
Bacterial Community Drives the Carbon Source Degradation during the Composting of Cinnamomum camphora Leaf Industrial Extracted Residues
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling
2.2. Determination of Basic Physicochemical Properties and Enzyme Activities
2.3. The DNA Sequencing and Bioinformatic Analysis
2.4. The Statistics
3. Results
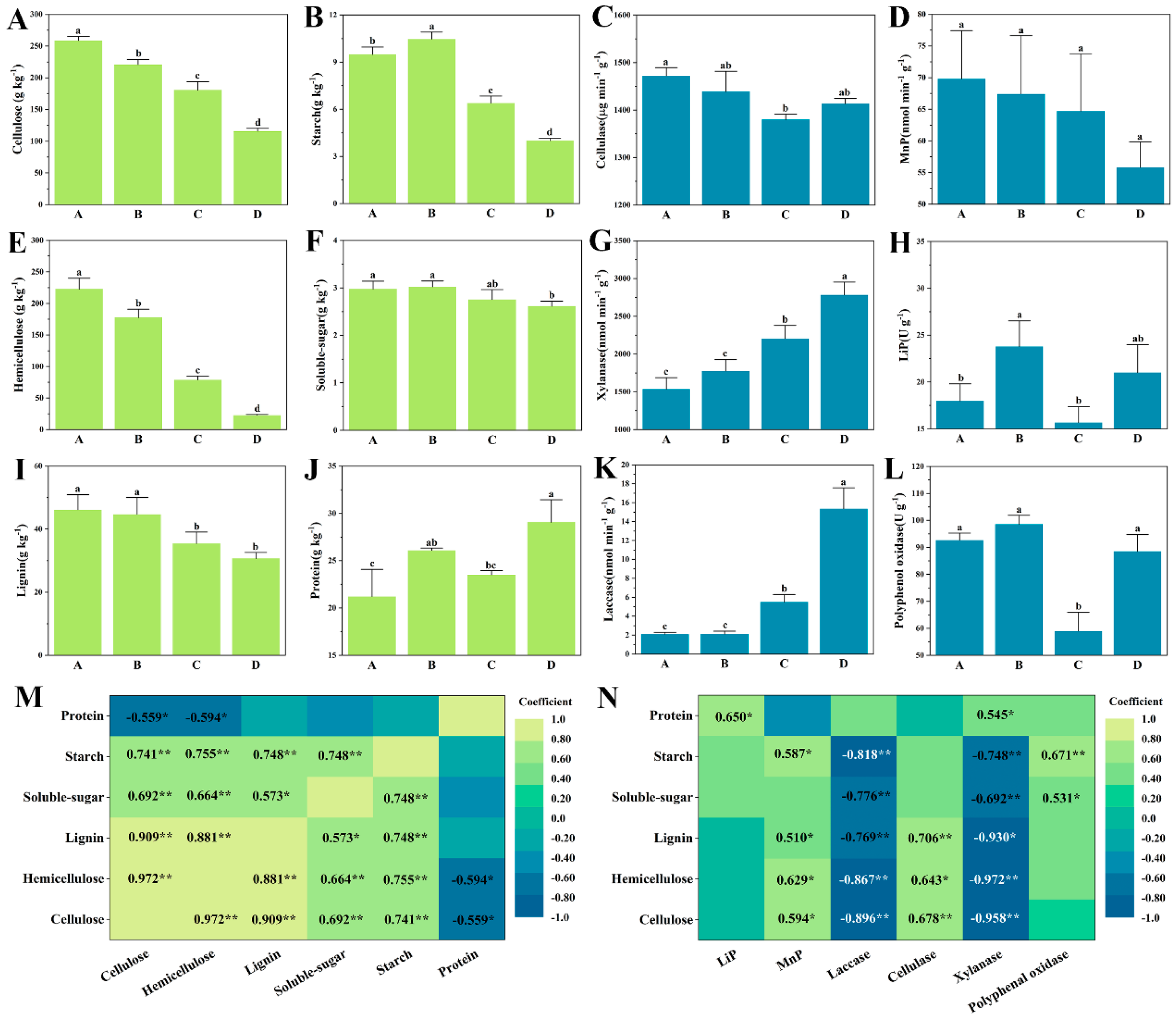
3.1. The Variation on Carbon Source and Enzymes
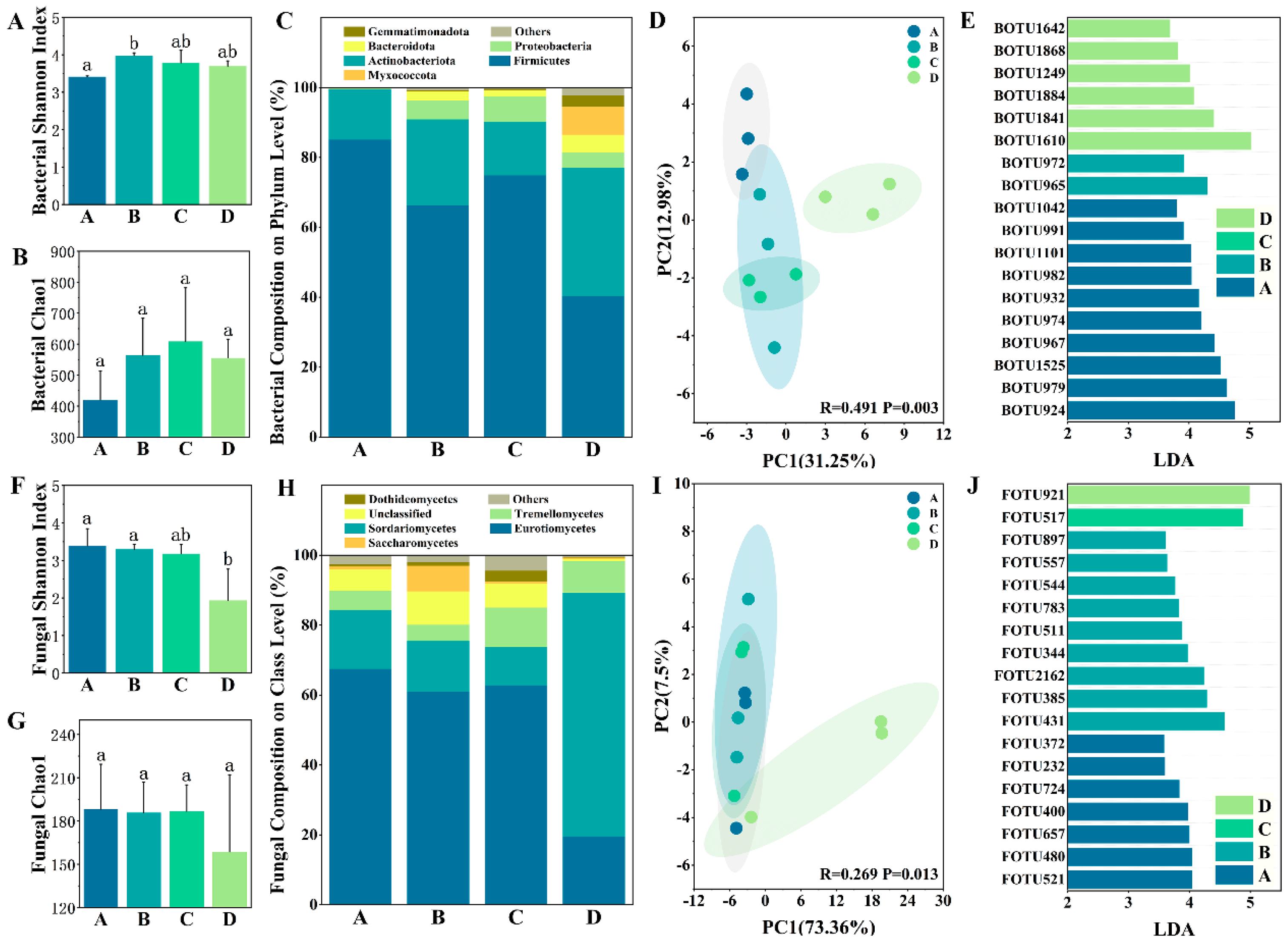
3.2. Microbial Community Composition Variation
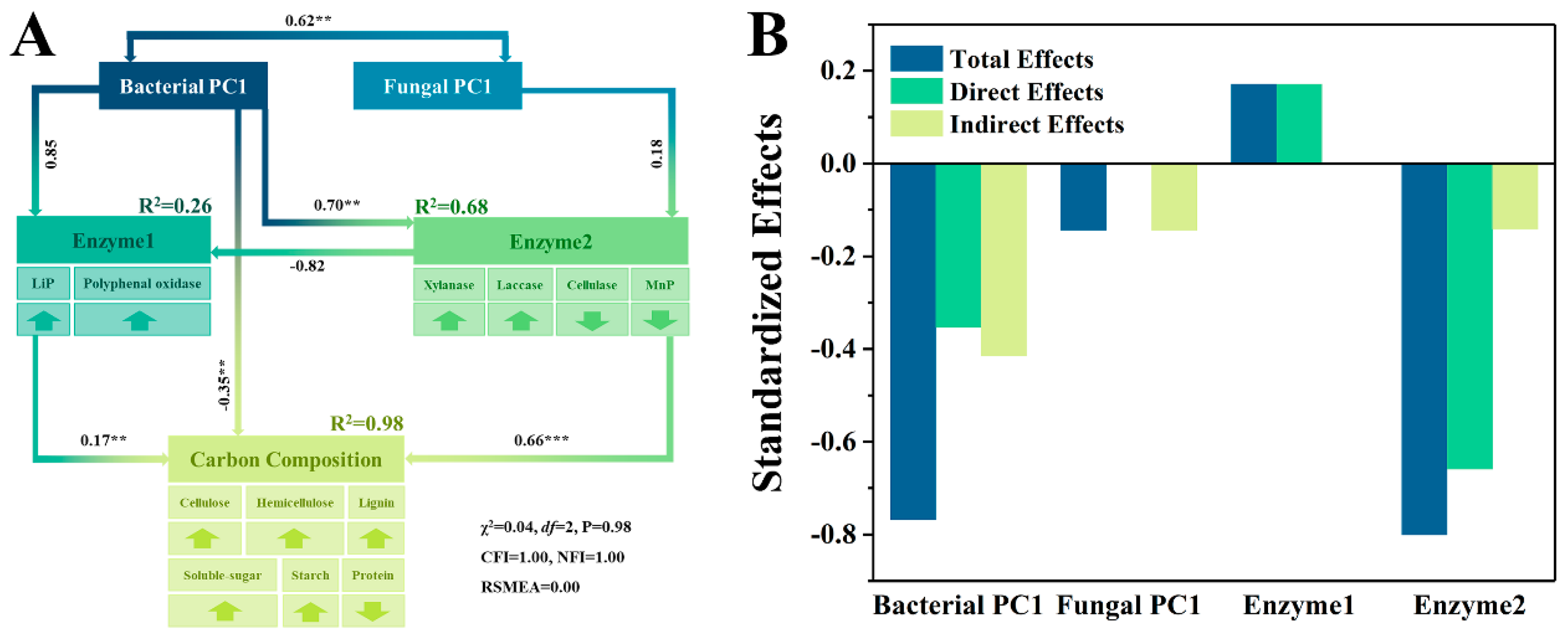
3.3. Contribution to the Variation of Carbon Source Composition
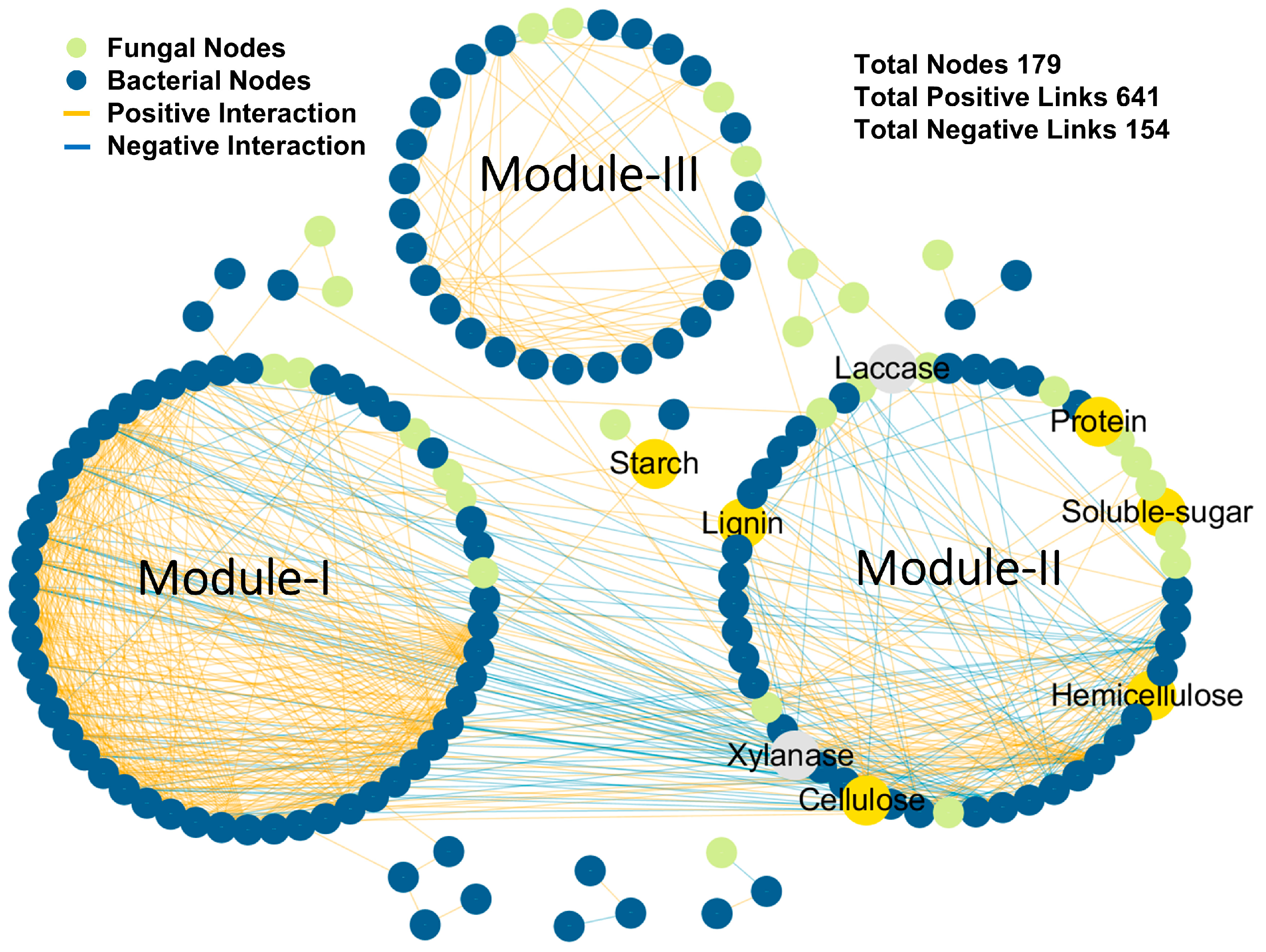
3.4. Network Analysis
4. Discussion
4.1. The Variation of Carbon Sources and Enzyme Activities during Composting
4.2. Relationship between Microbial Community and Carbon Source Degradation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rao, K.S.; Haran, R.H.; Rajpoot, V.S. Value addition: A novel strategy for quality enhancement of medicinal and aromatic plants. J. Appl. Res. Med. Aromat. Plants 2022, 31, 100415. [Google Scholar] [CrossRef]
- Charles Dorni, A.I.; Amalraj, A.; Gopi, S.; Varma, K.; Anjana, S.N. Novel cosmeceuticals from plants—An industry guided review. J. Appl. Res. Med. Aromat. Plants 2017, 7, 1–26. [Google Scholar] [CrossRef]
- SÁ, A.G.A.; Meneses, A.C.d.; Araújo, P.H.H.d.; Oliveira, D.d. A review on enzymatic synthesis of aromatic esters used as flavor ingredients for food, cosmetics and pharmaceuticals industries. Trends Food Sci. Technol. 2017, 69, 95–105. [Google Scholar] [CrossRef]
- Kumar, D.; Punetha, A.; Verma, P.P.S.; Padalia, R.C. Micronutrient based approach to increase yield and quality of essential oil in aromatic crops. J. Appl. Res. Med. Aromat. Plants 2022, 26, 100361. [Google Scholar] [CrossRef]
- Singh, P.A.; Bajwa, N.; Chinnam, S.; Chandan, A.; Baldi, A. An overview of some important deliberations to promote medicinal plants cultivation. J. Appl. Res. Med. Aromat. Plants 2022, 31, 100400. [Google Scholar] [CrossRef]
- Mo, L.; Bin, W.; Jiang, L.; Jiang, Y. The current developing state of chemical processes and utility industry of Cinnamomum camphora resources. Biomass Chem. Eng. (Chin.) 2021, 55, 15–22. [Google Scholar] [CrossRef]
- Gaur, V.K.; Sharma, P.; Sirohi, R.; Awasthi, M.K.; Dussap, C.-G.; Pandey, A. Assessing the impact of industrial waste on environment and mitigation strategies: A comprehensive review. J. Hazard. Mater. 2020, 398, 123019. [Google Scholar] [CrossRef]
- Agathokleous, E.; Barceló, D.; Rinklebe, J.; Sonne, C.; Calabrese, E.J.; Koike, T. Hormesis induced by silver iodide, hydrocarbons, microplastics, pesticides, and pharmaceuticals: Implications for agroforestry ecosystems health. Sci. Total Environ. 2022, 820, 153116. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, N.; Lin, Y.; Zhan, Y.; Ding, X.; Liu, Y.; Zhang, A.; Ding, G.; Xu, T.; Li, J. Recycling of nutrients from organic waste by advanced compost technology-A case study. Bioresour. Technol. 2021, 337, 125411. [Google Scholar] [CrossRef]
- Malherbe, S.; Cloete, T.E. Lignocellulose biodegradation: Fundamentals and applications. Rev. Environ. Sci. Biotechnol. 2002, 1, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Che, J.; Bai, Y.; Li, X.; Ye, J.; Liao, H.; Cui, P.; Yu, Z.; Zhou, S. Linking microbial community structure with molecular composition of dissolved organic matter during an industrial-scale composting. J. Hazard. Mater. 2021, 405, 124281. [Google Scholar] [CrossRef]
- Chen, Z.; Fu, Q.; Wen, Q.; Wu, Y.; Bao, H.; Guo, J. Microbial community competition rather than high-temperature predominates ARGs elimination in swine manure composting. J. Hazard. Mater. 2022, 423, 127149. [Google Scholar] [CrossRef]
- Guo, H.-N.; Liu, H.-T.; Wu, S. Immobilization pathways of heavy metals in composting: Interactions of microbial community and functional gene under varying C/N ratios and bulking agents. J. Hazard. Mater. 2022, 426, 128103. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, H.; Zhang, H.; Xun, L.; Chen, G.; Wang, L. Thermomyces lanuginosus is the dominant fungus in maize straw composts. Bioresour. Technol. 2015, 197, 266–275. [Google Scholar] [CrossRef]
- Perner, H.; Schwarz, D.; Bruns, C.; Mäder, P.; George, E. Effect of arbuscular mycorrhizal colonization and two levels of compost supply on nutrient uptake and flowering of pelargonium plants. Mycorrhiza 2007, 17, 469–474. [Google Scholar] [CrossRef]
- Huang, D.; Gao, L.; Cheng, M.; Yan, M.; Zhang, G.; Chen, S.; Du, L.; Wang, G.; Li, R.; Tao, J.; et al. Carbon and N conservation during composting: A review. Sci. Total Environ. 2022, 840, 156355. [Google Scholar] [CrossRef]
- Elisashvili, V.I.; Khardziani, T.S.; Tsiklauri, N.D.; Kachlishvili, E.T. Cellulase and xylanase activities in higher basidiomycetes. Biochemistry. Biokhimiia 1999, 64, 718–722. [Google Scholar]
- Tuor, U.; Winterhalter, K.; Fiechter, A. Enzymes of white-rot fungi involved in lignin degradation and ecological determinants for wood decay. J. Biotechnol. 1995, 41, 1–17. [Google Scholar] [CrossRef]
- Zeng, Z.; Guo, X.; Xu, P.; Xiao, R.; Huang, D.; Gong, X.; Cheng, M.; Yi, H.; Li, T.; Zeng, G. Responses of microbial carbon metabolism and function diversity induced by complex fungal enzymes in lignocellulosic waste composting. Sci. Total Environ. 2018, 643, 539–547. [Google Scholar] [CrossRef]
- Guan, Z.-B.; Song, C.-M.; Zhang, N.; Zhou, W.; Xu, C.-W.; Zhou, L.-X.; Zhao, H.; Cai, Y.-J.; Liao, X.-R. Overexpression, characterization, and dye-decolorizing ability of a thermostable, pH-stable, and organic solvent-tolerant laccase from Bacillus pumilus W3. J. Mol. Catal. B Enzym. 2014, 101, 1–6. [Google Scholar] [CrossRef]
- Li, X.; Zheng, Y. Lignin-enzyme interaction: Mechanism, mitigation approach, modeling, and research prospects. Biotechnol. Adv. 2017, 35, 466–489. [Google Scholar] [CrossRef]
- Sommers, L.E.; Nelson, D.W. Determination of Total Phosphorus in Soils: A Rapid Perchloric Acid Digestion Procedure. Soil Sci. Soc. Am. J. 1972, 36, 902–904. [Google Scholar] [CrossRef]
- Bradbury, I.K.; Malcolm, D. The effect of phosphorus and potassium on transpiration, leaf diffusive resistance and water-use efficiency in Sitka spruce (Picea sitchensis) seedlings. J. Appl. Ecol. 1977, 14, 631–641. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef]
- Zhou, H.; Ma, A.; Zhou, X.; Chen, X.; Zhang, J.; Zhang, Q.; Qi, X.; Liu, G.; Zhuang, G. Phosphorus Shapes Soil Microbial Community Composition and Network Properties During Grassland Expansion Into Shrubs in Tibetan Dry Valleys. Front. Plant Sci. 2022, 13, 848691. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Kai, F.; Zhang, Z.; Cai, W.; Liu, W.; Xu, M.; Yin, H.; Wang, A.; He, Z.; Ye, D. Biodiversity and species competition regulate the resilience of microbial biofilm community. Mol. Ecol. 2017, 26, 6170–6182. [Google Scholar]
- Deng, Y.; Zhou, J. Molecular Ecological Network of Microbial Communities. In Encyclopedia of Metagenomics: Genes, Genomes and Metagenomes: Basics, Methods, Databases and Tools; Nelson, K.E., Ed.; Springer: Boston, MA, USA, 2015; pp. 504–510. [Google Scholar]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911. [Google Scholar] [CrossRef]
- Kline, R.; Kline, R.B.; Kline, R. Principles and Practice of Structural Equation Modeling. J. Am. Stat. Assoc. 2011, 101. [Google Scholar]
- Xiao, B.; Sun, X.F.; Sun, R. Chemical, structural, and thermal characterizations of alkali-soluble lignins and hemicelluloses, and cellulose from maize stems, rye straw, and rice straw. Polym. Degrad. Stab. 2001, 74, 307–319. [Google Scholar] [CrossRef]
- Vidal-Beaudet, L.; Grosbellet, C.; Forget-Caubel, V.; Charpentier, S. Modelling long-term carbon dynamics in soils reconstituted with large quantities of organic matter. Eur. J. Soil Sci. 2012, 63, 787–797. [Google Scholar] [CrossRef]
- Liu, H.; Sun, J.; Leu, S.-Y.; Chen, S. Toward a fundamental understanding of cellulase-lignin interactions in the whole slurry enzymatic saccharification process. Biofuels Bioprod. Biorefining 2016, 10, 648–663. [Google Scholar] [CrossRef]
- Rodríguez Couto, S.; Toca Herrera, J.L. Industrial and biotechnological applications of laccases: A review. Biotechnol. Adv. 2006, 24, 500–513. [Google Scholar] [CrossRef]
- Scheel, T.; Höfer, M.; Ludwig, S.; Hölker, U. Differential expression of manganese peroxidase and laccase in white-rot fungi in the presence of manganese or aromatic compounds. Appl. Microbiol. Biotechnol. 2000, 54, 686–691. [Google Scholar] [CrossRef]
- Yu, Z.; Gwak, K.-S.; Treasure, T.; Jameel, H.; Chang, H.-M.; Park, S. Effect of Lignin Chemistry on the Enzymatic Hydrolysis of Woody Biomass. ChemSusChem 2014, 7, 1942–1950. [Google Scholar] [CrossRef]
- Egea, T.C.; Rodrigues, A.; da Silva, R.; Gomes, E. Production of xylanase, cellulase and laccase by mesophilic fungi isolated from sugar cane soil. J. Biotechnol. 2010, 150, 419. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, R.; Li, Z.; Li, J.; Fang, Z.; Fang, W.; Zhang, X.; XIiao, Y. Semi-rational engineering of microbial laccase Lac15 for enhanced activity. Acta Microbiol. Sin. 2022, 62, 1501–1512. [Google Scholar]
- Han, D.-J.; Wang, Z.-H.; Zhou, N.; Liu, G.; Yang, S.; Wang, W. Screening and Degradation Effect of Lignin-degrading Bacteria in Termite Nurseries. Biotechnol. Bull. 2022, 3, 113–120. [Google Scholar]
- Zhang, Y.; Ren, J.; Zhang, W.; Wang, S.; Wang, S.; Ma, F.; Li, S.; Li, Y. Screening, Identification and Lignin-degradation Characteristics of Bacillus amyloliquefaciens MN-13. J. Agric. Biotechnol. 2021, 29, 1389–1399. [Google Scholar]
- Silva, M.E.F.; Lopes, A.R.; Cunha-Queda, A.C.; Nunes, O.C. Comparison of the bacterial composition of two commercial composts with different physicochemical, stability and maturity properties. Waste Manag. 2016, 50, 20–30. [Google Scholar] [CrossRef]
- Tang, J.-C.; Maie, N.; Tada, Y.; Katayama, A. Characterization of the maturing process of cattle manure compost. Process Biochem. 2006, 41, 380–389. [Google Scholar] [CrossRef]
- Duan, H.; Ji, M.; Xie, Y.; Shi, J.; Liu, L.; Zhang, B.; Sun, J. Exploring the Microbial Dynamics of Organic Matter Degradation and Humification during Co-Composting of Cow Manure and Bedding Material Waste. Sustainability 2021, 13, 13035. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Duan, Y.; Awasthi, S.K.; Liu, T.; Zhang, Z.; Kim, S.-H.; Pandey, A. Effect of biochar on emission, maturity and bacterial dynamics during sheep manure compositing. Renew. Energy 2020, 152, 421–429. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Chen, H.; Wang, Q.; Liu, T.; Duan, Y.; Awasthi, S.K.; Ren, X.; Tu, Z.; Li, J.; Zhao, J.; et al. Succession of bacteria diversity in the poultry manure composted mixed with clay: Studies upon its dynamics and associations with physicochemical and gaseous parameters. Bioresour. Technol. 2018, 267, 618–625. [Google Scholar] [CrossRef]
- Franke-Whittle, I.H.; Knapp, B.A.; Fuchs, J.; Kaufmann, R.; Insam, H. Application of COMPOCHIP Microarray to Investigate the Bacterial Communities of Different Composts. Microb. Ecol. 2009, 57, 510–521. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Li, W.; Zhang, S.; Wu, C.; Lv, L. Feasibility of co-composting of sewage sludge, spent mushroom substrate and wheat straw. Bioresour. Technol. 2017, 226, 39–45. [Google Scholar] [CrossRef]
- Kasuga, T.; White, T.J.; Taylor, J.W. Estimation of Nucleotide Substitution Rates in Eurotiomycete Fungi. Mol. Biol. Evol. 2002, 19, 2318–2324. [Google Scholar] [CrossRef]
- Wang, X.; Kong, Z.; Wang, Y.; Wang, M.; Liu, D.; Shen, Q. Insights into the functionality of fungal community during the large scale aerobic co-composting process of swine manure and rice straw. J. Environ. Manag. 2020, 270, 110958. [Google Scholar] [CrossRef]
- Meng, X.; Yan, J.; Zuo, B.; Wang, Y.; Yuan, X.; Cui, Z. Full-scale of composting process of biogas residues from corn stover anaerobic digestion: Physical-chemical, biology parameters and maturity indexes during whole process. Bioresour. Technol. 2020, 302, 122742. [Google Scholar] [CrossRef]
- Whitehead, T.R.; Johnson, C.N.; Patel, N.B.; Cotta, M.A.; Moore, E.R.B.; Lawson, P.A. Savagea faecisuis gen. nov., sp. nov., a tylosin- and tetracycline-resistant bacterium isolated from a swine-manure storage pit. Antonie Van Leeuwenhoek 2015, 108, 151–161. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OTU ID | Genus | Xylanase | Cellulose | Hemicellulose | Laccase | Lignin | Starch | Soluble Sugar | Protein |
|---|---|---|---|---|---|---|---|---|---|
| BOTU1023 | Lactobacillus | − | + | ||||||
| BOTU1051 | Corynebacterium | − | |||||||
| BOTU1114 | Brevibacterium | + | |||||||
| BOTU1137 | Aerosphaera | − | − | ||||||
| BOTU1145 | Enterococcus | + | |||||||
| BOTU1181 | Carnobacteriaceae | + | |||||||
| BOTU1236 | Limnochordaceae | − | |||||||
| BOTU1249 | Limnochordaceae | + | − | − | |||||
| BOTU1570 | Bacillaceae | − | |||||||
| BOTU1610 | Thermobifida | + | − | − | |||||
| BOTU1613 | Ammoniphilus | + | − | − | |||||
| BOTU1642 | Bacillaceae | + | − | − | |||||
| BOTU1663 | Paenibacillaceae | + | |||||||
| BOTU1715 | Limnochordaceae | + | − | − | |||||
| BOTU1769 | Limnochordaceae | + | − | − | |||||
| BOTU1790 | Novibacillus | + | |||||||
| BOTU890 | Jeotgalibaca | − | |||||||
| BOTU898 | Brevibacterium | − | |||||||
| BOTU924 | Atopostipes | − | + | + | + | ||||
| BOTU932 | Sporosarcina | − | + | + | |||||
| BOTU959 | Staphylococcus | − | |||||||
| BOTU963 | Facklamia | − | |||||||
| BOTU965 | Corynebacterium | − | |||||||
| BOTU967 | Pseudogracilibacillus | − | |||||||
| BOTU972 | Gallicola | − | + | ||||||
| BOTU974 | Gallicola | − | + | + | |||||
| BOTU979 | Savagea | − | + | + | |||||
| BOTU991 | Savagea | − | + | + | − | + | |||
| FOTU400 | Aspergillus | − | + | + | |||||
| FOTU425 | Scopulariopsis | + | |||||||
| FOTU511 | Pestalotiopsis | + | |||||||
| FOTU521 | Aspergillus | − | + | + | |||||
| FOTU727 | Penicillium | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Di, L.; Hua, X.; Deng, T.; Wang, X. Bacterial Community Drives the Carbon Source Degradation during the Composting of Cinnamomum camphora Leaf Industrial Extracted Residues. Microbiol. Res. 2023, 14, 229-242. https://doi.org/10.3390/microbiolres14010019
Zhou H, Di L, Hua X, Deng T, Wang X. Bacterial Community Drives the Carbon Source Degradation during the Composting of Cinnamomum camphora Leaf Industrial Extracted Residues. Microbiology Research. 2023; 14(1):229-242. https://doi.org/10.3390/microbiolres14010019
Chicago/Turabian StyleZhou, Hanchang, Lan Di, Xiaoju Hua, Tao Deng, and Xiaodong Wang. 2023. "Bacterial Community Drives the Carbon Source Degradation during the Composting of Cinnamomum camphora Leaf Industrial Extracted Residues" Microbiology Research 14, no. 1: 229-242. https://doi.org/10.3390/microbiolres14010019