
Hesperetin Ameliorates Inhibition of Neuronal and Oligodendroglial Cell Differentiation Phenotypes Induced by Knockdown of Rab2b, an Autism Spectrum Disorder-Associated Gene Product

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies, siRNAs, and DNA Primers
2.2. DNA Amplification
2.3. Cell Culture, Differentiation, and siRNA Transfection
2.4. Denatured Polyacrylamide Gel Electrophoresis and Immunoblotting
2.5. Statistics
2.6. Ethics
3. Results
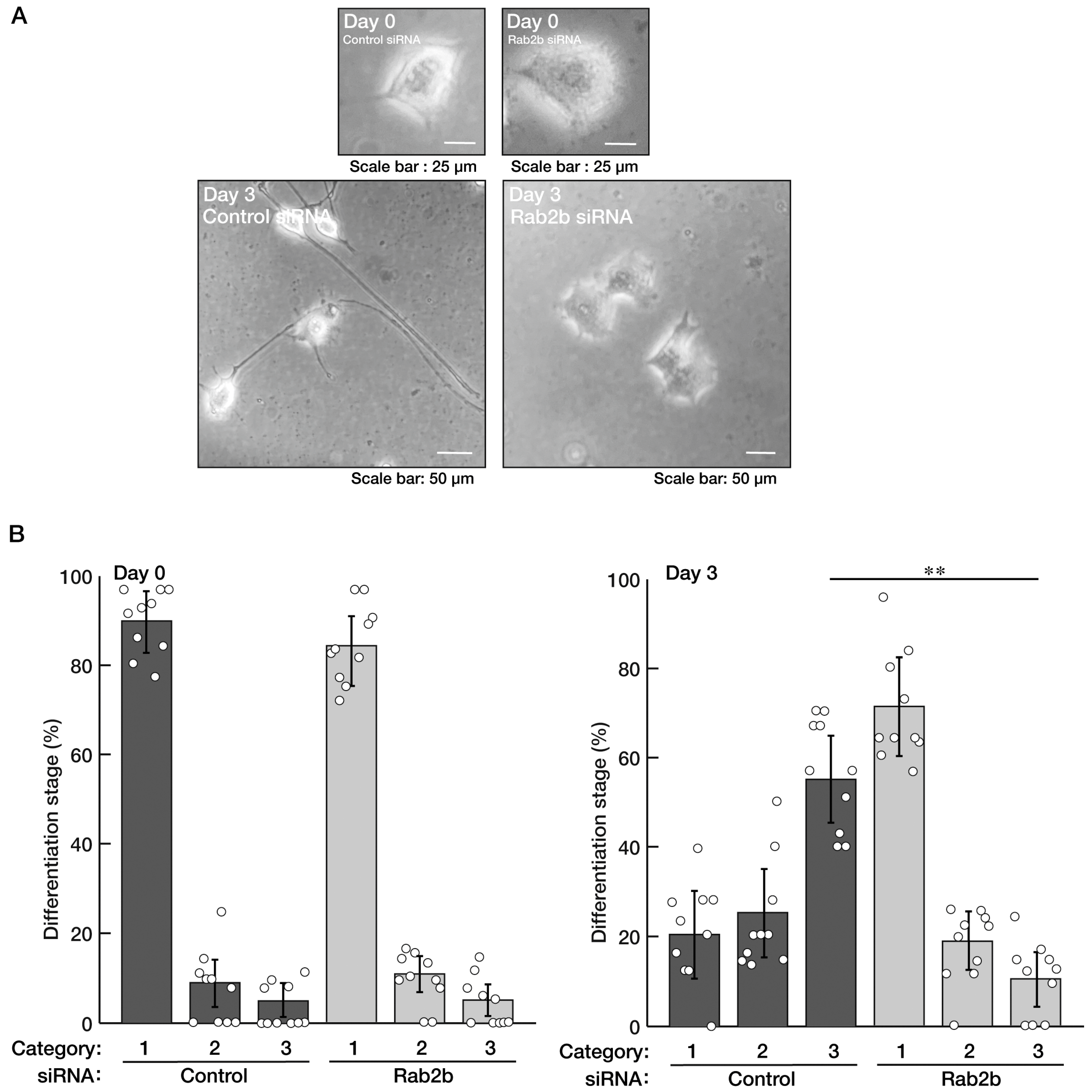
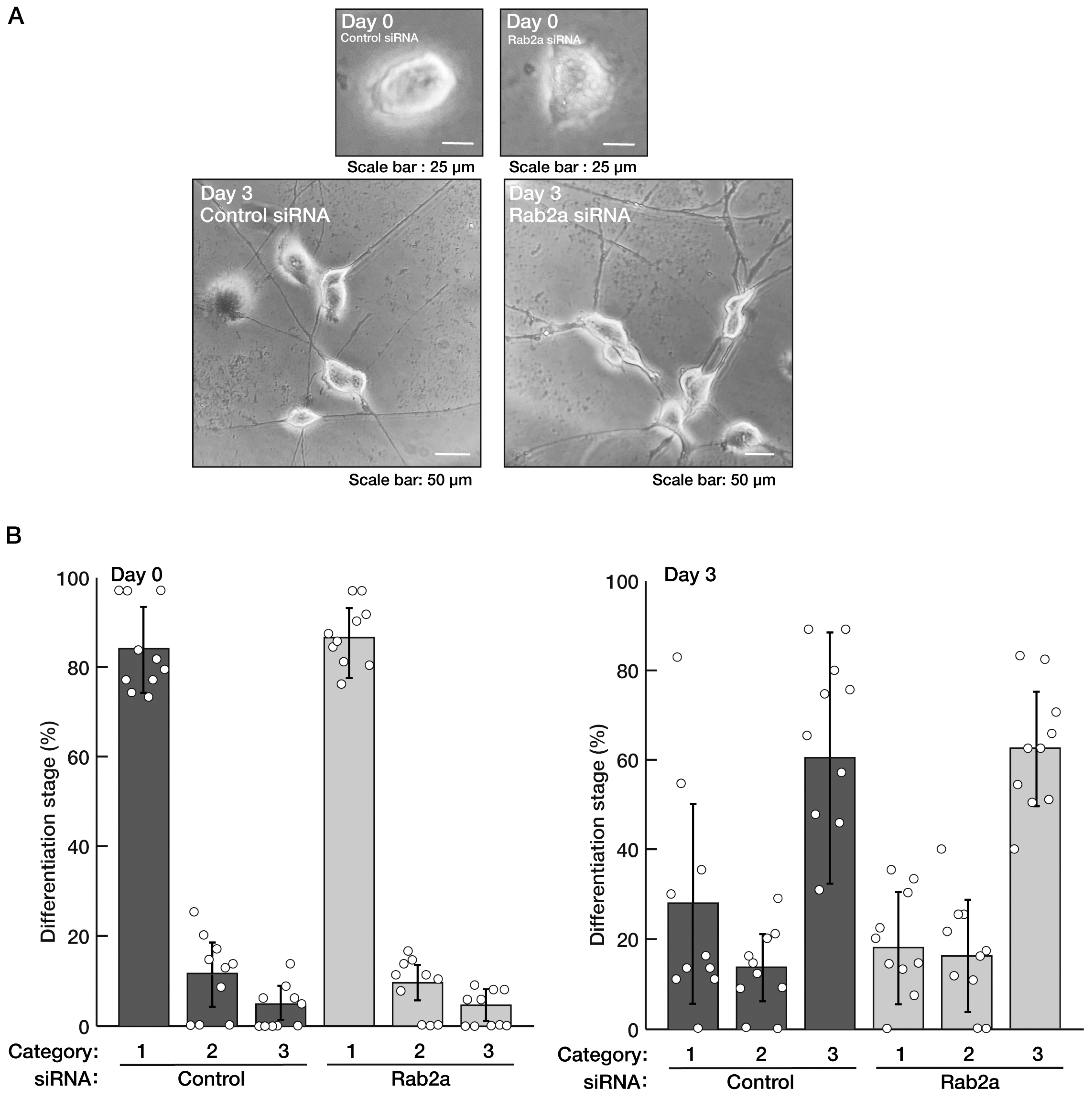
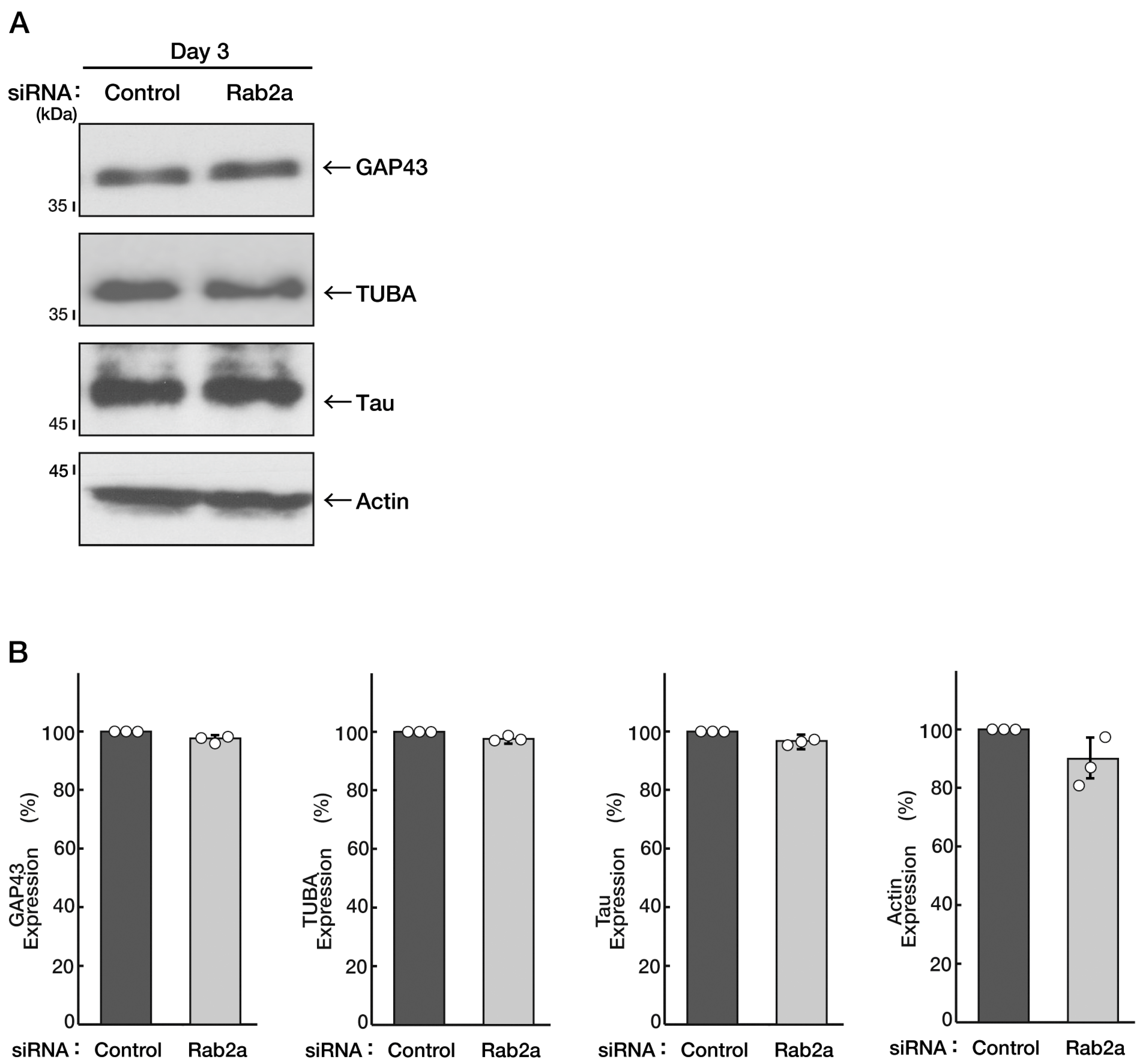
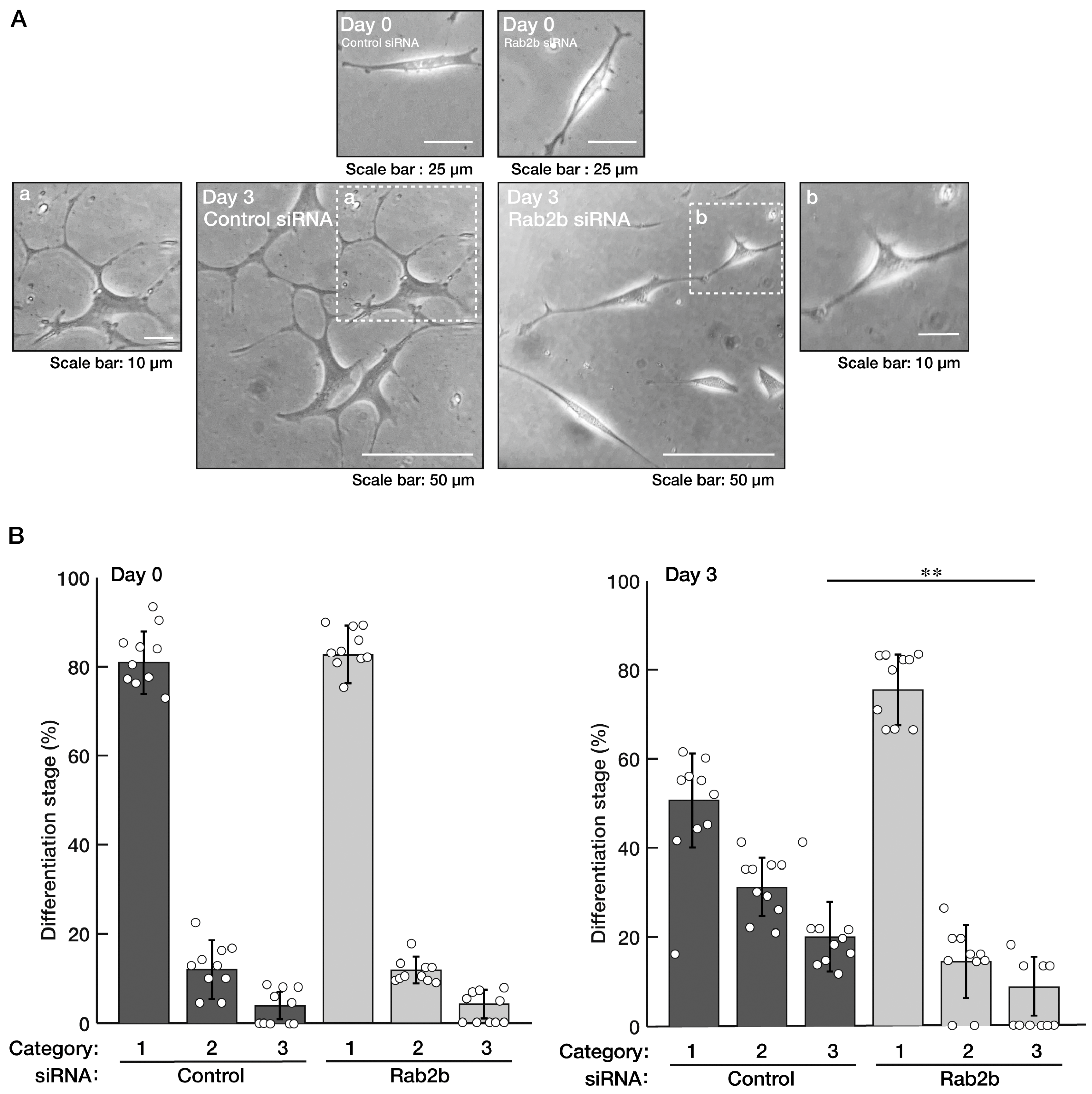
3.1. Knockdown of Rab2b but Not Rab2a Specifically Inhibits Neuronal Morphological Differentiation
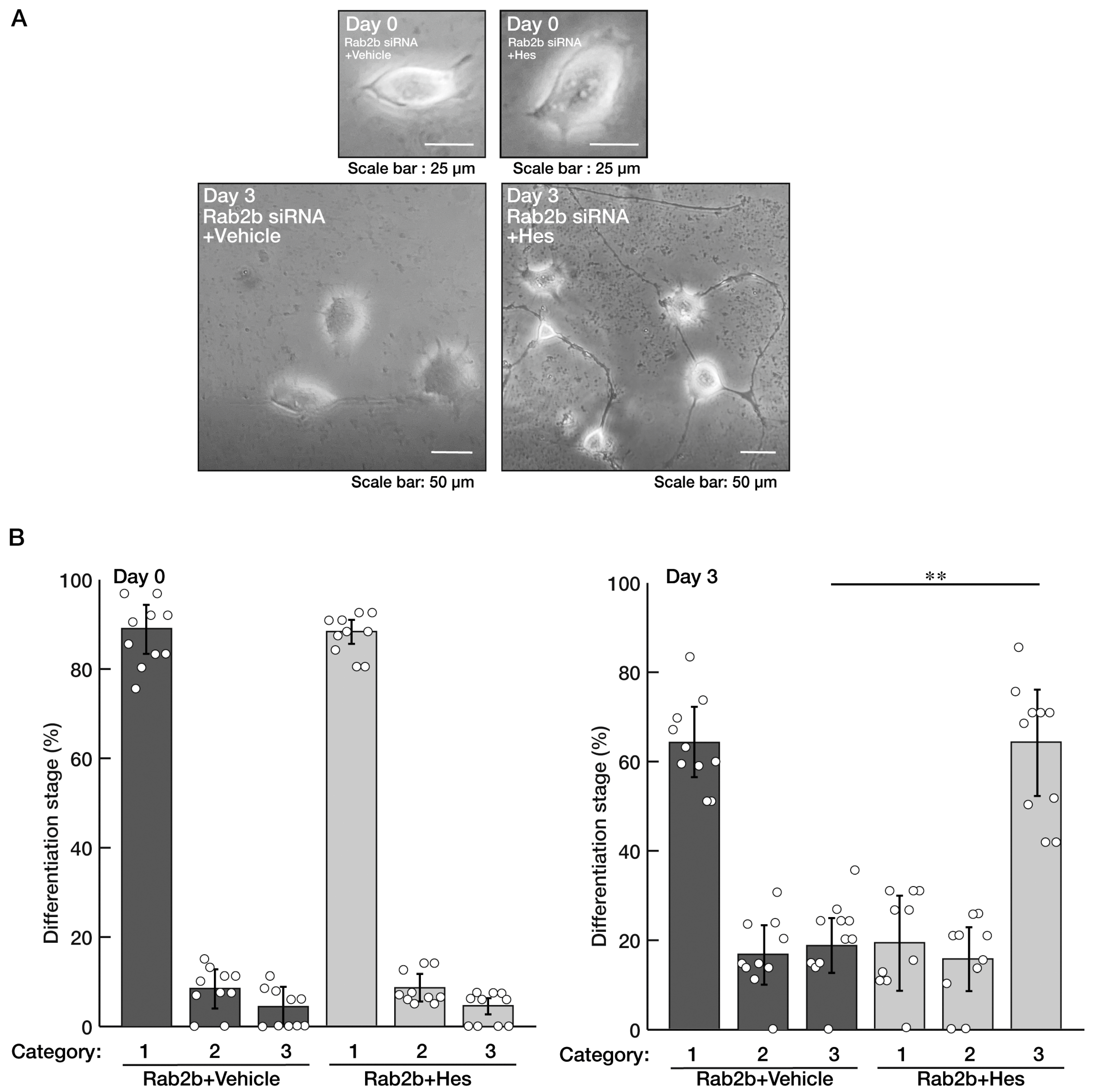
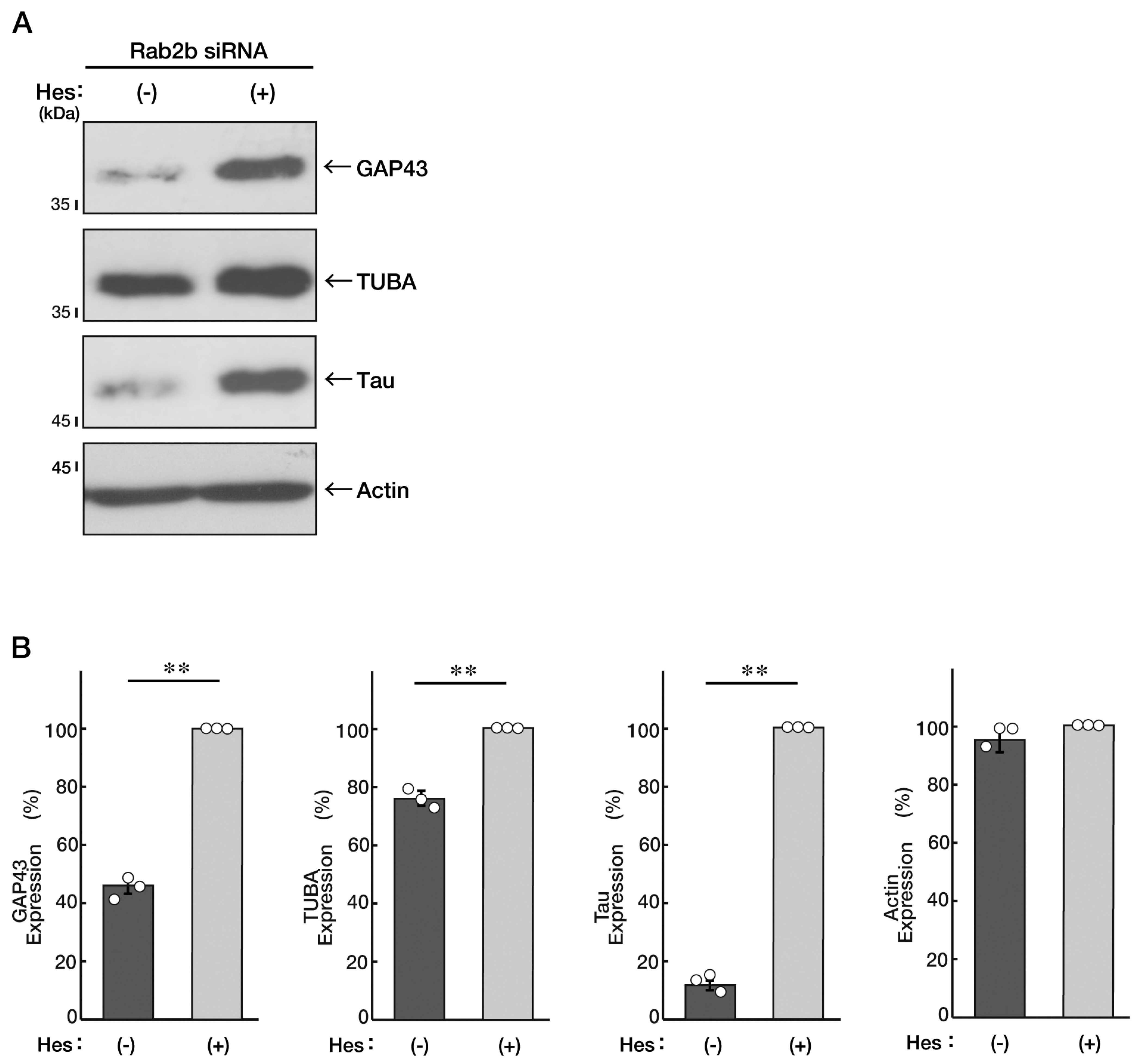
3.2. Hesperetin Recovers an Inhibitory Effect of Rab2b Knockdown on Neuronal Morphological Differentiation
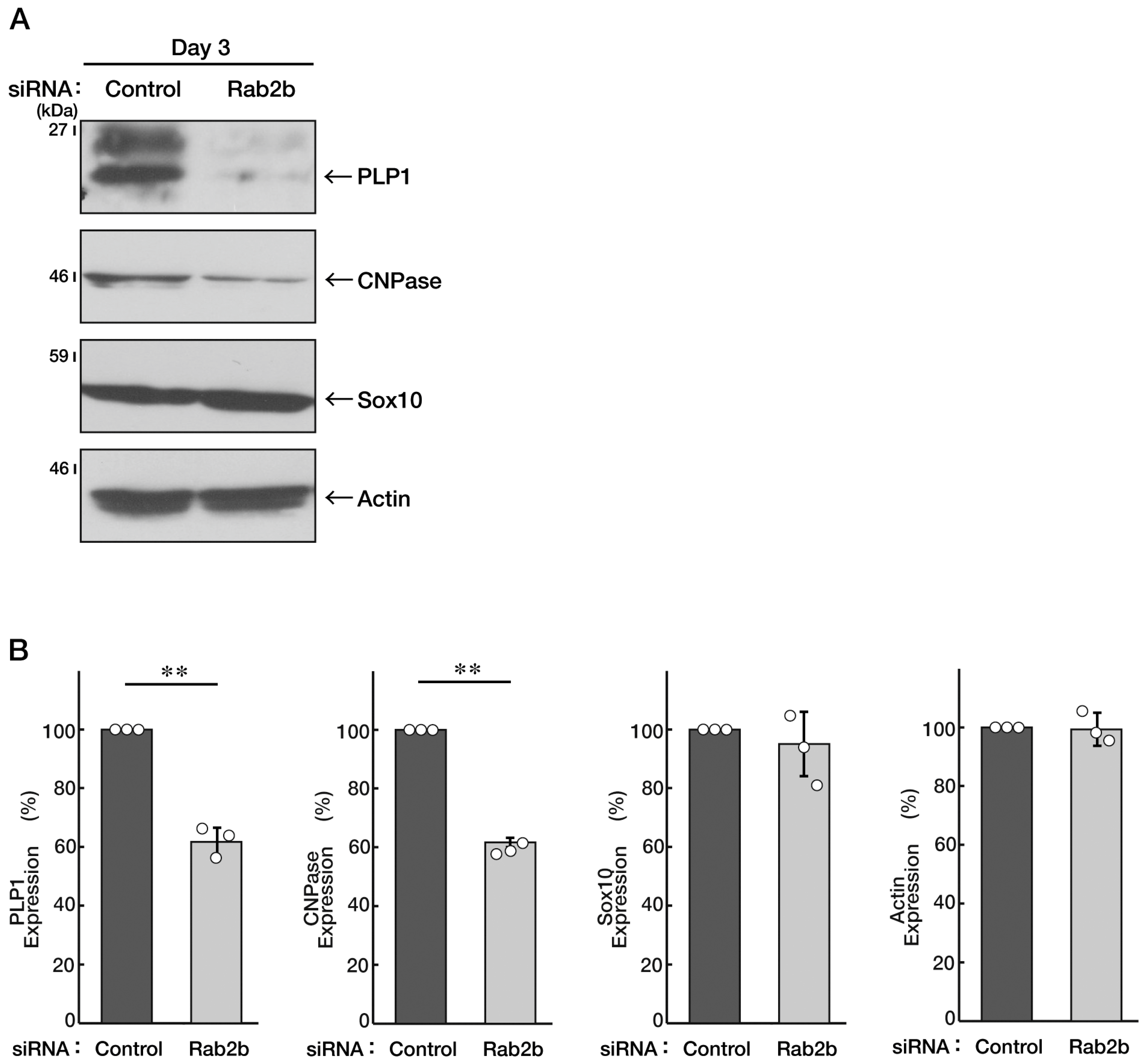
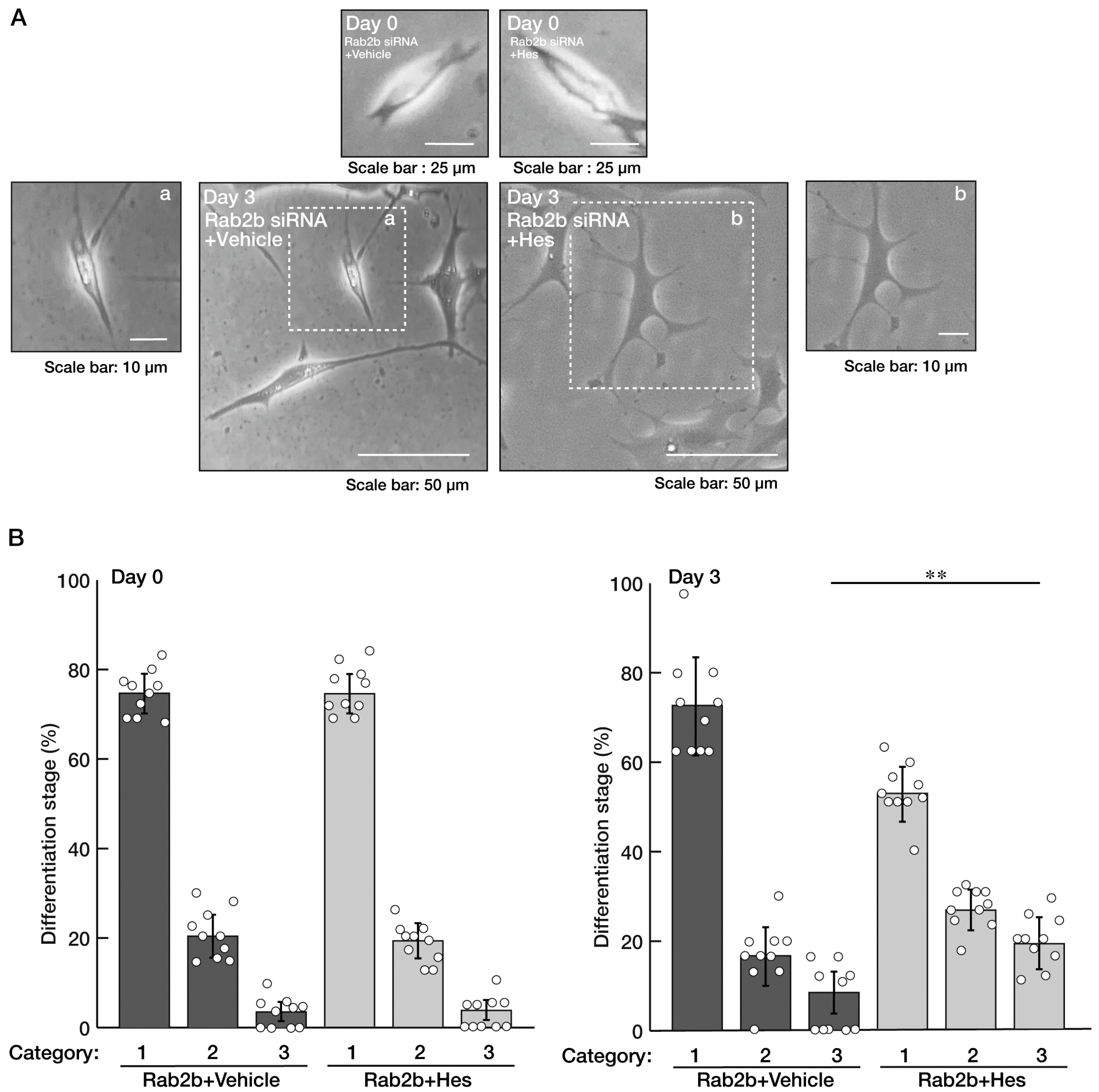
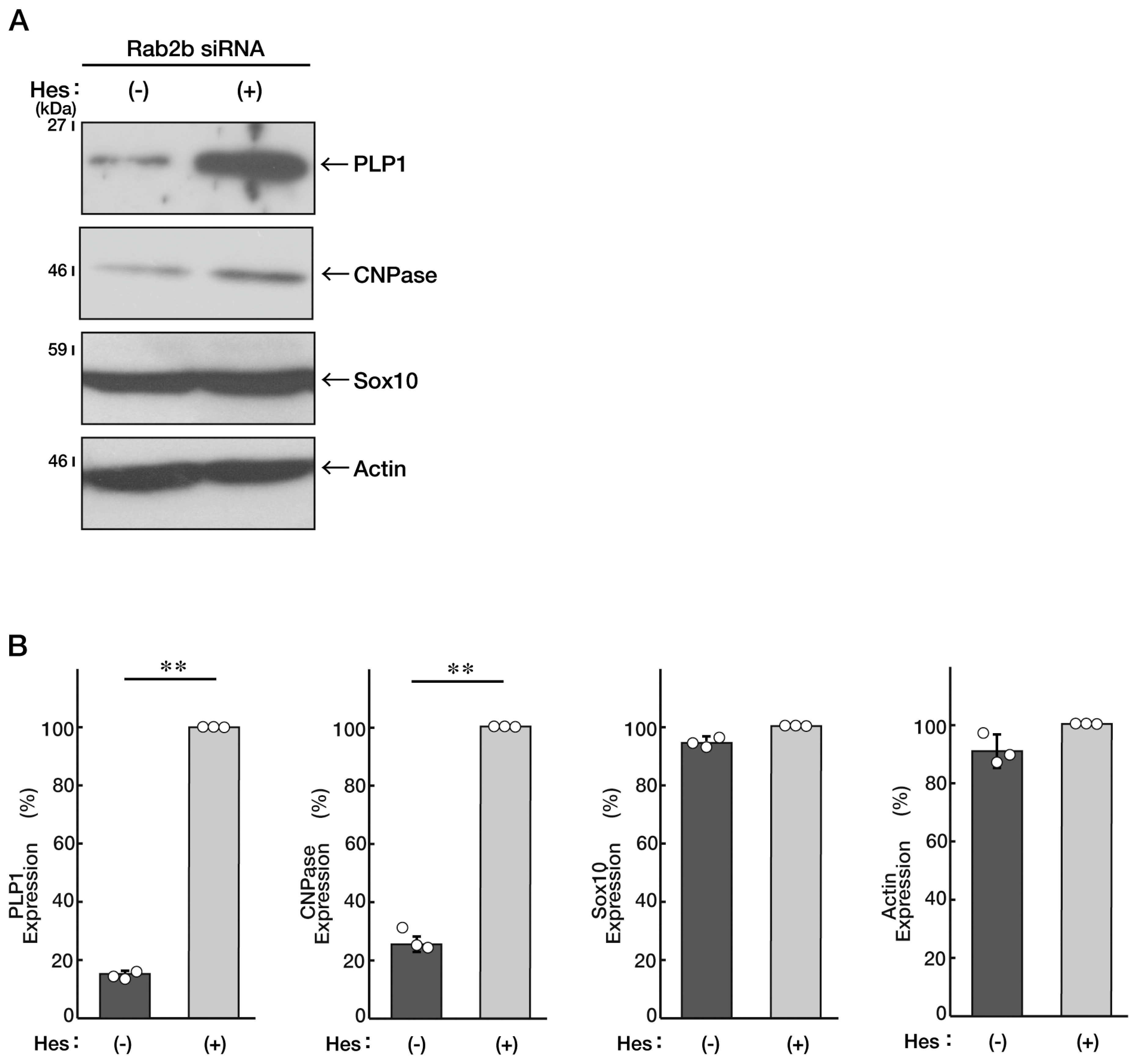
3.3. Knockdown of Rab2b Inhibits Oligodendroglial Morphological Differentiation, as Recovered by Hesperetin
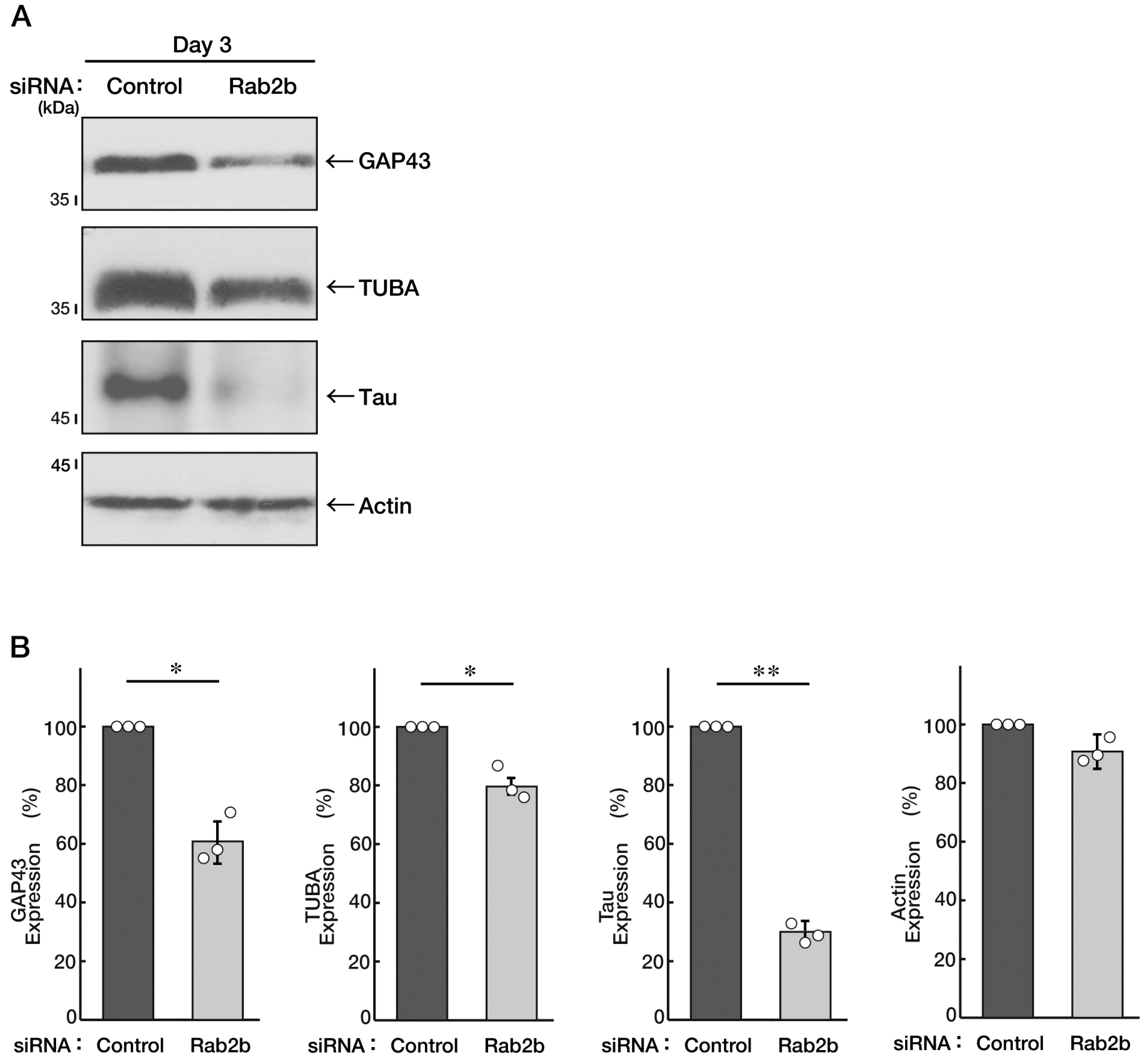
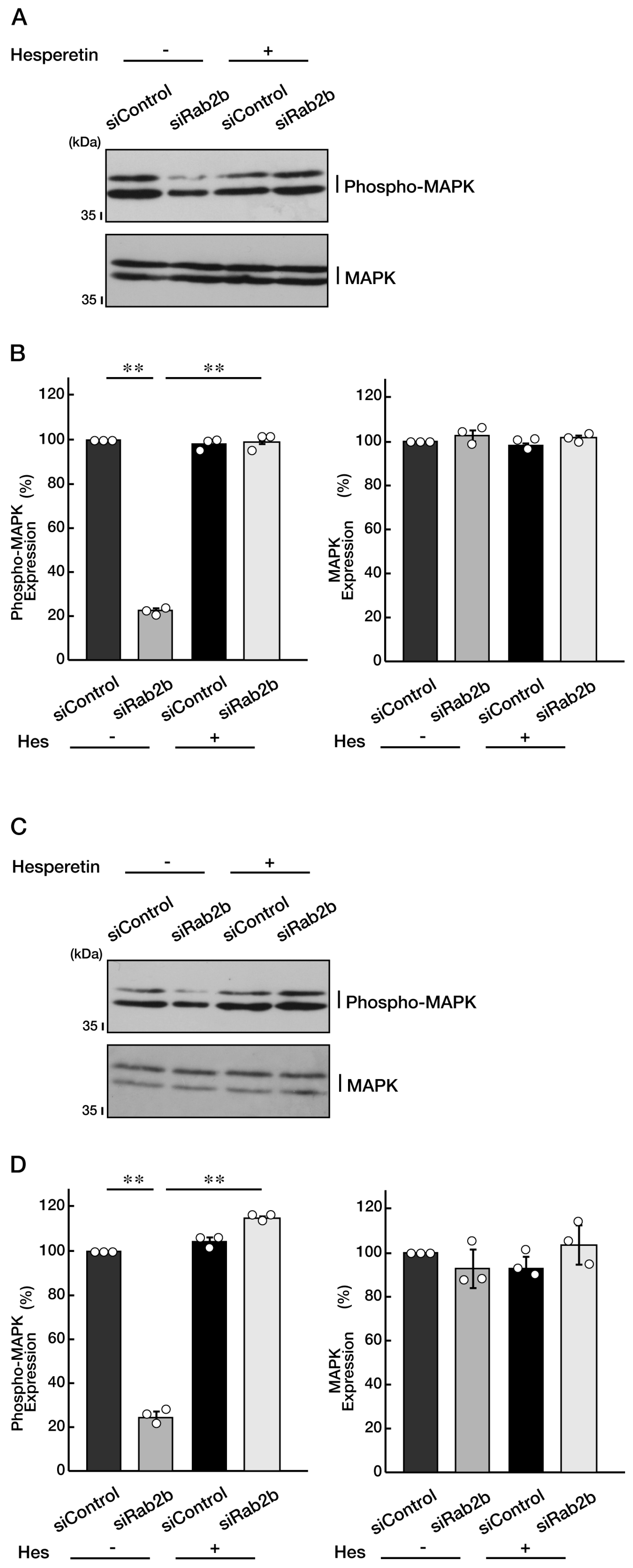
3.4. Knockdown of Rab2b Decreases Phosphorylation Levels of Mitogen-Activated Protein Kinase, as Recovered by Hesperetin
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, S.; Storrie, B. Are Rab proteins the link between Golgi organization and membrane trafficking? Cell. Mol. Life Sci. 2012, 69, 4093–4106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnár, K.; Kobolák, J.; Dinnyés, A. Golgi requires a new casting in the screenplay of mucopolysaccharidosis II cytopathology. Biol. Futur. 2022, 73, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Chun, D.K.; McEwen, J.M.; Burbea, M.; Kaplan, J.M. UNC-108/Rab2 regulates postendocytic trafficking in Caenorhabditis elegans. Mol. Biol. Cell 2008, 19, 2682–2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, V.K.; Lycas, M.D.; Schack, A.; Andersen, R.C.; Gether, U.; Kjaerulff, O. Rab2 drives axonal transport of dense core vesicles and lysosomal organelles. Cell Rep. 2021, 35, 108973. [Google Scholar] [CrossRef] [PubMed]
- Bernier, R.; Mao, A.; Yen, J. Psychopathology, families, and culture: Autism. Child Adolesc. Psychiatr. Clin. N. Am. 2010, 19, 855–867. [Google Scholar] [CrossRef]
- Pletscher-Frankild, S.; Pallejà, A.; Tsafou, K.; Binder, J.X.; Jensen, L.J. DISEASES: Text mining and data integration of disease-gene associations. Methods 2015, 74, 83–89. [Google Scholar] [CrossRef]
- Arnett, A.B.; Trinh, S.; Bernier, R.A. The state of research on the genetics of autism spectrum disorder: Methodological, clinical and conceptual progress. Curr. Opin. Psychol. 2019, 27, 1–5. [Google Scholar] [CrossRef]
- Peterson, J.L.; Earl, R.; Fox, E.A.; Ma, R.; Haidar, G.; Pepper, M.; Berliner, L.; Wallace, A.; Bernier, R. Trauma and autism spectrum disorder: Review, proposed treatment adaptations and future directions. J. Child Adolesc. Trauma 2019, 12, 529–547. [Google Scholar] [CrossRef]
- Berardis, D.D.; Filippis, S.D.; Masi, G.; Stefano Vicari Zuddas, A. A Neurodevelopment approach for a transitional model of early onset schizophrenia. Brain Sci. 2021, 11, 275. [Google Scholar] [CrossRef]
- Mazza, M.; Pino, M.C.; Mariano, M.; Tempesta, D.; Ferrana, M.; Berardis, D.D.; Masedu, F.; Valenti, M. Affective and cognitive empathy in adolescents with autism spectrum disorder. Front. Hum. Neurosci. 2014, 8, 791. [Google Scholar] [CrossRef] [Green Version]
- Hirose, M.; Ishizaki, T.; Watanabe, N.; Uehata, M.; Kranenburg, O.; Moolenaar, W.H.; Matsumura, F.; Maekawa, M.; Bito, H.; Narumiya, S. Molecular dissection of the Rho-associated protein kinase (p160ROCK)-regulated neurite remodeling in neuroblastoma N1E-115 cells. J. Cell Biol. 1998, 141, 1625–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, J.; Torii, T.; Kusakawa, S.; Sanbe, A.; Nakamura, K.; Takashima, S.; Hamasaki, H.; Kawaguchi, S.; Miyamoto, Y.; Tanoue, A. The mood stabilizer valproic acid improves defective neurite formation caused by Charcot-Marie-Tooth disease-associated mutant Rab7 through the JNK signaling pathway. J Neurosci. Res. 2010, 88, 3189–3197. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, H.; Seiwa, C.; Yoshioka, N.; Mizoguchi, K.; Yamamoto, M.; Asou, H.; Aiso, S. An extract of Chinpi, the dried peel of the citrus fruit unshiu, enhances axonal remyelination via promoting the proliferation of oligodendrocyte progenitor cells. Evid. Based Complement. Alternat. Med. 2016, 2016, 8692698. [Google Scholar] [CrossRef] [Green Version]
- Elyasi, L.; Jahanshahi, M.; Jameie, S.B.; Abadi, H.G.H.; Nikmahzar, E.; Khalili, M.; Jameie, M.; Jameie, M. 6-OHDA mediated neurotoxicity in SH-SY5Y cellular model of Parkinson disease suppressed by pretreatment with hesperidin through activating L-type calcium channels. J. Basic Clin. Physiol. Pharmacol. 2020, 32, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.H.; Ahmadnia, H.; Jelodar, S.K.; Ranjbar, M. Hesperetin nanoparticles attenuate anxiogenic-like behavior and cerebral oxidative stress through the upregulation of antioxidant enzyme expression in experimental dementia of Alzheimer’s type. Neurol. Res. 2020, 42, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, W.; Tan, R.; Xu, C.; Chen, X.; Li, S.; Liu, Y.; Qiu, H.; Cao, H.; Cheng, Q. The benefits of hesperidin in central nervous system disorders, based on the neuroprotective effect. Biomed. Pharmacother. 2023, 9, 114222. [Google Scholar] [CrossRef]
- Spinosa, M.R.; Progida, C.; De Luca, A.; Colucci, A.M.; Alifano, P.; Bucci, C. Functional characterization of Rab7 mutant proteins associated with Charcot-Marie-Tooth type 2B disease. J. Neurosci. 2008, 28, 1640–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, Y.C.; Peng, W.; Krainc, D. Lysosomal regulation of inter-mitochondrial contact fate and motility in Charcot-Marie-Tooth Type 2. Dev. Cell 2019, 50, 339–354. [Google Scholar] [CrossRef]
- Ahearn, I.M.; Haigis, K.; Bar-Sagi, D.; Philips, M.R. Regulating the regulator: Post-translational modification of RAS. Nat. Rev. Mol. Cell Biol. 2011, 13, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Cullis, J.; Das, S.; Bar-Sagi, D. K-Ras and tumor immunity: Friend or foe? Cold Spring Harb. Perspect. Med. 2018, 8, a031849. [Google Scholar] [CrossRef] [Green Version]
- Götz, T.W.B.; Puchkov, D.; Lysiuk, V.; Lützkendorf, J.; Nikonenko, A.G.; Quentin, C.; Lehmann, M.; Sigrist, S.J.; Petzoldt, A.G. Rab2 regulates presynaptic precursor vesicle biogenesis at the trans-Golgi. J. Cell Biol. 2021, 220, e202006040. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, M.; Fukuda, M. Small GTPase Rab2B and its specific binding protein Golgi-associated Rab2B interactor-like 4 (GARI-L4) regulate Golgi morphology. J. Biol. Chem. 2015, 290, 22250–22261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lőrincz, P.; Tóth, S.; Benkő, P.; Lakatos, Z.; Boda, A.; Glatz, G.; Zobel, M.; Bisi, S.; Hegedűs, K.; Takáts, S.; et al. Rab2 promotes autophagic and endocytic lysosomal degradation. J. Cell Biol. 2017, 216, 1937–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, N.; Watanabe, N.; Iibe, N.; Tatsumi, Y.; Hattori, K.; Takeuchi, Y.; Oizumi, H.; Ohbuchi, K.; Torii, T.; Miyamoto, Y.; et al. Hypomyelinating leukodystrophy-associated mutation of RARS leads it to the lysosome, inhibiting oligodendroglial morphological differentiation. Biochem. Biophys. Rep. 2019, 20, 100705. [Google Scholar] [CrossRef]
- Hattori, K.; Tago, K.; Memezawa, S.; Ochiai, A.; Sawaguchi, S.; Kato, Y.; Sato, T.; Tomizuka, K.; Ooizumi, H.; Ohbuchi, K.; et al. The infantile leukoencephalopathy-associated mutation of C11ORF73/HIKESHI proteins generates de novo interactive activity with Filamin A, inhibiting oligodendroglial cell morphological differentiation. Medicines 2021, 8, 9. [Google Scholar] [CrossRef]
- Zhang, J.; Lachance, V.; Schaffner, A.; Li, X.; Fedick, A.; Kaye, L.E.; Liao, J.; Rosenfeld, J.; Yachelevich, N.; Chu, M.L.; et al. A founder mutation in VPS11 causes an autosomal recessive leukoencephalopathy linked to autophagic defects. PLoS Genet. 2016, 12, e1005848. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, M.; Leimgruber, E.; Seguín-Estévez, Q.; Dunand-Sauthier, I.; Barras, E.; Reith, W. Expression of RAB4B, a protein governing endocytic recycling, is co-regulated with MHC class II genes. Nucleic Acids Res. 2007, 35, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Perrin, L.; Lacas-Gervais, S.; Gilleron, J.; Ceppo, F.; Prodon, F.; Benmerah, A.; Tanti, J.F.; Cormont, M. Rab4b controls an early endosome sorting event by interacting with the γ-subunit of the clathrin adaptor complex 1. J. Cell Sci. 2013, 126, 4950–4962. [Google Scholar] [CrossRef] [Green Version]
- Gilleron, J.; Bouget, G.; Ivanov, S.; Meziat, C.; Ceppo, F.; Vergoni, B.; Djedaini, M.; Soprani, A.; Dumas, K.; Jacquel, A.; et al. Rab4b deficiency in T cells promotes adipose Treg/Th17 imbalance, adipose tissue dysfunction, and insulin resistance. Cell Rep. 2018, 25, 3329–3341. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.B.; Dean, N.; Gao, X.D.; Fujita, M. MON2 guides wntless transport to the Golgi through recycling endosomes. Cell Struct. Funct. 2020, 45, 77–92. [Google Scholar] [CrossRef]
- Hu, Y.; Li, Y.; Huang, Y.; Jin, Z.; Wang, C.; Wang, H.; Xu, J. METTL3 regulates the malignancy of cervical cancer via post-transcriptional regulation of RAB2B. Eur. J. Pharmacol. 2020, 879, 173134. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Wu, Y.; Zhou, D.; Sun, Q.; Wang, W. miR-448 targets Rab2B and is pivotal in the suppression of pancreatic cancer. Oncol. Rep. 2018, 40, 1379–1389. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 2010, 1802, 396–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.R.; Gonda, X.; Tarazi, F.I. Autism spectrum disorder: Classification, diagnosis and therapy. Pharmacol. Ther. 2018, 35, 91–104. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent or Source | Company or Source | Cat. No. | Lot. No. | Concentration Used |
|---|---|---|---|---|
| Antibodies | ||||
| Anti-proteolipid protein 1 (PLP1) | Atlas Antibodies (Stockholm, Sweden) | HPA004128 | B115828 | Immunoblotting (IB), 1/500 |
| Anti-2′, 3′-cyclic nucleotide 3′-phosphodiesterase (CNPase) | Santa Cruz Biotechnology (Santa Cruz, CA, USA) | sc-166559 | F1514 | IB, 1/500 |
| Anti-SRY-related HMG-box protein 10 (SOX10) | Santa Cruz Biotechnology | sc-365692 | F1621 | IB, 1/100 |
| Anti-Actin (pan-b type) | MBL (Tokyo, Japan) | M177-3 | 008 | IB, 1/5000 |
| Anti-growth-associated protein 43 (GAP43) | Santa Cruz Biotechnology | sc-17790 | J0920 | IB, 1/8500 |
| Anti-Tubulin a chain 1a (TUBA1A, also called TUBA) | MBL | M175-3 | 001 | IB, 1/500,000 |
| Anti-Tau | Santa Cruz Biotechnology | sc-21796 | H2721 | IB, 1/250 |
| Anti-IgG (H+L chain) (mouse) pAb-HRP | MBL | 330 | 366 | IB, 1/5000 |
| Anti-IgG (H+L chain) (rabbit) pAb-HRP | MBL | 458 | 353 | IB, 1/5000 |
| Reagent or Source | Company or Source | Cat. No. | Lot. No. | Concentration Used |
|---|---|---|---|---|
| Key Chemicals | ||||
| Hesperetin | Santa Cruz Biotechnology | sc-202647 | D1921 | 1 or 10 μM |
| Dimethyl sulfoxide (DMSO, control vehicle) | FUJIFILM Wako Pure Chemical Corporation (Tokyo, Japan) | 047-29353 | CDN0170 | 0.10% |
| Reagent or Source | Company or Source | Cat. No. | Lot. No. | Concentration Used |
|---|---|---|---|---|
| Key Reagents | ||||
| ScreenFect TM siRNA Transfection Reagent | FUJIFILM Wako Pure Chemical Corporation | 292-75013 | CAM0357 | According to manufacturer’s instructions |
| ScreenFect TM Dilution Buffer | FUJIFILM Wako Pure Chemical Corporation | 194-18181 | SKF5794 | According to manufacturer’s instructions |
| ImmunoStar Zeta | FUJIFILM Wako Pure Chemical Corporation | 295-72404 | LEN3912 | According to manufacturer’s instructions |
| Chemi-Lumi One Ultra | Nacalai Tesque (Kyoto, Japan) | 11644-24 | L1P7669 | According to manufacturer’s instructions |
| Skim Milk Powder | FUJIFILM Wako Pure Chemical Corporation | 190-12865 | SKG4901 | According to manufacturer’s instructions |
| Western Blotting (WB) Stripping Solution | Nacalai Tesque | 05364-55 | L5M5218 | According to manufacturer’s instructions |
| Gflex DNA Polymerase | TaKaRa Bio (Shiga, Japan) | R060A | AL80564A | According to manufacturer’s instructions |
| 2×Gflex PCR Buffer (Mg2+, dNTP plus) | TaKaRa Bio | R060A | AL80564A | According to manufacturer’s instructions |
| ISOGEN | Nippon Gene (Tokyo, Japan) | 311-02501 | 75009K | According to manufacturer’s instructions |
| Sample Buffer Solution (plus 2-Mercaptoethanol) (4-fold mixtures) | FUJIFILM Wako Pure Chemical Corporation | 191-13272 | WDP4995 | According to manufacturer’s instructions |
| Pre-stained Protein Markers (Broad Range) for SDS-PAGE | Nacalai Tesque | 02525-35 | L9M9989 | According to manufacturer’s instructions |
| 5×Prime Script Master Mix | TaKaRa Bio | RR036A | AIE0440A | According to manufacturer’s instructions |
| Reagent or Source | Company or Source | Cat. No. | Lot. No. | Concentration Used |
|---|---|---|---|---|
| Cell Lines | ||||
| FBD-102b cells (mouse cells) | Dr. Yasuhiro Tomo-oka (Riken, Saitama, Japan and Tokyo University of Science, Chiba, Japan) | N/A | N/A | N/A |
| N1E-115 cells (mouse cells) | Dr. Daisuke Shiokawa (Tokyo Science University, Chiba, Japan) | N/A | N/A | N/A |
| Reagent or Source | Company or Source | Cat. No. | Lot. No. | Concentration Used |
|---|---|---|---|---|
| DNA Primers (5′ to 3′) | ||||
| Sense primer for actin (internal control) ATGGATGACGATATCGCTGCGCTGGTC Antisense primer for actin (internal control) CTAGAAGCACTTGCGGTGCACGATGGAG | This manuscript | N/A | N/A | 0.5 μM |
| Sense primer for Rab2a ATGGCGTACGCCTATCTCTTCAAGT Antisense primer for Rab2a TCAACAGCAGCCTCCCCCT | This manuscript | N/A | N/A | 0.5 μM |
| Sense primer for Rab2b ATGACTTACGCTTATCTCTTCAAGTACATCATCATC Antisense primer for Rab2b TCAGCAGCAGCCAGAGTCAG | This manuscript | N/A | N/A | 0.5 μM |
| Sense primer for Rab4a CCGGGATCCATGGCGCAGACCGCCATG Antisense primer for Rab4a CCGGGATCCCTAGCAGCCACACTCCTGTGCAC | This manuscript | N/A | N/A | 0.5 μM |
| Sense primer for Rab4b CCGAGATCTATGGCCGAGACCTACGACTTCCTC Antisense primer for Rab4b CCGAGATCTTCAGCAGCCACAGGGCTGAG | This manuscript | N/A | N/A | 0.5 μM |
| Reagent or Source | Company or Source | Cat. No. | Lot. No. | Concentration Used |
|---|---|---|---|---|
| siRNA Sequences (5′ to 3′) | ||||
| Sense chain for siLuciferase (Control siRNA) GCCAUUCUAUCCUCUAGAG-dTdT Antisense chain for siLuciferase (Control siRNA) CUCUAGAGGAUAGAAUGGC-dTdT | This manuscript | N/A | N/A | N/A |
| Sense chain for siRab2a-87th GAGGUUUCAGCCGGUGCAU-dTdT Antisense chain for siRab2a-87th AUGCACCGGCUGAAACCUC-dTdT | This manuscript | N/A | N/A | N/A |
| Sense chain for siRab2a-269th GGAGAGACACGUUCAACCA-dTdT Antisense chain for siRab2a-269th UGGUUGAACGUGUCUCUCC-dTdT | This manuscript | N/A | N/A | N/A |
| Sense chain for siRab2b-87th GCGGUUCCAGCCCGUGCAC-dTdT Antisense chain for siRab2b-87th GUGCACGGGCUGGAACCGC-dTdT | This manuscript | N/A | N/A | N/A |
| Sense chain for siRab2b-196th GAGUCCUUCCGUUCUAUCA-dTdT Antisense chain for siRab2b-196th UGAUAGAACGGAAGGACUC-dTdT | This manuscript | N/A | N/A | N/A |
| Sense chain for siRab4b-96th GUUCAAACAGGACUCCAAC-dTdT Antisense chain for siRab4b-96th GUUGGAGUCCUGUUUGAAC-dTdT | This manuscript | N/A | N/A | N/A |
| Sense chain for siRab4b-165th GACUGUGAAACUACAGAUU-dTdT Antisense chain for siRab4b-165th AAUCUGUAGUUUCACAGUC-dTdT | This manuscript | N/A | N/A | N/A |
| Sense chain for siRab4b-366th GAAGGACCUGGAUCCCGAG-dTdT Antisense chain for siRab4b-366th CUCGGGAUCCAGGUCCUUC-dTdT | This manuscript | N/A | N/A | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato, Y.; Shirai, R.; Ohbuchi, K.; Oizumi, H.; Yamamoto, M.; Miyata, W.; Iguchi, T.; Mimaki, Y.; Miyamoto, Y.; Yamauchi, J. Hesperetin Ameliorates Inhibition of Neuronal and Oligodendroglial Cell Differentiation Phenotypes Induced by Knockdown of Rab2b, an Autism Spectrum Disorder-Associated Gene Product. Neurol. Int. 2023, 15, 371-391. https://doi.org/10.3390/neurolint15010025
Kato Y, Shirai R, Ohbuchi K, Oizumi H, Yamamoto M, Miyata W, Iguchi T, Mimaki Y, Miyamoto Y, Yamauchi J. Hesperetin Ameliorates Inhibition of Neuronal and Oligodendroglial Cell Differentiation Phenotypes Induced by Knockdown of Rab2b, an Autism Spectrum Disorder-Associated Gene Product. Neurology International. 2023; 15(1):371-391. https://doi.org/10.3390/neurolint15010025
Chicago/Turabian StyleKato, Yukino, Remina Shirai, Katsuya Ohbuchi, Hiroaki Oizumi, Masahiro Yamamoto, Wakana Miyata, Tomoki Iguchi, Yoshihiro Mimaki, Yuki Miyamoto, and Junji Yamauchi. 2023. "Hesperetin Ameliorates Inhibition of Neuronal and Oligodendroglial Cell Differentiation Phenotypes Induced by Knockdown of Rab2b, an Autism Spectrum Disorder-Associated Gene Product" Neurology International 15, no. 1: 371-391. https://doi.org/10.3390/neurolint15010025