Procalcitonin and Adrenomedullin in Infectious Diseases


1
Kagawa School of Pharmaceutical Sciences, Tokushima Bunri University, Shido 1314-1, Sanuki 769-2193, Japan
2
Institute of Neuroscience, Tokushima Bunri University, Shido 1314-1, Sanuki 769-2193, Japan
*
Author to whom correspondence should be addressed.
Microbiol. Res. 2023, 14(1), 190-204; https://doi.org/10.3390/microbiolres14010016
Submission received: 29 December 2022
/
Revised: 27 January 2023
/
Accepted: 30 January 2023
/
Published: 1 February 2023
{kind=link}
{kind=link}
Abstract
:Calcitonin (CT) and adrenomedullin (ADM) are members of the CT family. Procalcitonin (PCT) is a prohormone of CT. Elevations in serum PCT and ADM levels are associated with severe sepsis and coronavirus disease 2019 (COVID-19). PCT enhances sepsis mortality and it binds to the CGRP receptor, which is a heterodimer of CT receptor-like receptor and receptor activity-modifying protein 1. The N-terminal truncated form of PCT, PCT3-116, is produced by the cleavage of PCT by dipeptidyl peptidase 4 (DPP-4) and is the main form of PCT in serum during sepsis, inducing microvascular permeability. Mid-regional pro-adrenomedullin (MR-proADM) is used instead of ADM as a biological indicator because ADM is rapidly degraded, and MR-proADM is released at the same rate as ADM. ADM reduces endothelial permeability and promotes endothelial stability. Endothelial dysfunction is responsible for multiple organ failure in sepsis and COVID-19 patients. Therefore, ADM may be an important molecule for improving the severity associated with sepsis and COVID-19. This review focuses on the current knowledge of PCT and ADM in sepsis and COVID-19.
1. Introduction
Calcitonin (CT) and adrenomedullin (ADM) are members of the CT family, which includes α-calcitonin gene-related peptide (αCGRP), βCGRP, and intermedin, also known as ADM2. Procalcitonin (PCT) is a prohormone of CT, and the elevation of PCT in the blood is associated with bacterial sepsis [1,2]. Pro-adrenomedullin (proADM) is a prohormone of ADM and contains proadrenomedullin N-terminal 20 peptide (PAMP), mid-regional pro-adrenomedullin (MR-proADM), ADM, and adrenotensin. The elevation of ADM in the blood is associated with hypertension [3], heart failure [4], chronic inflammatory lung diseases [5,6] and septic shock [7]. However, it is difficult to measure the expression level of ADM because of its rapid degradation [8]. In contrast, MR-proADM has a longer half-life than ADM. In addition, MR-proADM is released from the cleavage of proADM at the same rate as ADM. Therefore, measuring the expression level of MR-proADM can be used to estimate the expression level of ADM [9]. However, measuring MR-proADM itself is unable to distinguish between the biologically active ADM, which is amidated at N-terminus, and the biologically inactive ADM, which has a glycine-extended C-terminus. Therefore, a method to measure bioactive ADM (bio-ADM) was developed [10]. Both PCT and MD-proADM have been used as biomarkers of sepsis [11,12,13], with suppression of PCT [14,15,16] and administration of ADM both reducing the severity of sepsis [17]. Therefore, suppression of PCT and enhancement of ADM may reduce the severity of sepsis and other infectious diseases. Although various functions of ADM, such as suppression of vascular permeability, have been reported [18], the detailed physiological role of PCT has remained unclear. However, it has recently been reported that PCT increases cell permeability [19]. Therefore, the role of PCT and ADM in vascular permeability in sepsis is presumed to be important in the severity of infection. In addition, recent reports have shown the association between COVID-19 and PCT or ADM [20,21]. This review will focus on the mechanisms by which sepsis and other infections induce the expression of PCT and ADM and their roles in infectious disease with current knowledge of the physiological functions of PCT and ADM.
2. Procalcitonin and Adrenomedullin
2.1. Procalcitonin
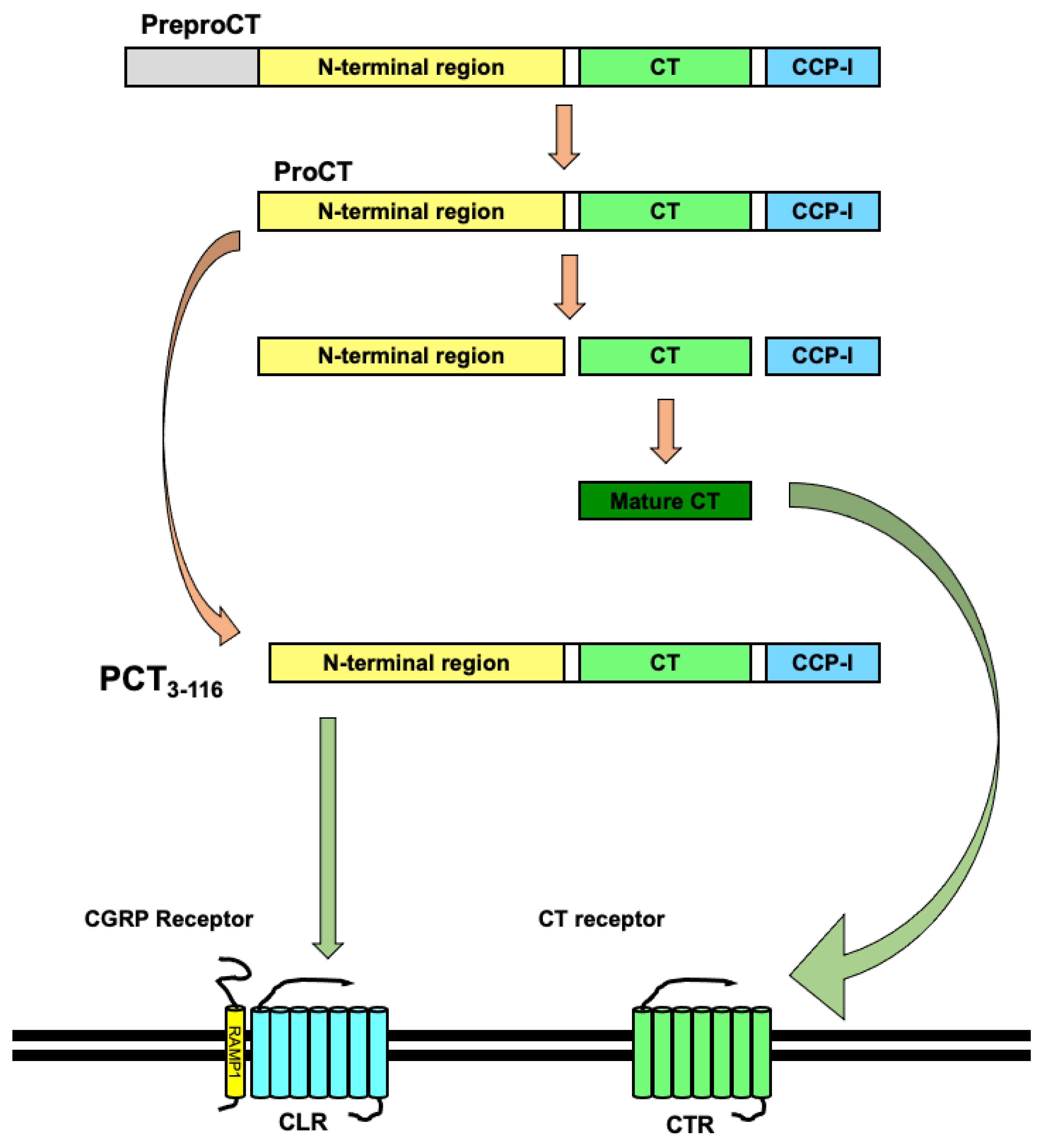
CT is a peptide hormone composed of 32 amino acids with a disulfide bridge in the N-terminus (Cys1 and Cys7) and amidation at the C-terminus. CT is mainly secreted from the thyroid gland’s parafollicular cells, which are also called C cells. CT is mainly considered to control serum calcium levels by inhibiting both the bone-resorbing activity of osteoclasts resorption and the renal tubular reabsorption of calcium in the kidney [22]. The human CT gene, CALCA, is located in chromosome 11 [23,24] and has two alternative splicing variants: CT and CGRP [25]. Exons I to IV of CALCA encode the 141-amino acid-long preproCT [26]. Exons I to III and V of CALCA encode the 128-amino acid-long preproCGRP [27]. After preproCT is cleaved and the signal peptide of 25 amino acids is detached, the 116-amino acid-long PCT is generated. The cleavage of PCT then produces aminoproCT, immature CT, and katacalcin, which is also known as CCP-I (calcitonin carboxypeptide-1). Then, the immature CT is converted into a mature CT through post-transcriptional modifications, namely a disulfide bridge between Cys1 and Cys7 and amidation of the C-terminus (Figure 1) [28,29,30].
CT receptor (CTR) and CTR-like receptor (CLR) are seven transmembrane G-protein-coupled receptors (GPCRs) [22,31]. These receptors associate with receptor activity-modifying proteins (RAMPs) and act as receptors for members of the CT family. RAMPs can associate with various GPCRs and modify GPCR signaling [32]. The receptor for CT is CTR. The receptor for CGRP is the combination of CLR and RAMP1. The receptor for ADM is the combination of CLR and RAMP2 or RAMP3. The receptor for amylin is the combination of CTR and RAMP1 or RAMP2, or RAMP3 [33,34,35,36]. Although CTR is the receptor for CT, PCT activates the CGRP receptor, which is a combination of CLR and RAMP1 [37]. PCT and its mRNA are increased in the response to bacterial sepsis and pyrogen in various human and rodent organs [1,2,38,39,40,41,42]. PCT is distributed in multiple organs, such as the brain, heart, lungs, liver, and kidney, and in the blood vessels of these organs in cases of patients who died of septic shock [40]. In addition, PCT levels were high in the liver, kidney, aorta, fat, ovaries, bladder and adrenal gland in lipopolysaccharide (LPS)-treated baboons [41] and postmortem PCT levels in the brain, kidney, and liver were high in LPS-treated rabbits [42]. In addition, CT-mRNA expression was also elevated in multiple organs, such as the brain, pituitary, lungs, heart, liver, and kidney, of the sepsis model animals [2,38,41]. PCT is a ligand for the CGRP receptor, formed from a heterodimer of CLR and RAMP1 [34]. The distribution of CLR in humans is similar to the distribution of PCT in septic shock. CLR is also expressed in multiple organs, such as the blood vessels, heart, and kidney. Although CLR was not detected over the epithelium of bronchi and bronchioles or bronchial glands, it was detected in pulmonary vessels [43]. PCT is produced in multiple organs and is able to enter the circulation. Therefore, it is understood that PCT can affect not only the producing organ itself, but also distal organs, by being released into the bloodstream from that organ. In addition, because both PCT and CLR are expressed in blood vessels, PCT may be involved in systemic inflammatory reaction syndrome (SIRS), by affecting the permeability of blood vessels throughout the body [44,45]. Under normal conditions, vascular permeability remains low. However, under conditions of induced inflammation, inflammatory mediators such as cytokines increase vascular permeability, causing leakage of plasma components and extravasation of leukocytes. However, excessive production of inflammatory mediators further increases vascular permeability and aggravates the pathogenesis [46].
Processing of PCT1-116 is catalyzed by dipeptidyl-peptidase 4 (DPP-4), which cleaves the N-terminal dipeptide Ala1-Pro2 of PCT1-116, resulting in the generation of a 114-amino acid-long, truncated form (PCT3-116) [47]. In addition, PCT3-116 is considered the main form of PCT in the serum during sepsis [48,49]. At the endothelial cell surface, the DPP-4-cleaved PCT3-116 activates the CGRP receptor (CLR heterodimerized with RAMP1) and induces microvascular permeability through the phosphorylation of vascular endothelial (VE)-cadherin [19]. In addition, increased vascular permeability during inflammatory responses is brought about by phosphorylation of VE-cadherin [50]. Furthermore, lipopolysaccharide induces VE-cadherin disruption [51,52]. VE-cadherin is the major adhesion molecule and constitutes vascular endothelial cell–cell adhesion. Vascular permeability is regulated by the formation of endothelial cell–cell adhesions. Thus, PCT can play an important role in the regulation of systemic vascular permeability via VE-cadherin.
2.2. Adrenomedullin
ADM is a peptide hormone composed of 52 amino acids that displays a disulfide bridge between Cys16 and Cys21 and amidation at the C-terminus. Although ADM is mainly expressed in the adrenal medulla, it is also found in a variety of organs, such as blood vessels, the heart, lungs, kidneys, and so on [53,54]. ADM has diverse physiological functions: hemodynamic improvement, organ protection, anti-inflammatory effects, tissue repair, and regeneration [18]. The human ADM gene, ADM, is located in chromosome 11 [55]. ADM is initially generated as preproADM, which has 185 amino acids. After cleavage of the 21-amino acid-long signal peptide from preproADM, a proADM of 164 amino acids is generated. Proadrenomedullin N-terminal 20 peptide (PAMP), of 20 amino acids, the 48-amino acid MR-proADM, the 52-amino acid ADM, and the 33-amino acid adrenotensin are all released by the cleavage of proADM [56]. Immature ADM becomes a mature ADM by post-transcriptional modification (disulfide bridge between Cys16 and Cys21 and amidation of C-terminus) [53] (Figure 2). No physiological function for MR-proADM has been found to date. In addition, alternative RNA processing of the ADM gene generates another mRNA coding PAMP and a truncated MR-proADM. Thus, the expression levels of PAMP and ADM are not parallel [57]. Circulating ADM is elevated by hypertension [3], heart failure [4], chronic inflammatory lung diseases [5,6] and septic shock [7]. In addition, ADM reduces endothelial permeability and promotes endothelial stability [58,59].
PAMP-12 or PAMP (9-20) is a 12-amino acid peptide is the processed form of PAMP. It activates chemokine receptors, namely the Mas-related G-protein coupled receptor member X2 (MRGPRX2 in humans and Mrgx2 in mice) and atypical chemokine receptor 3 (ACKR3), also known as the C-X-C chemokine receptor type 7 [60]. In addition, PAMP-12 has hypotensive action [61] and antimicrobial activity [62].
3. Infectious Diseases, Procalcitonin and Adrenomedullin
3.1. Procalcitonin and Adrenomedullin in Sepsis
3.1.1. Sepsis
The current definition of sepsis (Sepsis-3) is life-threatening organ dysfunction caused by a dysregulated host response to infection [63,64]. Organ dysfunction is considered an acute change with a sequential (sepsis-related) organ failure assessment (SOFA) score ≥ 2 points, which is associated with an in-hospital mortality greater than 10% [63,65]. Septic shock, a subtype of sepsis, is a severe form of sepsis affecting the circulatory system and producing cellular and metabolic abnormalities, with a higher risk of death than sepsis. Furthermore, the associated hypotension requires a vasopressor to maintain a mean arterial pressure ≥ 65 mm Hg and serum lactate > 2 mmol/L (>18 mg/dL) without hypovolemia [63]. International guidelines for managing sepsis and septic shock have been provided [66].
When tissues are stimulated or damaged by pathogens, such as bacteria and fungi, tissue abnormalities, such as circulatory disturbance, tissue infiltration, and tissue degeneration, occur. When the presence of these pathogens or pathological changes in tissues are sensed, inflammatory responses occur to eliminate the cause of the pathogens and to repair the abnormalities. The characteristic components of the pathogen or damaged tissues are recognized as molecular patterns by pattern recognition receptors (PRRs) [67,68,69]. Molecular patterns include pathogen-associated molecular pattern molecules (PAMPs), which are derived from infected pathogens, and damage-associated molecular pattern molecules (DAMPs), are derived from damaged cells. There are five classes of PRRs: toll-like receptors (TLRs), nucleotide-binding oligomerization domain-like receptors, RIG-I-like receptors (RLRs), C-type lectin receptors, and absent in melanoma-2-like receptors [70]. Stimulation of PRRs by pattern recognition molecules activates the intracellular signaling pathways of PRRs and induces cytokine production and secretion. As a result, inflammatory and infection defense responses are induced in immune cells and tissue components that express PRRs [68,71]. A variety of pro-inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interleukin-1 (IL-1), IL-6, IL-8, and IL-12, are elevated during the early stages of sepsis [64,72].
3.1.2. Procalcitonin in Sepsis
Serum PCT are increased by responding to bacterial sepsis and bacterial endotoxins and are produced in various organs in humans and rodents [1,2,38,39]. CT mRNA is also elevated in multiple organs by sepsis and endotoxins [1,2,38,39]. Moreover, procalcitonin increases sepsis mortality in rodents, and antibodies against PCT improve sepsis in pigs and rodents [14,15,16,73]. PCT is also induced by proinflammatory cytokines, such as IL-1, IL-2, IL-6, or TNF-α, which are induced by bacteria or their endotoxins [12,74]. In addition, the binding site for the transcription factor nuclear factor kappa B (NF-κB) is identified in the promoter region of the calcitonin gene and LPS induces the expression of PCT via the activation of NF-κB [75]. NF-κB is a member of the inducible transcription factor family and a crucial master transcription factor that regulates a variety of molecules for immune and inflammatory responses. The activation of NF-κB leads to the regulation of many genes involved in various processes of inflammatory responses, including the expression of proinflammatory cytokines. In addition, proinflammatory cytokines such as IL-1, IL-2, IL-6, and TNF-α, which induce PCT expression, also stimulate the activation of NF-κB [76].
PCT levels are higher in Gram-negative bacteremia than in Gram-positive bacteremia and fungemia [77,78,79,80]. The different PCT values induced by Gram-negative bacteria, Gram-positive bacteria, and fungi are considered to be due to the fact that each of them stimulate different receptors. The main membrane component of Gram-negative bacteria is LPS and that of Gram-positive bacteria is peptidoglycan (PGN) and lipoteichoic acid (LTA). LPS mainly activates TLR4. In contrast, PGN and LTA mainly activate TLR2. In addition, fungi mainly activate the C-type lectin receptor. However, the detailed mechanisms by which PCT values differ among infections caused by different pathogens have not been elucidated [80]. In contrast, it has been considered that the elevation of PCT is not due to viral infection. Interferons (IFNs) are significantly released by the infection of viruses [81,82,83]. IFNγ suppresses the expression of both PCT and its mRNA induced by IL-1 [84]. Thus, PCT has been considered as a diagnostic marker of sepsis caused by bacteria, not viruses. Therefore, it should be possible to monitor the usage of antibiotics because the PCT level is thought to indicate the level of bacterial infection [11,12].
However, recent studies have shown that PCT does not distinguish between bacterial and viral infection [85,86]. For example, PCT levels in patients with community-acquired pneumonia (CAP) do not reflect the expected difference between bacterial and viral pneumonia [86]. In addition, PCT levels are elevated by viral infection, organ failure, and mortality. Organ failure and mortality caused by influenza infection induce PCT without bacterial coinfection [85]. Furthermore, an association between PCT and coronavirus disease 2019 (COVID-19) has been reported [87,88,89,90,91,92,93,94].
PCT has been expected to be a diagnostic marker that indicates the likelihood of sepsis and septic shock. Early diagnosis of sepsis is one of the effective factors associated with poor prognosis of sepsis. Diagnostic markers provide information not only on the likelihood of disease, but also on the subsidence of the disease, improving the patient’s clinical evaluation. Monitoring PCT values is believed to aid in the appropriate use of antibiotics in clinical practice and in improving disease status. Therefore, efficient diagnosis of sepsis prevents misuse and overuse of antibiotics in clinical practice [95,96,97]. Various methods of measuring PCT levels have been developed for diagnosis, such as the enzyme-linked fluorescent immunoassay, chemiluminescence immunoassay, chemiluminescent microparticle immunoassay, time-resolved amplified cryptate emission immunoassay, electrochemiluminescence immunoassay, latex-enhanced immunoturbidimetric assay, and the lateral flow immunoassay, also known as immunochromatography assay or rapid antigen test [98,99,100]. In addition, the use of immunohistochemical assays for PCT in specific organs, such as the brain, heart, lungs, liver, and kidney, is useful for the postmortem diagnosis of sepsis [44]. Postmortem diagnosis leads to the investigation of the causes of the process of death and the prevention and treatment of that disease.
3.1.3. Adrenomedullin in Sepsis
Serum ADM levels are increased in sepsis and are associated with the severity of sepsis, organ failure, and mortality [101,102,103,104]. Furthermore, the expression of ADM is upregulated by LPS and proinflammatory cytokines, such as IL-1 and TNF [105]. In addition, ADM is also elevated by hypoxia (hypoxic responses) [106]. In sepsis, blood flow is impeded due to impaired circulatory function caused by vascular microthrombosis. As a result, oxygen supply to various organs is reduced, leading to hypoxia and multiorgan failure syndrome [107]. Many biological responses to hypoxia are regulated by the transcription factors hypoxia-inducible factors (HIFs), key regulators of hypoxia. HIFs regulate genes involved in a variety of biological events, including hematopoiesis, angiogenesis, inflammation, apoptosis, and autophagy. HIFs also regulate genes related to intracellular energy metabolism [108,109]. However, HIFs are negatively regulated by prolyl hydroxylase domain proteins (PHDs). Therefore, PHDs act as an oxygen concentration sensor molecule during hypoxia and serves as a master regulator of the response to hypoxia [110]. HIFs are transcription factors formed by a heterodimeric complex composed of two subunits, HIFα and HIF1β, which are also known as aryl hydrocarbon receptor nuclear translocator (Arnt). A heterodimeric complex of HIFα and HIF1β is responsible for the transcription of many hypoxia-related genes. The β-subunit of HIF (HIF1β) is always expressed, but the expression of the α-subunit (HIFα) changes with oxygen concentration. Under normal oxygen concentrations, specific proline residues in the N-terminal transcription activation domain (NTAD) of HIFα are hydroxylated by PHDs, which, in turn, is used as an indicator that HIFα is ubiquitinated by a ubiquitin ligase complex containing the protein product of the von Hippel-Lindau (pVHL) and degraded by the proteasome. Thus, HIFs are not able to form a heterodimer consisting of HIFα and HIF1β, leading to the suppression of the HIF-mediated hypoxic responses. In contrast, hypoxia suppresses the activity of PHD, which requires molecular oxygen for enzymatic activity, thus inhibiting the hydroxylation of HIFα by PHD. Therefore, HIFα is protected from proteasomal proteolysis by ubiquitination by the pVHL complex and accumulates intracellularly. This allows HIFα to bind to HIF1β/Arnt and form heterodimers. HIF promotes transcription of a group of hypoxia-related genes by binding to specific hypoxia response elements (HREs) on genomic DNA, thereby activating a HIF-mediated hypoxia response, including the elevation of ADM [106,108].
ADM suppresses endothelial permeability and promotes endothelial stability, leading to the reduction of hypoxia and leakage of plasma components and extravasation of leukocytes [58,59]. In addition, administration of ADM decreases mortality in a rat sepsis model [17]. It is considered that ADM may reduce sepsis-related mortality by inhibiting vascular permeability. ADM has two receptors: the AM1 receptor (CLR and RAMP2) and the AM2 receptor (CLR and RAMP3). It has been reported that AM1 receptors are involved in angiogenesis and vascular homeostasis, while AM2 receptors are involved in lymphatic vessel function [111,112]. ADM induces intracellular cAMP [113], leads to the activation of an exchange protein directly activated by cAMP (Epac1) [114]. Epac1 is a guanine nucleotide exchange factor for the small GTPase Rap1 [115]. Activation of Epac1 and Rap1 is responsible for the maintenance of endothelial barrier function [114,116]. In addition, Epac1 has been reported to regulate cell junction formation via VE-cadherin [117].
Although serum ADM levels are related to the severity of sepsis [101,102,103,104], it is difficult to measure serum ADM because the circulating ADM has a short half-life [8] and the ADM binding protein interferes with the measurement of ADM [118]. Thus, because MR-proADM is stable in the plasma [119], its levels are measured to indicate the severity of sepsis [9,13,120,121]. In addition, MR-proADM levels are associated with organ failure [122]. Administration of ADM reduces mortality in a rat sepsis model [17]. The N-terminus of ADM may not affect ADM receptors because ADM22–52 binds to the AM receptors [123]. Thus, Adrecizumab, an antibody that targets the N-terminus of ADM, increases circulating ADM, which leads to an increase in the activity of ADM [124].
3.2. Procalcitonin and Adrenomedullin in COVID-19
3.2.1. COVID-19
COVID-19, also called the novel coronavirus disease, is an infectious disease caused by the SARS-CoV-2 virus, also called the novel coronavirus. SARS-CoV-2 is related to SARS-CoV-1, which caused SARS in 2003, and to the Middle Eastern respiratory syndrome coronavirus (MERS-CoV), which caused MERS in 2012 [125]. COVID-19 is characterized by acute respiratory symptoms (sore throat, cough, and shortness of breath), fever, diarrhea, vomiting, loss of smell or taste, chills, muscle fatigue, and multiple organ failure [126,127,128]. COVID-19 causes severe viral pneumonia, which, in the most complicated cases, can lead to death [129]. SARS-CoV-2 has a genomic RNA of approximately 30,000 bases and produces proteins necessary for the replication and infection of SARS-CoV-2. The spike protein and envelope proteins are structural proteins that form viral particles. In addition to structural proteins, viral genomic RNA codes for nonstructural proteins (NSPs) crucial for replication and other processes, such as proteases and RNA-dependent RNA polymerase (RdRP). After SARS-CoV-2 enters the cell, it synthesizes SARS-CoV-2 proteins based on its viral genomic RNA. Cleavage of this SARS-CoV-2 protein results in the production of NSPs. NSP5 is a 3C-like protease (3CLpro), which is also called main protease (Mpro), and NSP3 is a papain-like protease (PLpro). 3CLpro and PLpro cleave 11 and 3 sites in the SARS-CoV-2 protein, respectively. 3CLpro is activated by dimerization to cleave the SARS-CoV-2 protein and release NSPs, including RdRP (NSP12), NSP 7, and NSP8 [130]. The complex of RdRP, NSP 7, and NSP8 is responsible for the replication of SARS-CoV-2 genomic RNA [131]. RdRP is an RNA-dependent RPA polymerase, and NSP7 and NSP8 play a role in guiding the long viral genomic RNA to RdRP. PLpro also plays an important role in prohibiting antiviral immunity in infected host cells. PLpro deconjugates interferon-stimulated gene 15 (ISG15) from the interferon regulatory factor (IRF3), leading to the inhibition of the interferon response [132]. However, the ISG15 released by PLpro is secreted into extracellular spaces and may induce proinflammatory cytokines, which may cause a cytokine storm [133]. Therefore, molecules that inhibit SARS-CoV-2 proteases and RdRP are attracting attention as potential drugs against SARS-CoV-2 [134,135,136,137].
The COVID-19 patients hospitalized in the intensive care unit met the Sepsis-3 septic shock definition and displayed organ dysfunctions [138]. The clinical presentation of a severe or critically ill COVID-19 patient is described as viral sepsis [139]. In addition, SARS CoV-2 evokes the disruption of intestinal barrier function, leading to the leakage of gut bacteria and endotoxin into blood and lymph vessels [140].
3.2.2. Procalcitonin and Adrenomedullin in COVID-19
PCT and ADM are diagnostic biomarkers for detecting the severity associated with COVID-19. The levels of PCT were elevated in patients with severe COVID-19 [87,88,89,90,91,92,93,94]. As in the case of CAP patients [86], the COVID-19 patients demonstrated an elevated PCT value despite a low prevalence of bacterial complications [91,141]. PCT levels were increased in patients with severe COVID-19, although PCT has been considered a diagnostic marker of sepsis caused by bacteria, not viruses. When viruses infect host cells, PRRs, such as RLRs and TLRs, sense viruses and promote the induction of type I IFNs (IFN-α, IFN-β) and type III IFN (IFN-λ). Both type I and III IFNs are innate cytokines that play an important role in defense against viruses [142]. However, the infection of SARS-CoV-2 leads to low levels of type I and type III interferons with increased cytokines [143]. Moreover, Orf9b in SARS-CoV-2 localizes to mitochondrial membranes and interacts with the mitochondrial import receptor Tom70 (translocase of the mitochondrial outer membrane 70), leading to the suppression of IFN-β expression [144]. On the other hand, PLpro from SARS-CoV-2 can release ISG15, leading to the induction of proinflammatory cytokines [133]. In addition, PLpro deconjugates ISG15 from IRF3, which inhibits interferon responses [132]. PCT is induced by proinflammatory cytokines and is suppressed by IFN. Therefore, the elevation of PCT in patients with COVID-19 may be due to the effects of the induction of proinflammatory cytokines in addition to the low level expression of IFNs and suppression of IFN response by SARS-CoV-2.
MR-proADM and bio-ADM serum levels are associated with severity and mortality in COVID-19 patients [21,145,146,147,148]. Furthermore, ADM levels are potent indicators of endothelial dysfunction and endotheliitis in patients diagnosed with COVID-19 [149,150,151]. Endothelial dysfunction is associated with multiple organ failure in COVID-19 [152,153]. Also, ADM plays an essential role in reducing endothelial permeability and promoting endothelial stability [58,59], as evidenced by phase 2a clinical trial involving an intravenous ADM infusion for treating COVID-19 patients [154].
4. Concluding Remarks
PCT and ADM have been considered as biological markers for sepsis. Both serum PCT and ADM levels are increased during severe sepsis and in COVID-19 patients. PCT is considered to be responsible severity and mortality of sepsis. In addition, PCT3-116 is considered as the main form of PCT in serum of patients with sepsis [48,49] and induces microvascular permeability [19]. In contrast, ADM lowers endothelial permeability and enhances endothelial stability [58,59]. Moreover, ADM reduces the mortality associated with sepsis and is used as a therapeutic molecule for COVID-19 [154]. Inhibiting the activity of PCT and promoting the activity of ADM can improve the severity and mortality associated with sepsis and in COVID-19 patients. Thus, developing effective drugs and antibodies targeting PCT and/or ADM to treat sepsis and COVID-19 patients is expected.
5. Future Directions
Both PCT and MD-proADM have been used as biomarkers of infectious diseases, with PCT reported to be involved in the severity of sepsis and ADM reported to decrease the severity of sepsis, contrary to PCT. Various functions have been reported for ADM, such as inhibition of vascular permeability, but the detailed physiological role of PCT has remained unclear. However, it was recently reported that PCT enhances cell permeability via phosphorylation of VE-cadherin. Thus, the opposing effects of PCT and ADM in sepsis are presumably due to the opposing effects of PCT and ADM on vascular permeability. Therefore, suppressing blood levels of bioactive PCTs or maintaining effective blood levels of bioactive ADMs may reduce the severity of sepsis. Detailed functional analysis of the physiological roles of PCT and ADM will be useful in reducing the severity of sepsis and other infectious diseases, and further research is expected to be developed. In addition, both PCT and ADM are expected to play a role as bioactive substances that contribute to treatment, in addition to their conventional role as biomarkers for clinical testing.
Author Contributions
Y.K. conceptualized and wrote this manuscript. H.N. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Tokushima Bunri University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Assicot, M.; Gendrel, D.; Carsin, H.; Raymond, J.; Guilbaud, J.; Bohuon, C. High serum procalcitonin concentrations in patients with sepsis and infection. Lancet 1993, 341, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Muller, B.; White, J.C.; Nylen, E.S.; Snider, R.H.; Becker, K.L.; Habener, J.F. Ubiquitous expression of the calcitonin-i gene in multiple tissues in response to sepsis. J. Clin. Endocrinol. Metab. 2001, 86, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Kohno, M.; Hanehira, T.; Kano, H.; Horio, T.; Yokokawa, K.; Ikeda, M.; Minami, M.; Yasunari, K.; Yoshikawa, J. Plasma adrenomedullin concentrations in essential hypertension. Hypertension 1996, 27, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Nishikimi, T.; Saito, Y.; Kitamura, K.; Ishimitsu, T.; Eto, T.; Kangawa, K.; Matsuo, H.; Omae, T.; Matsuoka, H. Increased plasma levels of adrenomedullin in patients with heart failure. J. Am. Coll. Cardiol. 1995, 26, 1424–1431. [Google Scholar] [CrossRef]
- Meng, D.Q.; Li, X.J.; Song, X.Y.; Xin, J.B.; Yang, W.B. Diagnostic and prognostic value of plasma adrenomedullin in COPD exacerbation. Respir. Care 2014, 59, 1542–1549. [Google Scholar] [CrossRef]
- Mandal, J.; Roth, M.; Papakonstantinou, E.; Fang, L.; Savic, S.; Tamm, M.; Stolz, D. Adrenomedullin mediates pro-angiogenic and pro-inflammatory cytokines in asthma and COPD. Pulm. Pharmacol. Ther. 2019, 56, 8–14. [Google Scholar] [CrossRef]
- Hirata, Y.; Mitaka, C.; Sato, K.; Nagura, T.; Tsunoda, Y.; Amaha, K.; Marumo, F. Increased circulating adrenomedullin, a novel vasodilatory peptide, in sepsis. J. Clin. Endocrinol. Metab. 1996, 81, 1449–1453. [Google Scholar]
- Meeran, K.; O’Shea, D.; Upton, P.D.; Small, C.J.; Ghatei, M.A.; Byfield, P.H.; Bloom, S.R. Circulating adrenomedullin does not regulate systemic blood pressure but increases plasma prolactin after intravenous infusion in humans: A pharmacokinetic study. J. Clin. Endocrinol. Metab. 1997, 82, 95–100. [Google Scholar] [CrossRef]
- Struck, J.; Tao, C.; Morgenthaler, N.G.; Bergmann, A. Identification of an Adrenomedullin precursor fragment in plasma of sepsis patients. Peptides 2004, 25, 1369–1372. [Google Scholar] [CrossRef]
- Weber, J.; Sachse, J.; Bergmann, S.; Sparwasser, A.; Struck, J.; Bergmann, A. Sandwich Immunoassay for Bioactive Plasma Adrenomedullin. J. Appl. Lab. Med. 2017, 2, 222–233. [Google Scholar] [CrossRef]
- Fugit, R.V.; McCoury, J.B.M.; Bessesen, M.T. Procalcitonin for sepsis management: Implementation within an antimicrobial stewardship program. Am. J. Health Syst. Pharm. 2022, 2022, zxac341. [Google Scholar] [CrossRef]
- Vijayan, A.L.; Vanimaya; Ravindran, S.; Saikant, R.; Lakshmi, S.; Kartik, R. Procalcitonin: A promising diagnostic marker for sepsis and antibiotic therapy. J. Intensive Care 2017, 5, 51. [Google Scholar] [CrossRef]
- Piccioni, A.; Saviano, A.; Cicchinelli, S.; Valletta, F.; Santoro, M.C.; de Cunzo, T.; Zanza, C.; Longhitano, Y.; Tullo, G.; Tilli, P.; et al. Proadrenomedullin in Sepsis and Septic Shock: A Role in the Emergency Department. Medicina 2021, 57, 920. [Google Scholar] [CrossRef]
- Martinez, J.M.; Wagner, K.E.; Snider, R.H.; Nylen, E.S.; Muller, B.; Sarani, B.; Becker, K.L.; White, J.C. Late immunoneutralization of procalcitonin arrests the progression of lethal porcine sepsis. Surg. Infect. 2001, 2, 193–202; discussion 193–202. [Google Scholar] [CrossRef]
- Tavares, E.; Maldonado, R.; Minano, F.J. Immunoneutralization of endogenous aminoprocalcitonin attenuates sepsis-induced acute lung injury and mortality in rats. Am. J. Pathol. 2014, 184, 3069–3083. [Google Scholar] [CrossRef]
- Wagner, K.E.; Martinez, J.M.; Vath, S.D.; Snider, R.H.; Nylen, E.S.; Becker, K.L.; Muller, B.; White, J.C. Early immunoneutralization of calcitonin precursors attenuates the adverse physiologic response to sepsis in pigs. Crit. Care Med. 2002, 30, 2313–2321. [Google Scholar] [CrossRef]
- Temmesfeld-Wollbruck, B.; Brell, B.; David, I.; Dorenberg, M.; Adolphs, J.; Schmeck, B.; Suttorp, N.; Hippenstiel, S. Adrenomedullin reduces vascular hyperpermeability and improves survival in rat septic shock. Intensive Care Med. 2007, 33, 703–710. [Google Scholar] [CrossRef]
- Ashizuka, S.; Kita, T.; Inatsu, H.; Kitamura, K. Adrenomedullin: A Novel Therapeutic for the Treatment of Inflammatory Bowel Disease. Biomedicines 2021, 9, 1068. [Google Scholar] [CrossRef]
- Brabenec, L.; Muller, M.; Hellenthal, K.E.M.; Karsten, O.S.; Pryvalov, H.; Otto, M.; Holthenrich, A.; Matos, A.L.L.; Weiss, R.; Kintrup, S.; et al. Targeting Procalcitonin Protects Vascular Barrier Integrity. Am. J. Respir. Crit. Care Med. 2022, 206, 488–500. [Google Scholar] [CrossRef]
- Hu, R.; Han, C.; Pei, S.; Yin, M.; Chen, X. Procalcitonin levels in COVID-19 patients. Int. J. Antimicrob. Agents 2020, 56, 106051. [Google Scholar] [CrossRef]
- Sozio, E.; Tascini, C.; Fabris, M.; D’Aurizio, F.; De Carlo, C.; Graziano, E.; Bassi, F.; Sbrana, F.; Ripoli, A.; Pagotto, A.; et al. MR-proADM as prognostic factor of outcome in COVID-19 patients. Sci. Rep. 2021, 11, 5121. [Google Scholar] [CrossRef] [PubMed]
- Naot, D.; Musson, D.S.; Cornish, J. The Activity of Peptides of the Calcitonin Family in Bone. Physiol. Rev. 2019, 99, 781–805. [Google Scholar] [CrossRef] [PubMed]
- Kittur, S.D.; Hoppener, J.W.; Antonarakis, S.E.; Daniels, J.D.; Meyers, D.A.; Maestri, N.E.; Jansen, M.; Korneluk, R.G.; Nelkin, B.D.; Kazazian, H.H., Jr. Linkage map of the short arm of human chromosome 11: Location of the genes for catalase, calcitonin, and insulin-like growth factor II. Proc. Natl. Acad. Sci. USA 1985, 82, 5064–5067. [Google Scholar] [CrossRef] [PubMed]
- Hoovers, J.M.; Redeker, E.; Speleman, F.; Hoppener, J.W.; Bhola, S.; Bliek, J.; van Roy, N.; Leschot, N.J.; Westerveld, A.; Mannens, M. High-resolution chromosomal localization of the human calcitonin/CGRP/IAPP gene family members. Genomics 1993, 15, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, M.G.; Mermod, J.J.; Amara, S.G.; Swanson, L.W.; Sawchenko, P.E.; Rivier, J.; Vale, W.W.; Evans, R.M. Production of a novel neuropeptide encoded by the calcitonin gene via tissue-specific RNA processing. Nature 1983, 304, 129–135. [Google Scholar] [CrossRef]
- Le Moullec, J.M.; Jullienne, A.; Chenais, J.; Lasmoles, F.; Guliana, J.M.; Milhaud, G.; Moukhtar, M.S. The complete sequence of human preprocalcitonin. FEBS Lett. 1984, 167, 93–97. [Google Scholar] [CrossRef]
- Gkonos, P.J.; Born, W.; Jones, B.N.; Petermann, J.B.; Keutmann, H.T.; Birnbaum, R.S.; Fischer, J.A.; Roos, B.A. Biosynthesis of calcitonin gene-related peptide and calcitonin by a human medullary thyroid carcinoma cell line. J. Biol. Chem. 1986, 261, 14386–14391. [Google Scholar] [CrossRef]
- Kiriakopoulos, A.; Giannakis, P.; Menenakos, E. Calcitonin: Current concepts and differential diagnosis. Ther. Adv. Endocrinol. Metab. 2022, 13, 20420188221099344. [Google Scholar] [CrossRef]
- Kokkoris, S.; Andrews, P.; Webb, D.J. Role of calcitonin gene-related peptide in cerebral vasospasm, and as a therapeutic approach to subarachnoid hemorrhage. Front. Endocrinol. 2012, 3, 135. [Google Scholar] [CrossRef]
- Mulinacci, F.; Poirier, E.; Capelle, M.A.; Gurny, R.; Arvinte, T. Enhanced physical stability of human calcitonin after methionine oxidation. Eur. J. Pharm. Biopharm. 2011, 78, 229–238. [Google Scholar] [CrossRef]
- Kiriyama, Y.; Tsuchiya, H.; Murakami, T.; Satoh, K.; Tokumitsu, Y. Calcitonin induces IL-6 production via both PKA and PKC pathways in the pituitary folliculo-stellate cell line. Endocrinology 2001, 142, 3563–3569. [Google Scholar] [CrossRef]
- Serafin, D.S.; Harris, N.R.; Nielsen, N.R.; Mackie, D.I.; Caron, K.M. Dawn of a New RAMPage. Trends Pharmacol. Sci. 2020, 41, 249–265. [Google Scholar] [CrossRef]
- Kiriyama, Y.; Nochi, H. Role and Cytotoxicity of Amylin and Protection of Pancreatic Islet beta-Cells from Amylin Cytotoxicity. Cells 2018, 7, 95. [Google Scholar] [CrossRef]
- McLatchie, L.M.; Fraser, N.J.; Main, M.J.; Wise, A.; Brown, J.; Thompson, N.; Solari, R.; Lee, M.G.; Foord, S.M. RAMPs regulate the transport and ligand specificity of the calcitonin-receptor-like receptor. Nature 1998, 393, 333–339. [Google Scholar] [CrossRef]
- Hay, D.L.; Garelja, M.L.; Poyner, D.R.; Walker, C.S. Update on the pharmacology of calcitonin/CGRP family of peptides: IUPHAR Review 25. Br. J. Pharmacol. 2018, 175, 3–17. [Google Scholar] [CrossRef]
- Klein, K.R.; Matson, B.C.; Caron, K.M. The expanding repertoire of receptor activity modifying protein (RAMP) function. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 65–71. [Google Scholar] [CrossRef]
- Sexton, P.M.; Christopoulos, G.; Christopoulos, A.; Nylen, E.S.; Snider, R.H., Jr.; Becker, K.L. Procalcitonin has bioactivity at calcitonin receptor family complexes: Potential mediator implications in sepsis. Crit. Care Med. 2008, 36, 1637–1640. [Google Scholar] [CrossRef]
- Kiriyama, Y.; Nomura, Y.; Tokumitsu, Y. Calcitonin gene expression induced by lipopolysaccharide in the rat pituitary. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E1380–E1384. [Google Scholar] [CrossRef]
- Dandona, P.; Nix, D.; Wilson, M.F.; Aljada, A.; Love, J.; Assicot, M.; Bohuon, C. Procalcitonin increase after endotoxin injection in normal subjects. J. Clin. Endocrinol. Metab. 1994, 79, 1605–1608. [Google Scholar]
- Maiese, A.; Del Nonno, F.; Dell’Aquila, M.; Moauro, M.; Baiocchini, A.; Mastracchio, A.; Bolino, G. Postmortem diagnosis of sepsis: A preliminary immunohistochemical study with an anti-procalcitonin antibody. Leg. Med. 2017, 28, 1–5. [Google Scholar] [CrossRef]
- Morgenthaler, N.G.; Struck, J.; Chancerelle, Y.; Weglohner, W.; Agay, D.; Bohuon, C.; Suarez-Domenech, V.; Bergmann, A.; Muller, B. Production of procalcitonin (PCT) in non-thyroidal tissue after LPS injection. Horm. Metab. Res. 2003, 35, 290–295. [Google Scholar] [PubMed]
- Attia, A.M.; Abo El-Atta, H.M.; El-sherbiny, M.; El-Shahat, E.E. Evaluation of procalcitonin postmortem levels in some models of death: An experimental study. J. Forensic. Leg. Med. 2016, 37, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Hagner, S.; Stahl, U.; Knoblauch, B.; McGregor, G.P.; Lang, R.E. Calcitonin receptor-like receptor: Identification and distribution in human peripheral tissues. Cell Tissue Res. 2002, 310, 41–50. [Google Scholar] [CrossRef] [PubMed]
- La Russa, R.; Maiese, A.; Viola, R.V.; De Matteis, A.; Pinchi, E.; Frati, P.; Fineschi, V. Searching for highly sensitive and specific biomarkers for sepsis: State-of-the-art in post-mortem diagnosis of sepsis through immunohistochemical analysis. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738419855226. [Google Scholar] [CrossRef] [PubMed]
- Joffre, J.; Hellman, J.; Ince, C.; Ait-Oufella, H. Endothelial Responses in Sepsis. Am. J. Respir. Crit. Care Med. 2020, 202, 361–370. [Google Scholar] [CrossRef]
- Park-Windhol, C.; D’Amore, P.A. Disorders of Vascular Permeability. Annu. Rev. Pathol. 2016, 11, 251–281. [Google Scholar] [CrossRef]
- Wrenger, S.; Kahne, T.; Bohuon, C.; Weglohner, W.; Ansorge, S.; Reinhold, D. Amino-terminal truncation of procalcitonin, a marker for systemic bacterial infections, by dipeptidyl peptidase IV (DP IV). FEBS Lett. 2000, 466, 155–159. [Google Scholar] [CrossRef]
- Weglohner, W.; Struck, J.; Fischer-Schulz, C.; Morgenthaler, N.G.; Otto, A.; Bohuon, C.; Bergmann, A. Isolation and characterization of serum procalcitonin from patients with sepsis. Peptides 2001, 22, 2099–2103. [Google Scholar] [CrossRef]
- Struck, J.; Strebelow, M.; Tietz, S.; Alonso, C.; Morgenthaler, N.G.; van der Hoeven, J.G.; Pickkers, P.; Bergmann, A. Method for the selective measurement of amino-terminal variants of procalcitonin. Clin. Chem. 2009, 55, 1672–1679. [Google Scholar] [CrossRef]
- Sidibe, A.; Imhof, B.A. VE-cadherin phosphorylation decides: Vascular permeability or diapedesis. Nat. Immunol. 2014, 15, 215–217. [Google Scholar] [CrossRef]
- Chan, Y.H.; Harith, H.H.; Israf, D.A.; Tham, C.L. Differential Regulation of LPS-Mediated VE-Cadherin Disruption in Human Endothelial Cells and the Underlying Signaling Pathways: A Mini Review. Front. Cell Dev. Biol. 2019, 7, 280. [Google Scholar] [CrossRef]
- Cao, J.; Schnittler, H. Putting VE-cadherin into JAIL for junction remodeling. J. Cell Sci. 2019, 132, jcs222893. [Google Scholar] [CrossRef] [Green Version]
- Hinson, J.P.; Kapas, S.; Smith, D.M. Adrenomedullin, a multifunctional regulatory peptide. Endocr. Rev. 2000, 21, 138–167. [Google Scholar]
- Ichiki, Y.; Kitamura, K.; Kangawa, K.; Kawamoto, M.; Matsuo, H.; Eto, T. Distribution and characterization of immunoreactive adrenomedullin in human tissue and plasma. FEBS Lett. 1994, 338, 6–10. [Google Scholar] [CrossRef]
- Ishimitsu, T.; Kojima, M.; Kangawa, K.; Hino, J.; Matsuoka, H.; Kitamura, K.; Eto, T.; Matsuo, H. Genomic structure of human adrenomedullin gene. Biochem. Biophys. Res. Commun. 1994, 203, 631–639. [Google Scholar] [CrossRef]
- Gumusel, B.; Chang, J.K.; Hyman, A.; Lippton, H. Adrenotensin: An ADM gene product with the opposite effects of ADM. Life Sci. 1995, 57, PL87–PL90. [Google Scholar] [CrossRef]
- Martinez, A.; Hodge, D.L.; Garayoa, M.; Young, H.A.; Cuttitta, F. Alternative splicing of the proadrenomedullin gene results in differential expression of gene products. J. Mol. Endocrinol. 2001, 27, 31–41. [Google Scholar] [CrossRef]
- Dunworth, W.P.; Fritz-Six, K.L.; Caron, K.M. Adrenomedullin stabilizes the lymphatic endothelial barrier in vitro and in vivo. Peptides 2008, 29, 2243–2249. [Google Scholar] [CrossRef]
- Hippenstiel, S.; Witzenrath, M.; Schmeck, B.; Hocke, A.; Krisp, M.; Krull, M.; Seybold, J.; Seeger, W.; Rascher, W.; Schutte, H.; et al. Adrenomedullin reduces endothelial hyperpermeability. Circ. Res. 2002, 91, 618–625. [Google Scholar] [CrossRef]
- Meyrath, M.; Palmer, C.B.; Reynders, N.; Vanderplasschen, A.; Ollert, M.; Bouvier, M.; Szpakowska, M.; Chevigne, A. Proadrenomedullin N-Terminal 20 Peptides (PAMPs) Are Agonists of the Chemokine Scavenger Receptor ACKR3/CXCR7. ACS Pharmacol. Transl. Sci. 2021, 4, 813–823. [Google Scholar] [CrossRef]
- Kuwasako, K.; Kitamura, K.; Ishiyama, Y.; Washimine, H.; Kato, J.; Kangawa, K.; Eto, T. Purification and characterization of PAMP-12 (PAMP[9-20]) in porcine adrenal medulla as a major endogenous biologically active peptide. FEBS Lett. 1997, 414, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Ajish, C.; Yang, S.; Kumar, S.D.; Shin, S.Y. Proadrenomedullin N-terminal 20 peptide (PAMP) and its C-terminal 12-residue peptide, PAMP(9-20): Cell selectivity and antimicrobial mechanism. Biochem. Biophys. Res. Commun. 2020, 527, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- van der Poll, T.; van de Veerdonk, F.L.; Scicluna, B.P.; Netea, M.G. The immunopathology of sepsis and potential therapeutic targets. Nat. Rev. Immunol. 2017, 17, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.L.; Moreno, R.; Takala, J.; Willatts, S.; De Mendonca, A.; Bruining, H.; Reinhart, C.K.; Suter, P.M.; Thijs, L.G. The SOFA (Sepsis-related Organ Failure Assessment) score to describe organ dysfunction/failure. On behalf of the Working Group on Sepsis-Related Problems of the European Society of Intensive Care Medicine. Intensive Care Med. 1996, 22, 707–710. [Google Scholar] [CrossRef]
- Oczkowski, S.; Alshamsi, F.; Belley-Cote, E.; Centofanti, J.E.; Hylander Moller, M.; Nunnaly, M.E.; Alhazzani, W. Surviving Sepsis Campaign Guidelines 2021: Highlights for the practicing clinician. Pol. Arch. Intern. Med. 2022, 132, 16290. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr. Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb. Symp. Quant. Biol. 1989, 54 Pt 1, 1–13. [Google Scholar] [CrossRef]
- Brubaker, S.W.; Bonham, K.S.; Zanoni, I.; Kagan, J.C. Innate immune pattern recognition: A cell biological perspective. Annu. Rev. Immunol. 2015, 33, 257–290. [Google Scholar] [CrossRef]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal Transduct. Target Ther. 2021, 6, 291. [Google Scholar] [CrossRef]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef]
- Jarczak, D.; Kluge, S.; Nierhaus, A. Sepsis-Pathophysiology and Therapeutic Concepts. Front. Med. 2021, 8, 628302. [Google Scholar] [CrossRef]
- Nylen, E.S.; Whang, K.T.; Snider, R.H., Jr.; Steinwald, P.M.; White, J.C.; Becker, K.L. Mortality is increased by procalcitonin and decreased by an antiserum reactive to procalcitonin in experimental sepsis. Crit. Care Med. 1998, 26, 1001–1006. [Google Scholar] [CrossRef]
- Oberhoffer, M.; Stonans, I.; Russwurm, S.; Stonane, E.; Vogelsang, H.; Junker, U.; Jager, L.; Reinhart, K. Procalcitonin expression in human peripheral blood mononuclear cells and its modulation by lipopolysaccharides and sepsis-related cytokines in vitro. J. Lab. Clin. Med. 1999, 134, 49–55. [Google Scholar] [CrossRef]
- Bai, Y.; Lu, J.; Cheng, Y.; Zhang, F.; Fan, X.; Weng, Y.; Zhu, J. NF-small ka, CyrillicB increases LPS-mediated procalcitonin production in human hepatocytes. Sci. Rep. 2018, 8, 8913. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Oussalah, A.; Ferrand, J.; Filhine-Tresarrieu, P.; Aissa, N.; Aimone-Gastin, I.; Namour, F.; Garcia, M.; Lozniewski, A.; Gueant, J.L. Diagnostic Accuracy of Procalcitonin for Predicting Blood Culture Results in Patients With Suspected Bloodstream Infection: An Observational Study of 35,343 Consecutive Patients (A STROBE-Compliant Article). Medicine 2015, 94, e1774. [Google Scholar] [CrossRef]
- Li, S.; Rong, H.; Guo, Q.; Chen, Y.; Zhang, G.; Yang, J. Serum procalcitonin levels distinguish Gram-negative bacterial sepsis from Gram-positive bacterial and fungal sepsis. J. Res. Med. Sci. 2016, 21, 39. [Google Scholar]
- Brodska, H.; Malickova, K.; Adamkova, V.; Benakova, H.; Stastna, M.M.; Zima, T. Significantly higher procalcitonin levels could differentiate Gram-negative sepsis from Gram-positive and fungal sepsis. Clin. Exp. Med. 2013, 13, 165–170. [Google Scholar] [CrossRef]
- Xu, H.G.; Tian, M.; Pan, S.Y. Clinical utility of procalcitonin and its association with pathogenic microorganisms. Crit. Rev. Clin. Lab. Sci. 2022, 59, 93–111. [Google Scholar] [CrossRef]
- Garcia-Sastre, A. Ten Strategies of Interferon Evasion by Viruses. Cell Host. Microbe. 2017, 22, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Nan, Y.; Nan, G.; Zhang, Y.J. Interferon induction by RNA viruses and antagonism by viral pathogens. Viruses 2014, 6, 4999–5027. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol 2015, 15, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Linscheid, P.; Seboek, D.; Nylen, E.S.; Langer, I.; Schlatter, M.; Becker, K.L.; Keller, U.; Muller, B. In vitro and in vivo calcitonin I gene expression in parenchymal cells: A novel product of human adipose tissue. Endocrinology 2003, 144, 5578–5584. [Google Scholar] [CrossRef]
- Gautam, S.; Cohen, A.J.; Stahl, Y.; Valda Toro, P.; Young, G.M.; Datta, R.; Yan, X.; Ristic, N.T.; Bermejo, S.D.; Sharma, L.; et al. Severe respiratory viral infection induces procalcitonin in the absence of bacterial pneumonia. Thorax 2020, 75, 974–981. [Google Scholar] [CrossRef]
- Kamat, I.S.; Ramachandran, V.; Eswaran, H.; Guffey, D.; Musher, D.M. Procalcitonin to Distinguish Viral From Bacterial Pneumonia: A Systematic Review and Meta-analysis. Clin. Infect. Dis. 2020, 70, 538–542. [Google Scholar] [CrossRef]
- Fabre, V.; Karaba, S.; Amoah, J.; Robinson, M.; Jones, G.; Dzintars, K.; Katz, M.; Landrum, B.M.; Qasba, S.; Gupta, P.; et al. The role of procalcitonin results in antibiotic decision-making in coronavirus disease 2019 (COVID-19). Infect. Control Hosp. Epidemiol. 2022, 43, 570–575. [Google Scholar] [CrossRef]
- Zhang, J.J.; Dong, X.; Cao, Y.Y.; Yuan, Y.D.; Yang, Y.B.; Yan, Y.Q.; Akdis, C.A.; Gao, Y.D. Clinical characteristics of 140 patients infected with SARS-CoV-2 in Wuhan, China. Allergy 2020, 75, 1730–1741. [Google Scholar] [CrossRef]
- Lippi, G.; Plebani, M. Laboratory abnormalities in patients with COVID-2019 infection. Clin. Chem. Lab. Med. 2020, 58, 1131–1134. [Google Scholar] [CrossRef]
- Aon, M.; Alsaeedi, A.; Alzafiri, A.; Ibrahim, M.M.; Al-Shammari, A.; Al-Shammari, O.; Tawakul, M.; Taha, S.; Alherz, N.; Alshammari, J.; et al. The Association between Admission Procalcitonin Level and The Severity of COVID-19 Pneumonia: A Retrospective Cohort Study. Medicina 2022, 58, 1389. [Google Scholar] [CrossRef]
- Vanhomwegen, C.; Veliziotis, I.; Malinverni, S.; Konopnicki, D.; Dechamps, P.; Claus, M.; Roman, A.; Cotton, F.; Dauby, N. Procalcitonin accurately predicts mortality but not bacterial infection in COVID-19 patients admitted to intensive care unit. Ir. J. Med. Sci. 2021, 190, 1649–1652. [Google Scholar] [CrossRef]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef]
- Chen, T.; Wu, D.; Chen, H.; Yan, W.; Yang, D.; Chen, G.; Ma, K.; Xu, D.; Yu, H.; Wang, H.; et al. Clinical characteristics of 113 deceased patients with coronavirus disease 2019: Retrospective study. BMJ 2020, 368, m1091. [Google Scholar] [CrossRef]
- Xu, J.B.; Xu, C.; Zhang, R.B.; Wu, M.; Pan, C.K.; Li, X.J.; Wang, Q.; Zeng, F.F.; Zhu, S. Associations of procalcitonin, C-reaction protein and neutrophil-to-lymphocyte ratio with mortality in hospitalized COVID-19 patients in China. Sci. Rep. 2020, 10, 15058. [Google Scholar] [CrossRef]
- Schuetz, P.; Beishuizen, A.; Broyles, M.; Ferrer, R.; Gavazzi, G.; Gluck, E.H.; Gonzalez Del Castillo, J.; Jensen, J.U.; Kanizsai, P.L.; Kwa, A.L.H.; et al. Procalcitonin (PCT)-guided antibiotic stewardship: An international experts consensus on optimized clinical use. Clin. Chem. Lab. Med. 2019, 57, 1308–1318. [Google Scholar] [CrossRef]
- Wirz, Y.; Meier, M.A.; Bouadma, L.; Luyt, C.E.; Wolff, M.; Chastre, J.; Tubach, F.; Schroeder, S.; Nobre, V.; Annane, D.; et al. Effect of procalcitonin-guided antibiotic treatment on clinical outcomes in intensive care unit patients with infection and sepsis patients: A patient-level meta-analysis of randomized trials. Crit. Care 2018, 22, 191. [Google Scholar] [CrossRef]
- Meier, M.A.; Branche, A.; Neeser, O.L.; Wirz, Y.; Haubitz, S.; Bouadma, L.; Wolff, M.; Luyt, C.E.; Chastre, J.; Tubach, F.; et al. Procalcitonin-guided Antibiotic Treatment in Patients With Positive Blood Cultures: A Patient-level Meta-analysis of Randomized Trials. Clin. Infect. Dis. 2019, 69, 388–396. [Google Scholar] [CrossRef]
- Lippi, G.; Salvagno, G.L.; Gelati, M.; Pucci, M.; Lo Cascio, C.; Demonte, D.; Faggian, D.; Plebani, M. Two-center comparison of 10 fully-automated commercial procalcitonin (PCT) immunoassays. Clin. Chem. Lab. Med. 2019, 58, 77–84. [Google Scholar] [CrossRef]
- Kylanpaa-Back, M.L.; Takala, A.; Kemppainen, E.; Puolakkainen, P.; Haapiainen, R.; Repo, H. Procalcitonin strip test in the early detection of severe acute pancreatitis. Br. J. Surg. 2001, 88, 222–227. [Google Scholar] [CrossRef]
- Kutz, A.; Hausfater, P.; Oppert, M.; Alan, M.; Grolimund, E.; Gast, C.; Alonso, C.; Wissmann, C.; Kuehn, C.; Bernard, M.; et al. Comparison between B.R.A.H.M.S PCT direct, a new sensitive point-of-care testing device for rapid quantification of procalcitonin in emergency department patients and established reference methods—A prospective multinational trial. Clin. Chem. Lab. Med. 2016, 54, 577–584. [Google Scholar] [CrossRef]
- Marino, R.; Struck, J.; Maisel, A.S.; Magrini, L.; Bergmann, A.; Di Somma, S. Plasma adrenomedullin is associated with short-term mortality and vasopressor requirement in patients admitted with sepsis. Crit. Care 2014, 18, R34. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.X.; Li, C.S. Prognostic value of adrenomedullin in septic patients in the ED. Am. J. Emerg. Med. 2013, 31, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Caironi, P.; Latini, R.; Struck, J.; Hartmann, O.; Bergmann, A.; Maggio, G.; Cavana, M.; Tognoni, G.; Pesenti, A.; Gattinoni, L.; et al. Circulating Biologically Active Adrenomedullin (bio-ADM) Predicts Hemodynamic Support Requirement and Mortality During Sepsis. Chest 2017, 152, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Mebazaa, A.; Geven, C.; Hollinger, A.; Wittebole, X.; Chousterman, B.G.; Blet, A.; Gayat, E.; Hartmann, O.; Scigalla, P.; Struck, J.; et al. Circulating adrenomedullin estimates survival and reversibility of organ failure in sepsis: The prospective observational multinational Adrenomedullin and Outcome in Sepsis and Septic Shock-1 (AdrenOSS-1) study. Crit. Care 2018, 22, 354. [Google Scholar] [CrossRef] [Green Version]
- Sugo, S.; Minamino, N.; Shoji, H.; Kangawa, K.; Kitamura, K.; Eto, T.; Matsuo, H. Interleukin-1, tumor necrosis factor and lipopolysaccharide additively stimulate production of adrenomedullin in vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 1995, 207, 25–32. [Google Scholar] [CrossRef]
- Garayoa, M.; Martinez, A.; Lee, S.; Pio, R.; An, W.G.; Neckers, L.; Trepel, J.; Montuenga, L.M.; Ryan, H.; Johnson, R.; et al. Hypoxia-inducible factor-1 (HIF-1) up-regulates adrenomedullin expression in human tumor cell lines during oxygen deprivation: A possible promotion mechanism of carcinogenesis. Mol. Endocrinol. 2000, 14, 848–862. [Google Scholar] [CrossRef]
- Chang, J.C. Sepsis and septic shock: Endothelial molecular pathogenesis associated with vascular microthrombotic disease. Thromb. J. 2019, 17, 10. [Google Scholar] [CrossRef]
- Lee, P.; Chandel, N.S.; Simon, M.C. Cellular adaptation to hypoxia through hypoxia inducible factors and beyond. Nat. Rev. Mol. Cell Biol. 2020, 21, 268–283. [Google Scholar] [CrossRef]
- Luo, Z.; Tian, M.; Yang, G.; Tan, Q.; Chen, Y.; Li, G.; Zhang, Q.; Li, Y.; Wan, P.; Wu, J. Hypoxia signaling in human health and diseases: Implications and prospects for therapeutics. Signal Transduct. Target Ther. 2022, 7, 218. [Google Scholar] [CrossRef]
- Chowdhury, R.; Leung, I.K.; Tian, Y.M.; Abboud, M.I.; Ge, W.; Domene, C.; Cantrelle, F.X.; Landrieu, I.; Hardy, A.P.; Pugh, C.W.; et al. Structural basis for oxygen degradation domain selectivity of the HIF prolyl hydroxylases. Nat. Commun. 2016, 7, 12673. [Google Scholar] [CrossRef]
- Yamauchi, A.; Sakurai, T.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Kawate, H.; Igarashi, K.; Toriyama, Y.; Tanaka, M.; Liu, T.; Xian, X.; et al. Functional differentiation of RAMP2 and RAMP3 in their regulation of the vascular system. J. Mol. Cell Cardiol. 2014, 77, 73–85. [Google Scholar] [CrossRef]
- Cui, N.; Sakurai, T.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Kawate, H.; Tanaka, M.; Tanaka, M.; Wei, Y.; Kakihara, S.; Zhao, Y.; et al. Adrenomedullin-RAMP2 and -RAMP3 Systems Regulate Cardiac Homeostasis during Cardiovascular Stress. Endocrinology 2021, 162, bqab001. [Google Scholar] [CrossRef]
- Ishizaka, Y.; Ishizaka, Y.; Tanaka, M.; Kitamura, K.; Kangawa, K.; Minamino, N.; Matsuo, H.; Eto, T. Adrenomedullin stimulates cyclic AMP formation in rat vascular smooth muscle cells. Biochem. Biophys. Res. Commun 1994, 200, 642–646. [Google Scholar] [CrossRef]
- Fukuhara, S.; Sakurai, A.; Sano, H.; Yamagishi, A.; Somekawa, S.; Takakura, N.; Saito, Y.; Kangawa, K.; Mochizuki, N. Cyclic AMP potentiates vascular endothelial cadherin-mediated cell-cell contact to enhance endothelial barrier function through an Epac-Rap1 signaling pathway. Mol. Cell Biol. 2005, 25, 136–146. [Google Scholar] [CrossRef] [Green Version]
- de Rooij, J.; Zwartkruis, F.J.; Verheijen, M.H.; Cool, R.H.; Nijman, S.M.; Wittinghofer, A.; Bos, J.L. Epac is a Rap1 guanine-nucleotide-exchange factor directly activated by cyclic AMP. Nature 1998, 396, 474–477. [Google Scholar] [CrossRef]
- Cullere, X.; Shaw, S.K.; Andersson, L.; Hirahashi, J.; Luscinskas, F.W.; Mayadas, T.N. Regulation of vascular endothelial barrier function by Epac, a cAMP-activated exchange factor for Rap GTPase. Blood 2005, 105, 1950–1955. [Google Scholar] [CrossRef]
- Kooistra, M.R.; Corada, M.; Dejana, E.; Bos, J.L. Epac1 regulates integrity of endothelial cell junctions through VE-cadherin. FEBS Lett. 2005, 579, 4966–4972. [Google Scholar] [CrossRef]
- Pio, R.; Martinez, A.; Unsworth, E.J.; Kowalak, J.A.; Bengoechea, J.A.; Zipfel, P.F.; Elsasser, T.H.; Cuttitta, F. Complement factor H is a serum-binding protein for adrenomedullin, and the resulting complex modulates the bioactivities of both partners. J. Biol. Chem. 2001, 276, 12292–12300. [Google Scholar] [CrossRef]
- Morgenthaler, N.G.; Struck, J.; Alonso, C.; Bergmann, A. Measurement of midregional proadrenomedullin in plasma with an immunoluminometric assay. Clin. Chem. 2005, 51, 1823–1829. [Google Scholar] [CrossRef]
- Onal, U.; Valenzuela-Sanchez, F.; Vandana, K.E.; Rello, J. Mid-Regional Pro-Adrenomedullin (MR-proADM) as a Biomarker for Sepsis and Septic Shock: Narrative Review. Healthcare 2018, 6, 110. [Google Scholar] [CrossRef]
- Elke, G.; Bloos, F.; Wilson, D.C.; Brunkhorst, F.M.; Briegel, J.; Reinhart, K.; Loeffler, M.; Kluge, S.; Nierhaus, A.; Jaschinski, U.; et al. The use of mid-regional proadrenomedullin to identify disease severity and treatment response to sepsis—A secondary analysis of a large randomised controlled trial. Crit. Care 2018, 22, 79. [Google Scholar] [CrossRef] [PubMed]
- Andres, C.; Andaluz-Ojeda, D.; Cicuendez, R.; Nogales, L.; Martin, S.; Martin-Fernandez, M.; Almansa, R.; Calvo, D.; Esteban-Velasco, M.C.; Vaquero-Roncero, L.M.; et al. MR- proADM to detect specific types of organ failure in infection. Eur. J. Clin. Investig. 2020, 50, e13246. [Google Scholar] [CrossRef] [PubMed]
- Watkins, H.A.; Au, M.; Bobby, R.; Archbold, J.K.; Abdul-Manan, N.; Moore, J.M.; Middleditch, M.J.; Williams, G.M.; Brimble, M.A.; Dingley, A.J.; et al. Identification of key residues involved in adrenomedullin binding to the AM1 receptor. Br. J. Pharmacol. 2013, 169, 143–155. [Google Scholar] [CrossRef]
- Geven, C.; van Lier, D.; Blet, A.; Peelen, R.; Ten Elzen, B.; Mebazaa, A.; Kox, M.; Pickkers, P. Safety, tolerability and pharmacokinetics/pharmacodynamics of the adrenomedullin antibody adrecizumab in a first-in-human study and during experimental human endotoxaemia in healthy subjects. Br. J. Clin. Pharmacol. 2018, 84, 2129–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Ahmad Farouk, I.; Lal, S.K. COVID-19: A Review on the Novel Coronavirus Disease Evolution, Transmission, Detection, Control and Prevention. Viruses 2021, 13, 202. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Nazerian, Y.; Ghasemi, M.; Yassaghi, Y.; Nazerian, A.; Hashemi, S.M. Role of SARS-CoV-2-induced cytokine storm in multi-organ failure: Molecular pathways and potential therapeutic options. Int. Immunopharmacol. 2022, 113, 109428. [Google Scholar] [CrossRef]
- White-Dzuro, G.; Gibson, L.E.; Zazzeron, L.; White-Dzuro, C.; Sullivan, Z.; Diiorio, D.A.; Low, S.A.; Chang, M.G.; Bittner, E.A. Multisystem effects of COVID-19: A concise review for practitioners. Postgrad. Med. 2021, 133, 20–27. [Google Scholar] [CrossRef]
- Thomas, E.; Delabat, S.; Carattini, Y.L.; Andrews, D.M. SARS-CoV-2 and Variant Diagnostic Testing Approaches in the United States. Viruses 2021, 13, 2492. [Google Scholar] [CrossRef]
- Suarez, D.; Diaz, N. SARS-CoV-2 Main Protease: A Molecular Dynamics Study. J. Chem. Inf. Model. 2020, 60, 5815–5831. [Google Scholar] [CrossRef]
- Hillen, H.S.; Kokic, G.; Farnung, L.; Dienemann, C.; Tegunov, D.; Cramer, P. Structure of replicating SARS-CoV-2 polymerase. Nature 2020, 584, 154–156. [Google Scholar] [CrossRef]
- Shin, D.; Mukherjee, R.; Grewe, D.; Bojkova, D.; Baek, K.; Bhattacharya, A.; Schulz, L.; Widera, M.; Mehdipour, A.R.; Tascher, G.; et al. Papain-like protease regulates SARS-CoV-2 viral spread and innate immunity. Nature 2020, 587, 657–662. [Google Scholar] [CrossRef]
- Munnur, D.; Teo, Q.; Eggermont, D.; Lee, H.H.Y.; Thery, F.; Ho, J.; van Leur, S.W.; Ng, W.W.S.; Siu, L.Y.L.; Beling, A.; et al. Altered ISGylation drives aberrant macrophage-dependent immune responses during SARS-CoV-2 infection. Nat. Immunol. 2021, 22, 1416–1427. [Google Scholar] [CrossRef]
- Ng, T.I.; Correia, I.; Seagal, J.; DeGoey, D.A.; Schrimpf, M.R.; Hardee, D.J.; Noey, E.L.; Kati, W.M. Antiviral Drug Discovery for the Treatment of COVID-19 Infections. Viruses 2022, 14, 961. [Google Scholar] [CrossRef]
- Kiriyama, Y.; Nochi, H. Inhibitory effects of olive-derived phytochemicals on SARS-CoV-2 3C-like protease (3CLpro). BPB Rep. 2023, 6, 8–10. [Google Scholar] [CrossRef]
- Chary, M.A.; Barbuto, A.F.; Izadmehr, S.; Tarsillo, M.; Fleischer, E.; Burns, M.M. COVID-19 Therapeutics: Use, Mechanism of Action, and Toxicity (Xenobiotics). J. Med. Toxicol. 2023, 19, 26–36. [Google Scholar] [CrossRef]
- Bijelic, K.; Hitl, M.; Kladar, N. Phytochemicals in the Prevention and Treatment of SARS-CoV-2-Clinical Evidence. Antibiotics 2022, 11, 1614. [Google Scholar] [CrossRef]
- Karakike, E.; Giamarellos-Bourboulis, E.J.; Kyprianou, M.; Fleischmann-Struzek, C.; Pletz, M.W.; Netea, M.G.; Reinhart, K.; Kyriazopoulou, E. Coronavirus Disease 2019 as Cause of Viral Sepsis: A Systematic Review and Meta-Analysis. Crit. Care Med. 2021, 49, 2042–2057. [Google Scholar] [CrossRef]
- Li, H.; Liu, L.; Zhang, D.; Xu, J.; Dai, H.; Tang, N.; Su, X.; Cao, B. SARS-CoV-2 and viral sepsis: Observations and hypotheses. Lancet 2020, 395, 1517–1520. [Google Scholar] [CrossRef]
- Assimakopoulos, S.F.; Eleftheriotis, G.; Lagadinou, M.; Karamouzos, V.; Dousdampanis, P.; Siakallis, G.; Marangos, M. SARS CoV-2-Induced Viral Sepsis: The Role of Gut Barrier Dysfunction. Microorganisms 2022, 10, 1050. [Google Scholar] [CrossRef]
- Conlon, A.S.C.; Chopra, Z.; Cahalan, S.; Cinti, S.; Rao, K. Effects of procalcitonin on antimicrobial treatment decisions in patients with coronavirus disease 2019 (COVID-19). Infect. Control Hosp. Epidemiol. 2022, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lazear, H.M.; Schoggins, J.W.; Diamond, M.S. Shared and Distinct Functions of Type I and Type III Interferons. Immunity 2019, 50, 907–923. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e1039. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.W.; Zhang, H.N.; Meng, Q.F.; Xie, J.; Li, Y.; Chen, H.; Zheng, Y.X.; Wang, X.N.; Qi, H.; Zhang, J.; et al. SARS-CoV-2 Orf9b suppresses type I interferon responses by targeting TOM70. Cell Mol. Immunol. 2020, 17, 998–1000. [Google Scholar] [CrossRef]
- Spoto, S.; Agro, F.E.; Sambuco, F.; Travaglino, F.; Valeriani, E.; Fogolari, M.; Mangiacapra, F.; Costantino, S.; Ciccozzi, M.; Angeletti, S. High value of mid-regional proadrenomedullin in COVID-19: A marker of widespread endothelial damage, disease severity, and mortality. J. Med. Virol. 2021, 93, 2820–2827. [Google Scholar] [CrossRef]
- Simon, T.P.; Stoppe, C.; Breuer, T.; Stiehler, L.; Dreher, M.; Kersten, A.; Kluge, S.; Karakas, M.; Zechendorf, E.; Marx, G.; et al. Prognostic Value of Bioactive Adrenomedullin in Critically Ill Patients with COVID-19 in Germany: An Observational Cohort Study. J. Clin. Med. 2021, 10, 1667. [Google Scholar] [CrossRef]
- Mangioni, D.; Oggioni, M.; Chatenoud, L.; Liparoti, A.; Uceda Renteria, S.; Alagna, L.; Biscarini, S.; Bolis, M.; Di Modugno, A.; Mussa, M.; et al. Prognostic Value of Mid-Region Proadrenomedullin and In Vitro Interferon Gamma Production for In-Hospital Mortality in Patients with COVID-19 Pneumonia and Respiratory Failure: An Observational Prospective Study. Viruses 2022, 14, 1683. [Google Scholar] [CrossRef]
- Papasidero, I.D.; Valli, G.; Marin, D.; Del Sasso, A.; De Magistris, A.; Cennamo, E.; Casalboni, S.; De Marco, F.; Rocchi, R.; Ndogmo Beumo, B.; et al. Utility of Measuring Circulating Bio-Adrenomedullin and Proenkephalin for 30-Day Mortality Risk Prediction in Patients with COVID-19 and Non-COVID-19 Interstitial Pneumonia in the Emergency Department. Medicina 2022, 58, 1852. [Google Scholar] [CrossRef]
- Garcia de Guadiana-Romualdo, L.; Calvo Nieves, M.D.; Rodriguez Mulero, M.D.; Calcerrada Alises, I.; Hernandez Olivo, M.; Trapiello Fernandez, W.; Gonzalez Morales, M.; Bolado Jimenez, C.; Albaladejo-Oton, M.D.; Fernandez Ovalle, H.; et al. MR-proADM as marker of endotheliitis predicts COVID-19 severity. Eur. J. Clin. Investig. 2021, 51, e13511. [Google Scholar] [CrossRef]
- Garcia de Guadiana-Romualdo, L.; Martinez Martinez, M.; Rodriguez Mulero, M.D.; Esteban-Torrella, P.; Hernandez Olivo, M.; Alcaraz Garcia, M.J.; Campos-Rodriguez, V.; Sancho-Rodriguez, N.; Galindo Martinez, M.; Alcaraz, A.; et al. Circulating MR-proADM levels, as an indicator of endothelial dysfunction, for early risk stratification of mid-term mortality in COVID-19 patients. Int. J. Infect. Dis. 2021, 111, 211–218. [Google Scholar] [CrossRef]
- Zaninotto, M.; Mion, M.M.; Marchioro, L.; Padoan, A.; Plebani, M. Endothelial dysfunction and Mid-Regional proAdrenomedullin: What role in SARS-CoV-2 infected Patients? Clin. Chim. Acta 2021, 523, 185–190. [Google Scholar] [CrossRef]
- Dupont, A.; Rauch, A.; Staessens, S.; Moussa, M.; Rosa, M.; Corseaux, D.; Jeanpierre, E.; Goutay, J.; Caplan, M.; Varlet, P.; et al. Vascular Endothelial Damage in the Pathogenesis of Organ Injury in Severe COVID-19. Arterioscler Thromb. Vasc. Biol. 2021, 41, 1760–1773. [Google Scholar] [CrossRef]
- Pelle, M.C.; Zaffina, I.; Luca, S.; Forte, V.; Trapanese, V.; Melina, M.; Giofre, F.; Arturi, F. Endothelial Dysfunction in COVID-19: Potential Mechanisms and Possible Therapeutic Options. Life 2022, 12, 1605. [Google Scholar] [CrossRef]
- Kita, T.; Kitamura, K. Adrenomedullin Therapy in Moderate to Severe COVID-19. Biomedicines 2022, 10, 533. [Google Scholar] [CrossRef]
Figure 1.
Calcitonin (CT), procalcitonin (PCT), and receptors of CT and PCT. The cleavage of preproCT and the release of the signal peptide produces PCT. The cleavage of PCT generates aminopro- CT, immature CT, and calcitonin carboxypeptide-1 (CCP-I). Immature CT is converted into mature CT by post-transcriptional modifications (disulfide bridge between Cys1 and Cys7 and amidation of C-terminus) and binds to the CT receptor (CTR). PCT is converted into PCT3-116, which binds to the CGRP receptor, which is composed of a heterodimer of the CTR-like receptor (CLR) and receptor activity-modifying protein 1 (RAMP1).
Figure 1.
Calcitonin (CT), procalcitonin (PCT), and receptors of CT and PCT. The cleavage of preproCT and the release of the signal peptide produces PCT. The cleavage of PCT generates aminopro- CT, immature CT, and calcitonin carboxypeptide-1 (CCP-I). Immature CT is converted into mature CT by post-transcriptional modifications (disulfide bridge between Cys1 and Cys7 and amidation of C-terminus) and binds to the CT receptor (CTR). PCT is converted into PCT3-116, which binds to the CGRP receptor, which is composed of a heterodimer of the CTR-like receptor (CLR) and receptor activity-modifying protein 1 (RAMP1).

Figure 2.
Adrenomedullin (ADM), pro-adrenomedullin N-terminal 20 peptide (PAMP), and receptors of ADM and PAMP. The cleavage of preproADM and the release of the signal peptide produce proADM. The cleavage of proADM generates mid-regional proadrenomedullin (MR-proADM), ADM, and adrenotensin. Immature ADM is converted into mature CT by post-transcriptional modifications (disulfide bridge between Cys16 and Cys21 and amidation of C-terminus) and binds to ADM receptors AM receptor 1 and AM receptor 2. PAMP is converted into PAMP-12, which binds to Mas-related G-protein coupled receptor member X2 (MRGPRX2 in humans and Mrgx2 in mice) and atypical chemokine receptor 3 (ACKR3).
Figure 2.
Adrenomedullin (ADM), pro-adrenomedullin N-terminal 20 peptide (PAMP), and receptors of ADM and PAMP. The cleavage of preproADM and the release of the signal peptide produce proADM. The cleavage of proADM generates mid-regional proadrenomedullin (MR-proADM), ADM, and adrenotensin. Immature ADM is converted into mature CT by post-transcriptional modifications (disulfide bridge between Cys16 and Cys21 and amidation of C-terminus) and binds to ADM receptors AM receptor 1 and AM receptor 2. PAMP is converted into PAMP-12, which binds to Mas-related G-protein coupled receptor member X2 (MRGPRX2 in humans and Mrgx2 in mice) and atypical chemokine receptor 3 (ACKR3).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kiriyama, Y.; Nochi, H. Procalcitonin and Adrenomedullin in Infectious Diseases. Microbiol. Res. 2023, 14, 190-204. https://doi.org/10.3390/microbiolres14010016
AMA Style
Kiriyama Y, Nochi H. Procalcitonin and Adrenomedullin in Infectious Diseases. Microbiology Research. 2023; 14(1):190-204. https://doi.org/10.3390/microbiolres14010016
Chicago/Turabian StyleKiriyama, Yoshimitsu, and Hiromi Nochi. 2023. "Procalcitonin and Adrenomedullin in Infectious Diseases" Microbiology Research 14, no. 1: 190-204. https://doi.org/10.3390/microbiolres14010016