
Omics of an Enigmatic Marine Amoeba Uncovers Unprecedented Gene Trafficking from Giant Viruses and Provides Insights into Its Complex Life Cycle
Abstract
:1. Introduction
2. Materials and Methods
2.1. Life Cycle Observation, Single-Cell Transcriptome, and Genome Sequencing
2.2. De Novo Genome Assembly and Polishing
2.3. Contaminant Removal
2.4. Gene Prediction, LGT Analysis, and Identification of Sex-Related Genes
3. Results
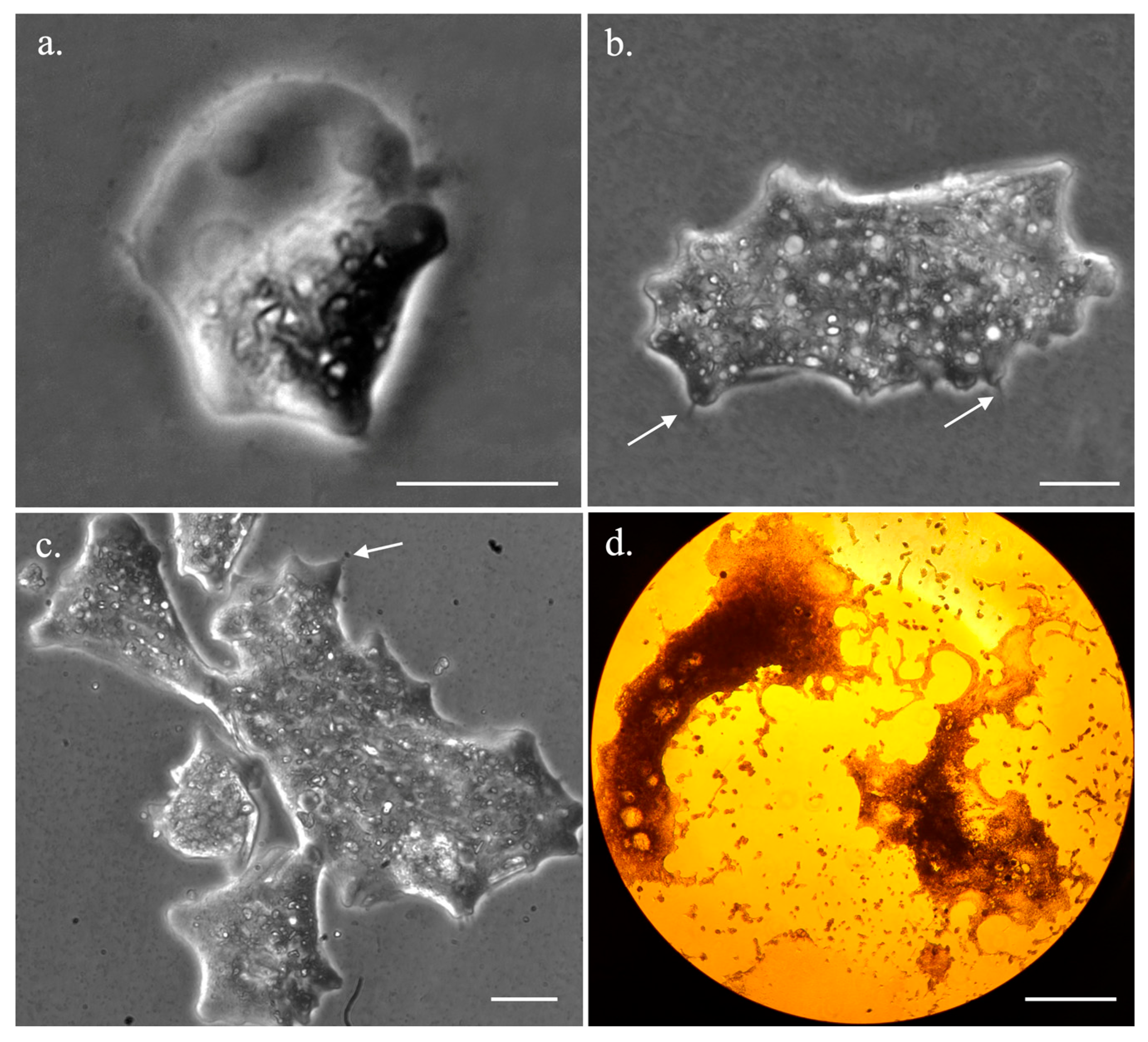
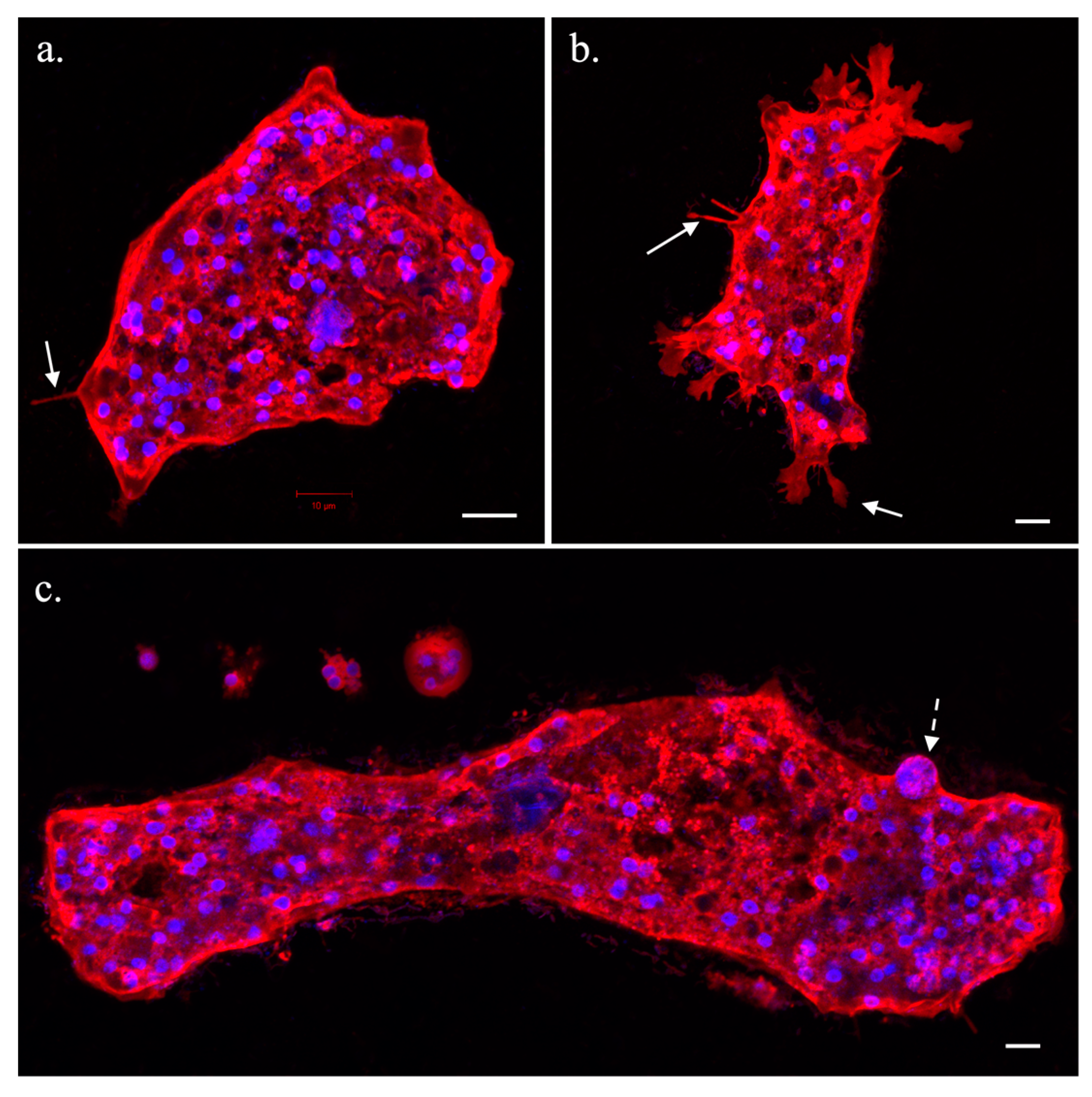
3.1. Trichophearium Life Cycle Observation
3.2. Genome Composition, Morphotype Divergences and Gene Prediction
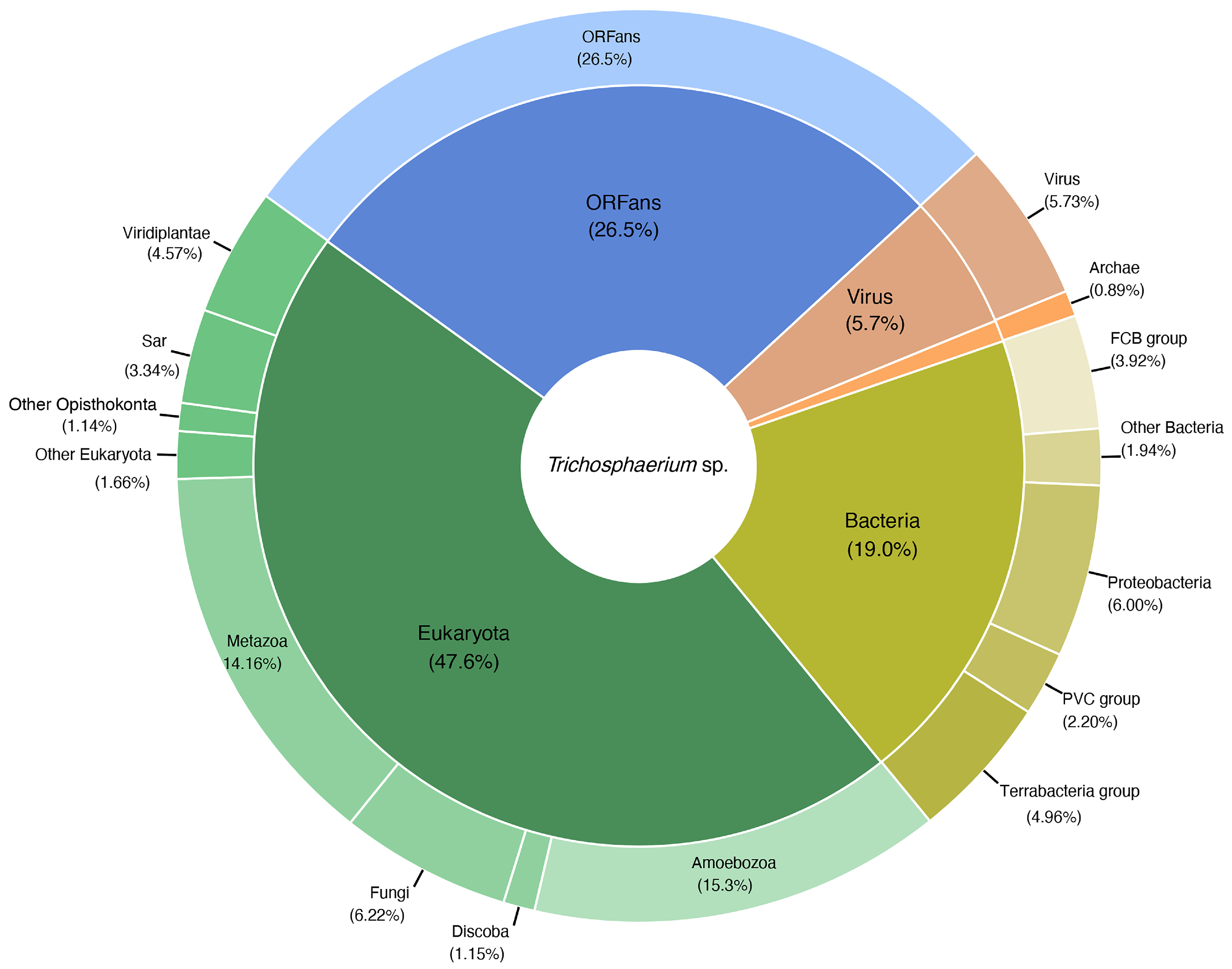
3.3. Taxonomic Distribution of Trichosphaerium sp. Predicted Gene Models
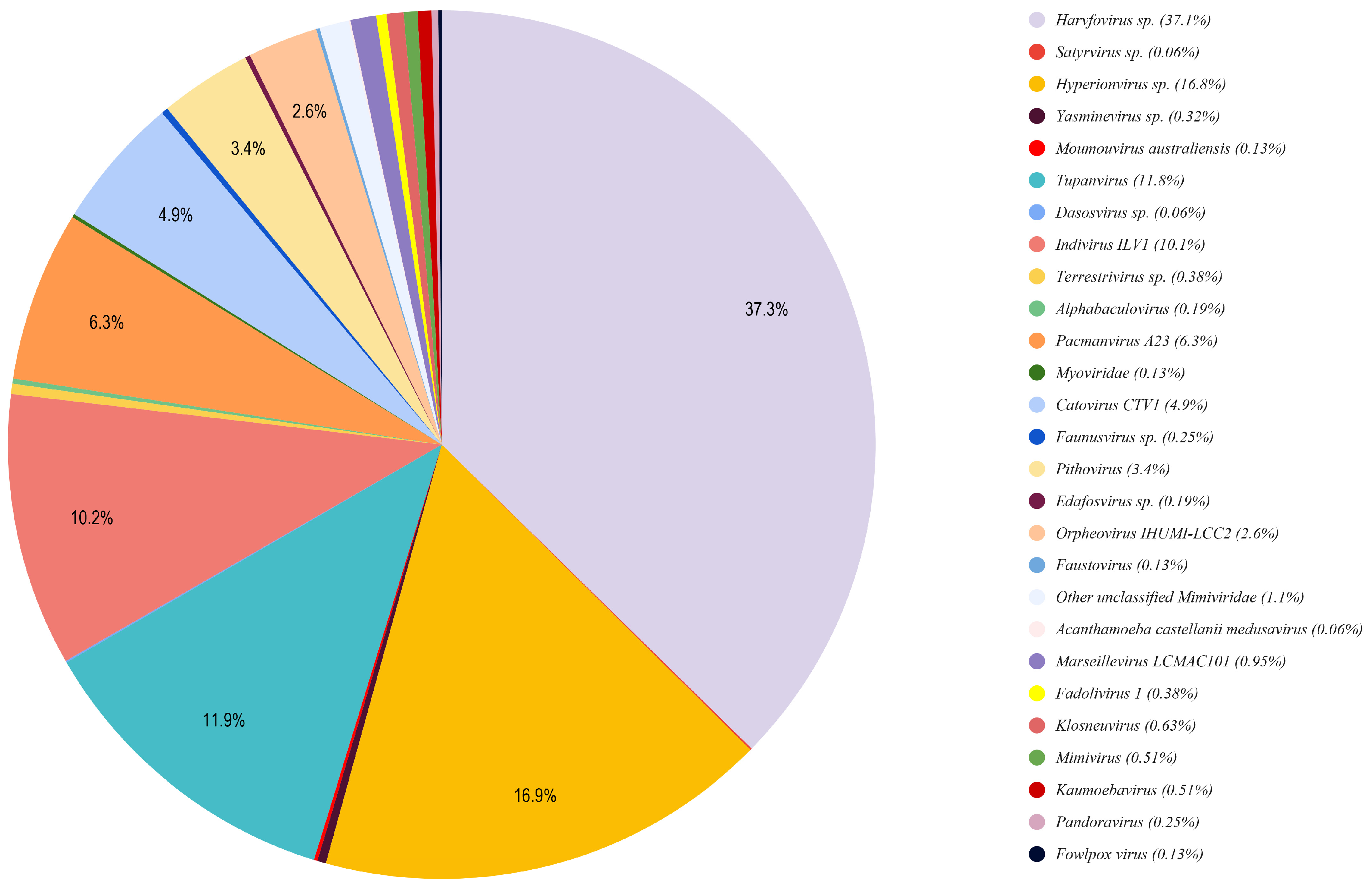
3.4. Elevated Levels of Lateral Gene Transfer (LGT) in the Trichosphaerium sp. Draft Genome
3.5. Comparison of Morphotypes Using Transcriptome Data
3.6. Genes Involved in Sexual Development of Trichosphaerium sp.
4. Discussion
4.1. Understanding Genetics of Morphological Polymorphism in Trichosphaerium sp.
4.2. Exploration of the Life Cycle of Trichosphaerium sp. Using Single Cell Transcriptomics
4.3. Variations of Sexual Genetics of Amoebozoa—SPO11 Evolution
4.4. Evidence of Elevated Gene Trafficking in Trichosphaerium sp. Genome
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loftus, B.; Anderson, I.; Davies, R.; Alsmark, U.C.M.; Samuelson, J.; Amedeo, P.; Roncaglia, P.; Berriman, M.; Hirt, R.P.; Mann, B.J.; et al. The genome of the protist parasite Entamoeba histolytica. Nature 2005, 433, 865–868. [Google Scholar] [CrossRef] [PubMed]
- Eichinger, L.; Pachebat, J.A.; Glockner, G.; Rajandream, M.A.; Sucgang, R.; Berriman, M.; Song, J.; Olsen, R.; Szafranski, K.; Xu, Q.; et al. The genome of the social amoeba Dictyostelium discoideum. Nature 2005, 435, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Carlton, J.M.; Hirt, R.P.; Silva, J.C.; Delcher, A.L.; Schatz, M.; Zhao, Q.; Wortman, J.R.; Bidwell, S.L.; Alsmark, U.C.; Besteiro, S.; et al. Draft genome sequence of the sexually transmitted pathogen Trichomonas vaginalis. Science 2007, 315, 207–212. [Google Scholar] [CrossRef]
- Eisen, J.A.; Coyne, R.S.; Wu, M.; Wu, D.; Thiagarajan, M.; Wortman, J.R.; Badger, J.H.; Ren, Q.; Amedeo, P.; Jones, K.M.; et al. Macronuclear genome sequence of the ciliate Tetrahymena thermophila, a model eukaryote. PLoS Biol. 2006, 4, e286. [Google Scholar] [CrossRef] [PubMed]
- Ellegren, H. Genome sequencing and population genomics in non-model organisms. Trends Ecol. Evol. 2014, 29, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Maumus, F.; Blanc, G. Study of Gene Trafficking between Acanthamoeba and Giant Viruses Suggests an Undiscovered Family of Amoeba-Infecting Viruses. Genome Biol. Evol. 2016, 8, 3351–3363. [Google Scholar] [CrossRef]
- Andersson, J.O. Lateral gene transfer in eukaryotes. Cell. Mol. Life Sci. 2005, 62, 1182–1197. [Google Scholar] [CrossRef]
- Williams, T.A.; Foster, P.G.; Cox, C.J.; Embley, T.M. An archaeal origin of eukaryotes supports only two primary domains of life. Nature 2013, 504, 231–236. [Google Scholar] [CrossRef]
- Alsmark, C.; Foster, P.G.; Sicheritz-Ponten, T.; Nakjang, S.; Martin Embley, T.; Hirt, R.P. Patterns of prokaryotic lateral gene transfers affecting parasitic microbial eukaryotes. Genome Biol. 2013, 14, R19. [Google Scholar] [CrossRef]
- Irwin, N.A.T.; Pittis, A.A.; Richards, T.A.; Keeling, P.J. Systematic evaluation of horizontal gene transfer between eukaryotes and viruses. Nat. Microbiol. 2022, 7, 327–336. [Google Scholar] [CrossRef]
- Ku, C.; Martin, W.F. A natural barrier to lateral gene transfer from prokaryotes to eukaryotes revealed from genomes: The 70 % rule. BMC Biol. 2016, 14, 89. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.; Brochier-Armanet, C. Giant viruses, giant chimeras: The multiple evolutionary histories of Mimivirus genes. BMC Evol. Biol. 2008, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Schaudinn, F. Untersuchungen Über Den Generationswechsel Von Trichosphaerium Sieboldi; Schn Abh Königl Preuss Akad Wiss: Berlin, Germany, 1899; 93p. [Google Scholar]
- Sheehan, R.; Banner, F.T. Trichosphaerium—An extraordinary testate Rhizopod from coastal waters. Estuar. Coast. Mar. Sci. 1973, 1, 245–260. [Google Scholar] [CrossRef]
- Polne-Fuller, M.; Rogerson, A.; Amano, H.; Gibor, A. Digestion of seaweeds by the marine amoeba Trichosphaerium. Hydrobiologia 1990, 204, 409–413. [Google Scholar] [CrossRef]
- Rogerson, A.; Williams, A.G.; Wilson, P.C. Utilization of Macroalgal Carbohydrates By The Marine Amoeba Trichosphaerium Sieboldi. J. Mar. Biol. Assoc. United Kingd 1998, 78, 733–744. [Google Scholar] [CrossRef]
- Angell, R.W. Structure of Trichosphaerium micrum sp. n. J. Protozool. 1975, 22, 18–22. [Google Scholar] [CrossRef]
- Angell, R.W. Observations On Trichosphaerium-Platyxyrum sp. n. J. Protozool. 1976, 23, 357–364. [Google Scholar] [CrossRef]
- Schuster, F.L. Fine Structure of the Schizont Stage of the Testate Marine Ameba, Trichosphaerium sp. J. Eurkaryotic Microbiol. 1976, 23, 86–93. [Google Scholar]
- Page, F.C. Marine Gymnamoebae; Institute of Terrestrial Ecology: Cambridge, UK, 1983. [Google Scholar]
- Tekle, Y.I.; Grant, J.; Anderson, O.R.; Nerad, T.A.; Cole, J.C.; Patterson, D.J.; Katz, L.A. Phylogenetic placement of diverse amoebae inferred from multigene analyses and assessment of clade stability within ‘Amoebozoa’ upon removal of varying rate classes of SSU-rDNA. Mol. Phylogenetics Evol. 2008, 47, 339–352. [Google Scholar] [CrossRef]
- Smirnov, A.V.; Chao, E.; Nassonova, E.S.; Cavalier-Smith, T. A revised classification of naked lobose amoebae (Amoebozoa: Lobosa). Protist 2011, 162, 545–570. [Google Scholar] [CrossRef]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukes, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eurkaryotic Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Tice, A.K.; Spiegel, F.W.; Silberman, J.D.; Panek, T.; Cepicka, I.; Kostka, M.; Kosakyan, A.; Alcantara, D.M.C.; Roger, A.J.; et al. Between a Pod and a Hard Test: The Deep Evolution of Amoebae. Mol. Biol. Evol. 2017, 34, 2258–2270. [Google Scholar] [CrossRef] [PubMed]
- Tekle, Y.I.; Wang, F.; Wood, F.C.; Anderson, O.R.; Smirnov, A. New Insights on the Evolutionary Relationships Between the Major Lineages of Amoebozoa. Sci. Rep. 2022, 12, 11173. [Google Scholar] [CrossRef]
- Tekle, Y.I.; Wood, F.C. Longamoebia is not monophyletic: Phylogenomic and cytoskeleton analyses provide novel and well-resolved relationships of amoebozoan subclades. Mol. Phylogenet Evol. 2017, 114, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Cavalier-Smith, T.; Chao, E.E.; Lewis, R. 187-gene phylogeny of protozoan phylum Amoebozoa reveals a new class (Cutosea) of deep-branching, ultrastructurally unique, enveloped marine Lobosa and clarifies amoeba evolution. Mol. Phylogenet Evol. 2016, 99, 275–296. [Google Scholar] [CrossRef]
- Tekle, Y.I.; Anderson, O.R.; Lecky, A.F. Evidence of parasexual activity in “asexual amoebae” Cochliopodium spp. (Amoebozoa): Extensive cellular and nuclear fusion. Protist 2014, 165, 676–687. [Google Scholar] [CrossRef]
- Tekle, Y.I.; Wang, F.; Heidari, A.; Stewart, A.J. Differential Gene Expression Analysis and Cytological Evidence Reveal a Sexual Stage of an Amoeba with Multiparental Cellular and Nuclear Fusion. PLoS ONE 2020, 15, e0235725. [Google Scholar] [CrossRef]
- Tekle, Y.I.; Wood, F.C.; Katz, L.A.; Ceron-Romero, M.A.; Gorfu, L.A. Amoebozoans Are Secretly but Ancestrally Sexual: Evidence for Sex Genes and Potential Novel Crossover Pathways in Diverse Groups of Amoebae. Genome Biol. Evol. 2017, 9, 375–387. [Google Scholar] [CrossRef]
- Wood, F.C.; Heidari, A.; Tekle, Y.I. Genetic Evidence for Sexuality in Cochliopodium (Amoebozoa). J. Hered. 2017, 108, 769–779. [Google Scholar] [CrossRef]
- Tekle, Y.I.; Wang, F.; Tran, H.; Hayes, T.D.; Ryan, J.F. The draft genome of Cochliopodium minus reveals a complete meiosis toolkit and provides insight into the evolution of sexual mechanisms in Amoebozoa. Sci. Rep. 2022, 12, 9841. [Google Scholar] [CrossRef]
- Clarke, M.; Lohan, A.J.; Liu, B.; Lagkouvardos, I.; Roy, S.; Zafar, N.; Bertelli, C.; Schilde, C.; Kianianmomeni, A.; Burglin, T.R.; et al. Genome of Acanthamoeba castellanii highlights extensive lateral gene transfer and early evolution of tyrosine kinase signaling. Genome Biol. 2013, 14, R11. [Google Scholar] [CrossRef]
- Chelkha, N.; Hasni, I.; Louazani, A.C.; Levasseur, A.; La Scola, B.; Colson, P. Vermamoeba vermiformis CDC-19 draft genome sequence reveals considerable gene trafficking including with candidate phyla radiation and giant viruses. Sci. Rep. 2020, 10, 5928. [Google Scholar] [CrossRef]
- Smirnov, A.V.; Goodkov, A.V. An illustrated list of basic morphotypes of Gymnamoebia (Rhizopoda, Lobosea). Protistology 1999, 1, 20–29. [Google Scholar]
- Page, F.C. The classification of ‘naked’ amoebae (Phylum Rhizopoda). Arch. Protistenkd. 1987, 133, 199–217. [Google Scholar] [CrossRef]
- Page, F.C. A New Key to Freshwater and Soil Gymnamoebae; Freshwater Biological Association: Ambleside, Cumbria, UK, 1988. [Google Scholar]
- Nassonova, E.; Smirnov, A.; Fahrni, J.; Pawlowski, J. Barcoding amoebae: Comparison of SSU, ITS and COI genes as tools for molecular identification of naked lobose amoebae. Protist 2010, 161, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Tekle, Y.I. DNA barcoding in amoebozoa and challenges: The example of Cochliopodium. Protist 2014, 165, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Tekle, Y.I.; Wood, F.C. A practical implementation of large transcriptomic data analysis to resolve cryptic species diversity problems in microbial eukaryotes. BMC Evol. Biol. 2018, 18, 170. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, G. Atypical ploidy cycles, Spo11, and the evolution of meiosis. Semin. Cell. Dev. Biol. 2016, 54, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Maciver, S.K.; Koutsogiannis, Z.; de Obeso Fernandez Del Valle, A. ‘Meiotic genes’ are constitutively expressed in an asexual amoeba and are not necessarily involved in sexual reproduction. Biol. Lett. 2019, 15, 20180871. [Google Scholar] [CrossRef]
- Erdos, G.W.; Raper, K.B.; Vogen, L.K. Mating Types And Macrocyst Formation In Dictyostelium-Discoideum. Proc. Natl. Acad. Sci. USA 1973, 70, 1828–1830. [Google Scholar] [CrossRef]
- Goodfellow, L.P.; Belcher, J.H.; Page, F.C. A light- and electron-microscopical study of Sappinia diploidea, a sexual amoeba. Protistologica 1974, 2, 207–216. [Google Scholar]
- Schensnovich, V.B. On the life cycle of Entamoeba histolytica. Med. Parazitol. 1967, 36, 712–715. [Google Scholar]
- Malik, S.B.; Ramesh, M.A.; Hulstrand, A.M.; Logsdon, J.M., Jr. Protist homologs of the meiotic Spo11 gene and topoisomerase VI reveal an evolutionary history of gene duplication and lineage-specific loss. Mol. Biol. Evol. 2007, 24, 2827–2841. [Google Scholar] [CrossRef]
- Katz, L.A. Lateral gene transfers and the evolution of eukaryotes: Theories and data. Int. J. Syst. Evol. Microbiol. 2002, 52, 1893–1900. [Google Scholar]
- de Koning, A.P.; Brinkman, F.S.L.; Jones, S.J.M.; Keeling, P.J. Lateral gene transfer and metabolic adaptation i the human parasite Trichomonas vaginalis. Mol. Biol. Evol. 2000, 17, 1769–1773. [Google Scholar] [CrossRef] [PubMed]
- Brueckner, J.; Martin, W.F. Bacterial Genes Outnumber Archaeal Genes in Eukaryotic Genomes. Genome Biol. Evol. 2020, 12, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Audic, S.; Robert, C.; Abergel, C.; Renesto, P.; Ogata, H.; La Scola, B.; Suzan, M.; Claverie, J.M. The 1.2-megabase genome sequence of Mimivirus. Science 2004, 306, 1344–1350. [Google Scholar] [CrossRef]
- La Scola, B.; Audic, S.; Robert, C.; Jungang, L.; de Lamballerie, X.; Drancourt, M.; Birtles, R.; Claverie, J.M.; Raoult, D. A giant virus in amoebae. Science 2003, 299, 2033. [Google Scholar] [CrossRef]
- Schulz, F.; Roux, S.; Paez-Espino, D.; Jungbluth, S.; Walsh, D.A.; Denef, V.J.; McMahon, K.D.; Konstantinidis, K.T.; Eloe-Fadrosh, E.A.; Kyrpides, N.C.; et al. Giant virus diversity and host interactions through global metagenomics. Nature 2020, 578, 432–436. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Weinheimer, A.R.; Martinez-Gutierrez, C.A.; Aylward, F.O. Widespread endogenization of giant viruses shapes genomes of green algae. Nature 2020, 588, 141–145. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, L.D.; Young, E.; Delaney, J.; Ruhnau, F.; Ritchie, K.B.; Paul, J.H. High frequency of horizontal gene transfer in the oceans. Science 2010, 330, 50. [Google Scholar] [CrossRef]
- Mihara, T.; Koyano, H.; Hingamp, P.; Grimsley, N.; Goto, S.; Ogata, H. Taxon Richness of “Megaviridae” Exceeds those of Bacteria and Archaea in the Ocean. Microbes Environ. 2018, 33, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Forterre, P.; Prangishvili, D. The major role of viruses in cellular evolution: Facts and hypotheses. Curr. Opin. Virol. 2013, 3, 558–565. [Google Scholar] [CrossRef]
- Nasir, A.; Kim, K.M.; Caetano-Anolles, G. Giant viruses coexisted with the cellular ancestors and represent a distinct supergroup along with superkingdoms Archaea, Bacteria and Eukarya. BMC Evol. Biol. 2012, 12, 156. [Google Scholar] [CrossRef] [PubMed]
- Gornik, S.G.; Ford, K.L.; Mulhern, T.D.; Bacic, A.; McFadden, G.I.; Waller, R.F. Loss of nucleosomal DNA condensation coincides with appearance of a novel nuclear protein in dinoflagellates. Curr. Biol. 2012, 22, 2303–2312. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.A.; Feschotte, C. Co-option of endogenous viral sequences for host cell function. Curr. Opin. Virol. 2017, 25, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, G.; Funk, M.; Vernochet, C.; Leal, F.; Tarazona, O.A.; Meurice, G.; Heidmann, O.; Dupressoir, A.; Miralles, A.; Ramirez-Pinilla, M.P.; et al. An endogenous retroviral envelope syncytin and its cognate receptor identified in the viviparous placental Mabuya lizard. Proc. Natl. Acad. Sci. USA 2017, 114, E10991–E11000. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Trichosphaerium |
|---|---|
| Genome size (bp) | 70,866,254 |
| GC content (%) | 37.69 |
| DNA scaffolds | 710 |
| Longest scaffold length (bp) | 1,255,013 |
| Shortest scaffold length (bp) | 1087 |
| Mean scaffold length (bp) | 99,812 |
| N50 (bp) | 262,981 |
| Total number of predicted transcripts | 27,369 |
| Proportion of transcripts with a size ≧300 bp | 26,430 |
| Genes assigned to Cluster Orthologous Groups (COGs) | 14,927 |
| Non-ORFan genes | 20,515 |
| ORFan genes | 7489 |
| Mean number of introns/gene | 8.7 |
| Mean number of exons/gene | 7.7 |
| Mean intron size (bp) | 75.4 |
| Mean exon size (bp) | 154.3 |
| Gene model BUSCO completeness (complete + partial) | 93.3% |
| Gene | Trichophaerium Gene ID | Small YT42 | Small YT43 | Small YT44 | Medium YT45 | Medium YT46 | Medium YT47 | Large YT48 | Large YT49 |
|---|---|---|---|---|---|---|---|---|---|
| Meiosis-specific | |||||||||
| SPO11-a | TRSP29784 | − | − | − | − | − | − | + | − |
| SPO11-b | TRSP6954 | − | − | − | − | − | − | − | − |
| DMC1 | TRSP36706 | − | − | − | − | + | − | − | − |
| HOP1 | TRSP9892 | − | − | − | − | − | − | − | + |
| HOP2 | TRSP22418 | − | − | − | + | + | + | + | + |
| MER3 | TRSP9895 | − | − | − | − | − | − | − | − |
| MND1 | TRSP36706 | − | − | − | − | + | + | + | + |
| MSH4 | TRSP24797 | − | − | − | − | − | − | − | − |
| MSH5 | TRSP39996 | − | − | − | + | − | + | − | − |
| ZIP1 | TRSP5893 | − | − | − | + | + | + | + | − |
| PCH2 | TRSP13443 | + | − | − | − | + | + | + | − |
| REC8 | TRSP31982 | − | − | − | − | + | + | + | + |
| HAP2 | TRSP10046 | − | − | − | + | + | + | + | + |
| ZIP4-a | TRSP31710 | − | − | − | − | − | − | + | − |
| ZIP4-b | TRSP29703 | − | − | − | − | + | + | + | + |
| Plasmogamy | |||||||||
| PRM1-a | TRSP15481 | − | − | − | + | + | + | − | − |
| KEX2 | TRSP14824 | − | − | − | − | + | + | + | + |
| CD9 | TRSP23326 | + | − | + | + | + | + | + | + |
| TPM1 | TRSP40945 | + | − | − | + | − | + | + | + |
| MYO2 | TRSP10218 | − | − | − | − | − | − | − | − |
| BNI1 | TRSP7161 | − | − | − | − | − | + | + | + |
| RVS161 | TRSP41670 | + | − | − | + | + | + | + | + |
| Karyogamy/Nuclear Congression | |||||||||
| KAR2 | TRSP20746 | + | − | − | + | + | + | + | + |
| CINI1-a | TRSP22439 | − | − | − | − | − | + | − | − |
| CINI2-a | TRSP25341 | + | − | − | + | + | + | + | + |
| KAR4 | TRSP2890 | + | − | − | + | − | + | + | + |
| SEC63 | TRSP27336 | − | − | − | + | + | − | + | + |
| BIK1-a-1 | TRSP6231 | − | − | − | + | + | + | + | + |
| BIK1-a-2 | TRSP44203 | + | − | − | + | + | + | + | + |
| CIN4 | TRSP41855 | − | − | − | + | + | + | + | + |
| KAR3 | TRSP32958 | − | − | − | − | + | − | − | + |
| SEC72-a | TRSP29205 | − | − | + | + | − | + | + | + |
| CDC4 | TRSP9903 | + | − | + | + | + | + | + | + |
| CDC34 | TRSP9743 | + | − | − | + | + | + | + | + |
| JEM1-a | TRSP11422 | − | − | − | − | − | − | − | + |
| CDC28 | TRSP6885 | + | − | + | + | + | + | + | + |
| KEM1 | TRSP9215 | − | − | − | − | + | − | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tekle, Y.I.; Tran, H.; Wang, F.; Singla, M.; Udu, I. Omics of an Enigmatic Marine Amoeba Uncovers Unprecedented Gene Trafficking from Giant Viruses and Provides Insights into Its Complex Life Cycle. Microbiol. Res. 2023, 14, 656-672. https://doi.org/10.3390/microbiolres14020047
Tekle YI, Tran H, Wang F, Singla M, Udu I. Omics of an Enigmatic Marine Amoeba Uncovers Unprecedented Gene Trafficking from Giant Viruses and Provides Insights into Its Complex Life Cycle. Microbiology Research. 2023; 14(2):656-672. https://doi.org/10.3390/microbiolres14020047
Chicago/Turabian StyleTekle, Yonas I., Hanh Tran, Fang Wang, Mandakini Singla, and Isimeme Udu. 2023. "Omics of an Enigmatic Marine Amoeba Uncovers Unprecedented Gene Trafficking from Giant Viruses and Provides Insights into Its Complex Life Cycle" Microbiology Research 14, no. 2: 656-672. https://doi.org/10.3390/microbiolres14020047