
Pharmacological Effects of Lactobacillus casei ATCC 7469 Fermented Soybean and Green Microalgae, Chlorella vulgaris, on Diabetic Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Preparation of the Soy Flour Fermentation Extracts
2.3. Algae Cultivation
2.4. Preparation of Chlorella vulgaris Extract
2.5. Experimental Design for Rats Feeding with Lactobacillus Casei ATCC 7469 Fermented Soy Flour Extract and Chlorella vulgaris Extract
2.5.1. Animals
2.5.2. Termination of the Experiment
2.5.3. Blood Sampling and Biochemical Parameters
2.5.4. Oral Glucose Tolerance Test (OGTT)
2.5.5. Histopathological Examination
2.6. Statistical Analyses
3. Results and Discussion
3.1. Effect on Lipid Profile
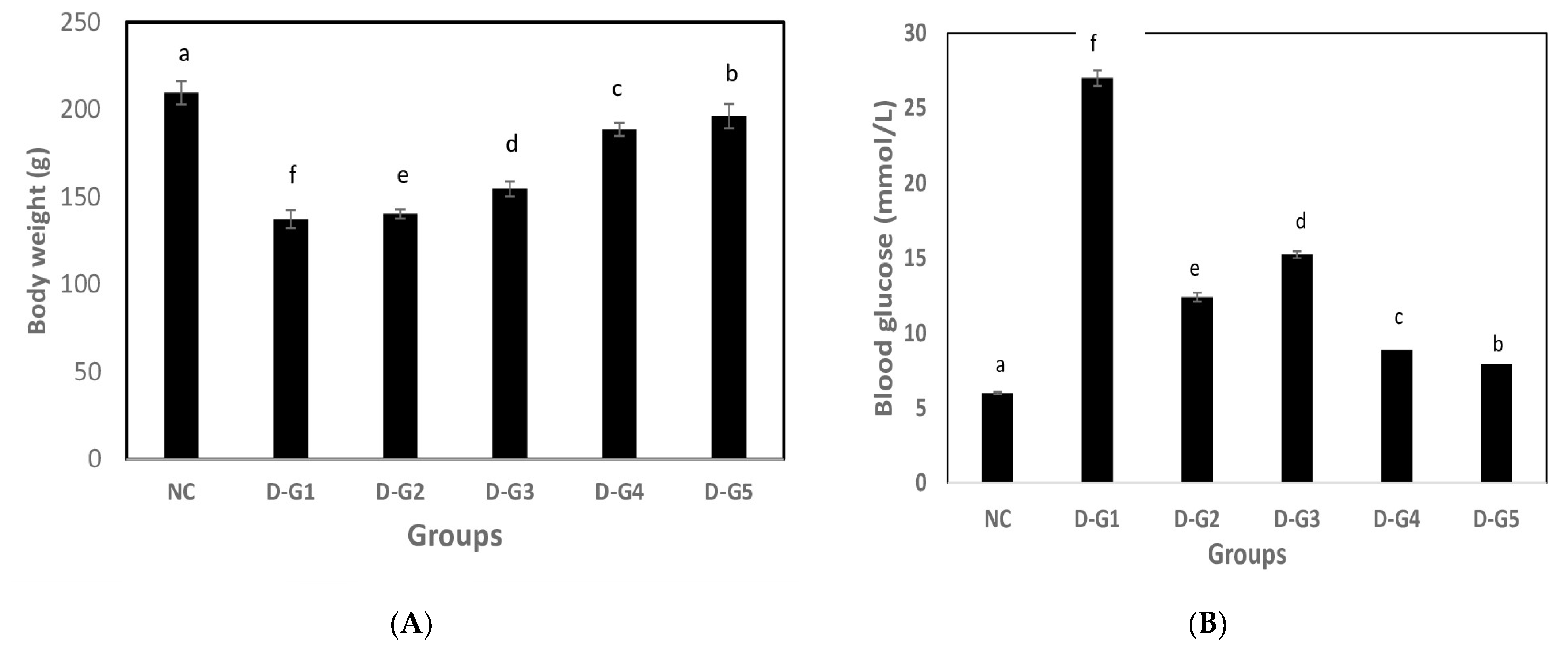
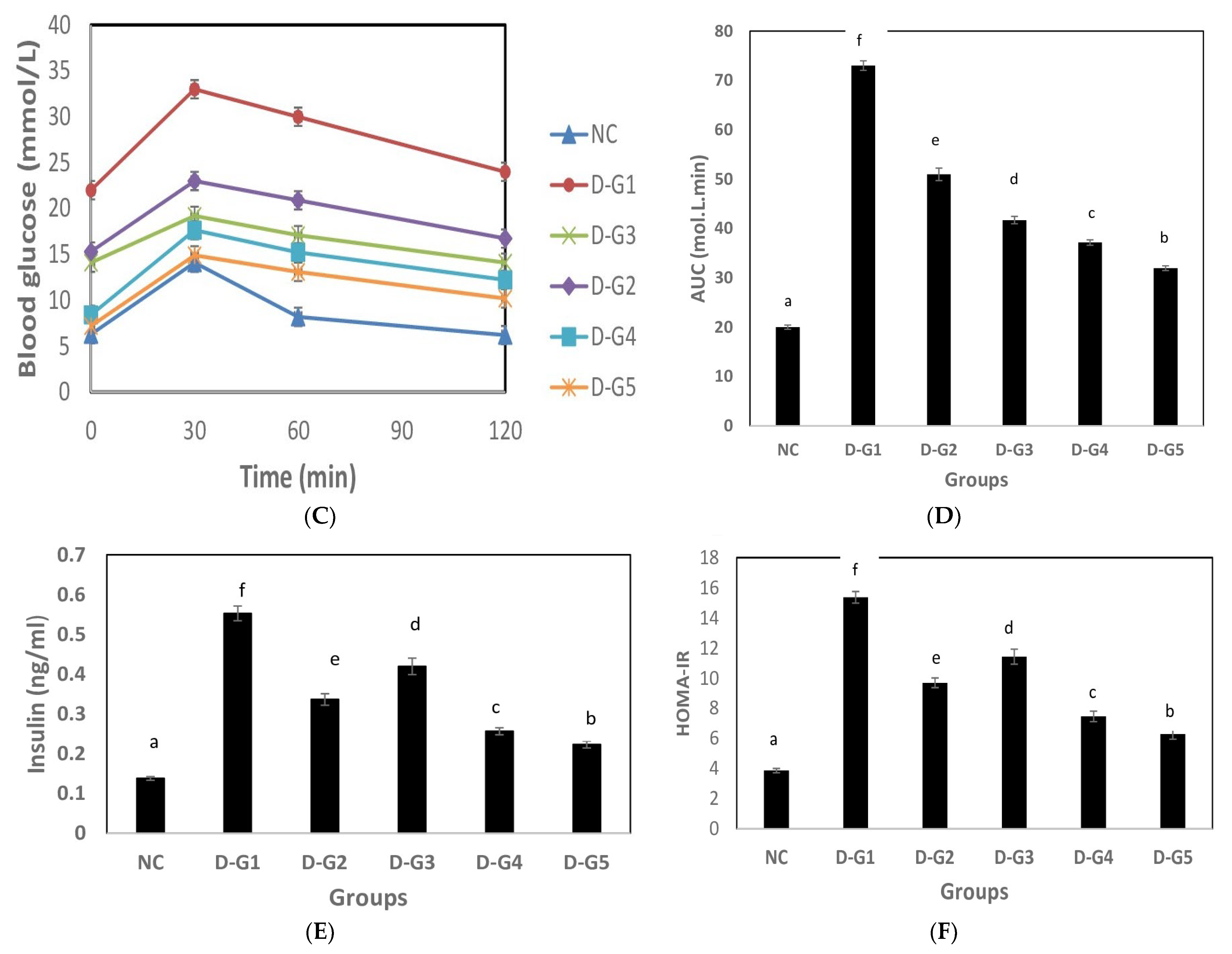
3.2. Hypoglycemic Effect
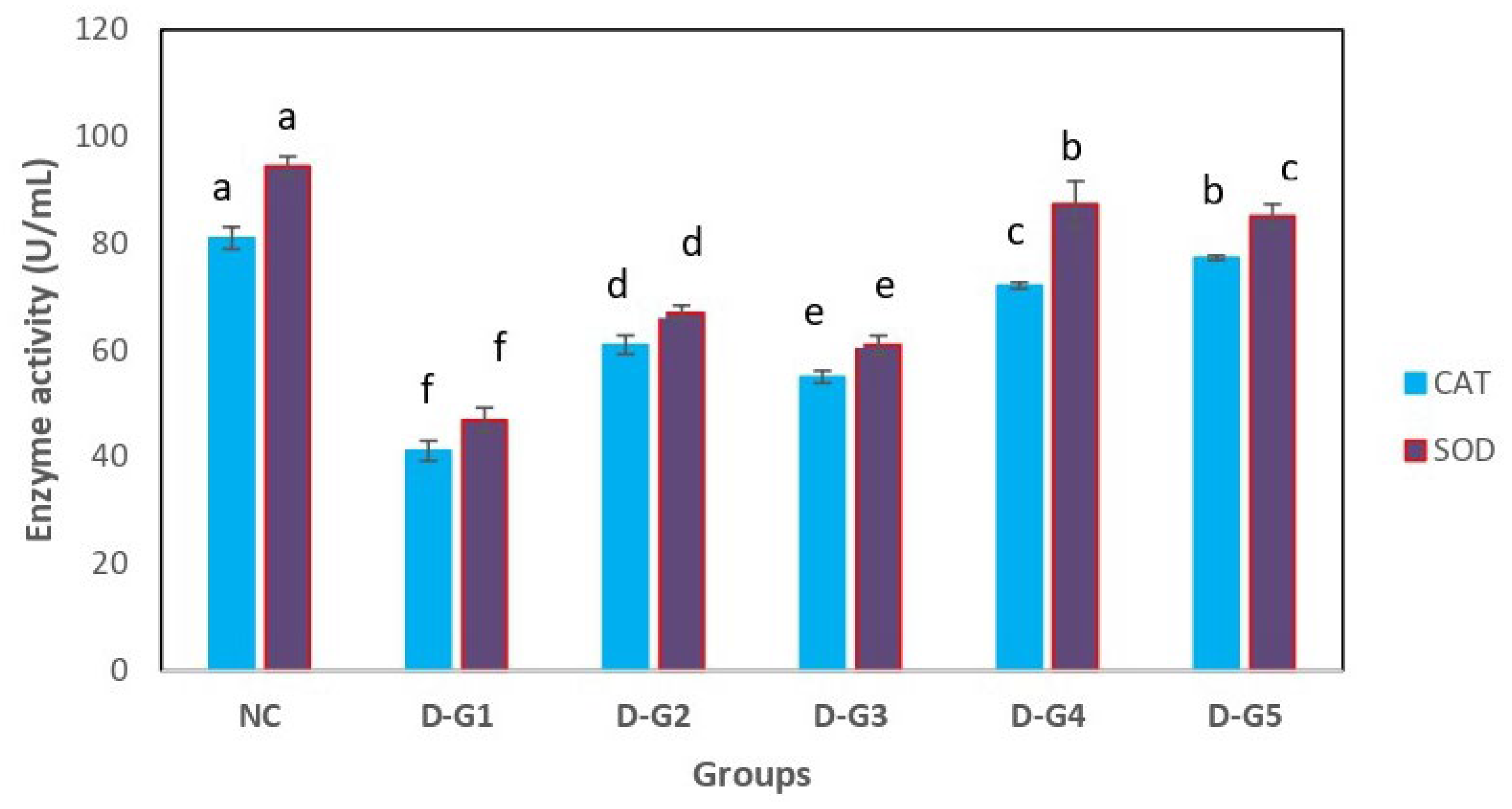
3.3. Effect on Antioxidant Parameters
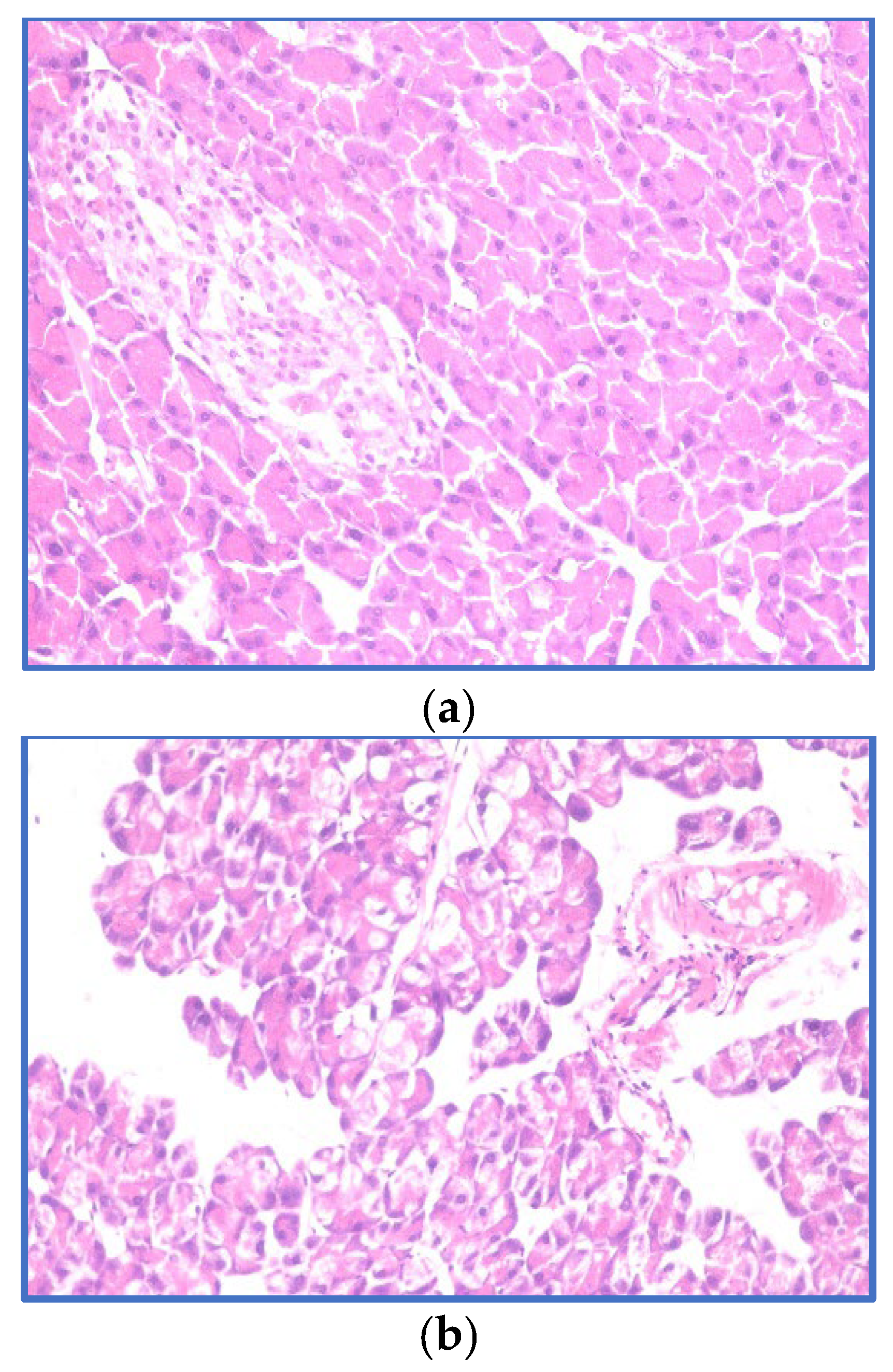
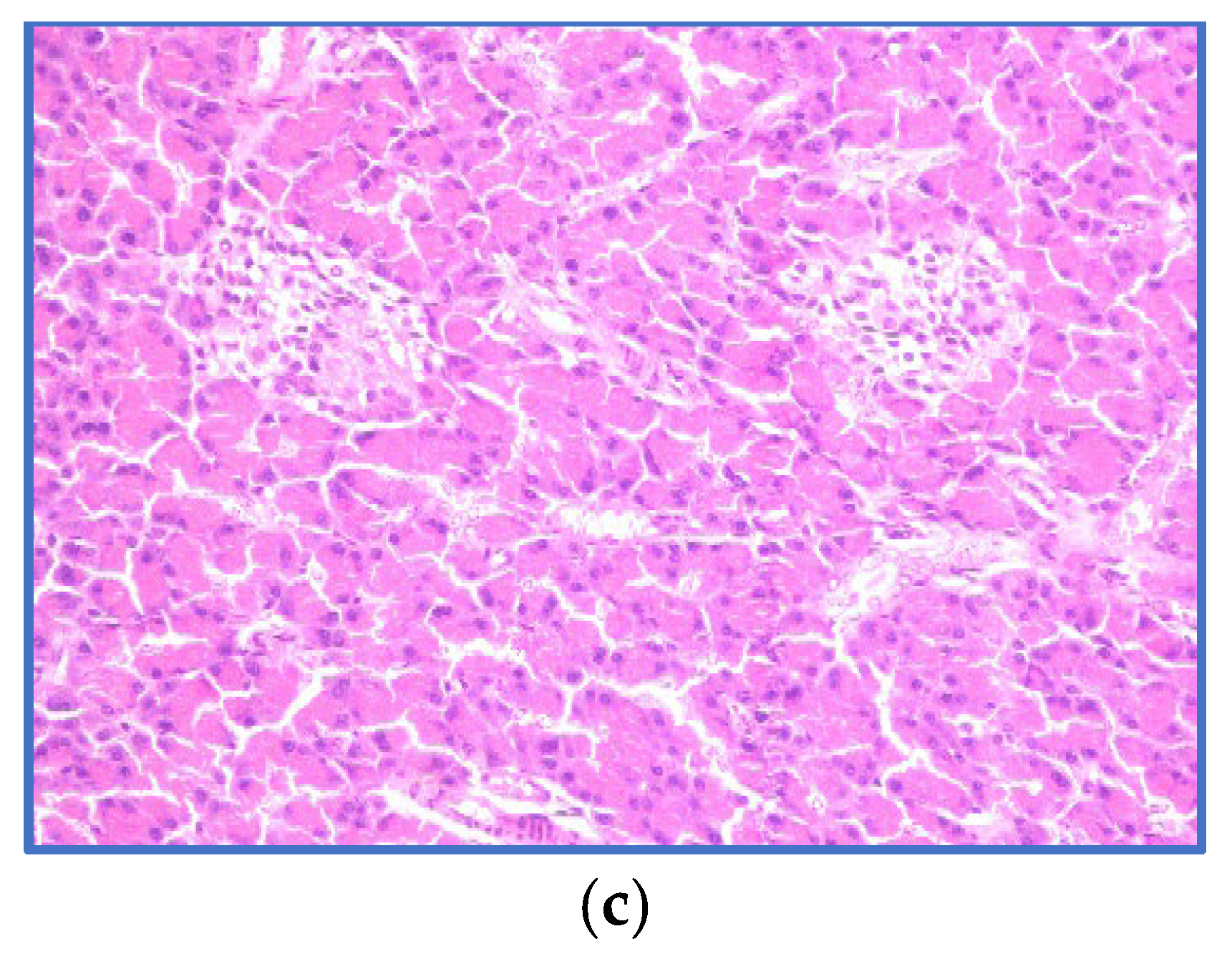
3.4. Histopathological Examination of the Pancreas
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takeda, Y.; Fujita, Y.; Honjo, J.; Sakagami, H.; Takiyama, Y.; Makino, Y.; Abiko, A.; Kieffer, T.J.; Haneda, M. Reduction of both beta cell death and alpha cell proliferation by dipeptidyl peptidase-4 inhibition in a streptozotocin-induced model of diabetes in mice. Diabetologia 2012, 55, 404. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jiang, Z.; Guo, C. New hope for type 2 diabetics: Targeting insulin resistance through the immune modulation of stem cells. Autoimmun Rev. 2011, 11, 137–142. [Google Scholar] [CrossRef]
- Mcclean, P.L.; Vadivel, P.; Emilie, F.; Christian, H.L. The diabetes drug liraglutide prevents degenerative processes in a mouse model of Alzheimer’s disease. J. Neurosci. 2011, 31, 6587–6594. [Google Scholar] [CrossRef] [PubMed]
- Sola, D.; Rossi, L.; Schianca, G.P.C.; Maffioli, P.; Bigliocca, M. Sulfonylureas and their use in clinical practice. Arch. Med. Sc. 2015, 4, 840–848. [Google Scholar] [CrossRef]
- Rizos, C.V.; Elisaf, M.; Mikhailidis, D.P.; Liberopoulos, E.N. How safe is the use of thiazolidinediones in clinical practice? Expert Opin. Drug Saf. 2009, 8, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Abd El Latif, M.A.; Mohamed, N.H.; Zaki, N.L.; Abbas, M.S.; Sobhy, H.M. Effects of Soybean Isoflavone on Lipid Profiles and Antioxidant Enzyme Activity in Streptozotocin Induced Diabetic Rats. Glob. J. Pharmacol. 2014, 8, 378–384. [Google Scholar]
- Chang, J.H.; Kim, M.S.; Kim, T.W.; Lee, S.S. Effects of soybean supplementation on blood glucose, plasma lipid levels, and erythrocyte antioxidant enzyme activity in type 2 diabetes mellitus patients. Nutr. Res. Pract. 2008, 2, 152–157. [Google Scholar] [CrossRef]
- Shu, G.; Shi, X.; Chen, H.; Ji, Z.; Meng, J. Optimization of nutrient composition for producing ACE inhibitory peptides from goat milk fermented by Lactobacillus bulgaricus LB6, Probiotics Antimicrob. Proteins 2019, 11, 723–729. [Google Scholar]
- Wagner, J.D.; Zhang, L.; Shadoan, M.K.; Kavanagh, K.; Chen, H.; Tresnasari, K.; Kaplan, J.R.; Adams, M.R. Effects of soy protein and isoflavones on insulin resistance and adiponectin in male monkeys. Metabolism 2008, 57, 24–31. [Google Scholar] [CrossRef]
- Sanjukta, S.; Rai, A.K.; Muhammed, A.; Jeyaram, K.; Talukdar, N.C. Enhancement of antioxidant properties of two soybean varieties of Sikkim Himalayan region by proteolytic Bacillus subtilis fermentation. J. Funct. Foods 2015, 14, 650–658. [Google Scholar] [CrossRef]
- Kim, J.H.; Jia, Y.; Lee, J.G.; Nam, B.; Lee, J.H.; Shin, K.S.; Hurh, B.S.; Choi, Y.H.; Lee, S.J. Hypolipidemic and antiinflammation activities of fermented soybean fibers from meju in C57BL/6 J mice. Phytother. Res. 2014, 28, 1335–1341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Wu, Y.F.; Wang, Y.S.; Wang, X.Z.; Piao, C.H.; Liu, J.M.; Liu, Y.L.; Wang, Y.H. The protective effects of probiotic-fermented soymilk on high fat diet-induced hyperlipidemia and liver injury. J. Funct. Foods 2017, 30, 220–227. [Google Scholar] [CrossRef]
- Long, X.S.; Liao, S.T.; Li, E.N.; Pang, D.R.; Li, Q.; Liu, S.C.; Hu, T.G.; Zou, Y.X. The hypoglycemic effect of freeze-dried fermented mulberry mixed with soybean on type 2 diabetes mellitus. Food Sci. Nutr. 2021, 9, 3641–3654. [Google Scholar] [CrossRef]
- Yu, S.; Wang, W.; Li, S.; Li, J.; Zhao, R.; Liu, D.; Wu, J. Glucoregulatory Properties of Fermented Soybean Products. Fermentation 2023, 9, 254. [Google Scholar] [CrossRef]
- Hu, T.; Chen, R.; Qian, Y.; Ye, K.; Long, X.; Park, K.Y.; Zhao, X. Antioxidant effect of Lactobacillus fermentum HFY02-fermented soy milk on D-galactose-induced aging mouse mode. Food Sci. Hum. Wellness 2022, 11, 1362–1372. [Google Scholar] [CrossRef]
- Stramarkou, M.; Papadaki, S.; Kyriakopoulou, K.; Krokida, M. Effect of drying and extraction conditions on the recovery of bioactive compounds from Chlorella vulgaris. J. Appl. Phycol. 2017, 29, 2947–2960. [Google Scholar] [CrossRef]
- Bito, T.; Okumura, E.; Fujishima, M.; Watanabe, F. Potential of Chlorella as a Dietary Supplement to Promote Human Health. Nutrients 2020, 12, 2524. [Google Scholar] [CrossRef]
- Daugherty, B.L. Histamine H4 antagonism: A therapy for chronic allergy? Br. J. Pharmacol. 2014, 142, 5. [Google Scholar] [CrossRef]
- Lee, H.S.; Park, H.J.; Kim, M.K. Effect of Chlorella vulgaris on lipid metabolism in Wistar rats fed high fat diet. Nutr. Res. Pract. 2008, 2, 204–210. [Google Scholar] [CrossRef]
- Jeong, H.; Kwon, H.J.; Kim, M.K. Hypoglycemic effect of Chlorella vulgaris intake in type 2 diabetic Goto-Kakizaki and normal Wistar rats. Nutr. Res. Pract. 2009, 3, 23–30. [Google Scholar] [CrossRef]
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae: Identification, Enumeration and Use as Bioindicators, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Stainer, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (Order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Dong, J.; Liang, Q.; Niu, Y.; Jiang, S.; Zhou, L.I.; Wang, J.; Ma, C.; Kang, W. Effects of nigella sativa seed polysaccharides on type 2 diabetic mice and gut microbiota. Int. J. Biol. Macromol. 2020, 159, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Choi, H. Dietary red chilli (Capsicum frutescens L.) is insulinotropic rather than hypoglycemic in type 2 diabetes model of rats. Phytother. Res. 2008, 22, 1025–1029. [Google Scholar] [CrossRef] [PubMed]
- Assinewe, V.A.; Baum, B.R.; Gagnon, D.; Arnason, J.T. Phytochemistry of Wild Populations of Panax quinquefolius L. (North American Ginseng). J. Agric. Food Chem. 2003, 51, 4549–4553. [Google Scholar] [CrossRef]
- Fossati, P.; Prencipe, L. Serum triglycerides are determined colorimetrically with a Nenzyme that produces hydrogen peroxide. Clin. Chem. 1982, 28, 2077–2080. [Google Scholar] [CrossRef] [PubMed]
- Deeg, R.; Ziegenohrm, J. Kinetic enzymatic method for automated determination of total cholesterol in serum. J. Clin. Chem. 1983, 29, 1798–1802. [Google Scholar] [CrossRef]
- Burstein, M.; Selvenick, H.R.; Morfin, R. Rapid method for the isolation of lipoproteins from human serum by precipitation with polyanions. J. Lipid Res. 1970, 11, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Norbert, W.T. Clinical Guide to Laboratory Tests, 3rd ed.; W.B. Saunders Company: Philadelphia, PA, USA, 1995. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Cohen, G.; Dembiec, D.; Marcus, J. Measurement of catalase activity in tissue. Anal. Biochem. 1970, 34, 30–38. [Google Scholar] [CrossRef]
- Bancroft, G.D.; Stevens, A.; Turner, D.R. Theory and Practice of Pathological Technique, 4th ed.; Churchill Livingston: New York, NY, USA, 1996. [Google Scholar]
- Snedcor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; The Iowa State University Press: Ames, IA, USA, 1982; p. 507. [Google Scholar]
- Zhao, J.; Wu, Y.Y.; Rong, X.L.; Zheng, C.W.; Guo, J. Anti-Lipolysis Induced by Insulin in Diverse Pathophysiologic Conditions of Adipose Tissue. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Prangthipa, P.; Surasianga, R.; Charoensiria, R.; Leardkamolkarn, V.; Komindr, S.; Yamborisuta, U.; Vanavichitd, A.; Kongkachuichai, R. Amelioration of hyperglycemia, hyperlipidemia, oxidative stress and inflammation in steptozotocin-induced diabetic rats fed a high fat diet by riceberry supplement. J. Funct. Foods 2013, 5, 195–203. [Google Scholar] [CrossRef]
- Cherng, J.Y.; Shih, M.F. Potential hypoglycemic effects of Chlorella in streptozotocin-induced diabetic mice. Life Sci. 2005, 78, 980–990. [Google Scholar]
- Zhao, C.; Wu, Y.J.; Yang, C.F.; Liu, B.; Huang, Y.F. Hypotensive, hypoglycemic and hypolipidemic effects of bioactive compounds from microalgae and marine microorganisms. Int. J. Food Sci. Technol. 2015, 50, 1705–1717. [Google Scholar] [CrossRef]
- Chen, J.; Gong, S.; Wan, X.; Gao, X.; Wang, C.; Zeng, F.; Zhao, C.; Liu, B.; Huang, Y. Hypolipidemic properties of Chlorella pyrenoidosa organic acids via AMPK/HMGCR/SREBP-1c pathway in vivo. Food Sci. Nutr. 2021, 9, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, F.; Marini, H.; Bitto, A.; Altavilla, D.; Polito, F.; Adamo, E.B.; D’Anna, R.; Arcoraci, V.; Burnett, B.P.; Minutoli, L.; et al. Genistein in the metabolic syndrome: Results of a randomized clinical trial. J. Clin. Endocrinol. Metab. 2013, 98, 3366–3374. [Google Scholar] [CrossRef]
- Moradi, M.; Daneshzad, E.; Azadbakht, L. The effects of isolated soy protein, isolated soy isoflavones and soy protein containing isoflavones on serum lipids in postmenopausal women: A systematic review and meta-analysis. Crit. Rev. Food Sci. Nutr. 2020, 60, 3414–3428. [Google Scholar] [CrossRef]
- Singh, B.P.; Vij, S.; Hati, S. Functional significance of bioactive peptides derived from soybean. Peptides 2014, 54, 171–179. [Google Scholar] [CrossRef]
- Wang, J.Y.; Dang, N.; Sun, P.; Xia, J.; Zhang, C.; Pang, S. +e effects of metformin on fibroblast growth factor 19, 21 and fibroblast growth factor receptor 1 in high-fat diet and streptozotocin induced diabetic rats. Endocr. J. 2017, 64, 543–552. [Google Scholar] [CrossRef]
- Tsai, C.C.; Lin, P.-P.; Hsieh, Y.-M.; Zhang, Z.-Y.; Wu, H.-C.; Huang, C.-C. Cholesterol-Lowering Potentials of Lactic Acid Bacteria Based on Bile-Salt Hydrolase Activity and Effect of Potent Strains on Cholesterol Metabolism In Vitro and In Vivo. Sci. World J. 2014, 2014, 690752. [Google Scholar] [CrossRef]
- Hamouda, R.A.; Hamza, H.A.; Salem, M.L.; Kamal, S.; Alhasani, R.H.; Alsharif, I.; Mahrous, H.; Abdella, A. Synergistic Hypolipidemic and Immunomodulatory Activity of Lactobacillus and Spirulina platensis. Fermentation 2022, 8, 220. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y.L. Bile acid metabolism and signaling in liver disease and therapy. Liver Res. 2017, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Pegah, A.; Abbasi-Oshaghi, E.; Khodadadi, I.; Mirzaei, F.; Tayebina, H. Probiotic, and resveratrol normalize GLP-1 levels and oxidative stress in the intestine of diabetic rats. Metab. Open 2021, 10, 100093. [Google Scholar] [CrossRef] [PubMed]
- Gayoso-Diz, P.; Otero-González, A.; Rodriguez-Alvarez, M.X.; Gude, F.; García, F.; De Francisco, A. Insulin resistance (HOMA-IR) cut-off values and the metabolic syndrome in a general adult population: Effect of gender and age: EPIRCE cross-sectional study. BMC Endocr. Disord. 2013, 13, 47. [Google Scholar] [CrossRef]
- Vecina, J.F.; Oliveira, A.; Araújo, T.; Baggio, S.R.; Torello, C.O.; Saad, M.J.A.; Queiroz, M.L.D.S. Chlorella modulates insulin signaling pathway and prevents high-fat diet-induced insulin resistance in mice. Life Sci. 2014, 95, 45–52. [Google Scholar] [CrossRef]
- Noriega-Lopez, L.; Tovar, A.R.; Gonzalez-Granillo, M.; Hernandez-Pando, R.; Escalante, B.; Santillan-Doherty, P.; Torres, N. Pancreatic insulin secretion in rats fed a soy protein high fat diet depends on the interaction between the amino acid pattern and isoflavones. J. Biol. Chem. 2007, 282, 20657–20666. [Google Scholar] [CrossRef]
- Liu, D.; Zhen, W.; Yang, Z.; Carter, J.D.; Si, H.; Reynolds, K.A. Genistein acutely stimulates insulin secretion in pancreatic β-cells through cAMP-dependent protein kinase pathway. Daibetes 2006, 55, 1043–1050. [Google Scholar] [CrossRef]
- Rasouli, H.; Hosseini-Ghazvini, S.M.; Adibi, H.; Khodarahmi, R. Differential α-amylase/α-glucosidase inhibitory activities of plant-derived phenolic compounds: A virtual screening perspective for the treatment of obesity and diabetes. Food Funct. 2017, 8, 1942–1954. [Google Scholar] [CrossRef]
- Gomes, A.C.; Bueno, A.A.; de Souza RG, M.; Mota, J.F. Gut microbiota, probiotics, and diabetes. Nutr. J. 2014, 13, 60. [Google Scholar] [CrossRef]
- Ma, X.; Hua, J.; Li, Z. Probiotics improve high fat diet-induced hepatic steatosis and insulin resistance by increasing hepatic NKT cells. J. Hepatol. 2008, 49, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, H.; Mahmood, N.; Kumar, M.; Varikuti, S.R.; Challa, H.R.; Myakala, S.P. Effect of probiotic (VSL# 3) and omega-3 on lipid profile, insulin sensitivity, inflammatory markers, and gut colonization in overweight adults: A randomized, controlled trial. Mediat. Inflamm. 2014, 2014, 348959. [Google Scholar]
- Vallianou, N.; Liu, J.; Dalamaga, M. What are the key points in the association between the gut microbiome and nonalcoholic fatty liver disease? Metabol Open 2019, 1, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Jafarnejad, A.; Bathale, S.Z.; Nakhjavani, M.; Hyassan, M.Z. Effect of spermine on lipid profile and HDL functionality in the streptozotocin-induced diabetic rat model. Life Sci. 2008, 82, 301–307. [Google Scholar] [CrossRef]
- Du, X.; Edelstein, D.; Obici SHigham, N.; Zou, M.-H.; Brownlee, M. Insulin resistance reduces arterial prostacyclin synthase and eNOS activities by increasing endothelial fatty acid oxidation. J. Clin. Investig. 2006, 116, 1071–1082. [Google Scholar] [CrossRef]
- Damasceno, D.C.; Volpato, G.T.; Calderon, I.M.P.; Rudg, M.V.C.R. Oxidative stress and diabetes in pregnant rats. Anim. Reprod. Sci. 2002, 72, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Songisepp, E.; Kals, J.; Kullisaar, T.; Mändar, R.; Hütt, P.; Zilmer, M.; Mikelsaar, M. Evaluation of the functional efficacy of an antioxidative probiotic in healthy volunteers. Nutr. J. 2005, 4, 22. [Google Scholar] [CrossRef]
- Thomas, B. The global burden of diabetic kidney disease: Time trends and gender gaps. Curr. Diab. Rep. 2019, 19, 18. [Google Scholar] [CrossRef]
- Li, R.; Bilik, D.; Brown, M.B.; Zhang, P.; Ettner, S.L.; Ackermann, R.T.; Crosson, J.C.; Herman, W.H. Medical costs associated with type 2 diabetes complications and comorbidities. Am. J. Manag. Care 2013, 19, 421–430. [Google Scholar] [PubMed]
- Roobab, U.; Batool, Z.; Manzoor, M.F.; Shabbir, M.A.; Khan, M.R.; Aadil, R.M. Sources, formulations, advanced delivery and health benefits of probiotics. Curr. Opin. Food Sci. 2020, 32, 17–28. [Google Scholar] [CrossRef]
- Bahreini-Esfahani, N.; Moravejolahkami, A.R. Can synbiotic dietary pattern predict lactobacillales strains in breast milk? Breastfeed Med. 2020, 15, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Itoh, M.; Oh-Ishi, S.; Hatao, H.; Leeuwenburgh, C.; Selman, C.; Ohno, H.; Kizaki, T.; Nakamura, H.; Matsuoka, T. Effects of dietary calcium restriction and acute exercise on the antioxidant enzyme system and oxidative stress in rat diaphragm. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Yoon, G.A.; Park, S. Antioxidant action of soy isoflavones on oxidative stress and antioxidant enzyme activities in exercised rats. Nutr. Res. Pract. 2014, 8, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Dangles, O.; Dufour, C.; Manach, C.; Morand, C.; Remesy, C. Binding of flavonoids to plasma proteins. In Methods in Enzymology 335; Academic Press: Cambridge, MA, USA, 2001; pp. 319–333. [Google Scholar]
- Shimada, M.; Hasegawa, T.; Nishimura, C.; Kan, H.; Kanno, T.; Nakamura, T.; Matsubayashi, T. Anti-hypertensive effect of γ-aminobutyric acid (GABA)-rich Chlorella on high-normal blood pressure and borderline hypertension in placebo controlled double-blind study. Clin. Exp. Hypertens 2009, 31, 342–354. [Google Scholar] [CrossRef]
- Ismail, M.; Hossain, M.; Tanu, A.R.; Shekhar, H.U. Effect of spirulina intervention on oxidative stress, antioxidant status, and lipid profile in chronic obstructive pulmonary disease patients. BioMed Res. Int. 2015, 2015, 897327. [Google Scholar] [CrossRef] [PubMed]
- Treitinger, A.; Spada, C.; Verdi, J.C.; Miranda, A.F.; Oliveira, O.V.; Silveira, M.V.; Moriel, P.; Abdalla, D.S. Decreased antioxidant defence in individuals infected by the human immunodeficiency virus. Eur. J. Clin. Investig. 2000, 30, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Yuan, O.; Yu, R.; Zhang, J.; Ma, H.; Chen, S. Ameliorative Effects of Probiotic Lactobacillus paracasei NL41 on Insulin Sensitivity, Oxidative Stress, and Beta-Cell Function in a Type 2 Diabetes Mellitus Rat Model. Mol. Nutr. Food Res. 2019, 63, 1900457. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable with Units | NC | D-G1 | D-G2 | D-G3 | D-G4 | D-G5 |
|---|---|---|---|---|---|---|
| TC (mg/dL) | 91.1 ± 1.02 a | 213 ± 2.1 f | 171 ± 1.16 e | 143.64 ± 2.1 d | 95 ± 1.21 c | 115 ± 0.5 b |
| TG (mg/dL) | 71.5 ± 2.1 a | 192.33 ± 1.7 f | 145 ± 1.34 e | 117.14 ± 1.32 d | 82 ± 1.19 b | 87 ± 1.23 c |
| LDL (mg/dL) | 37.3 ± 0.9 a | 82.06 ± 1.17 f | 63.9 ± 1.8 d | 68.4 ± 2.16 e | 47 ± 1.3 b | 51 ± 1.15 c |
| VLDL (mg/dL) | 15.9 ± 0.2 a | 31 ± 0.56 e | 23.18 ± 0.58 c | 28.46 ± 1.33 d | 18.4 ± 0.24 b | 19.8 ± 0.38 b |
| HDL (mg/dL) | 28.2 ± 0.18 a | 13.19 ± 0.9 f | 21.6 ± 0.16 d | 19.3 ± 0.18 e | 26.52± 0.15 c | 24 ± 0.19 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdella, A.; Abou-Elazm, F.I.; El-Far, S.W. Pharmacological Effects of Lactobacillus casei ATCC 7469 Fermented Soybean and Green Microalgae, Chlorella vulgaris, on Diabetic Rats. Microbiol. Res. 2023, 14, 614-626. https://doi.org/10.3390/microbiolres14020043
Abdella A, Abou-Elazm FI, El-Far SW. Pharmacological Effects of Lactobacillus casei ATCC 7469 Fermented Soybean and Green Microalgae, Chlorella vulgaris, on Diabetic Rats. Microbiology Research. 2023; 14(2):614-626. https://doi.org/10.3390/microbiolres14020043
Chicago/Turabian StyleAbdella, Asmaa, Fatma I. Abou-Elazm, and Shaymaa Wagdy El-Far. 2023. "Pharmacological Effects of Lactobacillus casei ATCC 7469 Fermented Soybean and Green Microalgae, Chlorella vulgaris, on Diabetic Rats" Microbiology Research 14, no. 2: 614-626. https://doi.org/10.3390/microbiolres14020043