
Update on Mycochemical Profile and Selected Biological Activities of Genus Schizophyllum Fr. 1815



1
Department of Biology and Ecology, Faculty of Sciences, University of Novi Sad, TrgDositejaObradovića 2, 21000 Novi Sad, Serbia
2
Department of Chemistry, Biochemistry and Environmental Protection, Faculty of Sciences, University of Novi Sad, TrgDositejaObradovića 3, 21000 Novi Sad, Serbia
*
Author to whom correspondence should be addressed.
Microbiol. Res. 2023, 14(1), 409-429; https://doi.org/10.3390/microbiolres14010031
Submission received: 22 February 2023
/
Revised: 13 March 2023
/
Accepted: 16 March 2023
/
Published: 18 March 2023
(This article belongs to the Special Issue The Fungal Sense of Nonself)
Abstract
:The aim of this systematic review was to investigate new research on the antioxidant, anti-acetylcholine, antimicrobial, and antitumor activity of genus Schizophyllum, as well as to describe the mycochemical profiles. A summary was made on the published studies in the five-year period from 2017 to 2022, with the focus on the most investigated species of this genus, S. commune. Data were obtained through various scientific online databases, including Google Scholar, Semantic Scholar, PubMed, Science Direct, Elsevier, and Wiley Online Library using specific keywords. Out of 918 records published between 2017 and 2022, a total of 44 peer-reviewed studies were included in qualitative synthesis. Most examined compounds were glucans isolated from the submerged cultivation of S. commune, even though many studies reported proteins, phenolics, and some other secondary metabolites such as flavonoids, saponins, steroids, tannins, triterpenoids, etc. Schizophyllan (SPG), one of the most studied β-glucans isolated from S. commune, has been utilized in clinical trials to treat patients receiving anticancer therapy as an immunopotentiator. Considering the enormous biopotential of genus Schizophyllum, specifically S. commune and S. radiatum, additional attention should be paid to identify the biomolecules more accurately and focus on their antitumor and anti-acetylcholinesterase properties, since they proved to have great prospects in the pharmaceutical and nutraceutical industries.
Keywords:
Schizophyllum; S. commune; S. radiatum; antioxidant; antimicrobial; polysaccharides; phenolics1. Introduction
One of the most widely distributed fungal species on the Earth belongs to the genus Schizophyllum Fr 1815. It is a member of the Schizophyllacea family, order Agaricales, and phylum Basidiomycota [1]. Within the family Schizophyllaceae, order Agaricales, we distinguish two genera, Auriculariopsis and Schizophyllum.
Schizophyllum Fr., a widespread genus, has a very easily identifiable macromorphology, with characteristic lamellar hymenophores whose radial eccentric lamellae are longitudinally split at the edges [2]. Within this fungal genus, 15 different species have been described so far in the Index Fungorum database [3], including S. album Rick, S. amplum (Lév.) Nakasone, S. commune Fr., S. brasiliense W.B. Cooke, Sc. brevilamellatum Linder, S. fasciatum Pat., S. leprieurii Linder, S. murrayi Massee, S. palmatum Jungh. ex W.B. Cooke, S. umbrinum Berk., S. radiatum Fr., S. variabile Sorokin, S. lobatum Went, S. mexicanum Pat, and S. miia (Scop.) Fr. According to Mycobank, the genera Schizophyllum includes 21 species [4]. However, ex-type strains for many of the described species do not exist, and only a small number of strains recognized by the experts have been deposited into several public culture collections (i.e., S. commune, S. fasciatum, S. radiatum, and S. umbrinum) [5]. S. commune and S. radiatum were earlier considered conspecific due to morphological and ITS rDNA genetic similarity; however, multigenetic analyses confirmed that both species are similar and closely related but of independent taxa [1,6].
Taking into consideration over ten species and their great morphological variability, most of the records presented are generally considered as synonyms of S. commune, which represents the most important, well-known, and most-researched species of this genus. A study from 1961 reported over 3714 specimens of genus Schizophyllum, where over 96% were attributed to S. commune, 2% were assigned to S. fasciatum, 2% to S. umbrinum, while S. leprieurii was represented only within four collections, S. Brasiliense within two, and S. palmatum within a single collection [7]. Other species have not been researched as much, considering that they are rarely found in nature and little attention was paid to them during field research [2,6,7]. There are only scarce data related to the morphology, taxonomy, phylogeny, and distribution of S. brasiliense, S. fasciatum, S. leprieurii, S. lobatum, S. palmatum, S. umbrinum, and S. variabile [7]. The species S. fasciatum, S. umbrinum, and S. leprieurii have limited distribution and were found mostly in countries near the Caribbean Sea and Gulf of Mexico, while S. brasiliense was recorded in Brazil and S. palmatum was recorded only in Japan [7]. During the phylogenetic analysis of genus Schizophyllum from Central Argentina, S. umbrinum and S. leprieuri were described in this country as well [2]. However, Carreño-Ruiz et al. [8], after an examination of the fungal collection (the Herbarium of the Biological Sciences Academic Division of the Juarez Autonomous University of Tabasco, Mexico) and fresh basidiomes collected during field investigations, recently determined that S. radiatum was the most abundant specimen (62), followed by S. commune (28) and S. umbrinum (3). At the same time, Sammut et al. [6] conducted a morphological and genetic study of S. amplum from Estonia and Malta, which is the first public record of this species in this part of Europe.
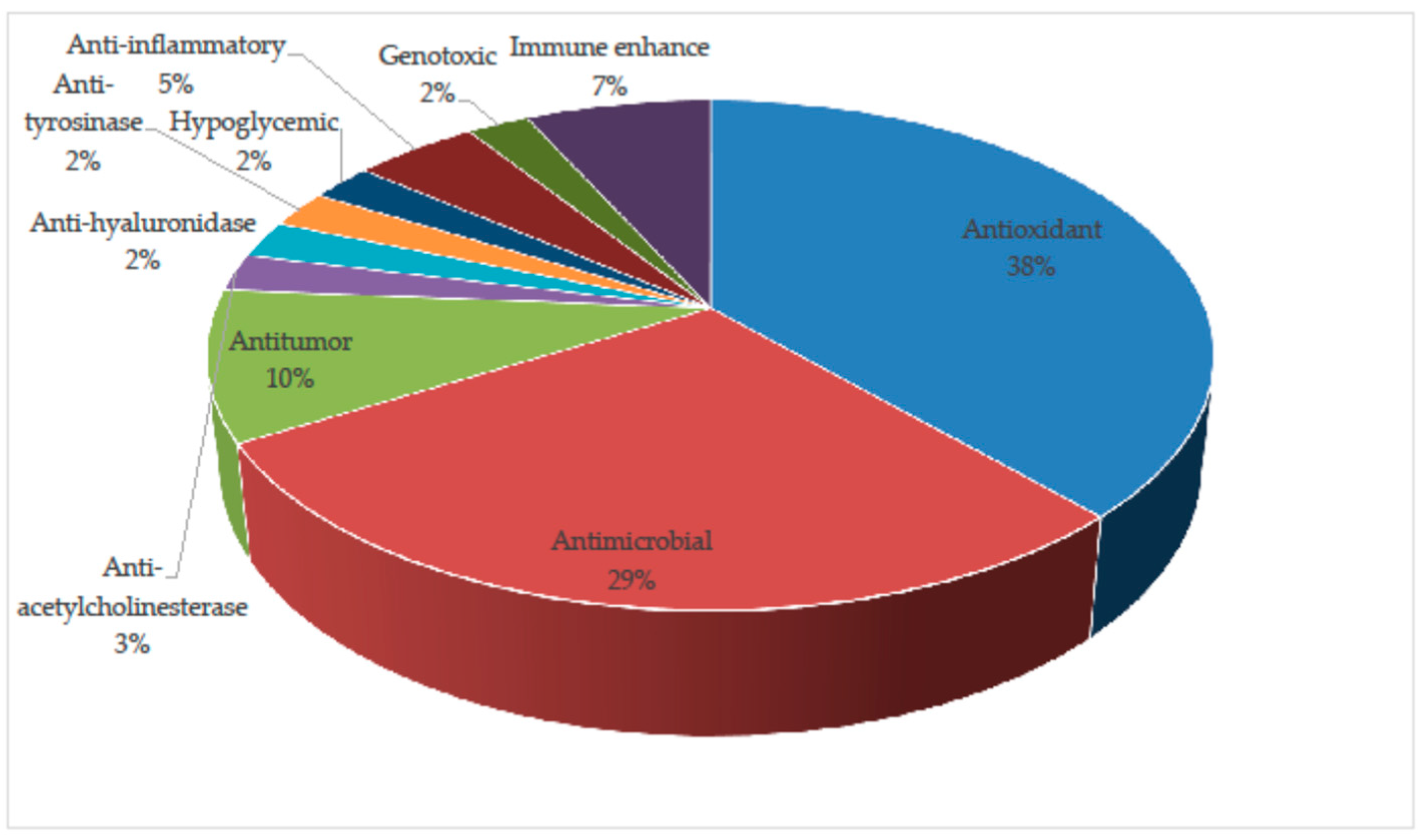
Among the research regarding the bioactivity of Schizophyllum, only one species—S. commune—has been studied in terms of its biological activities (Figure 1) and mycochemical characterization, with the exception of one publication that examined the antitumor activity of the polysaccharides (PSHs) of S. radiatum [1].
Reports about these edible fungi, until recently considered inedible due to its small size and toughness, indicate that S. commune has antidiabetic, anti-inflammatory, antimicrobial (AM), antioxidant (AO), antitumor, anti-tyrosinase, immunomodulatory, neuroprotective activity, etc. [9,10,11,12,13,14,15,16]. Various extracts and isolated substances, including PSHs, polysaccharide–protein complexes, proteoglycans, protein, phenolics (PHs), and flavonoids from fungal fermentation broth, mycelia, and fruiting bodies, have been reported to contribute to these activities [9,10,11,15,16]. In particular, PSHs seem to be effective immune-modulators and anticancer agents in addition to having other positive effects (such as wound healing, antinematode activity, and others) [17]. Moreover, schizophyllan (SPG), one of the most studied β-glucans isolated from S. commune, has been utilized in clinical trials to treat patients receiving anticancer therapy as an immunopotentiator and is currently being commercially developed by a number of Japanese pharmaceutical companies [1].
The purpose of this systematic review of the genus Schizophyllum is to systematize and describe the AO, anti-acetylcholinesterase, AM, and antitumor activity as well as thechemical characterization reported in the period from 2017 to 2022. This update is focused on S. commune, given that there is a lack of research data related to the activities and mycochemistry of other species belonging to this genus. Moreover, this review provides a baseline for future research studies.
2. Methodology
This study is a systematic review of the genus Schizophyllum, regarding the selected bioactivities and the mycochemical profiles published in the last five years (2017–2022), except 3 studies published prior to 2017, which were used as data source, given that data on the mentioned species are rare or nonexistent. Data were obtained through various scientific online databases, including Google Scholar, Semantic Scholar, PubMed, Science Direct, Elsevier, and Wiley Online Library using keywords such as Schizophyllum, Schizophyllumamplum, Schizophyllum commune, Schizophyllumfasciatum, Schizophyllumradiatum, Schizophyllumumbrinum, schizophyllan, antioxidant, etc. Only relevant studies that included the biochemistry and specifically selected bioactivities were included. A total of 918 records published between 2017 and 2022 were identified through the database search, and after duplicates and irrelevant research were removed, 256 studies were screened by title, abstract, and keywords. A total of 82 studies were assessed for eligibility by reading the full text, and afterwards, studies were removed due to detected activities not being the subject of this work. Furthermore, studies that were not peer-reviewed were also rejected, leaving a total of 44 studies which were included in the qualitative synthesis.
3. Mycochemistry
The majority ofthe research on the targeted activities of S. commune describes the effects of extracts while identifying the chemicals that are responsible for the activity. However, some studies lack this data, so it is difficult to determine which compounds are responsible for the testedactivities. Mycochemical screening of the ethanolic (EtOH) extracts of S. commune revealed the presence of saponin, tannins, alkaloids, flavonoids, terpenoids, proteins, and carbohydrates [18]. The same year, Karaman et al. [16] summarized that alkaloids, iminolactones, PHs, PSH, steroids, terpenoids, vitamins, and some other functional ingredients were found in S. commune (Figure 2 and Figure 3). Debnath et al. [9] found that the moisture content of S. commune samples originated from India was determined at 87.76% d.w., ash (3.5%), while the nutrition profile was presented through the total carbohydrate content (TCC) (42.0%), followed by crude fiber (30.0%), crude protein (15.55%), crude fat (9.0%), and lipids (0.4%). In the study from 2022, proximate analysis results confirmed that this fungus was rich in moisture (60.60%), nitrogen-free (25.15%), and had a crude protein content of 7.63% and an ash content of 6.17%, while the crude fiber and fat materials were 0.18% and 0.27%, respectively [18]. Besides S. commune, a nutritional composition of S. radiatum was conducted as well [1]. The TCC was in the range of 73–79% for water-soluble exopolysaccharides (SEPSs) and water-soluble intrapolysaccharides (SIPSs), respectively. In addition, the protein content was quantified only in SIPSs, with an amount of 1 ± 0.002% [1].
3.1. Primary Metabolites
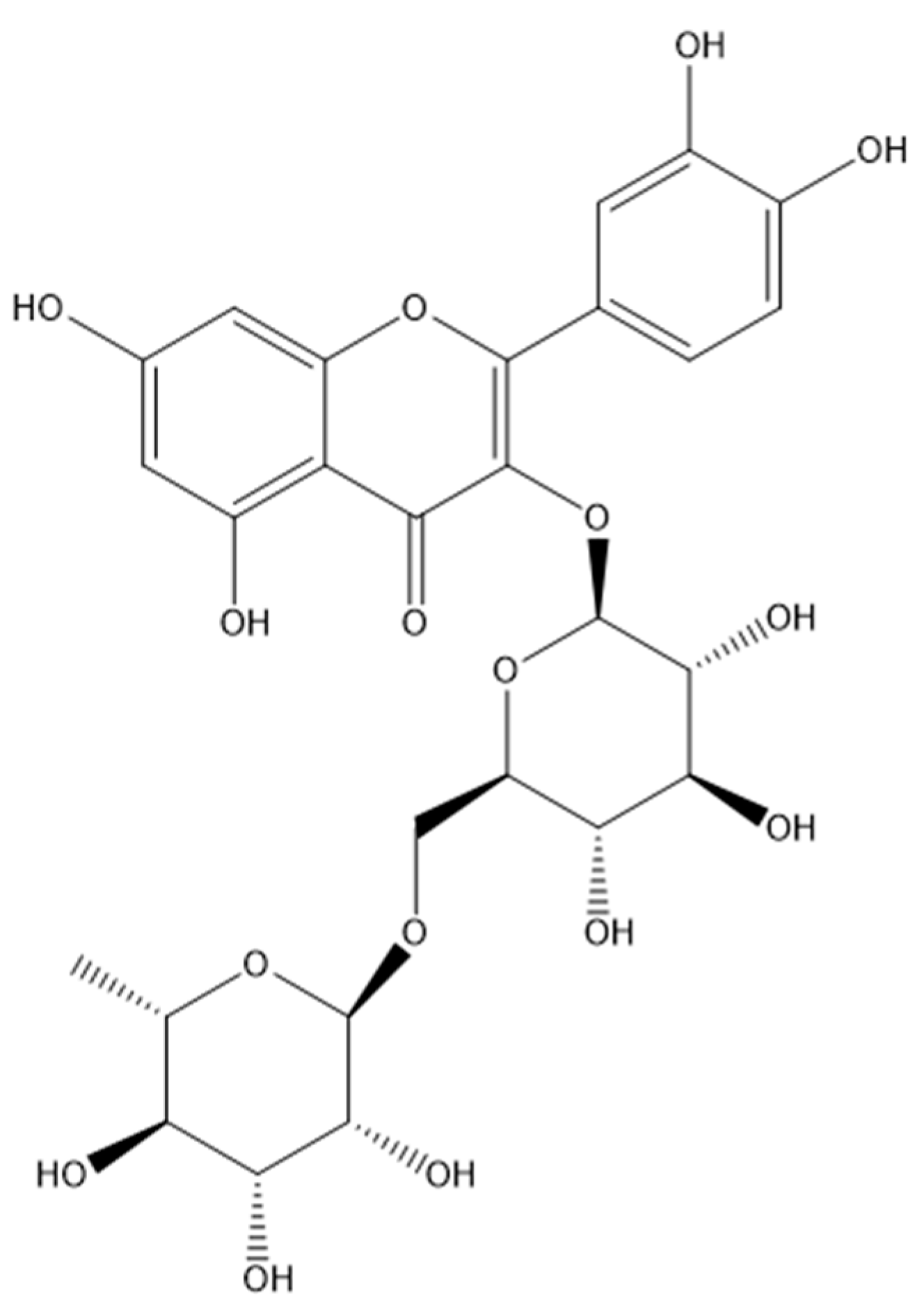
The biological activity of PSHs is closely linked to the structure, where β-glucans from S. commune play a major role [19]. SPG, β-1.3(1.6)-glucan (Figure 2), is an exopolysaccharide (EPS) isolated from S. commune and appreciated as a versatile metabolite that is useful in various industries, including the food industry and pharmaceutical industry, due to a wide range of biological effects, including antitumor, immunomodulatory, and anti-inflammatory activities [20]. However, few studies have addressed the bioactivity and molecular structure of PSHs derived from S. commune [10,11,12,13,15,21,22].
In their study, Smirnou et al. [23] worked on the procedure for obtaining highly pure SPG cleaved into shorter fragments by using ultrasonication. The ultrasonicated SPG (uSPG) was obtained by the submerged cultivation of S. commune in a bioreactor. It contained less than 0.3% protein and 0.078 IU/mg endotoxin. Although the authors found that the β-glucan concentration in the medium continued to increase, cultivation longer than 144 h was not economically acceptable [23]. In addition, the authors highlighted that the application of ultrasound-assisted downstream processing was an efficient methodology for the manufacturing of low-MW SPG, which are applicable to cosmetics and medicine. Moreover, ultrasonication, when applied on a complete culture broth with mycelium, brought certain advantages to the SPG production. It reduced the product losses downstream and made the production of highly purified PSH possible due to the efficient usage of ultrafiltration [23].
In their research, Du et al. [10,11] characterized the EPSs of S. commune. In the first study, [10] used an ultrasonic treatment to produce EPSs with various molecular weights (MWs). The authors found EPSs with different MWs, including low MW (197 kDa), medium MW (936 kDa), and high MW (1437 kDa), respectively. In addition, it was found that high- and medium-MW EPSs had triple- and single-helix conformations, while the low-MW ESP exhibited the random-coiled conformation [10]. In the same year, Du et al. [11], using DEAE-52 cellulose and Sephadex G-150 chromatography, isolated, purified, and characterized an EPS with a molecular weight of 2900 kDa from a submerged mycelial fermentation of S. commune. It was found that the EPS was a type of heteropolysaccharide, β-(1→3)-D-glucan, based on the IR spectrum. It was composed of carbohydrates (89.0%), proteins (2.2%), and uronic acid (7.52%); therefore, the EPS could be observed as a protein-bound polysaccharide based on the protein content. In addition, it was found that glucose was the main monosaccharide in the EPS, consisting of arabinose, galactose, glucose, mannose, rhamnose, ribose, and xylose [11]. Moreover, the authors found that the elemental analysis of the C, H, and N content (% w/w) was: C, 25.84%; H, 5.45%; N, 0.65%. Based on the low amounts of N, it is obvious that polysaccharide-rich samples contain only small amounts of it, and the authors suggested that N may have its origins in either chitin or proteins from the EPS [11].
Phuket et al. [13] isolated β-glucan from a S. commune strain from Thailand to determine the AO and total phenolic content (TP) (284.41± 1.22 mgGAE/g). On the other hand, Yelithao et al. [21] documented the chemical composition, monosaccharide composition, MWs of the crude PSHs, and two fractions from S. commune in relation to the immune-enhancing activity. The PSHs obtained in this study contained carbohydrates (50.3–65.8%), proteins (1.46–20.1%), and sulfates (1.33–7.01%), while gas chromatography–mass spectrometry (GC-MS) confirmed the presence of glucose (17.0–88.2%), mannose (2.70–55.2%), galactose (6.2–19.1%), and xylose (0.90–8.57%). Additionally, Yelithao et al. [21] reported MWs of crude PSHs (710.3 × 103, 170.6 × 103, and 136.3 × 103 g/mol), as well as MWs of two fractions (255.0 × 103, 44.8 × 103; 533.7 × 103, 93.5 × 103, 79.8 × 103, respectively).
Since the presumed antiviral substances in the fungal extracts include PSHs, or proteins and PHs, a correlation study was made between the anti-dengue activity of the aqueous extracts (Aq) of S. commune and these components [12]. This study showed that the main chemical composition of the Aq were glucans, with a determined content of α-glucan (1.4 ± 0.1%) and β-glucan (34.4 ± 0.4%), and protein (15 ± 0.4%), while the PHs were detected in traces (0.31 ± 0.05%). The concentration and type of detected compounds varied based on the extraction method used, but no significant correlation for anti-dengue activity was reported [12].
Nonting et al. [19] evaluated the dependence of water-extraction conditions and β-glucan content due to the high demand in cosmeceuticals application, and determined that the β-glucan content from the extract with microwave-irradiation pretreatment was increased by 27.93%. Furthermore, Chen et al. [22] determined the influence of the four different extraction methods on the physicochemical properties, AO, and hypoglycemic activity of S. commune PSHs. This research showed that the highest PSH (67.96%), protein (3.53%), and uronic acid (3.0%) content was obtained with hot-water extraction, as well as the highest MW (527.2 kDa). Another study confirmed these results, where the higher level of total PSH content (76.3 ± 1.2 mg GE/g extract) was observed in extracts subjected to hot-water extraction [24]. Meanwhile, Basso et al. [25] determined the β-glucan content of S. commune grown on different agro-industrial wastes, where substrates containing 94% grape residues showed the highest amount of PSHs (13.14%). The chemical composition of monosaccharides varied based on type of substrate used, with glucose from 8.53% to 30.03%, xylose from 4.13% to 10.95%, and arabinose from 0.24% to 0.60% [25]. Chen et al. [22] also detected ten monosaccharides, including ribose, rhamnose, glucuronic acid, galacturonic acid, glucose, galactose, xylose, arabinose, and fucose, in the PSHs extracted with hot water. However, due to the lowest AO activity obtained, hot-water extraction may not be the best choice for PSH extraction [22].
The submerged cultivation, extraction, and antitumor activity of the PSHs from S. radiatum were the focus of the study by López-Legarda et al. [1]. Elemental analysis of the dried mycelia of S. radiatum showed that the majority of the fungal biomass components were C and O (45.05% and 35.58%, respectively), followed by N and H (7.66% and 6.87%, respectively), while the presence of S was not determined. Extracts assigned as SIPS contained carbohydrates and total glucans in higher concentrations (79 ± 3% and 14 ± 2%, respectively) in comparison with SEPS (73 ± 6% and 11 ± 0.5%, respectively). Among the total detected glucans, β-glucans were the most represented in both PSH extracts (11–14%) in comparison with α-glucans (0.4–1.7%) (Table 1). Similar to some studies conducted on S. commune, the extracts have a larger concentration of β-glucans than of α-glucans, which supports the previously described link between α- and β-glucans seen in S. commune [15]. In addition, the high-performance liquid chromatography with the refraction-index-detection (HPLC-RI) technique showed that SIPS and SEPS are heteropolysaccharides composed mainly of arabinose, galactose, glucose, and xylose, similar to studies conducted on S. commune [10,22].
On the other hand, Mongkontanawat and Thumrongchote [35] worked on five pure mycelial culture strains of S. commune (Thailand), whereas 1-day-old dried S. commune fruiting bodies were used as samples. The authors applied two different β-glucan extraction methods: hot-water and hot-alkali extraction, which were compared with the control (native-MR). Results indicated that the total glucan content varied from 48.51 to 49.76%, whereas β-glucans were the most present, with a content of 47.94–49.20%, and α-glucans were present in the range of 0.41–0.57% [35]. It was discovered that the yield of β-glucan was unaffected by the extraction techniques. Moreover, FT-IR spectroscopy was used for the structural analysis of the extracted PSHs and showed that extraction conditions such as heat and alkalinity had an effect on the structure and AO activity of β-glucan from S. commune. The PSHs extracted from the fungal fruiting bodies were impacted by the extraction process, whereas the extracted β-glucan was affected by the extraction heat or pH. Additionally, it was found that the analyzed samples contained some PHs besides glucans, whereas TP was in the range of 2.54–2.64 mg GAE/g d.w. [35].
Enzymatic hydrolysiscreates peptides fromamino acids in proteins, which may have the strong ability to quench free radicals, thanks to which peptides are investigated as natural antioxidants in plants, and recently in fungal species. The relationship among radical-scavenging activity and amino acid composition, sequence, and the MW of S. commune peptides (Table 1) was determined, but no correlation was found between the degree of hydrolysis and ABTS radical-scavenging activity [14].
Mišković et al. [15] used FTIR for the determination of the spectra of PSH and EtOH extracts from Italian (IT) and Serbian (SRB) strains of S. commune. The method showed peaks characteristic for the presence of predominantly PSH molecules, small quantities of proteins, and some aromatic compounds, while the EtOH showed absorption bands characteristic for the carbohydrate polymers, polyphenolics, and proteins. Moreover, the authors analyzed the monosaccharide composition and found the presence of a large amount of D-glucose, with lower amounts of D-galactose and traces of D-mannose [15].
Аl Azad and Ai Ping [34] determined the total protein, total amino acid content (AAC), and amino acid profile of the Aq and organic extracts (50% acetone, 50% EtOH, and 50% methanol–MeOH) from two edible fungi, including S. commune. The different extraction solvents used in this study affected the yield of the examined extracts, whereas the selectivity was characterized by different strengths of the polarity. Based on the results, the Aq extracts obtained the highest dry extract yield in general, while the yield for the Aq extract of S. commune was 14.48 ± 1.2%. It was found that the amount of protein varied between the extracts of the same fungal species, whereas the Aq extract contained the highest amount of total proteins (2.17 ± 0.28%). Similar to total proteins, total amino acids were present in the highest amount in the Aq extract (443.84 mg/g) as well [34]. S. commune extracts contained a total of 17 different kinds of amino acids, with an AAC of 308.65–443.84 mg/g, and among them, glutamic acid was the most present in the acetone and EtOH extracts (150.65 ± 2.55 mg/g and 111.49 ± 1.36 mg/g, respectively). In addition, the authors revealed that these extract types contained the highest content of amino acids [34].
Debnath et al. [40] investigated seven distinct carbon and six different nitrogen sources to identify the ideal starting point for the synthesis of EPS using a submerged cultivation. It was found that S. commune produced higher levels of EPSs in the presence of xylose (4.100.26 g/L) and in the yeast-extract medium (4.26 ± 0.11 g/L), while the highest TCC was obtained in glucose (0.597 ± 0.06 g/100 g) [40]. Al-Salihi and Lau [44] determined that gamma irradiation increased the total sugar content due to the possible overexpression of genes encoding the PSH synthesis.
3.2. Secondary Metabolites
Essential mycochemicals in fungi, such asphenolics and flavonoids (Figure 3), have drawn increased attention because of their ability to prevent disease through their AO, anti-inflammatory, and AM activities. These biologically active compounds represent the main secondary metabolites found in fungi [13,16,45,46].
Deka et al. [26] analyzed the extracts of eight macrofungal species, including S. commune, and determined the presence of the following secondary metabolites: flavonoids, saponins, steroids, tannins, and triterpenoids (Table 1). Kaur et al. [27] documented the presence of flavonoids, triterpenoids, and PHs, while the ultra-high-performance liquid chromatography (UHPLC) analysis confirmed the presence gallic acid, catechin, chlorogenic acid, epicatechin, caffeic acid, coumaric acid, rutin, quercetin, and kaempferol PH compounds in the ethyl acetate extracts of S. commune.
Considering that earlier studies showed the antibacterial activity of S. commune, Woo et al. [28] isolated a new nerolidol mannoside from an AM chloroform:MeOH fraction of the submerged culture broth, designated as mannonerolidol, in addition to schizostatin and nerolidol.
On the other hand, some authors investigated the effect of different substrates or extraction methods on the TP and total flavonoid content (TF) [24,25,32]. S. commune grown in 94% cotton cake demonstrated the highest TP (299.31 ± 3.67 mg GAE/100 g substrate) compared to the other substrates from agroindustrial waste [25]. Furthermore, S. commune cultured onto wheat exhibited the highest levels of PH (8.56 ± 1.09 mg GAE/g d.w.), while the highest TP was detected on barley as a substrate (577.35 ± 29.93 μg CE/g d.w.), compared to other edible cereal media used as a substrate [32]. Abd Razak et al. [24] obtained slightly higher TPs in fruiting-body extracts subjected to hot-water extraction (14.65 ± 0.01 mg GAE/g extract) compared to cold-water extraction (14.22 ± 0.02 mg GAE/g extract).
Comparison of the TFs in the fruiting bodies of confronting fungi Hericiumcoralloides, Fomes fomentarius, and S. commune showed that cocultivation is followed by the outstanding increase of the TFs (from 7- to 20.6-fold) compared to the monoculture conditions of S. commune as a response to stress factors [31].
The TFs of the S. commune hot-water extracts were lower than their TPs, while the gamma-irradiated strains showed higher levels of flavonoids compared to the parental strain [44]. The study demonstrated that the natural strain W possessed the highest TP and TF among the different S. commune strains from Malaysia and Thailand. In another study, Debnath et al. [40] determined the TP (2.00 ± 0.05 mg GAE/g) and TF (0.04 ± 0.00 mg QE/g) in S. commune from India. The same year, Saetang et al. [41] compared the TP in SPG, supernatant from the separation of SPG extraction, and fungal essence extracted with deionized water. The results showed the highest TP in the fungal essence (14.07 ± 0.09 mg GAE/g d.w.), followed by the supernatant (12.85 ±0.12 mg GAE/g d.w.) and the SPG (6.22 ± 0.06 mg GAE/g d.w.) [41].
Boonthatui et al. [33] determined the quantitative changes in the production of selected bioactive compounds (flavonoid, β-glucan, and PH) and their AO activities in the first and second harvests of S. commune. For two subsequent growing cycles, the authors divided fresh fungi samples into three groups and harvested them separately: (1) immature fruiting body (IF); (2) mature fruiting body with immature spores (MFISs); (3) mature fruiting body with mature spores (MFMSs). The findings showed that the PH compounds, flavonoids, and β-1,3- and β-1,3-1,6-glucans were considerably higher in fully grown fungi containing spores of each lifecycle. Regardless of the cycle, the fungal-bioactive-chemical content increased as it grew [33].
Four substances, gallic, p-hydroxybenzoic, protocatechuic, and quinic acids, were measured by liquid chromatography–mass spectrometry (LC-MS/MS) in the EtOH and PSH extracts of IT and SRB S. commune strains [15]. Among the quantified compounds, p-hydroxybenzoic acid was the most present (1.87–22.19 µg/g d.w.), and its presence was confirmed in all tested samples. Among the four quantified compounds, gallic acid was the most abundant compound in the fermentation broth (F) extract of the SRB strain, with an amount of 75.77 µg/g d.w. The authors found that the EtOH extracts in general contained most of the quantified phenolic acids, whereas F contained a higher PH content than the mycelial biomass (M). Besides the LC-MS/MS profile, the authors measured the TP, although the TP levels in both strains were high and quite identical. In addition, the SRB extracts contained a higher amount (84.60 ± 1.64 mg GAE/g d.w. for F and 82.62 ± 0.99 mg GAE/g d.w. for M, respectively). Moreover, the authors showed that the F extracts of both strains contained a higher TP content compared to the M extracts [15].
In their study, Wirth et al. [36] claimed that volatile organic compounds (VOCs) such as ketones, sesquiterpenes, and ethanol are released by S. commune. Among them, sesquiterpene β-bisabolol and ethanol are involved in the S. commune biological activity against plant-pathogenic fungi that cause plant diseases [36]. Moreover, the S. commune strain thn produces large amounts of (-)-(1R,2S)-β-bisabolol, which makes it possible to analyze sesquiterpenes’ role as AM metabolites. The sesquiterpenes (-)-(1R,2S)-β-bisabolol, β-bisabolene, and (E)-γ-bisabolene were dominant in GC-MS of different S. commune strains [36]. Volatile sulfur compounds were identified as the major VOCs produced by S. commune, and they can be helpful as biomarkers to diagnose different states, such as allergic bronchopulmonary mycosis caused by S. commune [37]. Among the identified compounds, dimethyl disulfide (DMDS), dimethyl trisulfide (DMTriS), dimethyl tetrasulfide (DMTetraS), methyl ethyl disulfide (MEDS), diethyl disulfide (DEDS), and H2S were determined as the major VOCs of S. commune [37].
The constant search for new antibiotics within the secondary metabolites of fungi led to the discovery of fusidane-type antibiotics. These compounds are represented by fusidic acid, helvolic acid, and cephalosporin P1, and share similar core structures, but are produced by fungi [47]. Chen et al. [22] isolated sixteen active fusidane-type compounds from the ethyl acetate extract of the solid-state fermented S. commune.
When compared to current medicines for many disorders, metallic nanoparticle systems have a significant potential to offer effective treatment with few adverse effects [48]. In recent years, silver has grown in prominence as a crucial component for producing metallic nanoparticles, whereas silver nanoparticles (AgNPs) have undergone extensive assessment for the creation of future diagnostic and treatment systems [42]. In relation to this, Tosun et al. [42] synthesized AgNPs with green chemistry by microwave irradiation using the edible fungal species Geoporasumneriana and S. commune.
4. Biological Activities
4.1. Antioxidant Activity
Edible fungi represent a fantastic source of antioxidants due to the accumulation of different secondary metabolites, including PHs, flavonoids, steroids, terpenoids, carotenoids, polyketides, and quinones [46].
Debnath et al. [9] analyzed the AO activity of the MeOH extract of S. commune using a DPPH assay and showed the scavenging effect rapidly increased from 0.5 mg/mL to 8.0 mg/mL, showing an IC50 value at 1.4 mg/mL. The highest scavenging ability was determined at a concentration of 8.0 mg/mL (91.85%) [9], while a 2022 study of S. commune from a submerged culture determined the highest activity (92.59%) at a 16 mg/mL concentration, with a chelating effect of 91.11% [40]. In comparison with standard compounds (ascorbic acid and BHA), the extract showed a lower scavenging ability at a concentration of 0.125 mg/mL (inhibition of 95.37% and 97.83%, respectively) [9,40]. Moreover, the scavenging activity of β-glucan isolated from the S. commune strain from Thailand was confirmed by ABTS (IC50 = 0.829 ± 0.006 mg/mL) and DPPH (IC50 = 0.724 ± 0.021 mg/mL) assays [13]. Since, in this study, the TP in β-glucan extracts were documented [13], we can assume that the complexes of primary and secondary metabolites, in this case PSHs and PHs, are responsible for the obtained activity, which coincides with previously published research [15]. This is also supported by studies where peptides and PHs found in S. commune from India and Thailand positively correlated with ABTS scavenging activity [14,32].
Chen et al. [22] obtained a concentration-dependent trend of the reducing power, DPPH, and hydroxyl radical-scavenging abilities (range from 0.5 to 3.0 mg/mL) of the PSHs extracted using different extraction methods. Abd Razak et al. [24] demonstrated the AO activity of hot- and cold-water extracts by DPPH, SOA, and FRAP assays. Results of this study showed that the cold-water extract of S. commune had the highest DPPH (69.79 ± 0.13%) and SOA inhibition activity (94.82 ± 1.29%) [24]. Since the ultrasound-assisted hot-water extraction exhibited the highest PSH and TP, as well as cold extraction, they are proposed as the best extraction techniques due to the highest levels of AO activity [22,24]. The highest DPPH scavenging activity was recorded for S. commune grown on 94% cotton cake (60.74 ± 0.7%) and wheat substrate (IC50 = 3.83 ± 0.33 mg/mL), while the strongest FRAP activity was obtained from barley (IC50 = 2.14 mg/mL) [25,32].
Additionally, the introduction of the stress of direct fungi confrontation led to an increased AO activity, as well as the levels of MDA detected by lipid peroxidation in the fruiting bodies of S. commune [31]. Boonthatui et al. [33] studied the AO potential of S. commune fruiting bodies at different stages of maturity in both generations and confirmed the scavenging activity of different maturity samples with the DPPH, АBTS, and ferric-reducing AO power assays. The highest DPPH, ABTS, and ferric-reducing AO power in the first cycle of harvesting was determined as IC50 = 0.69 ± 0.02 mg/mL, IC50 = 2.59 ± 0.13 mg/mL, and EC50 = 1.86 ± 0.17 mg/mL, respectively, while for the second cycle, it was measured as IC50 = 0.67 ± 0.08, IC50 = 2.18 ± 0.36, and EC50 = 1.15 ± 0.07 mg/mL, respectively.
Moreover, it was observed that fully grown cultivated fungi strongly correlated with their TP and TF, whereas bioactive compounds and AO activities of the second-harvest fungi were higher than for the first batch [33]. In the same year, Mišković et al. [15] worked on the examination of AO activity using six different in vitro assays (DPPH, ABTS, FRAP, OH, NO, and SOA). Among the analyzed extracts, the PSH extract after 7 days of submerged cultivation exhibited the most significant AO activity, opposite to the EtOH extracts, where the 14- and 21-day incubation periods stood out [15]. Moreover, it was found that there are statistically significant differences between the PSH and EtOH samples from the SRB strain in the DPPH and FRAP assays. The authors concluded that the majority of the submerged samples demonstrated significant AO activity, which is directly related to the traits of the strain’s growth because it can indicate the point of entry into the secondary metabolism [15].
Mongkontanawat and Thumrongchote [35] measured the AO potential of β-glucan extracts using DPPH and ABTS assays, and found small differences between the activities of the five analyzed S. commune strains (Table 1).
One year later, the AO activity of the SPG, the supernatant from the separation of the extraction, and the fungal essence was examined [41]. Results revealed that the split-gill fungal essence exhibited prominent potential for the inhibition of the ABTS radical (98.18%) at 5.00 mg/mL (IC50 = 0.73 ± 0.02 mg of dry extract/mL) [41]. On the other hand, Al Azad and Ping [38] compared the AO activity of this fungi extracted in Aq and distinct organic solvents (acetone, MeOH, and EtOH). The highest DPPH scavenging activity was seen in the Aq extract (IC50 = 1.52 mg/mL), while the greatest reducing power was exhibited by the acetone extract (IC50 = 0.22 mg/mL) [38]. Furthermore, the results measured by the ABTS and DPPH assays in the Al-Salihi and Lau study [44] indicate the higher scavenging activities of hot-water extract of S. commune compared to the Aq extract of P. pulmonarius. The study suggested that PFs and flavonoids are the main contributors to higher activity since a positive correlation was observed. They also presented that gamma-irradiated strains exhibited a superior scavenging activity and reducing power in all assays other than the natural strain. The authors explained that mutation triggered by irradiation might have caused changes in the gene expression and led to the increased production of secondary metabolites with AO activity [44].
4.2. Anti-Acetylcholinesterase Activity
Free-radical production and quenching imbalance may result in a variety of illnesses, including neurodegenerative conditions such as Alzheimer’s disease [15]. Acetylcholinesterase inhibitors have gained popularity among researchers because there is no cure for this condition or a way to stop or reverse its progression. As a result, a variety of organisms, including fungi, have been studied in an effort to develop a physiologically active substance that could replace the acetylcholinesterase inhibitors effectively. To the best of our knowledge, the anti-acetylcholinesterase activity of S. commune was only investigated in one study from 2021 [15]. Mišković et al. [15] demonstrated that the PSH extracts showed remarkable neuroprotective results, launching this type of research, and establishing S. commune as an important source of AChE inhibitors. In their study, the greatest AChE activity was seen in the M28 SRB PSH extract (IC90 = 79.73 ± 26.34 μg/mL), while the IT was more active in the case for the F14 EtOH extracts (IC50 = 0.8 ± 0.6 μg/mL). It is important to note that the activity of the M28 SRB PSH extract is comparable with the activity of a commercially approved Alzheimer’s disease drug donepezil (IC90 = 87.92 μg/mL). Anti-AChE activity was documented for phenolic compounds and flavonoids [49], while results from our previous study strongly pointed out the possibility of the synergistic effect of both primary (mixture of polysaccharide fraction, probably SPG) and secondary metabolites (phenolics) [15].
4.3. Antimicrobial Activity
Medicinal fungi have great prospects in the drug and nutraceutical industries, as they possess a wide range of pharmacological properties, including antiviral (AV), antifungal (AF), and antibacterial (AB) activity [16]. Finding AB metabolites from various sources, including medicinal fungi, is crucial in the search for novel antibiotics that can fight the antibiotic resistance of a number of bacterial infections.
Debnath et al. [9] analyzed the AM activity of the MeOH extract of S. commune by using by the disc-diffusion method on Gram-positive bacterial strains (Bacillus subtilis and Staphylococcus aureus) and Gram-negative bacterial strains such as Escherichia coli, Pseudomonas aeruginosa, and Xanthomonas campestris. Based on the results, the MeOH extract showed the highest inhibition against B. subtilis and X. campestris in comparison with those of E. coli and S. aureus. S. commune’s MeOH extract had no effect on P. aeruginosa, which was confirmed in a 2022 study by Debnath et al. [40] as well. Meanwhile, the study by Deka et al. [26] analyzed the AM activity of the EtOH and Aq extracts of macrofungal species, including S. commune, against B. subtilis, E. coli, P. aeruginosa, Salmonella typhi, and S. aureus. The analyzed extracts showed various degrees of AM effects against the tested microorganisms, whereas the EtOH extract showed satisfactory results, with an MIC range of 10–25 mg/mL. In addition, the S. commune extracts showed wider inhibition zones in the disc-diffusion method. The activities of commercial drugs, when compared to the analyzed extracts, were found to be lower, whereas the microbial-inhibition activity was found to be in the range of 2.28–7.33 mm. The highest activity was shown against E. coli [26].
AB activity against S. aureus was confirmed in another study, together with AF activity against Candida albicans, as reported by Mandal [50]. The significant AF activity of schizostatin against Rhizoctonia solani, Diaporthe sp., Botrytis cinerea, and Alternaria solani was reported, while a new nerolidol mannoside (mannonerolidol) isolated from the same culture broth of S. commune displayed moderate AF activities against R. solani and Diaporthe sp., with clear zone diameters of 11.4 and 10.9 mm, respectively [28]. In the same study, schizostatin exhibited AB activity against B. subtilis and S. aureus, while nerolidol did not exhibit any AM activity.
Khardziani et al. [30] conducted AB screening of S. commune grown on agar plates in the submerged and solid-state fermentation of five lignocellulosic growth substrates. Strong activity against S. aureus (13 mm inhibition zone), S. enteritidis (11 mm), E. coli (12 mm), P. aeruginosa (19 mm), and S. epidermitidis (12 mm) was observed [30]. Moreover, S. commune filtrates inhibited the growth of B. cereus and E. cloacae [29]. The testing of the AB activity after submerged and solid-state cultivation revealed that the highest inhibition activity against both Gram-negative and Gram-positive strains (94 and 98% growth inhibition, respectively) was in the presence of mandarin pulp in the substrate [30]. This study also documented the effect of the carbon and nitrogen source on the AB activity, where it was shown that xylose ensured the maximum inhibition for S. aureus and glucose for E. coli, as well as that KNO3 supplementation enhancing the AB activity [30].
In their study, Mišković et al. [15] examined AM activity, measuring the minimal bactericidal concentration (MBC) and MIC of EtOH extracts. Only two extracts among all the tested SRB-strain samples exhibited AB activity, while the IT strain was not active. It was found that the F extract of the SRB strain after 21 days of cultivation expressed the highest AM activity, with MIC and MBC values lower than 0.31% against three bacterial strains (B. cereus, E. coli, and S. aureus), while the lowest was exhibited against P. aeruginosa (with MIC and MBC values of 5%). At the same time, Wirth et al. [36] determined that the presence of the bisabolene mixture from S. communethn mutants was considered potentially responsible for the growth inhibition of the analyzed bacteria and fungi based on the fact that their absence in S. commune wildtype monokaryon correlated with reduced effects. It was found that used mixture led to growth inhibition in Flammulinavelutipes, Ganoderma lucidum, and Kuehneromycesmutabilis, whereas the influence of (-)-(1R,2S)-β-bisabolol was even stronger on growth reduction for all analyzed mushrooms. The (-)-(1R,2S)-β-bisabolol and the mixture of bisabolenes did not show an effect on bacterial growth that would mimic the effect of the VOC of S. commune thn. In addition, the bisabolene mixture and (-)-(1R,2S)-β-bisabolol significantly reduced the swarming motility in the experiments on the analyzed Gram-negative bacteria Serratia marcescens.
Al Azad and Ping [38] compared the AB activity of S. commune extracted with Aq and three types of organic solvents. All tested extracts inhibited Vibrio harveyi growth, with an MIC lower than 1.25 mg/mL, while the MIC values of 2.5 mg/mL and 5 mg/mL in V. parahaemolyticus, as well as of 5 mg/mL and 10 mg/mL in V. anguilarium, were observed [38].
Fusidane-type active compounds, helvolinic acid and diketo-cephalosporin P1, isolated from S. commune ethyl acetate extracts, exhibited strong AB activity against Stenotrophomonas maltophilia, with MIC values of 4 µg/mL and 16 µg/mL, respectively [39].
In summary, S. commune exhibited a significant AB response; however, a stronger effect was observed against Gram-positive bacteria as opposed to Gram-negative ones [9,30,40,50]. Moreover, S. aureus and B. subtilis were more sensitive to MeOH extracts [40], which indicates that further research should be based on this type of solvent.
The prominent anti-dengue serotype 2 activity of the Aq soluble extracts of S. commune was documented in vitro with simultaneous (IC50 = 424.9 µg/mL) and penetration (IC50 = 279.3 µg/mL) assays [12]. Zhang et al. [43] summarized that hemolysin monomer and ribonuclease extracted from S. commune fruiting bodies inhibit the activity of HIV-1 reverse transcriptase, while SPG was associated with regulation of humoral immune response and improving antibody levels in patients with chronic hepatitis B. Furthermore, this polysaccharide (SPG) enhanced shrimpimmune levels against white spot syndrome virus [43].
In addition to the most common application of fungal extracts, AgNPs have a big potential in designing next-generation anticancer and AM agents. Tosun et al. [42] analyzed synthesized AgNPs nanoparticles as potential AM and anticancer agents. The AM potential of the AgNPs was determined as AB against pathogenic bacterial (Enterococcus faecalis, Klebsiella pneumonia, Pseudomonas aeruginosa, and Staphylococcus aureus) and fungal (Candida albicans, and C. utilis) strains, while the anticancer activities were studied on the four cancer cell lines (breast (MCF-7), lung (A549), colon (HT-29), and liver (HUH-7) cancer cells). The authors pointed out the fact that nanoparticles synthesized from fungi have great potential for the treatment of cancer and infectious diseases [42].
4.4. Antitumor Activity
Due to the diversity of their physicochemical properties and the intricacy of their antitumor effects, natural PSHs’ anticancer effectiveness has been challenging to compare with manufactured medicines [51]. Different fungal PSHs showed varying degrees of antitumor activity, and typically, were in correlation with the immunomodulatory activities of the identified compounds [51]. The antitumor activity of S. commune was analyzed in studies conducted before 2017, while in the period covered in this paper, we did not find studies that examined the mentioned activity of this species.
The submerged cultivation, extraction, and antitumor activity of PSHs from S. radiatum were the focus of the study by Lopez-Legarda et al. [1]. Different cancer cell lines (EL-4, MDA-MB-231, RAW 264.7, and U937) were analyzed for antitumor and immunostimulant activities. The findings imply that these PSHs both directly and indirectly inhibit tumor growth by stimulating immune cells such asmacrophages [1]. The highest antitumor activity was determined for the SIPS extract, while the best immunostimulant activity was obtained for the SEPS extract. The researchers concluded that, although the S. commune PSHs studied did not have a strong and direct antitumor effect as with other anticancer drugs, they had a considerable impact on the in vitro activation of macrophages. Probably, the in vivo the antitumor activity of SIPS and SEPS could be enhanced with the help of all the immunological machinery and gut microbiota [1].
5. Conclusions
In conclusion, research on the described bioactivites and mycochemistry identification has expanded in a five-year period, particularly in terms of the S. commune species, except for the antitumor activity. The most examined bioactivites were AO and AM, while the anti-AChE and antitumor activities received only passing consideration. Moreover, the most examined compounds were glucans, isolated from S. commune, even though many studies reported PHs and other secondary metabolites, including alkaloids, terpenoids, and steroids, detected as well. These findings suggest that the synergism of primary and secondary metabolites is responsible for the detected bioactivities of genus Schizophyllum. In general, more attention should be paid to the antitumor and anti-AChE activity, together with more accurate compound identification of genus Schizophyllum, specifically S. commune and S. radiatum, considering their huge biopotential. Nevertheless, S. commune should be considered as one of the first choices for functional food and pharmaceutical production.
Author Contributions
Conceptualization, J.M. and M.R.; methodology, J.M., M.R. and N.K.; validation, J.M., M.R., N.K. and M.K.; formal analysis, J.M. and M.R.; investigation, J.M., M.R. and N.K.; resources, J.M., M.R., N.K. and M.K.; data curation, J.M. and M.R.; writing—original draft preparation, J.M., M.R., N.K. and M.K.; writing—review and editing, M.K.; visualization, J.M. and M.R.; supervision, J.M., M.R. and M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AAC | total amino acid content |
| AB | antibacterial |
| ABTS | and 2,2-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid |
| AChE | acetylcholinesterase |
| AF | antifungal |
| AM | antimicrobial |
| AO | antioxidant |
| Aq | aqueous, water |
| AV | antiviral |
| BHA | butylated hydroxyanisole |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| EPS | exopolysaccharide |
| EtOH | ethanolic |
| F | fermentation broth |
| FRAP | ferric reducing–antioxidant power |
| GC-MS | gas chromatography–mass spectrometry |
| HPLC-RI | high-performance liquid chromatography with refraction index detection |
| IR | infrared |
| IT | Italy |
| LC-MS/MS | liquid chromatography–mass spectrometry |
| M | mycelial biomass |
| MBC | minimum bactericidal concentration |
| MeOH | methanol |
| MIC | minimum inhibitory concentration |
| MW | molecular weight |
| NO | nitric oxide radical |
| OH | hydroxyl radical |
| PH | phenolic |
| PSH | polysaccharide |
| SEPS | water-soluble exopolysaccharide |
| SIPS | water-soluble intrapolysaccharide |
| SOA | superoxide anion radical |
| SPG | schizophyllan |
| SRB | Serbia |
| TCC | total carbohydrate content |
| TF | total flavonoid content |
| TP | total phenolic content |
| VOC | volatile organic compounds |
References
- López-Legarda, X.; Rostro-Alanís, M.D.; Parra-Saldívar, R.; Villa-Pulgarín, J.A.; Segura-Sánchez, F. Submerged cultivation, characterization and in vitro antitumor activity of polysaccharides from Schizophyllumradiatum. Int. J. Biol. Macromol. 2021, 186, 919–932. [Google Scholar] [CrossRef] [PubMed]
- Robledo, G.L.; Popoff, O.F.; Amarilla, L.D.; Moncalvo, J.M.; Urcelay, C. Schizophyllumleprieurii and the Schizophyllumumbrinum lineage (Agaricales, Basidiomycota) in Argentina. Lilloa 2014, 51, 87–96. [Google Scholar]
- Available online: http://www.speciesfungorum.org/Names/Names.asp?strGenus=Schizophyllum (accessed on 30 January 2023).
- Available online: https://www.mycobank.org/ (accessed on 4 February 2023).
- Siqueira, J.P.; Sutton, D.A.; Gené, J.; García, D.; Guevara-Suarez, M.; Decock, C.A.; Wiederhold, N.P.; Guarro, J. Schizophyllumradiatum, an emerging fungus from human respiratory tract. J. Clin. Microbiol. 2016, 54, 2491–2497. [Google Scholar] [CrossRef] [Green Version]
- Sammut, C.; Alvarado, P.; Saar, I. Schizophyllumamplum (Agaricales, Schizophyllaceae): A rare Basidiomycete from Malta and Estonia. Ital. J. Mycol. 2019, 48, 50–56. [Google Scholar]
- Cooke, W.B. The genus Schizophyllum. Mycologia 1961, 53, 575–599. [Google Scholar] [CrossRef]
- Carreño-Ruiz, S.D.; Lázaro, A.A.; García, S.C.; Hernández, R.G.; Chen, J.; Navarro, G.K.; Fajardo, L.V.; Pérez, N.D.; Cruz, M.T.; Blanco, J.C.; et al. New record of Schizophyllum (Schizophyllaceae) from Mexico and the confirmation of its edibility in the humid tropics. Phytotaxa 2019, 413, 137–148. [Google Scholar] [CrossRef]
- Debnath, S.; Saha, A.K.; Panna, D. Biological activities of Schizophyllum commune Fr.: A wild edible mushroom of Tripura, North East India. J. Mycopathol. Res. 2017, 54, 469–475. [Google Scholar]
- Du, B.; Yang, Y.; Bian, Z.; Xu, B. Molecular weight and helix conformation determine intestinal anti-inflammatory effects of exopolysaccharide from Schizophyllum commune. Carbohydr. Polym. 2017, 172, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Yang, Y.D.; Bian, Z.X.; Xu, B.J. Characterization and anti-inflammatory potential of an exopolysaccharide from submerged mycelial culture of Schizophyllum commune. Front. Pharmacol. 2017, 8, 252. [Google Scholar] [CrossRef] [PubMed]
- Ellan, K.; Thayan, R.; Raman, J.; Hidari, K.I.; Ismail, N.; Sabaratnam, V. Anti-viral activity of culinary and medicinal mushroom extracts against dengue virus serotype 2: An in-vitro study. BMC Complement. Altern. Med. 2019, 19, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Phuket, S.R.N.; Sangkaew, T.; Chanapan, P.; Techaoei, S. Biological activity of SS-glucans from edible mushroom, Schizophyllum commune in Thailand. Int. J. Appl. Pharm. 2019, 11, 110–112. [Google Scholar] [CrossRef]
- Wongaem, A.; Reamtong, O.; Srimongkol, P.; Sangtanoo, P.; Saisavoey, T.; Karnchanatat, A. Antioxidant properties of peptides obtained from the split gill mushroom (Schizophyllum commune). J. Food. Sci. Technol. 2021, 58, 680–691. [Google Scholar] [CrossRef]
- Mišković, J.; Karaman, M.; Rašeta, M.; Krsmanović, N.; Berežni, S.; Jakovljević, D.; Piattoni, F.; Zambonelli, A.; Gargano, M.L.; Venturella, G. Comparison of two Schizophyllum commune strains in production of acetylcholinesterase inhibitors and antioxidants from submerged cultivation. J. Fungi 2021, 7, 115. [Google Scholar] [CrossRef]
- Karaman, M.; Čapelja, E.; Rašeta, M.; Rakić, M. Diversity, Chemistry, and Environmental Contamination of Wild Growing Medicinal Mushroom Species as Sources of Biologically Active Substances (Antioxidants, Anti-Diabetics, and AChE Inhibitors). In Biology, Cultivation and Applications of Mushrooms; Arya, A., Rusevska, K., Eds.; Springer: Singapore, 2022; Volume 8, pp. 203–257. [Google Scholar]
- Elkhateeb, W.A.; El Ghwas, D.E.; Gundoju, N.R.; Somasekhar, T.; Akram, M.; Daba, G.M. Chicken of the woods Laetiporussulphureus and Schizophyllum commune treasure of medicinal mushrooms. J. Microbiol. Biotechnol. 2021, 6, 000201. [Google Scholar]
- Acanto, R.B.; Van Helen, S.C.; Gimoto, P.H. Phytochemical screening, cytotoxic activity, and proximate analysis of split gill mushroom (Schizophyllum commune). J. Multidiscip. Res. 2022, 47, 15–29. [Google Scholar] [CrossRef]
- Nonting, S.; Satirapipathkul, C.; Charnvanich, D. The influences of extraction conditions on the content of β-glucan extracted from Schizophyllum commune processed-product residue. Int. J. Biosci. Biochem. Bioinform. 2020, 11, 8–13. [Google Scholar]
- Kumar, A.; Bharti, A.K.; Bezie, Y. Schizophyllum commune: A fungal cell-factory for production of valuable metabolites and enzymes. BioResources 2022, 17, 5420–5436. [Google Scholar] [CrossRef]
- Yelithao, K.; Surayot, U.; Lee, C.; Palanisamy, S.; Prabhu, N.M.; Lee, J.; You, S. Studies on structural properties and immune-enhancing activities of glycomannans from Schizophyllum commune. Carbohydr. Polym. 2019, 218, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yin, C.; Fan, X.; Ma, K.; Yao, F.; Zhou, R.; Gao, H. Characterization of physicochemical and biological properties of Schizophyllum commune polysaccharide extracted with different methods. Int. J. Biol. Macromol. 2020, 156, 1425–1434. [Google Scholar] [CrossRef]
- Smirnou, D.; Knotek, P.; Nešporová, K.; Šmejkalová, D.; Pavlík, V.; Franke, L.; Velebný, V. Ultrasound-assisted production of highly-purified β-glucan schizophyllan and characterization of its immune properties. Process Biochem. 2017, 58, 313–319. [Google Scholar] [CrossRef]
- Abd Razak, D.L.; Jamaluddin, A.; Abd Rashid, N.Y.; Sani, N.A.; Abdul Manan, M. Assessment of cosmeceutical potentials of selected mushroom fruit body extracts through evaluation of antioxidant, anti-hyaluronidase and anti-tyrosinase activity. J 2020, 3, 329–342. [Google Scholar]
- Basso, V.; Schiavenin, C.; Mendonça, S.; de Siqueira, F.G.; Salvador, M.; Camassola, M. Chemical features and antioxidant profile by Schizophyllum commune produced on different agroindustrial wastes and byproducts of biodiesel production. Food Chem. 2020, 329, 127089. [Google Scholar] [CrossRef]
- Deka, A.C.; Sarma, I.D.; Dey, S.; Sarma, T.C. Antimicrobial properties and phytochemical screening of some wild macrofungi of Rani-Garbhanga reserve forest area of Assam, India. Adv. Appl. Sci. Res. 2017, 8, 17–22. [Google Scholar]
- Kaur, M.; Chadha, P.; Kaur, S.; Kaur, A.; Kaur, R.; Yadav, A.K.; Kaur, R. Schizophyllum commune induced genotoxic and cytotoxic effects in Spodoptera litura. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Woo, E.E.; Kim, J.Y.; Kim, J.S.; Kwon, S.W.; Lee, I.K.; Yun, B.S. Mannonerolidol, a new nerolidol mannoside from culture broth of Schizophyllum commune. J. Antibiot. 2019, 72, 178–180. [Google Scholar] [CrossRef]
- Hastuty, A.; Mairani, R.; Rosada, K.K. Antibacterial and anti-biofilm activities of culture filtrates from Schizophyllum commune, Coniothyrium sp., and Fusarium sp. Makara J. Sci. 2020, 24, 4. [Google Scholar]
- Khardziani, T.; Metreveli, E.; Didebulidze, K.; Elisashvili, V.I. Screening of Georgian medicinal mushrooms for their antibacterial activity and optimization of cultivation conditions for the split gill medicinal mushroom, Schizophyllum commune BCC64 (Agaricomycetes). Int. J. Med. Mushrooms 2020, 22, 659–669. [Google Scholar] [CrossRef]
- Pasailiuk, M. Total flavonoid content, lipid peroxidation and total antioxidant activity of Hericiumcoralloides, Fomes fomentarius and Schizophyllum commune cultivated by the method of direct confrontation. IJM 2020, 49, 25–37. [Google Scholar]
- Boonthatui, Y.; Chongsuwat, R.; Kittisakulnam, S. Production of antioxidant bioactive compounds during mycelium growth of Schizophyllum commune on different cereal media. CMUJ. Nat. Sci. 2021, 20, e2021032. [Google Scholar] [CrossRef]
- Boonthatui, Y.; Thakhiew, W.; Kittisakulnam, S. Quantitative changes of phenolics, flavonoids, β-glucans and antioxidant activities during the first and second harvests of split-gill mushroom (Schizophyllum commune). Agr. Nat. Resour. 2021, 5, 995–1004. [Google Scholar]
- Al Azad, S.; Ai Ping, V.C. Comparison of protein and amino acids in the extracts of two edible mushroom, Pleurotussajor-caju and Schizophyllum commune. Adv. Biosci. Biotechnol. 2021, 12, 286–296. [Google Scholar] [CrossRef]
- Mongkontanawat, N.; Thumrongchote, D. Effect of strains and extraction methods on β-glucan production, antioxidant properties, and FTIR spectra from mushroom fruiting bodies of Schizophyllum commune Fr. in Thailand. Food Res. 2021, 5, 410–415. [Google Scholar] [CrossRef]
- Wirth, S.; Krause, K.; Kunert, M.; Broska, S.; Paetz, C.; Boland, W.; Kothe, E. Function of sesquiterpenes from Schizophyllum commune in interspecific interactions. PLoS ONE 2021, 16, e0245623. [Google Scholar] [CrossRef] [PubMed]
- Toyotome, T.; Takino, M.; Takaya, M.; Yahiro, M.; Kamei, K. Identification of volatile sulfur compounds produced by Schizophyllum commune. J. Fungi 2021, 7, 465. [Google Scholar] [CrossRef] [PubMed]
- Al Azad, S.; Ping, V.C.A. Antioxidant properties and antimicrobial activity in the extracts of two edible mushroom, Pleurotussajorcaju and Schizophyllum commune. Adv. Biosci. Biotechnol. 2022, 13, 352–361. [Google Scholar] [CrossRef]
- Chen, G.G.; Zhu, Q.F.; Long, X.M.; Lu, Q.; Li, K.Y.; Chen, Q.; Xu, G.B. Antibacterial activities of the chemical constituents of Schizophyllum commune MST7-3 collected from the coal area. Nat. Prod. Res. 2022, 36, 4645–4654. [Google Scholar] [CrossRef]
- Debnath, S.; Hore, S.; Das, P.; Saha, A.K. The evaluation of selected bioactivities of fungal mycelium of a wild Schizophyllum commune Fr. from submerged cultures. J. Microbiol. Biotechnol. Food Sci. 2022, 12, e3250. [Google Scholar] [CrossRef]
- Saetang, N.; Amornlerdpison, D.; Rattanapot, T.; Ramaraj, R.; Unpaprom, Y. Processing of split gill mushroom as a biogenic material for functional food purpose. Biocatal. Agric. Biotechnol. 2022, 41, 102314. [Google Scholar] [CrossRef]
- Tosun, N.G.; Kaplan, Ö.; Türkekul, İ.; Gökçe, İ.; Özgür, A. Green synthesis of silver nanoparticles using Schizophyllum commune and Geoporasumneriana extracts and evaluation of their anticancer and antimicrobial activities. Part. Sci. Technol. 2022, 40, 801–811. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, G.; Ling, J. Medicinal fungi with antiviral effect. Molecules 2022, 27, 4457. [Google Scholar] [CrossRef] [PubMed]
- Al-Salihi, A.S.R.; Lau, B.F. Antioxidant activities and chemical composition of hot water-soluble extracts and fractions of selected Schizophyllum commune Fr. strains. Int. J. Health Sci. 2022, 6, 3976–3994. [Google Scholar] [CrossRef]
- Rašeta, M.; Popović, M.; Beara, I.N.; Šibul, F.; Zengin, G.; Krstić, S.; Karaman, M. Anti-inflammatory, antioxidant and enzyme inhibition activities in correlation with mycochemical profile of selected indigenous Ganoderma spp. from Balkan region (Serbia). Chem. Biodivers. 2020, 17, e2000828. [Google Scholar] [CrossRef]
- Rašeta, M.J.; Rakić, M.S.; Čapelja, E.V.; Karaman, M.A. Update on research data on the nutrient composition of mushrooms and their potentials in future human diets. In Food Chemistry, Function and Analysis, Edible Fungi: Chemical Composition, Nutrition and Health Effects; Stojković, D., Barros, L., Eds.; Royal Society of Chemistry: London, UK, 2022; Volume 2, pp. 27–67. [Google Scholar]
- Li, X.; Cheng, J.; Liu, X.; Guo, X.; Liu, Y.; Fan, W.; Lu, L.; Ma, Y.; Liu, T.; Tao, S.; et al. Origin and evolution of fusidane-type antibiotics biosynthetic pathway through multiple horizontal gene transfers. Genome Biol. Evol. 2020, 12, 1830–1840. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jiang, J.; Meng, Y.; Aihemaiti, A.; Xu, Y.; Xiang, H.; Gao, Y.; Chen, X. Preparation, environmental application and prospect of biochar-supported metal nanoparticles: A review. J. Hazard. 2020, 388, 122026. [Google Scholar] [CrossRef] [PubMed]
- Szwajgier, D. Anticholinesterase activity of selected phenolic acids and flavonoids-interaction testing in model solutions. Ann. Agric. Environ. Med. 2015, 22, 690–694. [Google Scholar] [CrossRef]
- Mandal, P. Screening, isolation and documentation of pharmaceutical importance macro fungi of kulikraiganj wildlife sanctuary, west bengal, india Parimal. Rev. Res. 2019, 7631, 1–9. [Google Scholar]
- Li, N.; Wang, C.; Georgiev, M.I.; Bajpai, V.K.; Tundis, R.; Simal-Gandara, J.; Lu, X.; Xiao, J.; Tang, X.; Qiao, X. Advances in dietary polysaccharides as anticancer agents: Structure-activity relationship. Trends Food Sci. Technol. 2021, 111, 360–377. [Google Scholar] [CrossRef]
Figure 1.
Percentage representation of bioactivities identified in S. commune from 2017 to 2022.

Figure 2.
Schizophyllan from S. commune.

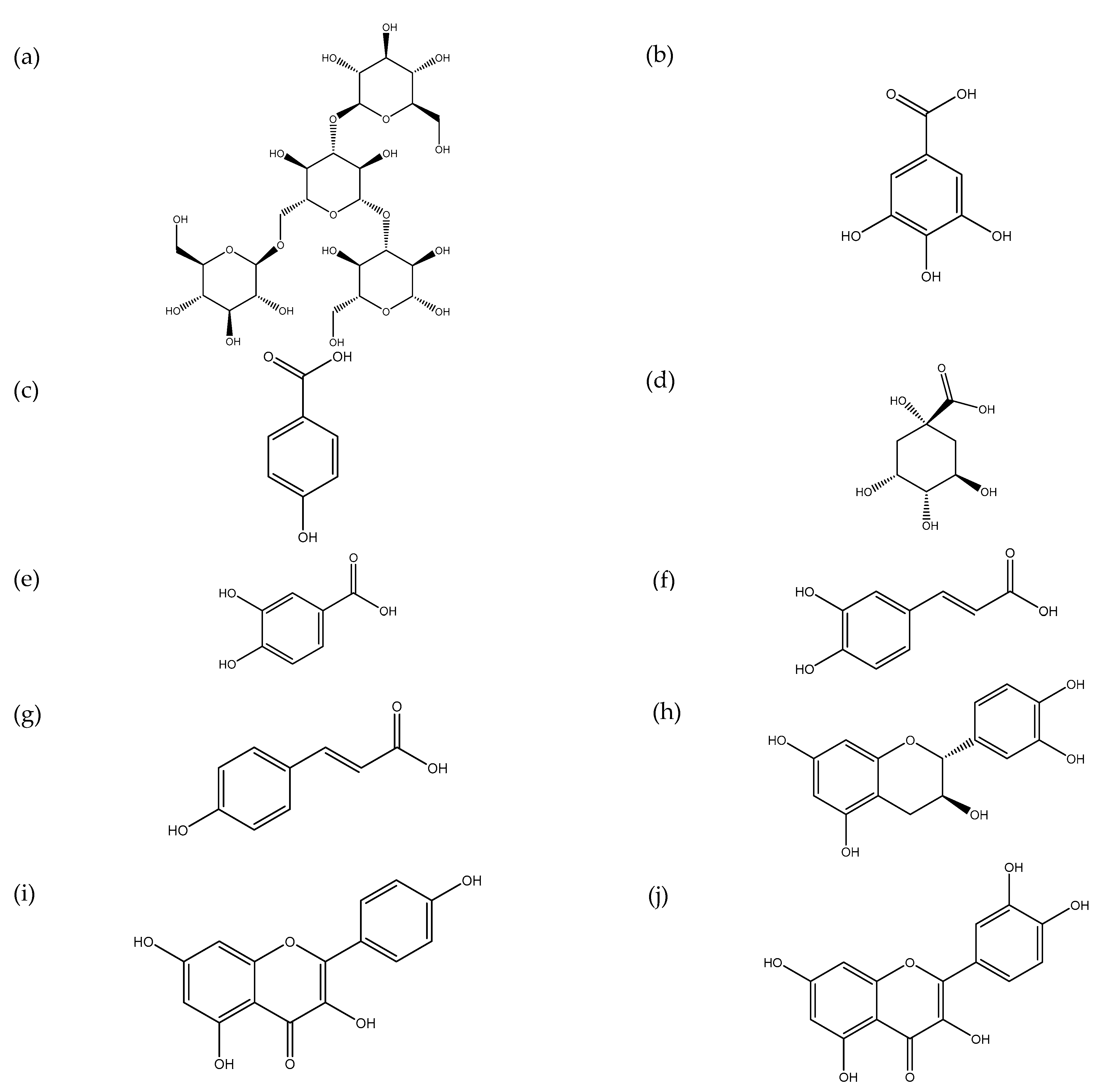
Figure 3.
Phenolic compounds from S. commune: (a) rutin; (b) gallic acid; (c) p-hydroxybenzoic acid; (d) quinic acid; (e) protocatechuic acid; (f) caffeic acid; (g) coumaric acid; (h) catechin; (i) kaempferol; (j) quercetin.
Figure 3.
Phenolic compounds from S. commune: (a) rutin; (b) gallic acid; (c) p-hydroxybenzoic acid; (d) quinic acid; (e) protocatechuic acid; (f) caffeic acid; (g) coumaric acid; (h) catechin; (i) kaempferol; (j) quercetin.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Mychochemical profile of primary and secondary metabolites and bioactivities of genus Schizophyllum Fr. (species name, region, year of publication, identified primary and secondary metabolites, content, and used references).
Table 1.
Mychochemical profile of primary and secondary metabolites and bioactivities of genus Schizophyllum Fr. (species name, region, year of publication, identified primary and secondary metabolites, content, and used references).
| Species | Region | Year of Publication | Primary Metabolites | Content/Molecular Weight | Secondary Metabolites | Content | Biological Activity | Reference |
|---|---|---|---|---|---|---|---|---|
| S. commune | India | 2017 | total carbohydrate | 42.0% | - | - | Antimicrobial and antioxidant | [9] |
| crude fiber | 30.0% | |||||||
| crude protein | 15.55% | |||||||
| crude fat | 9.0% | |||||||
| lipids | 0.4% | |||||||
| S. commune | China | 2017 | low-MW EPS | 197 kDa | - | - | Anti-inflammatory | [10] |
| medium-MW EPS | 936 kDa | |||||||
| high-MW EPS | 1437 kDa | |||||||
| S. commune | China | 2017 | EPS | 2900 kDa | - | - | Anti-inflammatory | [11] |
| S. commune | India | 2017 | - | - | flavonoids saponins steroids tannins triterpenoids | qualitative mycochemical analysis | Antimicrobial | [26] |
| S. commune | Czech Republic | 2017 | ultrasonicated schizophyllan | - | - | - | Immunomodulatory | [23] |
| S. commune | India | 2018 | proteins | qualitative mycochemical analysis | glycosides | qualitative mycochemical analysis | Antioxidant, cytotoxic and genotoxic | [27] |
| flavonoids: catechin epicatechin rutin quercetin kaempferol | ||||||||
| phenolics: gallic acid chlorogenic acid caffeic acid coumaric acid | ||||||||
| triterpenoids | ||||||||
| S. commune | Korea | 2019 | - | - | nerolidol mannoside, (mannonerolidol) | - | Antibacterial and antifungal | [28] |
| schizostatin | ||||||||
| nerolidol | ||||||||
| S. commune | Malaysia | 2019 | α-glucan | 1.4 ± 0.1% | total phenolics | 0.31 ± 0.05% | Antiviral (anti- dengue) | [12] |
| β-glucan | 34.4 ± 0.4% | |||||||
| protein | 15 ± 0.4% | |||||||
| S. commune | Thailand | 2019 | β-glucan | - | total phenolics | 284.41 ± 1.22 mg GAE/g d.w. | Antioxidant | [13] |
| S. commune | Laos | 2019 | crude-PSH MWs | 710.3 × 103 g/mol 170.6 × 103 g/mol 136.3 × 103 g/mol | - | - | Immune enhancing | [21] |
| fraction MWs | 255.0 × 103 g/mol 44.8 × 103 g/mol 533.7 × 103 g/mol 93.5 × 103 g/mol 79.8 × 103 g/mol | |||||||
| carbohydrates | 50.3–65.8% | |||||||
| proteins | 1.46–20.1% | |||||||
| sulphates | 1.33–7.01% | |||||||
| glucose | 17.0–88.2% | |||||||
| mannose | 2.70–55.2% | |||||||
| galactose | 6.2–19.1% | |||||||
| xylose | 0.90–8.57% | |||||||
| S. commune | Malaysia | 2020 | polysaccharides | 66.7–76.3 mg GE/g extract | total phenolics | 14.65 mg GAE/g extract | Antioxidant, anti-hyaluronidase, and anti-tyrosinase | [24] |
| total glucan | 34.72 ± 1.94% | |||||||
| α-glucan | 4.30 ± 0.40% | |||||||
| β-glucan | 29.97 ± 1.55% | |||||||
| S. commune | Brazil | 2020 | β-glucan | 13.14% | total phenolics | 299.31 ± 3.67 mg GAE/100 g | Antioxidant | [25] |
| S. commune | China | 2020 | polysaccharides | 45.09–67.96% | - | - | Antioxidant and hypoglycemic | [22] |
| protein | 2.67–3.53% | |||||||
| uronic acid | 1.92–3.0% | |||||||
| S. commune | Indonesia | 2020 | - | - | - | - | Antibacterial | [29] |
| S. commune | Georgia | 2020 | - | - | - | - | Antibacterial | [30] |
| S. commune | Thailand | 2020 | β-glucan | 5.04–32.94% | - | - | - | [19] |
| S. commune | Ukraine | 2020 | - | - | total flavonoids | 0.64 ± 0.16 QE/g | Antioxidant | [31] |
| S. commune | India | 2020 | peptides | - | Antioxidant | [14] | ||
| MYSEKHGSGGT | 1153.23 Da | |||||||
| PGTRGAIAASSPQV | 1311.44 Da | |||||||
| MVSTLAVLGIREP | 1385.67 Da | |||||||
| EKEAAELGKGSF | 1265.37 Da | |||||||
| MSVTLLLLFISLVWVTISGLN | 2319.84 Da | |||||||
| S. commune | Thailand | 2021 | - | - | total phenolics | 0.12–8.56 mg GAE/g d.w. | Antioxidant | [32] |
| total flavonoids | 53.06–577.35 μg CE/g d.w. | |||||||
| S. commune | Thailand | 2021 | β-1,3-glucans | 64.43–114.70 mg CE/g d.w. | total phenolics | 15.15–18.92 mg GAE/g d.w.), | Antioxidant | [33] |
| β-1,3-1,6-glucans | 5.42–7.79 mg SE/g d.w | total flavonoids | 422.54–910.54 CE/g d.w | |||||
| S. commune | Malaysia | 2021 | total protein | 1.30–2.17% | - | - | - | [34] |
| total amino acid | 308.65–443.84 mg/g | |||||||
| essential amino acid | 51.44–88.56 mg/g | |||||||
| nonessential amino acid | 151.48–265.32 mg/g | |||||||
| S. commune | Italy and Serbia | 2021 | Polysaccharides | - | total phenolics | 42.74–77.52 mg GAE/g d.w. (IT) | Antioxidant antibacterial, and antiacetylcholinesterase | [15] |
| 2.09–84.60 mg GAE/g d.w. (SRB) | ||||||||
| phenolics: | ||||||||
| p-hydroxybenzoic acid | 1.87–22.19 μg/g.d.w. | |||||||
| protocatechuic acid | 1.90–5.41 μg/g.d.w. | |||||||
| gallic acid | 75.77 μg/g d.w. | |||||||
| quinic acid | 4.29–20.06 μg/g d.w. | |||||||
| S. commune | Thailand | 2021 | α-glucan | 0.41–0.57% | total phenolics | 2.54–2.64 mg GAE/g d.w. | Antioxidant | [35] |
| β-glucan | 47.94–49.20% | |||||||
| S. commune | Germany | 2021 | - | - | sesquiterpenes: | - | Antibacterial and antifungal | [36] |
| (-)-(1R,2S)-β-bisabolol | ||||||||
| β-bisabolene | ||||||||
| (E)-γ-bisabolene | ||||||||
| S. commune | Japan | 2021 | - | - | volatile sulfur compounds: | - | - | [37] |
| dimethyl disulfide | ||||||||
| dimethyl trisulfide | ||||||||
| dimethyl tetrasulfide | ||||||||
| methyl ethyl disulfide | ||||||||
| diethyl disulfide | ||||||||
| H2S | ||||||||
| S. radiatum | Colombia | 2021 | total carbohydrate | 73–79% | - | - | Antitumor and immunostimulant | [1] |
| total glucans | 11–14% | |||||||
| α-glucan | 0.4–1.7% | |||||||
| β-glucan | 11–14% | |||||||
| proteins | 1% | |||||||
| S. commune | Philippines | 2022 | proteins | saponins tannins alkaloids, flavonoids, terpenoids | - | Cytotoxic | [18] | |
| carbohydrates | ||||||||
| S. commune | Malaysia | 2022 | - | - | - | - | Antioxidant and antimicrobial | [38] |
| S. commune | China | 2022 | - | - | fusidane-type antibiotics: | - | Antibacterial | [39] |
| (E)-4-(4-hydroxy-3-methylbut-2-en-1-yl) oxybenzoic acid | ||||||||
| methyl 4-(2,3-dihydroxy-3-methylbutoxy) benzoate | ||||||||
| ethyl 4-(2,3-dihydroxy-3-methylbutoxy) benzoate | ||||||||
| (R/S)-3-hydroxy-3-((R/S)-1-hydroxyethyl) indolin-2-one | ||||||||
| helvolinic acid | ||||||||
| 3,7-diketo-cephalosporin P1 | ||||||||
| (24R)-6β-hydroxy-24-ethylcholest-4-en-3-one | ||||||||
| dankasterone A | ||||||||
| (22E,24R)-3β-hydroxyergosta-7,22-diene-6-one | ||||||||
| (3β,22E)-ergosta-5,7,22-trien-3-ol | ||||||||
| juglansnoid B | ||||||||
| trans-ferulic acid | ||||||||
| (S)-3-hydroxy-3-(2-oxopropyl) indolin-2-one | ||||||||
| schizostatin | ||||||||
| (13S)-8-oxo-(9E,11E)-8-oxo-octadeca-9,11-dien-13-olide | ||||||||
| S. commune | India | 2022 | total carbohydrate | 0.597 ± 0.06 g/100 g d.w. | total phenolics | 2.00 ± 0.05 mg GAE/g d.w. | Antioxidant and antibacterial | [40] |
| total flavonoids | 0.04 ± 0.00 mg QE/g d.w. | |||||||
| S. commune | Thailand | 2022 | schizophyllan | - | total phenolics in schizophyllan, | 6.22 ± 0.06 mg GAE/g | Antioxidant | [41] |
| total phenolics in supernatant | 12.85 ± 0.12 mg GAE/g | |||||||
| total phenolics in mushroom essence | 14.07 ± 0.09 mg GAE/g of dry extract | |||||||
| S. commune | Turkey | 2022 | - | - | - | - | Antimicrobial and anticancer | [42] |
| S. commune | Review | 2022 | hemolysin monomer ribonuclease schizophyllan | 29 kDa 20 kDa | - | - | Antiviral | [43] |
| S. commune | Malaysia and Thailand | 2022 | total carbohydrate | 229.95–596.22 mg carbohydrate/g extract | total phenolics | 6.43–37.87 mg GAE/g extract | Antioxidant | [44] |
| total proteins | 395.75–539.69 mg protein/g extract | total flavonoids | 5.77–84.55 mg RE/g extract |
—no data; EPS—exopolysaccharide; MW—molecular weight; d.w.—dry weight; PSH—polysaccharide; schizophyllan (SPG); GAE—gallic acid equivalents; GE—glucose equivalents; QE—quercetin equivalents; CE—catechin equivalents; SE—schizophyllan equivalents; IT—Italy; SRB—Serbia; amino acid abbreviations: M—Met; Y—Tyr; S—Ser; E—Glu; K—Lys; H—His; G—Gly; S—Ser; T—Thr; P—Pro; R—Arg; A—Ala; I—Ile; Q—Gln; V—Val; L—Leu; F—Phe; W—Trp; N—Asn.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mišković, J.; Rašeta, M.; Krsmanović, N.; Karaman, M. Update on Mycochemical Profile and Selected Biological Activities of Genus Schizophyllum Fr. 1815. Microbiol. Res. 2023, 14, 409-429. https://doi.org/10.3390/microbiolres14010031
AMA Style
Mišković J, Rašeta M, Krsmanović N, Karaman M. Update on Mycochemical Profile and Selected Biological Activities of Genus Schizophyllum Fr. 1815. Microbiology Research. 2023; 14(1):409-429. https://doi.org/10.3390/microbiolres14010031
Chicago/Turabian StyleMišković, Jovana, Milena Rašeta, Nenad Krsmanović, and Maja Karaman. 2023. "Update on Mycochemical Profile and Selected Biological Activities of Genus Schizophyllum Fr. 1815" Microbiology Research 14, no. 1: 409-429. https://doi.org/10.3390/microbiolres14010031