
Effects of Cr Stress on Bacterial Community Structure and Composition in Rhizosphere Soil of Iris tectorum under Different Cultivation Modes
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sampling and Chemical Analysis
2.3. DNA Extraction, 16S rRNA Gene Sequencing, and Shotgun Metagenome Sequencing
2.4. Statistical Analysis
3. Results
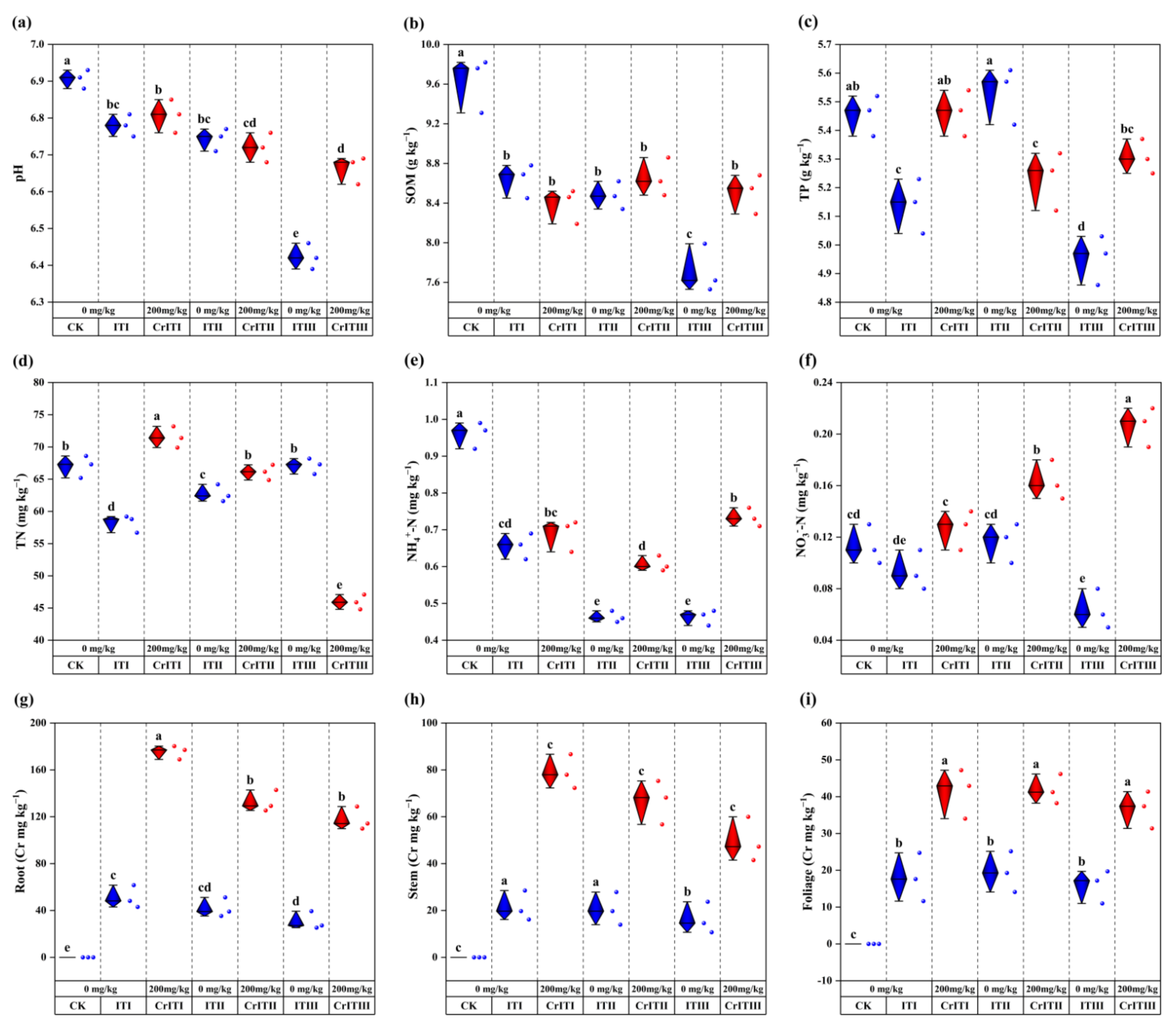
3.1. Effects of Cr stress on Soil Physicochemical Properties and Heavy Metal Accumulation in Plants
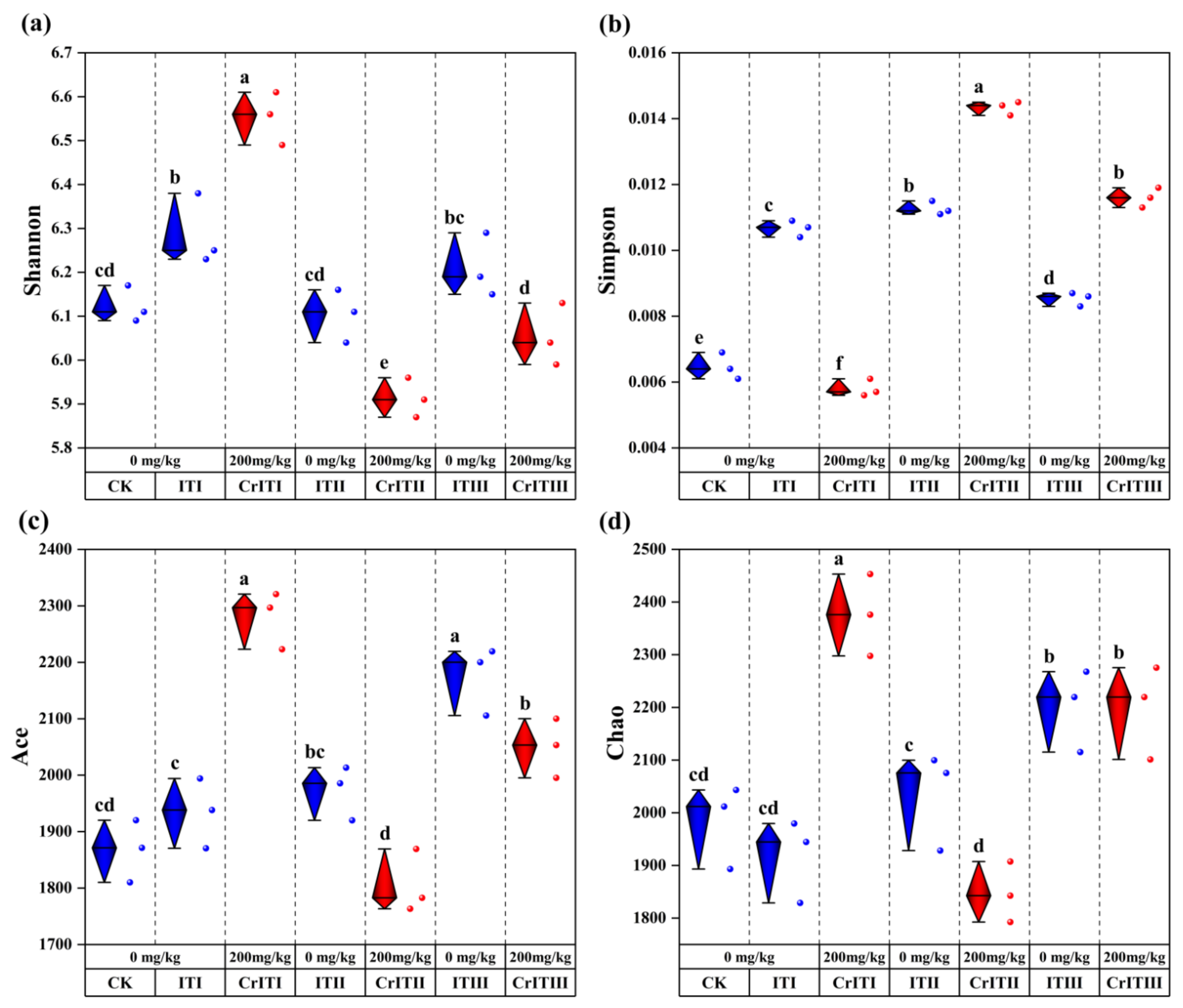
3.2. Rhizosphere Bacterial Diversity Index
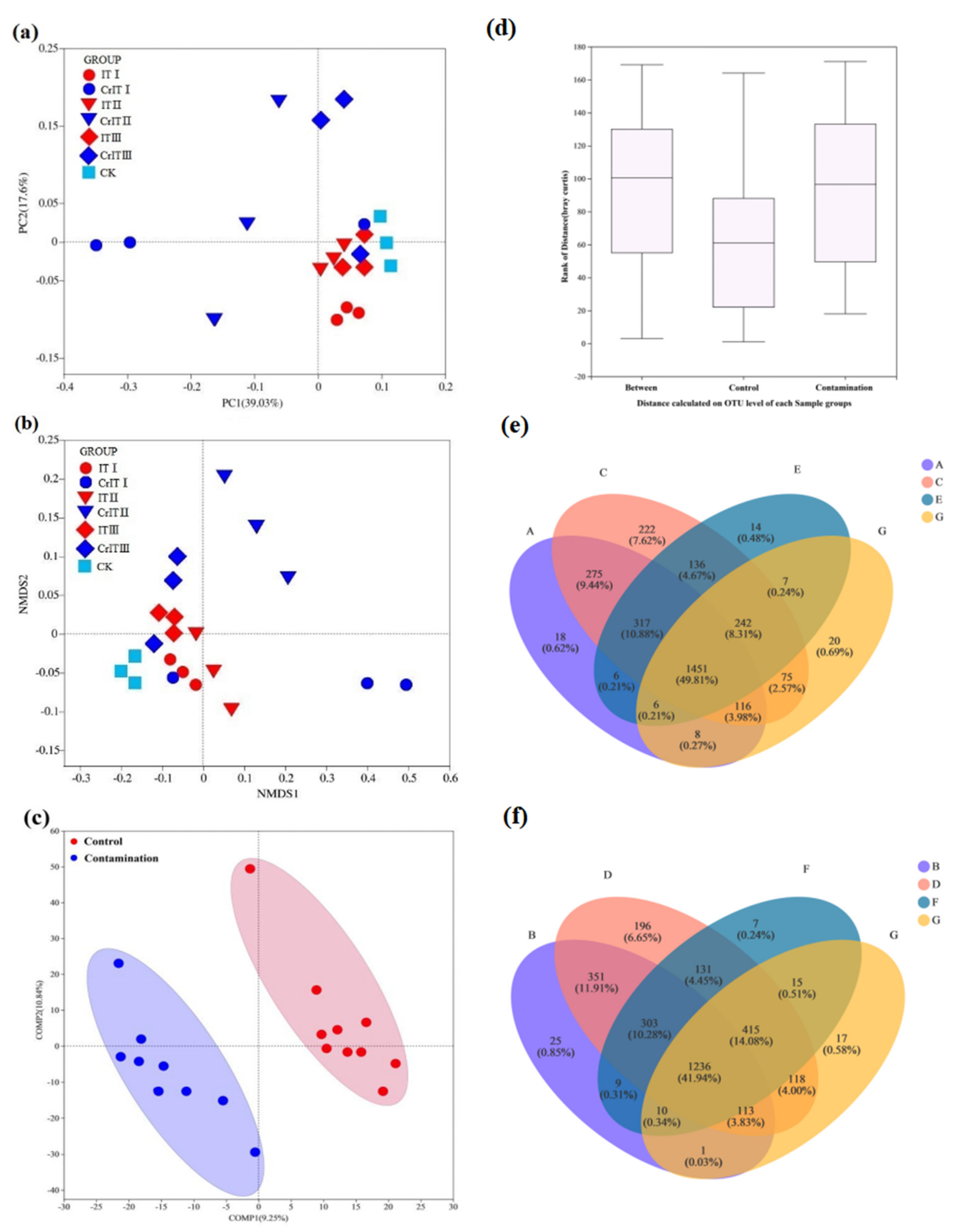
3.3. Bacterial Community Structure in Rhizosphere Soil
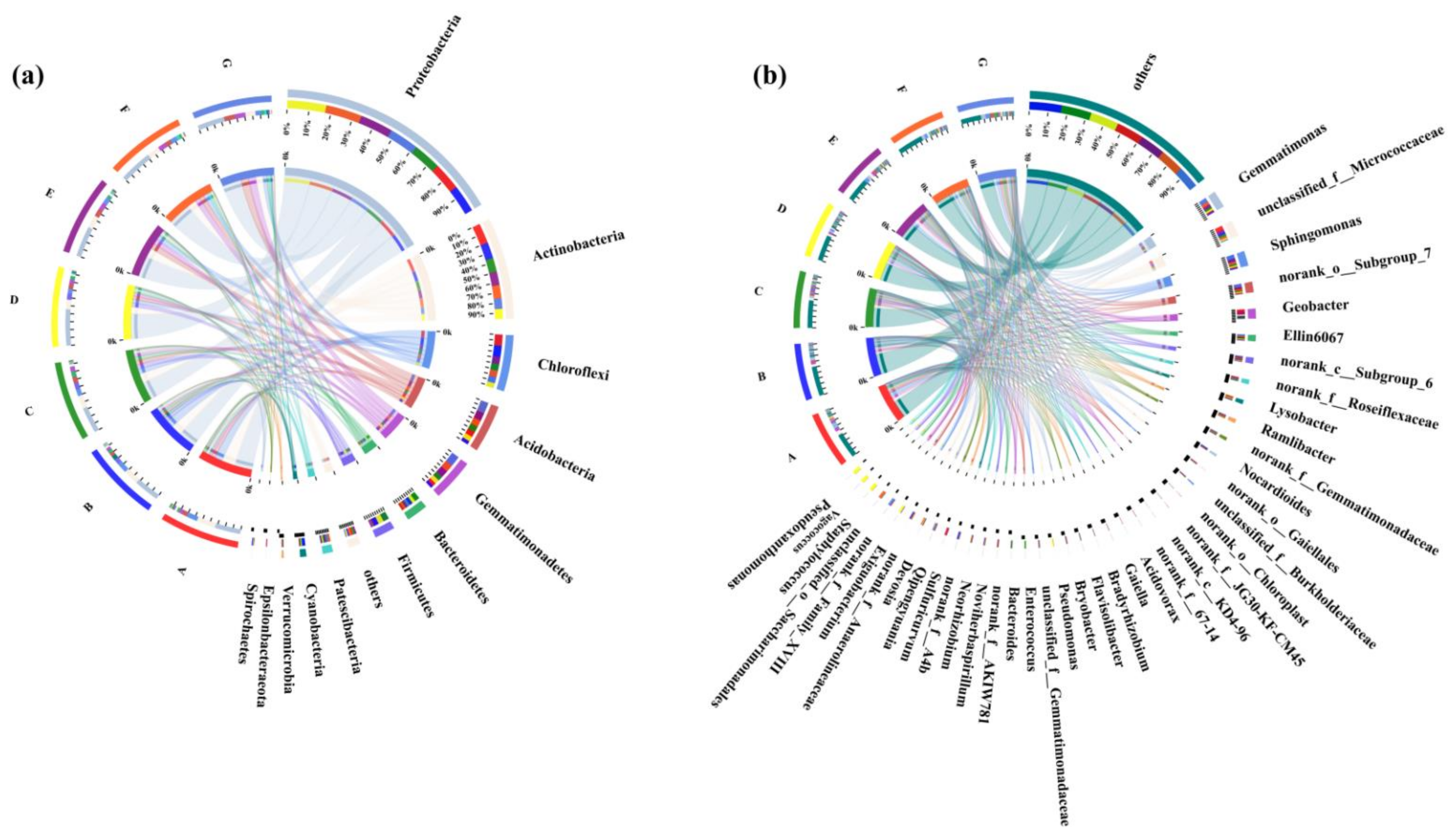
3.4. Bacterial Community Composition in Rhizosphere Soil
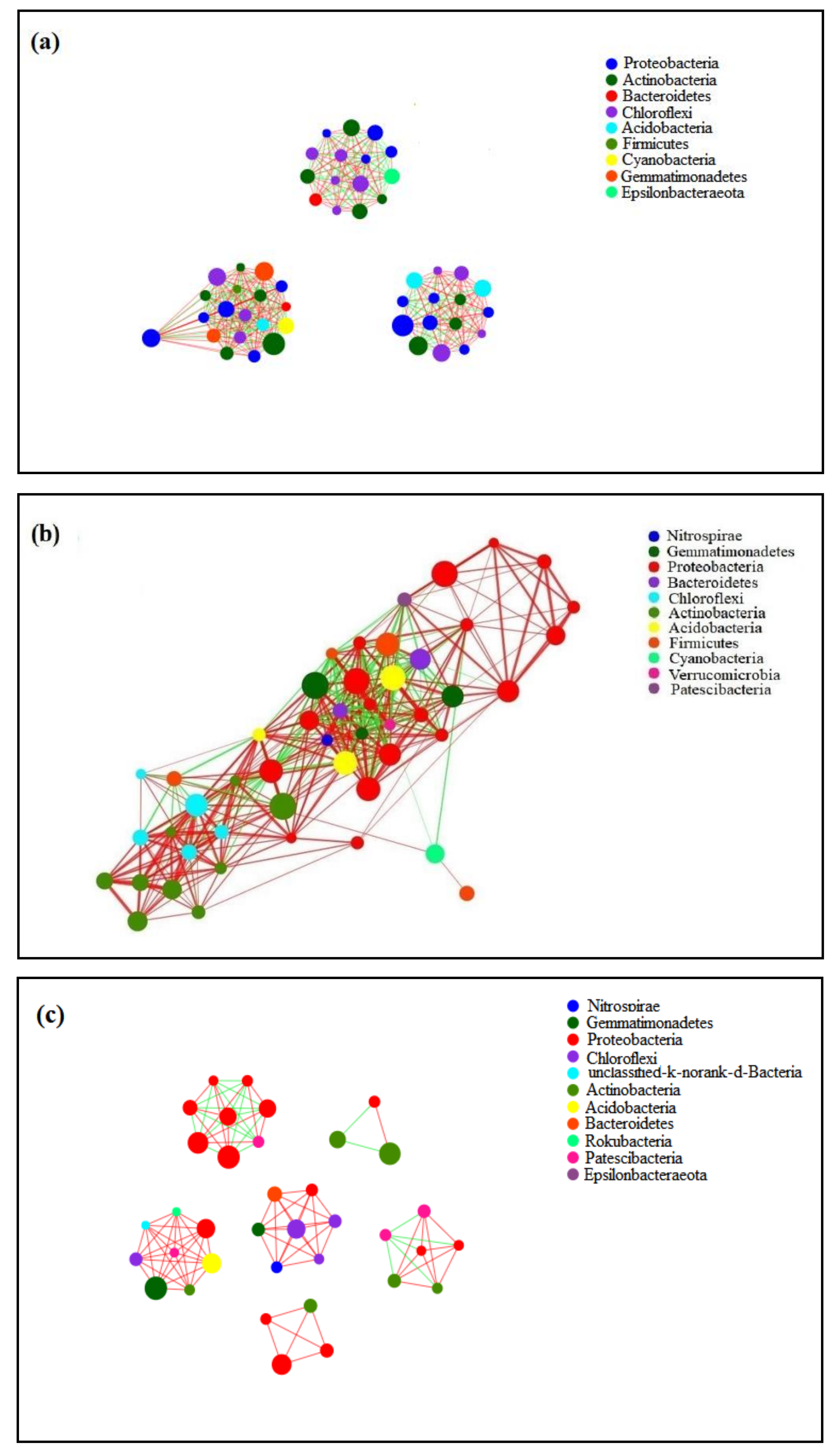
3.5. Interspecific Symbiotic Network of Soil Bacterial Communities
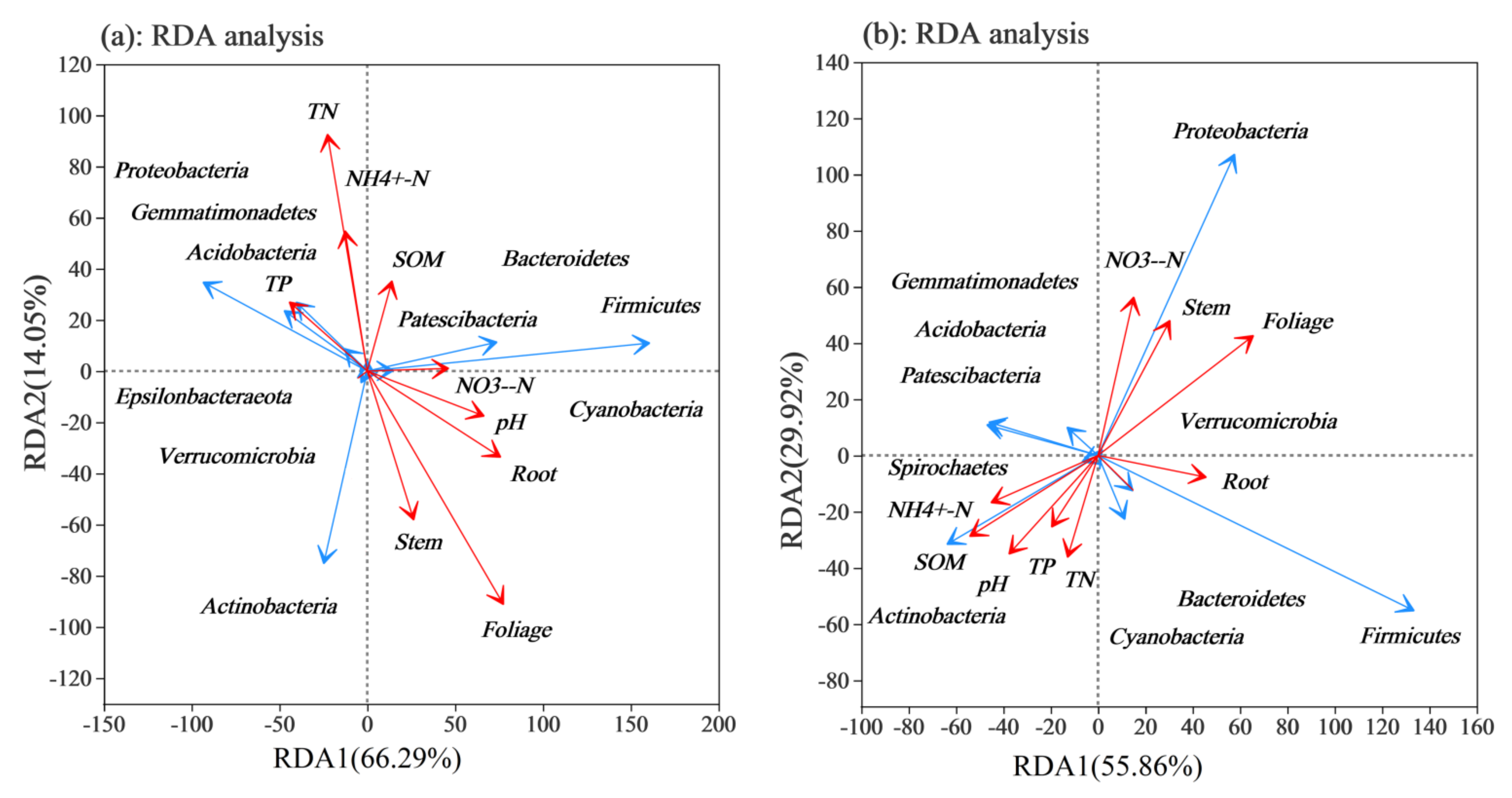
3.6. Correlation Analysis between Rhizosphere Soil Bacterial Community and Physicochemical Properties
4. Discussion
4.1. Physiological Response of I. tectorum to Cr Stress
4.2. Effects of Cr Stress on Soil Bacterial Community Structure
4.3. Effects of Cr Stress on Rhizosphere Bacteria
4.4. Effect of Different Cultivation Patterns on Rhizosphere Bacteria
4.5. Effects of Soil Physicochemical Properties on Rhizosphere Bacterial Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaheer, I.E.; Shafaqat, A.; Muhammad, H.S.; Muhammad, I.; Ghalia, S.H.A.; Basmah, M.A.; Muhammad, R.; Zohaib, A.; Muhammad, R.; Mona, H.S. Role of iron-lysine on morpho-physiological traits and combating chromium toxicity in rapeseed (Brassica napus L.) plants irrigated with different levels of tannery wastewater. Plant Physiol. Biochem. 2020, 155, 70–84. [Google Scholar] [CrossRef]
- Riaz, M.; Yasmeen, T.; Arif, M.S.; Ashraf, M.A.; Hussain, Q.; Shahzad, S.M. Var-iations in morphological and physiological traits of wheat regulated by chromium species in long-term tannery effluent irrigated soils. Chemosphere 2019, 222, 891–903. [Google Scholar] [CrossRef]
- Shen, L.; Saky, S.A.; Yang, Z.; Ho, S.H.; Chen, C.X.; Qin, L.; Zhang, G.L.; Wang, Y.P.; Lu, Y.H. The critical utilization of active heterotrophic microalgae for bioremoval of Cr(VI) in organics co-contaminated wastewater. Chemosphere 2019, 228, 536–544. [Google Scholar] [CrossRef]
- Muhammad, R.; Muhammad, K.; Fang, Y. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2020, 402, 123919. [Google Scholar]
- Kisholay, M.; Suchismita, D. Phytoremediation of soil treated with metalliferous leachate from an abandoned industrial site by Alternanthera sessilis and Ipomoea aquatica: Metal extraction and biochemical responses. Ecol. Eng. 2021, 170, 106349. [Google Scholar]
- Wang, B.C.; Zhu, S.X.; Li, W.J.; Tang, Q.; Luo, Y.H. Effects of chromium stress on the rhizosphere microbial community composition of Cyperus alternifolius. Ecotoxicol. Environ. Saf. 2021, 218, 112253. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P. Efficiency of bacteria and bacterial assisted phytoremediation of heavy metals: An update. Bioresour. Technol. 2021, 328, 124835. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; Abrantes, I.; Barra Caracciolo, A.; Bevivino, A.; Ciancio, A.; Grenni, P.; Hrynkiewicz, K.; Kredics, L.; Proenca, D.N. Belowground microbiota and the health of tree crops. Front. Microbiol. 2018, 9, 1006. [Google Scholar] [CrossRef]
- Badri, D.V.; Weir, T.L.; van der Lelie, D.; Vivanco, J.M. Rhizosphere chemical dialogues: Plant-microbe interactions. Curr. Opin. Biotechnol. 2009, 20, 642–650. [Google Scholar] [CrossRef]
- Xue, J.L.; Shi, K.; Chen, C.; Bai, Y.; Cui, Q.; Li, N.; Fu, X.; Qiao, Y. Evaluation of response of dynamics change in bioaugmentation process in diesel-polluted seawater via high-throughput sequencing: Degradation characteristic, community structure, functional genes. J. Hazard. Mater. 2021, 403, 123569. [Google Scholar] [CrossRef]
- Barra Caracciolo, A.; Terenzi, V. Rhizosphere Microbial Communities and Heavy Metals. Microorganisms 2021, 9, 1462. [Google Scholar] [CrossRef]
- Younas, F.; Niazi, N.K.; Bibi, I.; Afzal, M.; Hussain, K.; Shahid, M.; Aslam, Z.; Bashir, S.; Hussain, M.M.; Bundschuh, J. Constructed wetlands as a sustainable technology for wastewater treatment with emphasis on chromium-rich tannery wastewater. J. Hazard. Mater. 2022, 422, 126926. [Google Scholar] [CrossRef]
- Manoj, S.R.; Karthik, C.; Kadirvelu, K.; Arulselvi, P.I.; Shanmugasundaram, T.; Bruno, B.; Rajkumar, M. Understanding the molecular mechanisms for the enhanced phytoremediation of heavy metals through plant growth promoting rhizobacteria: A review. J. Environ. Manag. 2020, 254, 109779. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M. Enhancing phytoremediation of chromium-stressed soils through plant-growth-promoting bacteria. J. Genet. Eng. Biotechnol. 2015, 13, 51–58. [Google Scholar] [CrossRef]
- Xiao, W.; Ye, X.; Ye, Z.; Zhang, Q.; Zhao, S.; Chen, D. Responses of microbial community composition and function to biochar and irrigation management and the linkage to Cr transformation in paddy soil. Environ. Pollut. 2022, 304, 119232. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, N.E.A.; El-Khateeb, A.Y.; Ghoniem, A.A.; El-Hersh, M.S.; Saber, W.I. Innovative low-cost biosorption process of Cr6+ by Pseudomonas alcaliphila NEWG-2. Sci. Rep. 2020, 10, 14043. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; You, S.H.; Liu, L.H. Characterization of Microbial Communities, Identification of Cr(Ⅵ) Reducing Bacteria in constructed Wetland and Cr(Ⅵ) Removal Ability of Bacillus cereus. Sci. Rep. 2019, 9, 12873. [Google Scholar] [CrossRef]
- Wendkuuni, F.C.; Ann, D.; Diederik, P.L.R. Metal uptake by spontaneously grown Typha domingensis and introduced Chrysopogon zizanioides in a constructed wetland treating gold mine tailing storage facility seepage. Ecol. Eng. 2020, 158, 106037. [Google Scholar]
- Mufarrege, M.D.L.M.; di Luca, G.A.; Hadad, H.R.; Maine, M.A. Exposure of Typha domingensis to high concentrations of multi-metal and nutrient solutions: Study of tolerance and removal efficiency. Ecol. Eng. 2021, 159, 106118. [Google Scholar] [CrossRef]
- Shi, Y.M.; Wang, W.L.; Chen, S.H.; Lv, H.F.; Ma, C.D.; Ma, N.; Gao, J.Q. Study on the purification performance of constructed wetland with iris sibirica for treating simulated Cd contaminated river water. Environ. Sci. 2016, 34, 21–24+40. [Google Scholar]
- Liu, Q.Q.; Zhang, Y.X.; Wang, Y.; Wang, W.; Gu, C.; Huang, S.Z.; Yuan, H.Y.; Om, P.D. Quantitative proteomic analysis reveals complex regulatory and metabolic response of Iris lactea Pall. var. chinensis to cadmium toxicity. J. Hazard. Mater. 2020, 400, 123165. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.S.; Yang, G.Z.; Jiang, D.Z.; Liu, Z.H.; Sun, B.M. Basic statistics and characteristics of background values of soil elements in China. China Natl. Environ. Monit. Cent. 1991, 7, 1–6. [Google Scholar]
- Lin, Y.S.; Hua, X.M.; San, Y.H.; Zhang, X.F. Soil Environmental Quality Standards for Producing Areas of Agricultural Products; Nanjing Institute of Environmental Sciences, State Environmental Protection Administration: Jiangsu Province, Nanjing, China, 2007. [Google Scholar]
- Wang, A.-Y.; Huang, S.S.; Zhong, G.F.; Xu, G.B.; Liu, Z.X.; Shen, X.B. Effect of Cr(Ⅵ) Stress on Growth of Three Herbaceous Plants and Their Cr Uptake. Environ. Sci. 2012, 33, 2028–2037. [Google Scholar]
- Hu, S.; Hu, B.; Chen, Z.; Vosátka, M.; Vymazal, J. Arbuscular Mycorrhizal Fungi Modul. Chromium Distrib. Bioavailab. Semi-Aquat. Habitats. Chem. Eng. J. 2021, 420, 129925. [Google Scholar] [CrossRef]
- Zhang, X.X.; Zhang, R.J.; Gao, J.S.; Wang, X.C.; Fan, F.L.; Ma, X.T.; Yin, H.Q.; Zhang, C.W.; Feng, K.; Deng, Y. Thirty-one years of rice-rice-green manure rotations shape the rhizosphere microbial community and enrich beneficial bacteria. Soil Biol. Biochem. 2017, 104, 208–217. [Google Scholar] [CrossRef]
- Bao, S.D. Analytical Methods of Soil Agro-Chemistry; China Agriculture Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Seang-On, L.; Meeinkuirt, W.; Saengwilai, P.; Saminpanya, S.; Koedrith, K. Alleviating of cadmium stress in Thai rice cultivar (psl2) by inoculation of indigenous cadmium-resistant microbial consortia. Appl. Ecol. Environ. Res. 2019, 17, 14679–14697. [Google Scholar] [CrossRef]
- Bray, R.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Shahid, M.; Shamshad, S.; Rafiq, M.; Khalid, S.; Bibi, I.; Niazi, K.N.; Dumat, C.; Rashid, M.I. Chromium speciation, bioavailability, uptake, toxicity and detoxification in soil-plant system: A review. Chemosphere 2017, 178, 513–533. [Google Scholar] [CrossRef]
- Zong, H.; Liu, J.; Wang, F.; Song, N. Root morphological response of six peanut cultivars to chromium (VI) toxicity. Environ. Sci. Pollut. Res. 2020, 27, 18403–18411. [Google Scholar] [CrossRef]
- Yuan, Y.; Muhammad, I.; Muhammad, R.; Zhihua, D.; Md, M.H.; Yihui, Z.; Hengliang, H.; Shuxin, T. The role and its transcriptome mechanisms of cell wall polysaccharides in vanadium detoxication of rice. J. Hazard. Mater. 2022, 425, 127966. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Yasmeen, T.; Arif, M.S.; Riaz, M.; Shahzad, S.M.; Imran, Q.; Ali, I. Combined ability of chromium (Cr) tolerant plant growth promoting bacteria (PGPB) and salicylic acid (SA) in attenuation of chromium stress in maize plants. Plant Physiol. Biochem. 2016, 108, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Singh, C.K.; Singh, D.; Sarkar, S.K.; Prasad, S.K.; Ishwar Singh, N.L.S. Glycine betaine modulates chromium (VI)-induced morpho-physiological and biochemical responses to mitigate chromium toxicity in chickpea (Cicer arietinum L.) cultivars. Sci. Rep. 2022, 12, 8005. [Google Scholar] [CrossRef] [PubMed]
- Sokol, N.W.; Slessarev, E.; Marschmann, G.L.; Alexa, N.; Steven, J.; Blazewicz, E.L.; Brodie, M.K.; Firestone, M.M.; Foley, R.H.; Bruce, A.; et al. Life and death in the soil microbiome: How ecological processes influence biogeochemistry. Nat. Rev. Microbiol. 2022, 22, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Vijay, S.M.; Sunita, K.M.; Jay, P.V. Plant beneficial rhizospheric microorganism (PBRM) strategies to improve nutrients use efficiency: A review. Ecol. Eng. 2017, 107, 8–32. [Google Scholar]
- Yu, H.; An, Y.J.; Jin, D.C.; Jin, T.; Wang, X.R. Effects of Chromium Pollution on Soil Bacterial Community Structure and Assembly Processes. Environ. Sci. 2021, 42, 1197–1204. [Google Scholar]
- Sun, F.L.; Fan, L.L.; Wang, Y.S.; Huang, L.Y. Metagenomic analysis of the inhibitory effect of chromium on microbial communities and removal efficiency in A2O sludge. J. Hazard. Mater. 2019, 368, 523–529. [Google Scholar] [CrossRef]
- Guo, S.Y.; Xiao, C.Q.; Zhou, N.; Chi, R. Speciation, toxicity, microbial remediation and phytoremediation of soil chromium contamination. Environ. Chem. Lett. 2020, 19, 1413–1431. [Google Scholar] [CrossRef]
- Huang, L.; Wang, X.; Chi, Y.; Huang, L.; Li, W.C.; Ye, Z. Rhizosphere bacterial community composition affects cadmium and arsenic accumulation in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2021, 222, 112474. [Google Scholar] [CrossRef]
- Joutey, N.T.; Sayel, H.; Bahafid, W.; El, G.N. Mechanisms of hexavalent chromium resistance and removal by microorganisms. Rev. Environ. Contam. Toxicol. 2015, 233, 45–69. [Google Scholar]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef] [PubMed]
- He, C.W.; Gu, L.P.; Xu, Z.X.; He, H.; Fu, G.; Han, F.X.; Huang, B.; Pan, X.J. Cleaning chromium pollution in aquatic environments by biore-mediation, photocatalytic remediation, electrochemical remediation and coupled remediation systems. Environ. Chem. Lett. 2020, 18, 561–576. [Google Scholar] [CrossRef]
- Munees, A.; Mulugeta, K. Recent Trends in Microbial Biosorption of Heavy Metals: A Review. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2013, 1, 19–26. [Google Scholar]
- Sharma, N.; Sodhi, K.K.; Kumar, M.; Singh, D.K. Heavy metal pollution: Insights into chromium eco-toxicity and recent advancement in its remediation. Environmental Nanotechnology. Monit. Manag. 2021, 15, 100388. [Google Scholar] [CrossRef]
- Ramesh, C.P.; Seidu, M.; Michael, B.; Mallavarapu, M.; Ravi, N. Molecular characterization of chromium (VI) reducing potential in Gram positive bacteria isolated from contaminated sites. Soil Biol. Biochem. 2010, 42, 1857–1863. [Google Scholar]
- Li, X.G.; Jousset, A.; de Boer, W.; Carrion, V.J.; Zhang, T.L.; Wang, X.X.; Kuramae, E.E. Legacy of land use history determines reprogramming of plant physiology by soil microbiome. ISME J. 2019, 13, 738–751. [Google Scholar] [CrossRef]
- Hyeseon, C.; Franz, K.G.; Minsu, J. Evaluation of bacterial community in constructed wetlands treating different sources of wastewater. Ecol. Eng. 2022, 182, 106703. [Google Scholar]
- Zhou, Z.B.; Zhang, Y.J.; Zhang, F. Abundant and rare bacteria possess different diversity and function in crop monoculture and rotation systems across regional farmland. Soil Biol. Biochem. 2022, 171, 108742. [Google Scholar] [CrossRef]
- Yuan, Q.; Liu, P.; Lu, Y. Differential responses of nirK- and nirS-carrying bacteria to denitrifying conditions in the anoxic rice field soil. Env. Microbiol. 2012, 4, 113–122. [Google Scholar] [CrossRef]
- Shu, W.S.; Huang, L.N. Microbial diversity in extreme environments. Nat. Rev. Microbiol. 2022, 20, 219–235. [Google Scholar] [CrossRef]
- Garavaglia, L.; Cerdeira, S.B.; Vullo, D.L. Chromium (VI) biotransformation by β-and γ-Proteobacteria from natural polluted environments: A combined biological and hemical treatment for industrial wastes. J. Hazard. Mater. 2010, 175, 104–110. [Google Scholar] [CrossRef]
- Lopez, S.; Piutti, S.; Vallance, J.; Morel, J.-L.; Echevarria, G.; Benizri, E. Nickel drives bacterial community diversity in the rhizosphere of the hyperaccumulator Alyssum murale. Soil Biol. Biochem. 2017, 114, 121–130. [Google Scholar] [CrossRef]
- Eichorst, S.A.; Trojan, D.; Roux, S.; Herbold, C.; Rattei, T.; Woebken, D. Genomic insights into the Acidobacteria reveal strategies for their success in terrestrial environments. Environ. Microbiol. 2018, 20, 1041–1063. [Google Scholar] [CrossRef] [PubMed]
- Hug, L.A.; Castelle, C.J.; Wrighton, K.C.; Thomas, B.C.; Sharon, I.; Frischkorn, K.R.; Williams, K.H.; Tringe, S.G.; Banfield, J.F. Community genomic analyses constrain the dis-tribution of metabolic traits across the chloroflexi phylum and indicate roles in sedi-ment carbon cycling. Microbiome 2013, 1, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Z.; Sixi, Z.; Baojing, G.; Xiuqing, Y.; Guodong, X.; Baichun, W. Effects of Cr Stress on Bacterial Community Structure and Composition in Rhizosphere Soil of Iris tectorum under Different Cultivation Modes. Microbiol. Res. 2023, 14, 243-261. https://doi.org/10.3390/microbiolres14010020
Wei Z, Sixi Z, Baojing G, Xiuqing Y, Guodong X, Baichun W. Effects of Cr Stress on Bacterial Community Structure and Composition in Rhizosphere Soil of Iris tectorum under Different Cultivation Modes. Microbiology Research. 2023; 14(1):243-261. https://doi.org/10.3390/microbiolres14010020
Chicago/Turabian StyleWei, Zhao, Zhu Sixi, Gu Baojing, Yang Xiuqing, Xia Guodong, and Wang Baichun. 2023. "Effects of Cr Stress on Bacterial Community Structure and Composition in Rhizosphere Soil of Iris tectorum under Different Cultivation Modes" Microbiology Research 14, no. 1: 243-261. https://doi.org/10.3390/microbiolres14010020