
Protein Metabolism Changes and Alterations in Behavior of Trace Amine-Associated Receptor 1 Knockout Mice Fed a High-Fructose Diet
 , , ,
, , ,  , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Study Design
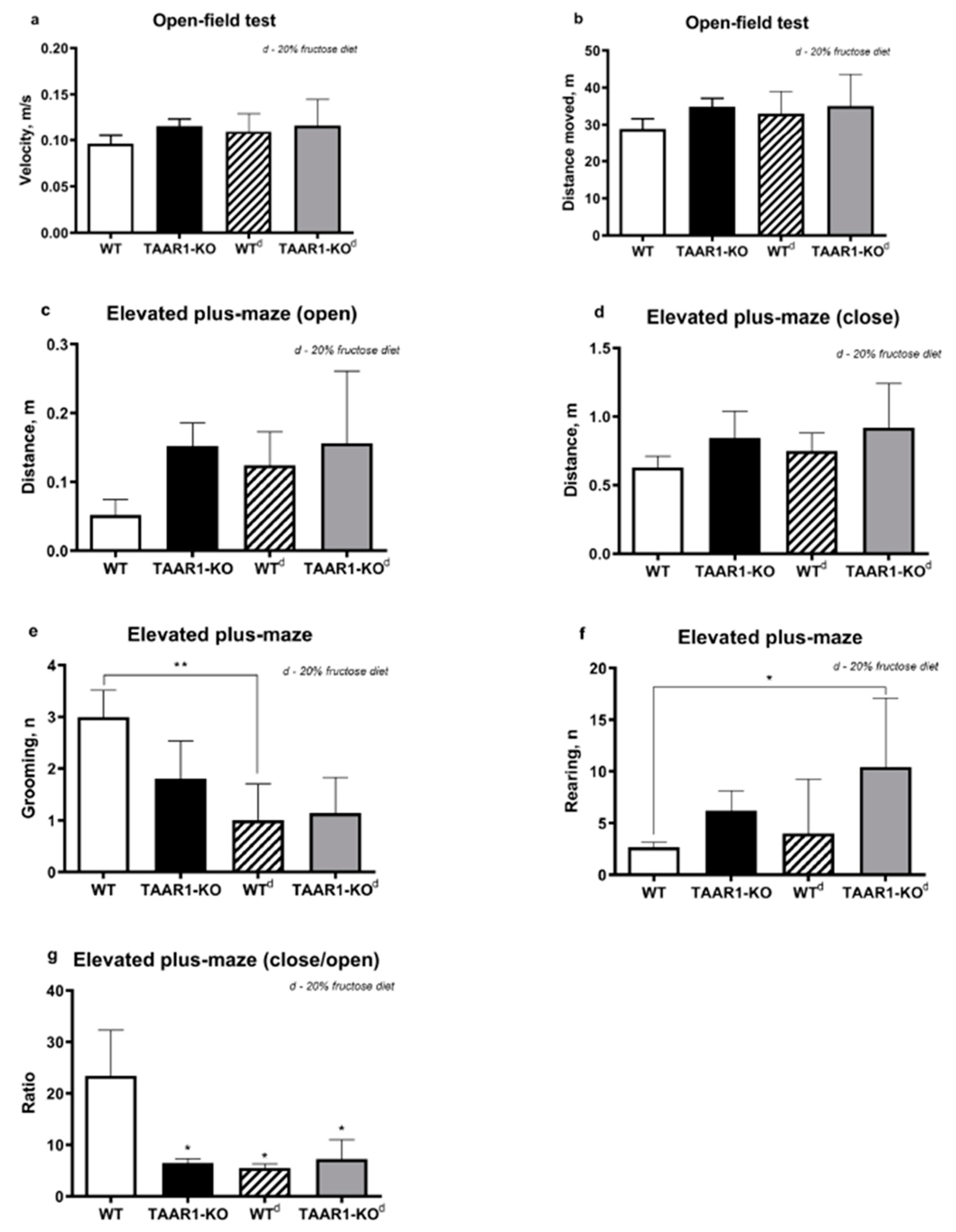
2.2. Behavioral Tests
2.2.1. Open Field Test (OF)
2.2.2. Elevated Plus Maze Test (EPM)
2.2.3. Grooming Microstructure Analysis
2.3. Measurement of Biochemical Parameters
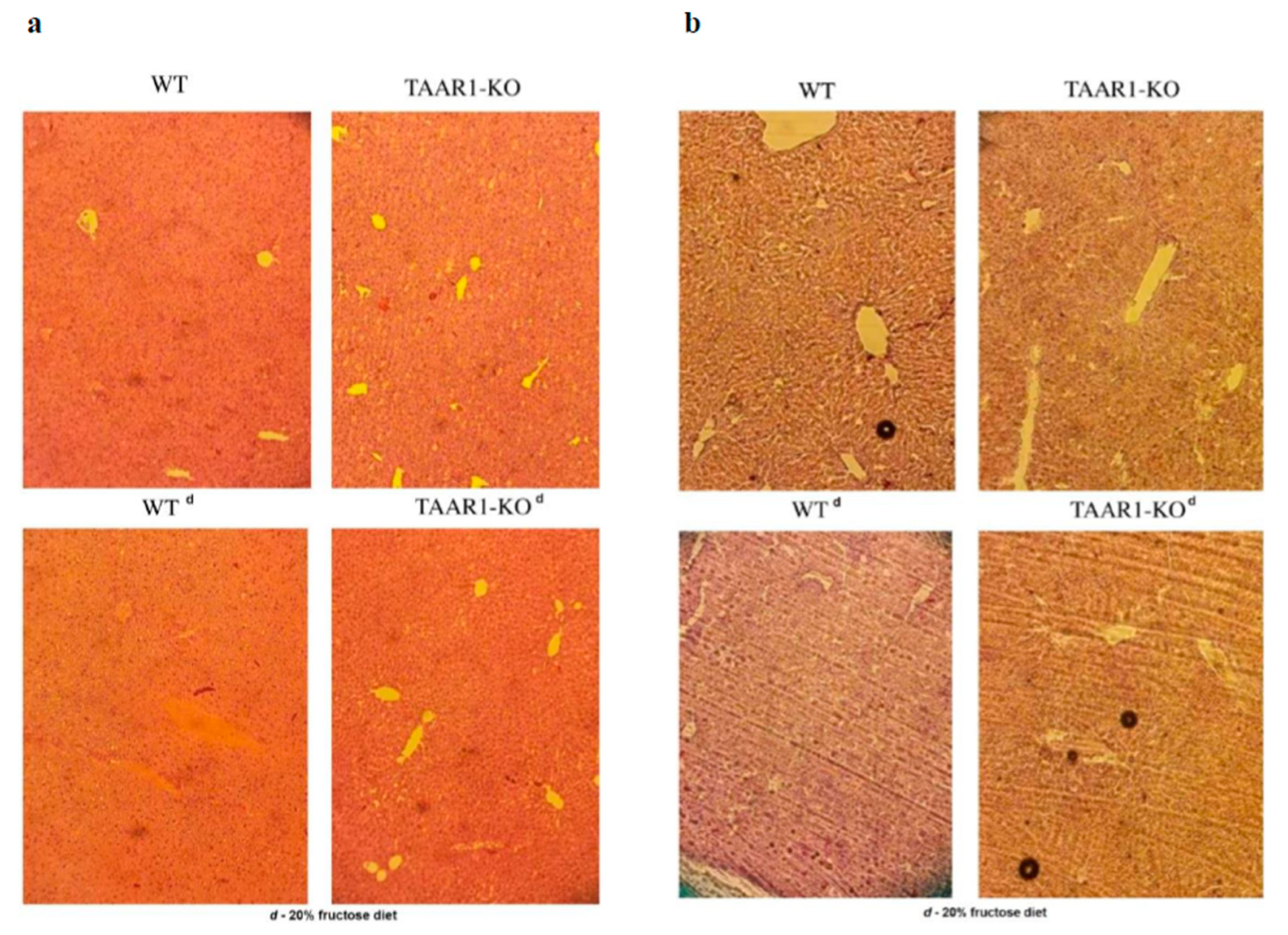
2.4. Histological Analysis
2.5. Statistical Analysis
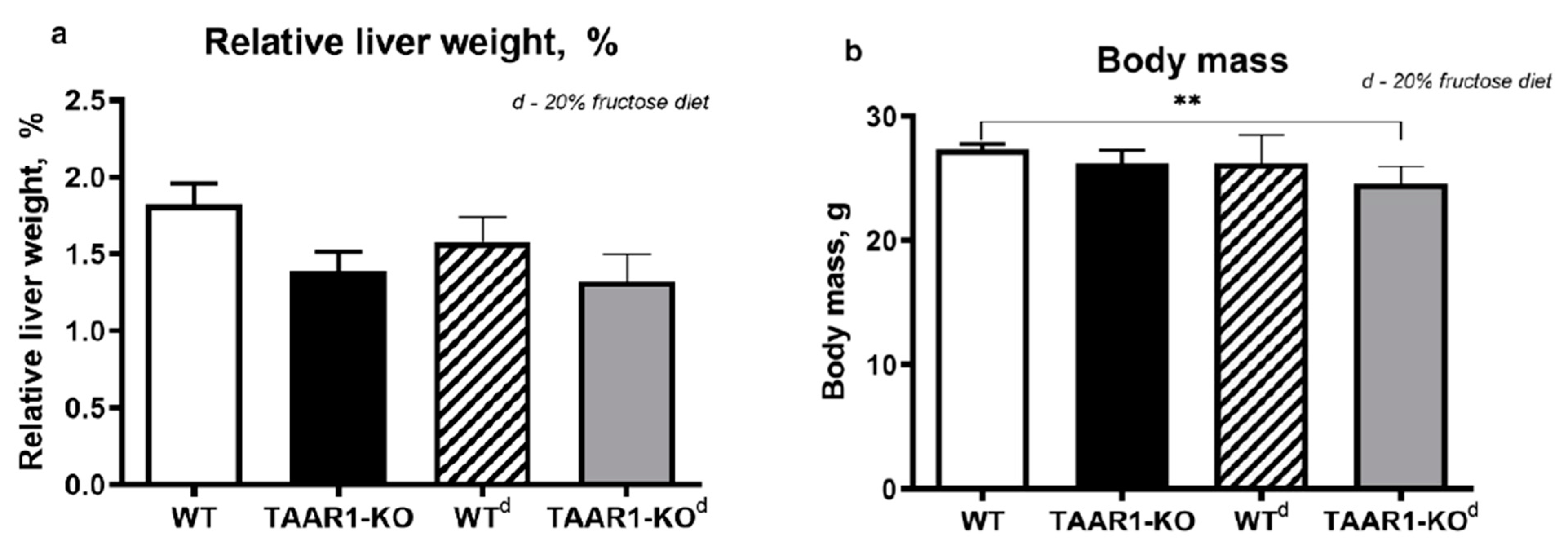
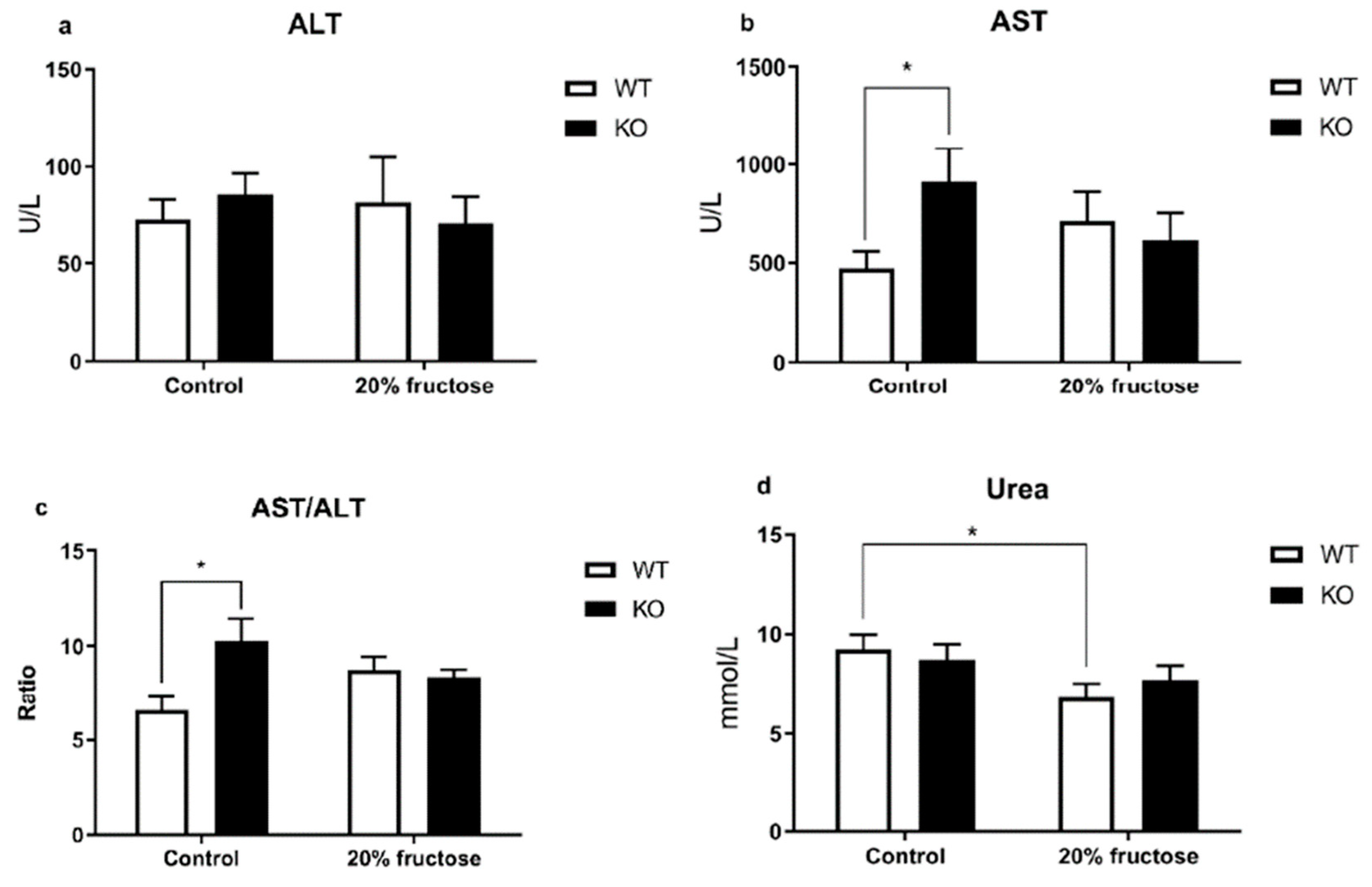
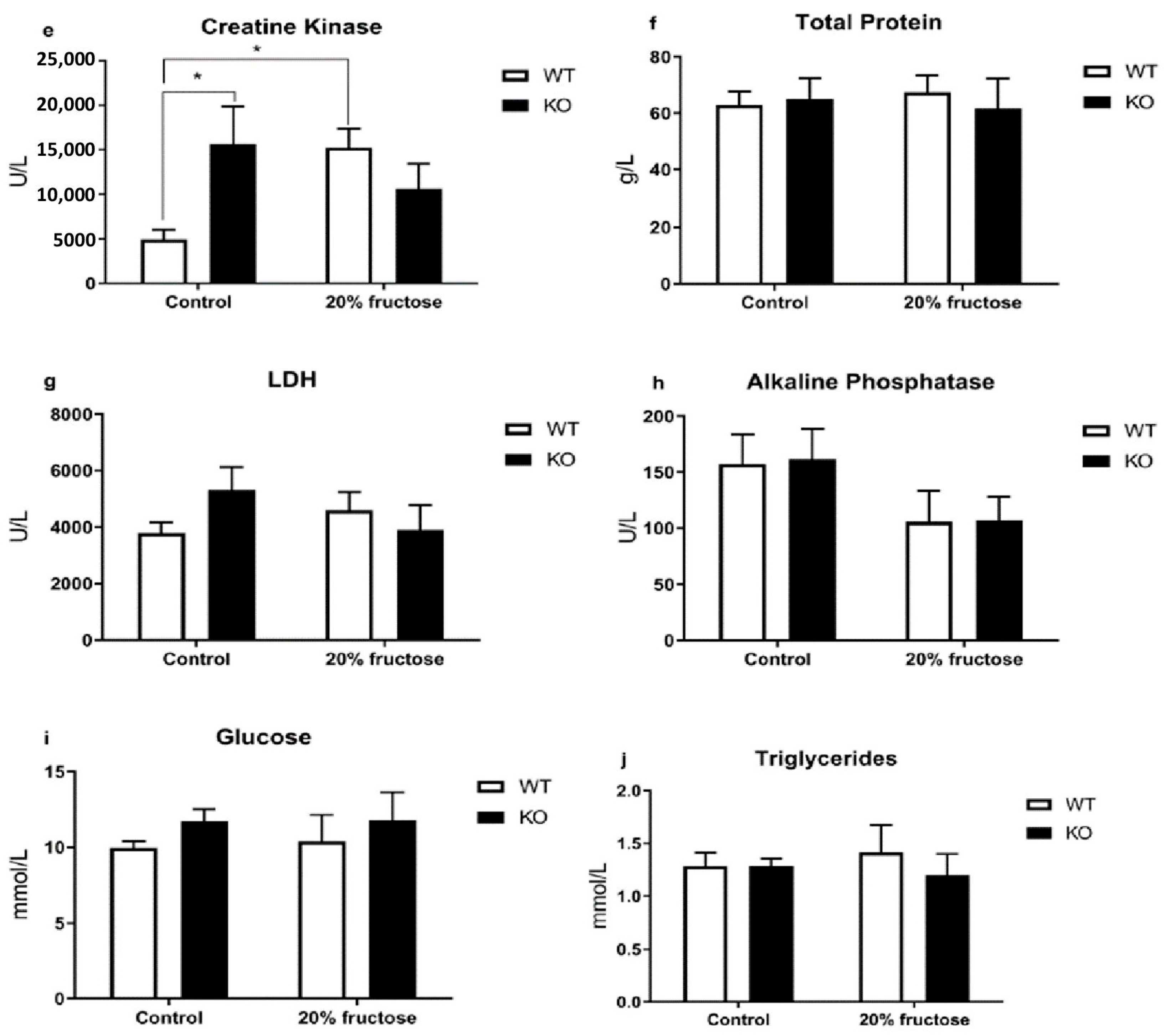
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berry, M.D. Mammalian central nervous system trace amines. Pharmacologic amphetamines, physiologic neuromodulators. J. Neurochem. 2004, 90, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace amines and their receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef] [PubMed]
- Boulton, A. Amines and theories in psychiatry. Lancet 1974, 304, 52–53. [Google Scholar] [CrossRef] [PubMed]
- Burchett, S.A.; Hicks, T.P. The mysterious trace amines: Protean neuromodulators of synaptic transmission in mammalian brain. Prog. Neurobiol. 2006, 79, 223–246. [Google Scholar] [CrossRef]
- Borowsky, B.; Adham, N.; Jones, K.A.; Raddatz, R.; Artymyshyn, R.; Ogozalek, K.L.; Durkin, M.M.; Lakhlani, P.P.; Bonini, J.A.; Pathirana, S.; et al. Trace amines: Identification of a family of mammalian G protein-coupled receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 8966–8971. [Google Scholar] [CrossRef]
- Bunzow, J.R.; Sonders, M.S.; Arttamangkul, S.; Harrison, L.M.; Zhang, G.; Quigley, D.I.; Darland, T.; Suchland, K.L.; Pasumamula, S.; Kennedy, J.L.; et al. Amphetamine, 3,4-methylenedioxymethamphetamine, lysergic acid diethylamide, and metabolites of the catecholamine neurotransmitters are agonists of a rat trace amine receptor. Mol. Pharmacol. 2001, 60, 1181–1188. [Google Scholar] [CrossRef]
- Lindemann, L.; Ebeling, M.; Kratochwil, N.A.; Bunzow, J.R.; Grandy, D.K.; Hoener, M.C. Trace amine-associated receptors form structurally and functionally distinct subfamilies of novel G protein-coupled receptors. Genomics 2005, 85, 372–385. [Google Scholar] [CrossRef]
- Lindemann, L.; Meyer, C.A.; Jeanneau, K.; Bradaia, A.; Ozmen, L.; Bluethmann, H.; Bettler, B.; Wettstein, J.G.; Borroni, E.; Moreau, J.-L.; et al. Trace Amine-Associated Receptor 1 Modulates Dopaminergic Activity. J. Pharmacol. Exp. Ther. 2008, 324, 948–956. [Google Scholar] [CrossRef]
- Leo, D.; Mus, L.; Espinoza, S.; Hoener, M.; Sotnikova, T.; Gainetdinov, R. Taar1-mediated modulation of presynaptic dopaminergic neurotransmission: Role of D2 dopamine autoreceptors. Neuropharmacology 2014, 81, 283–291. [Google Scholar] [CrossRef]
- Premont, R.T.; Gainetdinov, R.R.; Caron, M.G. Following the trace of elusive amines. Proc. Natl. Acad. Sci. USA 2001, 98, 9474–9475. [Google Scholar] [CrossRef]
- Khan, M.Z.; Nawaz, W. The emerging roles of human trace amines and human trace amine-associated receptors (hTAARs) in central nervous system. Biomed. Pharmacother. 2016, 83, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, W.; Mohammed, G.I.; Al-Eryani, D.A.; Saigl, Z.M.; Alyoubi, A.O.; Alwael, H.; Bashammakh, A.S.; O’Sullivan, C.K.; El-Shahawi, M.S. Biogenic Amines Formation Mechanism and Determination Strategies: Future Challenges and Limitations. Crit. Rev. Anal. Chem. 2020, 50, 485–500. [Google Scholar] [CrossRef] [PubMed]
- Leo, D.; Targa, G.; Espinoza, S.; Villers, A.; Gainetdinov, R.R.; Ris, L. Trace Amine Associate Receptor 1 (TAAR1) as a New Target for the Treatment of Cognitive Dysfunction in Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 7811. [Google Scholar] [CrossRef]
- Giménez-Palomo, A.; Vieta, E. The potential of TAAR1 agonists in bipolar disorder. Eur. Neuropsychopharmacol. 2022, 62, 4–6. [Google Scholar] [CrossRef]
- Karovicova, J.; Kohajdova, Z.; Simko, P.; Lukacova, D. Using capillary isotachophoresis for the determination of biogenic amines and D-isocitric acid in food products. Nahrung 2003, 47, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Geracitano, R.; Federici, M.; Bernardi, G.; Mercuri, N.B. On the effects of psychostimulants, antidepressants, and the antiparkinsonian drug levodopa on dopamine neurons. Ann. N. Y. Acad. Sci. 2006, 1074, 320–329. [Google Scholar] [CrossRef]
- Katolikova, N.V.; Vaganova, A.N.; Efimova, E.V.; Gainetdinov, R.R. Expression of Trace Amine-Associated Receptors in the Murine and Human Hippocampus Based on Public Transcriptomic Data. Cells 2022, 11, 1813. [Google Scholar] [CrossRef]
- Gainetdinov, R.R. Dopamine transporter mutant mice in experimental neuropharmacology. Naunyn Schmiedebergs Arch. Pharmacol. 2008, 377, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Yu, Y.; Han, Y.; Wei, J.; Collier, D.; Li, T.; Ma, X. The role of single nucleotide polymorphism of D2 dopamine receptor gene on major depressive disorder and response to antidepressant treatment. Psychiatry Res. 2012, 200, 1047–1050. [Google Scholar] [CrossRef]
- Moore, C.F.; Sabino, V.; Cottone, P. Trace Amine Associated Receptor 1 (TAAR1) Modulation of Food Reward. Front. Pharmacol. 2018, 9, 129. [Google Scholar] [CrossRef]
- Rutigliano, G.; Bandini, L.; Sestito, S.; Chiellini, G. 3-Iodothyronamine and Derivatives: New Allies against Metabolic Syndrome? Int. J. Mol. Sci. 2020, 21, 2005. [Google Scholar] [CrossRef]
- Guariento, S.; Tonelli, M.; Espinoza, S.; Gerasimov, A.S.; Gainetdinov, R.R.; Cichero, E. Rational design, chemical synthesis and biological evaluation of novel biguanides exploring species-specificity responsiveness of TAAR1 agonists. Eur. J. Med. Chem. 2018, 146, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Ferragud, A.; Howell, A.D.; Moore, C.F.; Ta, T.L.; Hoener, M.C.; Sabino, V.; Cottone, P. The Trace Amine-Associated Receptor 1 Agonist RO5256390 Blocks Compulsive, Binge-like Eating in Rats. Neuropsychopharmacology 2017, 42, 1458–1470. [Google Scholar] [CrossRef]
- Raony, Í.; Domith, I.; Lourenco, M.V.; Paes-de-Carvalho, R.; Pandolfo, P. Trace amine-associated receptor 1 modulates motor hyperactivity, cognition, and anxiety-like behavior in an animal model of ADHD. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 117, 110555. [Google Scholar] [CrossRef]
- Wu, R.; Liu, J.; Li, J.X. Trace amine-associated receptor 1 and drug abuse. Adv. Pharmacol. 2022, 93, 373–401. [Google Scholar] [PubMed]
- Decker, A.M.; Brackeen, M.F.; Mohammadkhani, A.; Kormos, C.M.; Hesk, D.; Borgland, S.L.; Blough, B.E. Identification of a Potent Human Trace Amine-Associated Receptor 1 Antagonist. ACS Chem. Neurosci. 2022, 13, 1082–1095. [Google Scholar] [CrossRef]
- Glyakina, A.V.; Pavlov, C.D.; Sopova, J.V.; Gainetdinov, R.R.; Leonova, E.I.; Galzitskaya, O.V. Search for Structural Basis of Interactions of Biogenic Amines with Human TAAR1 and TAAR6 Receptors. Int. J. Mol. Sci. 2021, 23, 209. [Google Scholar] [CrossRef]
- Mantas, I.; Millan, M.J.; Di Cara, B.; Groenink, L.; Veiga, S.; Cistarelli, L.; Brocco, M.; Bertrand, M.; Svenningsson, P.; Zhang, X. Trace Amine-Associated Receptor 1 Contributes to Diverse Functional Actions of O-Phenyl-Iodotyramine in Mice but Not to the Effects of Monoamine-Based Antidepressants. Int. J. Mol. Sci. 2021, 22, 8907. [Google Scholar] [CrossRef]
- Hopkins, S.C.; Ogirala, A.; Worden, M.; Koblan, K.S. Depicting Safety Profile of TAAR1 Agonist Ulotaront Relative to Reactions Anticipated for a Dopamine D2-Based Pharmacological Class in FAERS. Clin. Drug Investig. 2021, 41, 1067–1073. [Google Scholar] [CrossRef]
- Heffernan, M.L.R.; Herman, L.W.; Brown, S.; Jones, P.G.; Shao, L.; Hewitt, M.C.; Campbell, J.E.; Dedic, N.; Hopkins, S.C.; Koblan, K.S.; et al. Ulotaront: A TAAR1 Agonist for the Treatment of Schizophrenia. ACS Med. Chem. Lett. 2021, 13, 92–98. [Google Scholar] [CrossRef]
- Cisneros, I.E.; Ghorpade, A. Methamphetamine and HIV-1-induced neurotoxicity: Role of trace amine associated receptor 1 cAMP signaling in astrocytes. Neuropharmacology 2014, 85, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.M. The emerging role of trace amine-associated receptor 1 in the functional regulation of monoamine transporters and dopaminergic activity. J. Neurochem. 2011, 116, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Grandy, D.K. Trace amine-associated receptor 1-Family archetype or iconoclast? Pharmacol. Ther. 2007, 116, 355–390. [Google Scholar] [CrossRef]
- Alnefeesi, Y.; Tamura, J.K.; Lui, L.M.W.; Jawad, M.Y.; Ceban, F.; Ling, S.; Nasri, F.; Rosenblat, J.D.; McIntyre, R.S. Trace amine-associated receptor 1 (TAAR1): Potential application in mood disorders: A systematic review. Neurosci. Biobehav. Rev. 2021, 131, 192–210. [Google Scholar] [CrossRef] [PubMed]
- Raab, S.; Wang, H.; Uhles, S.; Cole, N.; Alvarez-Sanchez, R.; Künnecke, B.; Ullmer, C.; Matile, H.; Bedoucha, M.; Norcross, R.D.; et al. Incretin-like effects of small molecule trace amine-associated receptor 1 agonists. Mol. Metab. 2016, 5, 47–56. [Google Scholar] [CrossRef]
- Narang, D.; Tomlinson, S.; Holt, A.; Mousseau, D.D.; Baker, G.B. Trace amines and their relevance to psychiatry and neurology: A brief overview. Klin. Psikofarmakol. Bülteni. 2011, 1, 73–79. [Google Scholar] [CrossRef]
- Halff, E.F.; Rutigliano, G.; Garcia-Hidalgo, A.; Howes, O.D. Trace amine-associated receptor 1 (TAAR1) agonism as a new treatment strategy for schizophrenia and related disorders. Trends Neurosci. 2023, 46, 60–74. [Google Scholar] [CrossRef]
- de Bartolomeis, A.; Ciccarelli, M.; Vellucci, L.; Fornaro, M.; Iasevoli, F.; Barone, A. Update on novel antipsychotics and pharmacological strategies for treatment-resistant schizophrenia. Expert Opin. Pharmacother. 2022, 17, 1–18. [Google Scholar] [CrossRef]
- Krasavin, M.; Peshkov, A.A.; Lukin, A.; Komarova, K.; Vinogradova, L.; Smirnova, D.; Kanov, E.V.; Kuvarzin, S.R.; Murtazina, R.Z.; Efimova, E.V.; et al. Discovery and In Vivo Efficacy of Trace Amine-Associated Receptor 1 (TAAR1) Agonist 4-(2-Aminoethyl)-N-(3,5-dimethylphenyl)piperidine-1-carboxamide Hydrochloride (AP163) for the Treatment of Psychotic Disorders. Int. J. Mol. Sci. 2022, 23, 11579. [Google Scholar] [CrossRef]
- Kane, J.M. A New Treatment Paradigm: Targeting Trace Amine-Associated Receptor 1 (TAAR1) in Schizophrenia. J. Clin. Psychopharmacol. 2022, 42 (Suppl. S1), S1–S13. [Google Scholar] [CrossRef]
- Nair, P.C.; Chalker, J.M.; McKinnon, R.A.; Langmead, C.J.; Gregory, K.J.; Bastiampillai, T. Trace Amine-Associated Receptor 1 (TAAR1): Molecular and Clinical Insights for the Treatment of Schizophrenia and Related Comorbidities. ACS Pharmacol. Transl. Sci. 2022, 5, 183–188. [Google Scholar] [CrossRef]
- Correll, C.U.; Abi-Dargham, A.; Howes, O. Emerging Treatments in Schizophrenia. J. Clin. Psychiatry 2022, 83, 39683. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, M. The Breakthrough of TAAR1 Agonists for the Treatment of Neuropsychiatric Disorders: One Step Away. Curr. Med. Chem. 2022, 29, 4893–4895. [Google Scholar] [CrossRef] [PubMed]
- Dedic, N.; Dworak, H.; Zeni, C.; Rutigliano, G.; Howes, O.D. Therapeutic Potential of TAAR1 Agonists in Schizophrenia: Evidence from Preclinical Models and Clinical Studies. Int. J. Mol. Sci. 2021, 22, 13185. [Google Scholar] [CrossRef]
- Dodd, S.F.; Carvalho, A.; Puri, B.K.; Maes, M.; Bortolasci, C.C.; Morris, G.; Berk, M. Trace Amine-Associated Receptor 1 (TAAR1): A new drug target for psychiatry? Neurosci. Biobehav. Rev. 2021, 120, 537–541. [Google Scholar] [CrossRef]
- Grinchii, D.; Hoener, M.C.; Khoury, T.; Dekhtiarenko, R.; Bervanlou, R.N.; Jezova, D.; Dremencov, E. Effects of acute and chronic administration of trace amine-associated receptor 1 (TAAR1) ligands on in vivo excitability of central monoamine-secreting neurons in rats. Mol. Psychiatry 2022, 27, 4861–4868. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.T.; Wang, H.; Niu, W.P.; Zhang, C.C.; Zhang, Y.; Wang, X.D.; Si, T.M.; Su, Y.A. Role of trace amine-associated receptor 1 in the medial prefrontal cortex in chronic social stress-induced cognitive deficits in mice. Pharmacol. Res. 2021, 167, 105571. [Google Scholar] [CrossRef] [PubMed]
- Dedic, N.; Jones, P.G.; Hopkins, S.C.; Lew, R.; Shao, L.; Campbell, J.E.; Spear, K.L.; Large, T.H.; Campbell, U.C.; Hanania, T.; et al. SEP-363856, a novel psychotropic agent with a unique, non-D2 receptor mechanism of action. J. Pharm. Exp. 2019, 371, 1–14. [Google Scholar] [CrossRef]
- Ren, X.; Xiong, J.; Liang, L.; Chen, Y.; Zhang, G. Molecules. The Potential Antidepressant Action of Duloxetine Co-Administered with the TAAR1 Receptor Agonist SEP-363856 in Mice. Molecules 2022, 27, 2755. [Google Scholar] [CrossRef]
- Feemster, J.C.; Westerland, S.M.; Gossard, T.R.; Steele, T.A.; Timm, P.C.; Jagielski, J.T.; Strainis, E.; McCarter, S.J.; Hopkins, S.C.; Koblan, K.S.; et al. Treatment with the novel TAAR1 agonist ulotaront is associated with reductions in quantitative polysomnographic REM sleep without atonia in healthy human subjects: Results of a post-hoc analysis. Sleep Med. 2023, 101, 578–586. [Google Scholar] [CrossRef]
- Xiao, G.; Chen, Y.L.; Dedic, N.; Xie, L.; Koblan, K.S.; Galluppi, G.R. In Vitro ADME and Preclinical Pharmacokinetics of Ulotaront, a TAAR1/5-HT1A Receptor Agonist for the Treatment of Schizophrenia. Pharm. Res. 2022, 39, 837–850. [Google Scholar] [CrossRef] [PubMed]
- Højlund, M.; Correll, C.U. Ulotaront: A TAAR1/5-HT1A agonist in clinical development for the treatment of schizophrenia. Expert Opin. Investig. Drugs. 2022, 12, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Zhukov, I.S.; Kubarskaya, L.G.; Karpova, I.V.; Vaganova, A.N.; Karpenko, M.N.; Gainetdinov, R.R. Minor Changes in Erythrocyte Osmotic Fragility in Trace Amine-Associated Receptor 5 (TAAR5) Knockout Mice. Int. J. Mol. Sci. 2021, 22, 7307. [Google Scholar] [CrossRef]
- Murtazina, R.Z.; Zhukov, I.S.; Korenkova, O.M.; Popova, E.A.; Kuvarzin, S.R.; Efimova, E.V.; Kubarskaya, L.G.; Batotsyrenova, E.G.; Zolotoverkhaya, E.A.; Vaganova, A.N.; et al. Genetic Deletion of Trace-Amine Associated Receptor 9 (TAAR9) in Rats Leads to Decreased Blood Cholesterol Levels. Int. J. Mol. Sci. 2021, 22, 2942. [Google Scholar] [CrossRef]
- Zhukov, I.S.; Ptukha, M.A.; Zolotoverkhaja, E.A.; Sinitca, E.L.; Tissen, I.Y.; Karpova, I.V.; Volnova, A.B.; Gainetdinov, R.R. Evaluation of Approach to a Conspecific and Blood Biochemical Parameters in TAAR1 Knockout Mice. Brain Sci. 2022, 12, 614. [Google Scholar] [CrossRef]
- Liu, J.; Wu, R.; Johnson, B.; Zhang, Y.; Zhu, Q.; Li, J.X. Selective TAAR1 agonists induce conditioned taste aversion. Psychopharmacology 2022, 239, 3345–3353. [Google Scholar] [CrossRef]
- Regard, J.B.; Kataoka, H.; Cano, D.A.; Camerer, E.; Yin, L.; Zheng, Y.W.; Scanlan, T.S.; Hebrok, M.; Coughlin, S.R. Probing cell type-specific functions of Gi in vivo identifies GPCR regulators of insulin secretion. J. Clin. Investig. 2007, 117, 4034–4043. [Google Scholar] [CrossRef]
- Braulke, L.J.; Klingenspor, M.; De Barber, A.; Tobias, S.C.; Grandy, D.K.; Scanlan, T.S.; Heldmaier, G.J. 3-Iodothyronamine: A novel hormone controlling the balance between glucose and lipid utilisation. Comp. Physiol. B 2008, 178, 167–177. [Google Scholar] [CrossRef]
- Apryatin, S.A.; Shipelin, V.A.; Trusov, N.V.; Mzhelskaya, K.V.; Evstratova, V.S.; Kirbaeva, N.V.; Soto, J.S.; Fesenko, Z.S.; Gainetdinov, R.R.; Gmoshinski, I.V. Comparative analysis of the influence of a high-fat/high-carbohydrate diet on the level of anxiety and neuromotor and cognitive functions in Wistar and DAT-KO rats. Physiol. Rep. 2019, 7, e13987. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Tuohimaa, P. The grooming analysis algorithm discriminates between different levels of anxiety in rats: Potential utility for neurobehavioural stress research. J. Neurosci. Methods 2005, 143, 169–177. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Stewart, A.M.; Song, C.; Berridge, K.C.; Graybiel, A.M.; Fentress, J.C. Neurobiology of rodent self-grooming and its value for translational neuroscience. Nat. Rev. Neurosci. 2016, 17, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Evangelou, K.; Gorgoulis, V.G. Sudan Black B, The Specific Histochemical Stain for Lipofuscin: A Novel Method to Detect Senescent Cells. Methods Mol. Biol. 2017, 1534, 111–119. [Google Scholar]
- Chan, J.K.C.T. The Wonderful Colors of the Hematoyxlin−Eosin Stain in Diagnostic Surgical Pathology. Int. J. Surg. Pathol. 2014, 22, 12–32. [Google Scholar] [CrossRef] [PubMed]
- Zhukov, I.S.; Karpova, I.V.; Krotova, N.A.; Tissen, I.Y.; Demin, K.A.; Shabanov, P.D.; Budygin, E.A.; Kalueff, A.V.; Gainetdinov, R.R. Enhanced Aggression, Reduced Self-Grooming Behavior and Altered 5-HT Regulation in the Frontal Cortex in Mice Lacking Trace Amine-Associated Receptor 1 (TAAR1). Int. J. Mol. Sci. 2022, 23, 14066. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apryatin, S.A.; Zhukov, I.S.; Zolotoverkhaya, E.A.; Kuvarzin, S.R.; Khunagov, T.A.; Ushmugina, S.V.; Klimenko, V.M. Protein Metabolism Changes and Alterations in Behavior of Trace Amine-Associated Receptor 1 Knockout Mice Fed a High-Fructose Diet. Neurol. Int. 2023, 15, 339-351. https://doi.org/10.3390/neurolint15010022
Apryatin SA, Zhukov IS, Zolotoverkhaya EA, Kuvarzin SR, Khunagov TA, Ushmugina SV, Klimenko VM. Protein Metabolism Changes and Alterations in Behavior of Trace Amine-Associated Receptor 1 Knockout Mice Fed a High-Fructose Diet. Neurology International. 2023; 15(1):339-351. https://doi.org/10.3390/neurolint15010022
Chicago/Turabian StyleApryatin, Sergey A., Ilya S. Zhukov, Ekaterina A. Zolotoverkhaya, Saveliy R. Kuvarzin, Temirkan A. Khunagov, Sanelya V. Ushmugina, and Victor M. Klimenko. 2023. "Protein Metabolism Changes and Alterations in Behavior of Trace Amine-Associated Receptor 1 Knockout Mice Fed a High-Fructose Diet" Neurology International 15, no. 1: 339-351. https://doi.org/10.3390/neurolint15010022