol. 2016, 82, 4100–4111. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ryser, E.T. Quantitative transfer of Salmonella Typhimurium LT2 during mechanical slicing of tomatoes as impacted by multiple processing variables. Int. J. Food Microbiol. 2016, 234, 76–82. [Google Scholar] [CrossRef]
- Malone, M.; Bjarnsholt, T.; McBain, A.J.; James, G.A.; Stoodley, P.; Leaper, D.; Tachi, M.; Schultz, G.; Swanson, T.; Wolcott, R.D. The prevalence of biofilms in chronic wounds: A systematic review and meta-analysis of published data. J. Wound Care 2017, 26, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Buffet-Bataillon, S.; Tattevin, P.; Bonnaure-Mallet, M.; Jolivet-Gougeon. Emergence of resistance to antibacterial agents: The role of quaternary ammonium compounds—A critical review. Int. J. Antimicrob. Agents 2012, 39, 381–389. [Google Scholar] [CrossRef]
- Fukuzaki, S. Mechanisms of actions of sodium hypochlorite in cleaning and disinfection processes. Biocontrol Sci. 2006, 11, 147–157. [Google Scholar] [CrossRef]
- Chowdhury, A.; Aras, S.; Kabir, N.; Wadood, S.; Allison, A.; Chowdhury, S.; Fouladkhah, A.C. Susceptibility of pathogenic nontyphoidal Salmonella serovars and avirulent Salmonella LT2 to elevated hydrostatic pressure and citricidalTM. J. Tenn. Acad. Sci. 2021, 96, 49–54. [Google Scholar] [CrossRef]
- Kabir, M.N.; Aras, S.; Allison, A.; Adhikari, J.; Chowdhury, S.; Fouladkhah, A.C. Interactions of carvacrol, caprylic acid, habituation, and mild heat for pressure-based inactivation of O157 and non-O157 serogroups of Shiga toxin-producing Escherichia coli in acidic environment. Microorganisms 2019, 7, 145. [Google Scholar] [CrossRef]
- Aras, S.; Kabir, M.N.; Chowdhury, S.; Fouladkhah, A.C. Augmenting the pressure-based pasteurization of Listeria monocytogenes by synergism with nisin and mild heat. Int. J. Environ. Res. Public Health 2020, 17, 563. [Google Scholar] [CrossRef]
- Simpson Beauchamp, C.; Dourou, D.; Geornaras, I.; Yoon, Y.; Scanga, J.A.; Belk, K.E.; Smith, G.C.; Nychas, G.J.E.; Sofos, J.N. Transfer, attachment, and formation of biofilms by Escherichia coli O157: H7 on meat-contact surface materials. J. Food Sci. 2012, 77, M343–M347. [Google Scholar] [CrossRef] [PubMed]
- Wijman, J.G.; de Leeuw, P.P.; Moezelaar, R.; Zwietering, M.H.; Abee, T. Air-liquid interface biofilms of Bacillus cereus: Formation, sporulation, and dispersion. Appl. Environ. Microbiol. 2007, 73, 1481–1488. [Google Scholar] [CrossRef]
- Giaouris, E.D.; Nychas, G.J.E. The adherence of Salmonella Enteritidis PT4 to stainless steel: The importance of the air–liquid interface and nutrient availability. Food Microbiol. 2006, 23, 747–752. [Google Scholar] [CrossRef]
- United States Food and Drug Administration. Bacteriological Analytical Methods (FDA BAM). Aerobic Plate Count. 2001. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-3-aerobic-plate-count (accessed on 29 December 2022).
- Zhou, S.; Sheen, S.; Zhao, G.; Chuang, S.; Liu, L. Prediction of Salmonella inactivation in sliced tomato subject to high pressure processing and trans-cinnamaldehyde treatment using selective and non-selective growth media for survival evaluations. Food Control 2020, 118, 107441. [Google Scholar] [CrossRef]
- Allison, A.; Chowdhury, S.; Fouladkhah, A.C. Synergism of mild heat and high-pressure pasteurization against Listeria monocytogenes and natural microflora in phosphate-buffered saline and raw milk. Microorganisms 2018, 6, 102. [Google Scholar] [CrossRef]
- Kabir, M.N.; Aras, S.; George, J.; Wadood, S.; Chowdhury, S.; Fouladkhah, A.C. High-pressure and thermal-assisted pasteurization of habituated, wild-type, and pressure-stressed Listeria monocytogenes, Listeria innocua, and Staphylococcus aureus. LWT 2021, 137, 110445. [Google Scholar] [CrossRef]
- Allison, A.; Daniels, E.; Chowdhury, S.; Fouladkhah, A.C. Effects of elevated hydrostatic pressure against mesophilic background microflora and habituated Salmonella serovars in orange juice. Microorganisms 2018, 6, 23. [Google Scholar] [CrossRef]
- Aviles, B.; Klotz, C.; Smith, T.; Williams, R.; Ponder, M. Survival of Salmonella enterica serotype Tennessee during simulated gastric passage is improved by low water activity and high fat content. J. Food Prot. 2013, 76, 333–337. [Google Scholar] [CrossRef]
- Wilmes-Riesenberg, M.R.; Foster, J.W.; Curtiss, R. An altered rpoS allele contributes to the avirulence of Salmonella typhimurium LT2. Infect. Immun. 1997, 65, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Smolinski, H.S.; Wang, S.; Ren, L.; Chen, Y.; Kowalcyk, B.; Thomas, E.; Doren, J.V.; Ryser, E.T. Transfer and redistribution of Salmonella Typhimurium LT2 and Escherichia coli O157: H7 during pilot-scale processing of baby spinach, cilantro, and romaine lettuce. J. Food Prot. 2018, 81, 953–962. [Google Scholar] [CrossRef]
- Allison, A.; Fouladkhah, A.C. Sensitivity of wild-type and rifampicin-resistant O157 and non-O157 Shiga toxin-producing Escherichia coli to elevated hydrostatic pressure and lactic acid in ground meat and meat homogenate. PLoS ONE 2021, 16, e0246735. [Google Scholar] [CrossRef] [PubMed]
- Kwok, T.Y.; Ma, Y.; Chua, S.L. Biofilm dispersal induced by mechanical cutting leads to heightened foodborne pathogen dissemination. Food Microbiol. 2022, 102, 103914. [Google Scholar] [CrossRef] [PubMed]
- Dantas, S.T.; Rossi, B.F.; Bonsaglia, E.C.; Castilho, I.G.; Hernandes, R.T.; Fernandes, A.; Rall, V.L. Cross-contamination and biofilm formation by Salmonella enterica serovar Enteritidis on various cutting boards. Foodborne Pathog. Dis. 2018, 15, 81–85. [Google Scholar] [CrossRef] [PubMed]
- The U.S. Food and Drug Administration. Bad Bug Book: Handbook of Foodborne Pathogenic Microorganisms and Natural Toxins. 2017. Available online: https://www.fda.gov/files/food/published/Bad-Bug-Book-2nd-Edition-%28PDF%29.pdf (accessed on 29 December 2022).
- The U.S. Centers for Disease Control and Prevention. Oral Health. 2021. Available online: https://www.cdc.gov/oralhealth/index.html (accessed on 29 December 2022).
- Fouladkhah, A. Epidemiology–Laboratory Interactions for Developing Resilience Against Future Infectious Diseases. Research Outreach 2022, p. 129. Available online: https://researchoutreach.org/articles/epidemiology-laboratory-interactions-developing-resilience-future-infectious-diseases/ (accessed on 29 December 2022).
- Aviles, B.; Klotz, C.; Eifert, J.; Williams, R.; Ponder, M. Biofilms promote survival and virulence of Salmonella enterica sv. Tennessee during prolonged dry storage and after passage through an in vitro digestion system. Int. J. Food Microbiol. 2013, 162, 252–259. [Google Scholar] [CrossRef]
- Lee, S.; Chen, J. Identification of the genetic elements involved in biofilm formation by Salmonella enterica serovar Tennessee using mini-Tn10 mutagenesis and DNA sequencing. Food Microbiol. 2022, 106, 104043. [Google Scholar] [CrossRef]
- Jahan, F.; Chinni, S.V.; Samuggam, S.; Reddy, L.V.; Solayappan, M.; Yin, L.S. The Complex Mechanism of the Salmonella typhi Biofilm Formation That Facilitates Pathogenicity: A Review. Int. J. Mol. Sci. 2022, 2, 6462. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asefaw, S.; Aras, S.; Kabir, M.N.; Wadood, S.; Chowdhury, S.; Fouladkhah, A.C. Public Health Importance of Preventive Measures for Salmonella Tennessee and Salmonella Typhimurium Strain LT2 Biofilms. Microbiol. Res. 2023, 14, 714-726. https://doi.org/10.3390/microbiolres14020051
Asefaw S, Aras S, Kabir MN, Wadood S, Chowdhury S, Fouladkhah AC. Public Health Importance of Preventive Measures for Salmonella Tennessee and Salmonella Typhimurium Strain LT2 Biofilms. Microbiology Research. 2023; 14(2):714-726. https://doi.org/10.3390/microbiolres14020051
Chicago/Turabian StyleAsefaw, Simen, Sadiye Aras, Md Niamul Kabir, Sabrina Wadood, Shahid Chowdhury, and Aliyar Cyrus Fouladkhah. 2023. "Public Health Importance of Preventive Measures for Salmonella Tennessee and Salmonella Typhimurium Strain LT2 Biofilms" Microbiology Research 14, no. 2: 714-726. https://doi.org/10.3390/microbiolres14020051